
Grazing and Mowing Affect the Carbon-to-Nitrogen Ratio of Plants by Changing the Soil Available Nitrogen Content and Soil Moisture on the Meadow Steppe, China


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
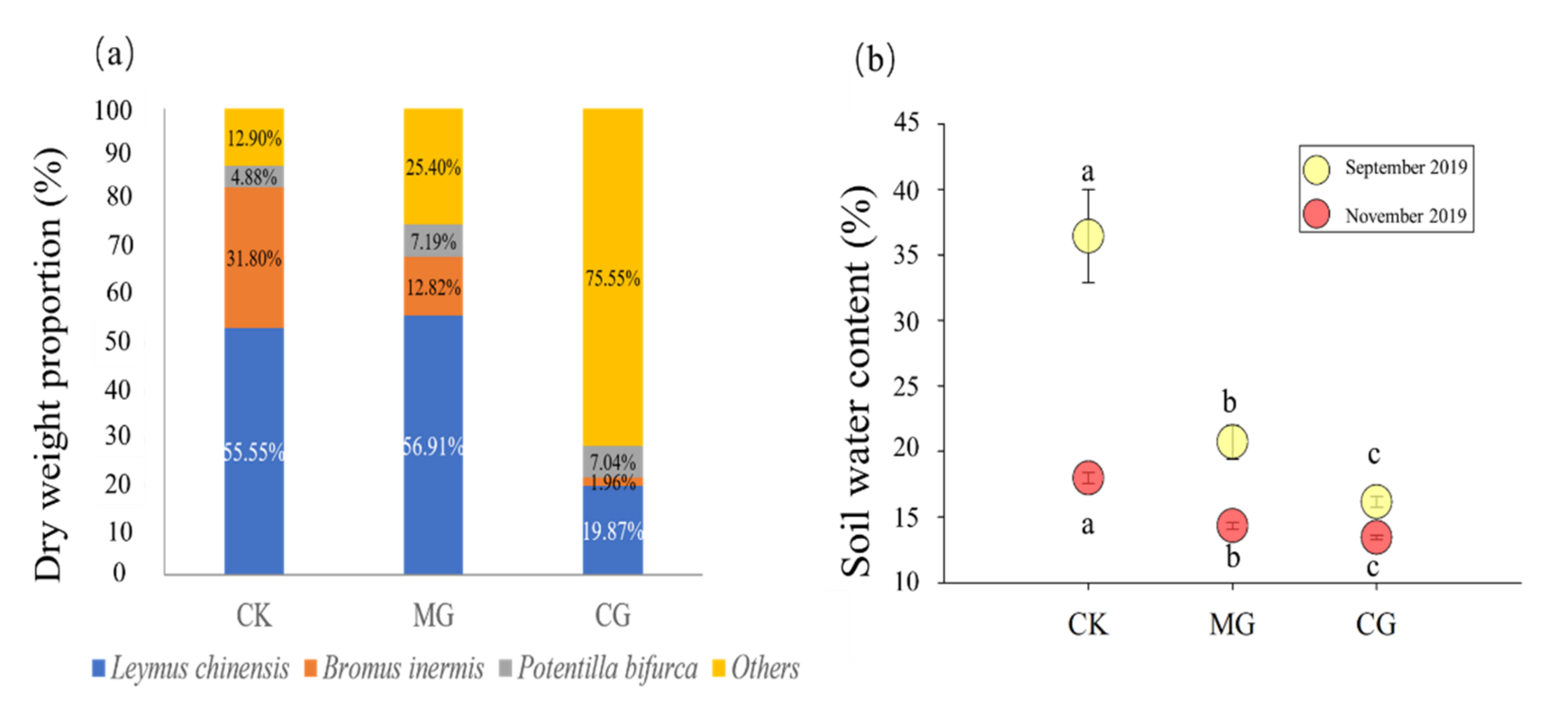
2.1. Dry Weight Proportions of Three Main Plants and Soil Water Content
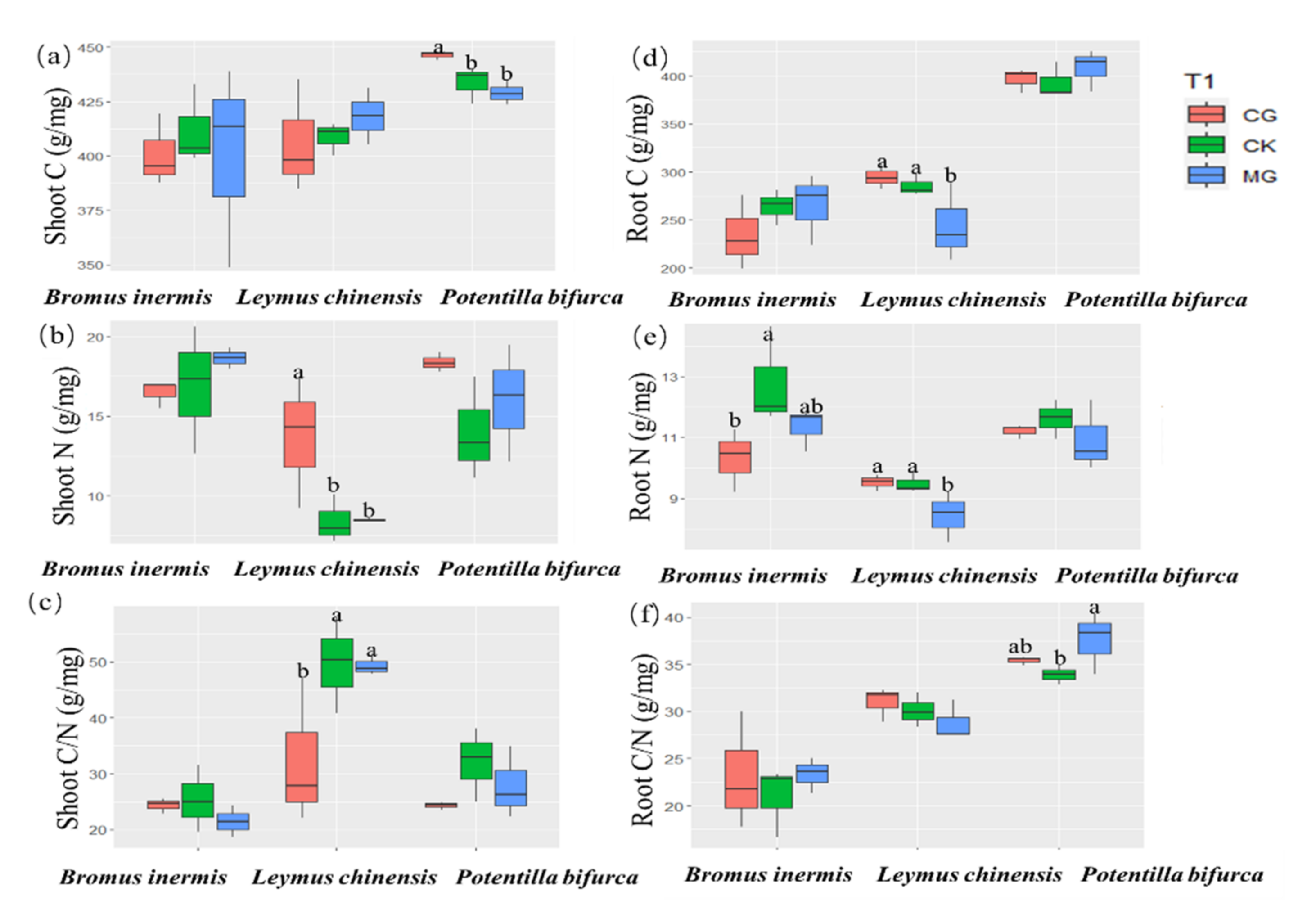
2.2. Responses of the Carbon to Nitrogen Ratios of Three Dominant Plants
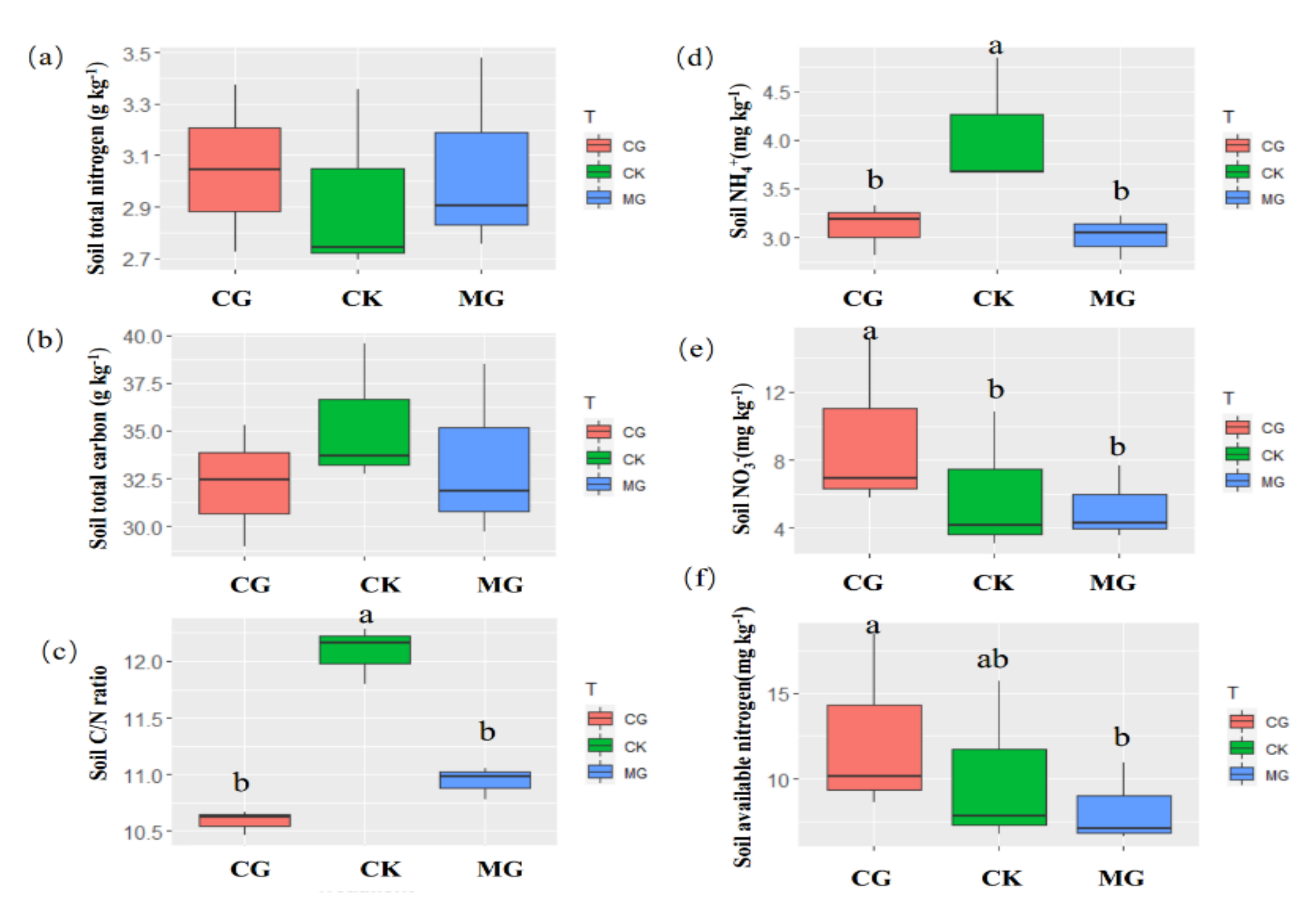
2.3. Soil Nutrient and the C/N Ratio
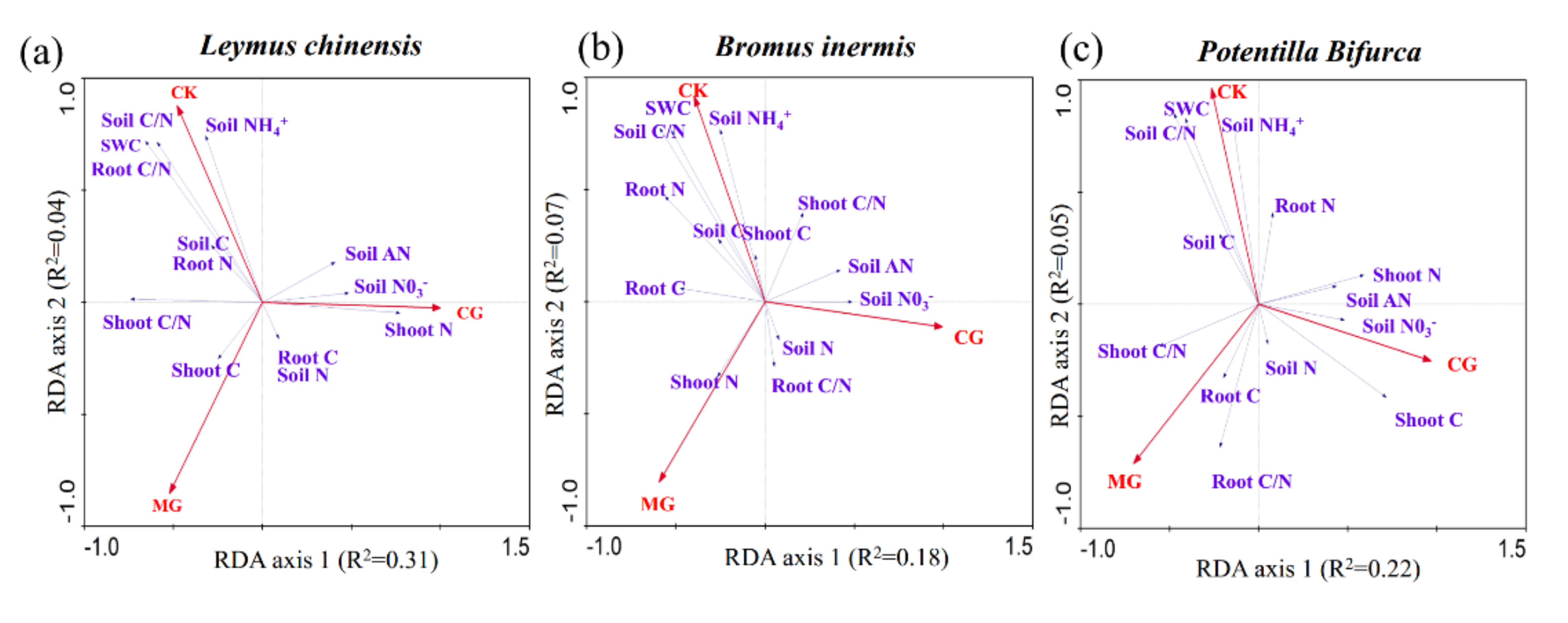
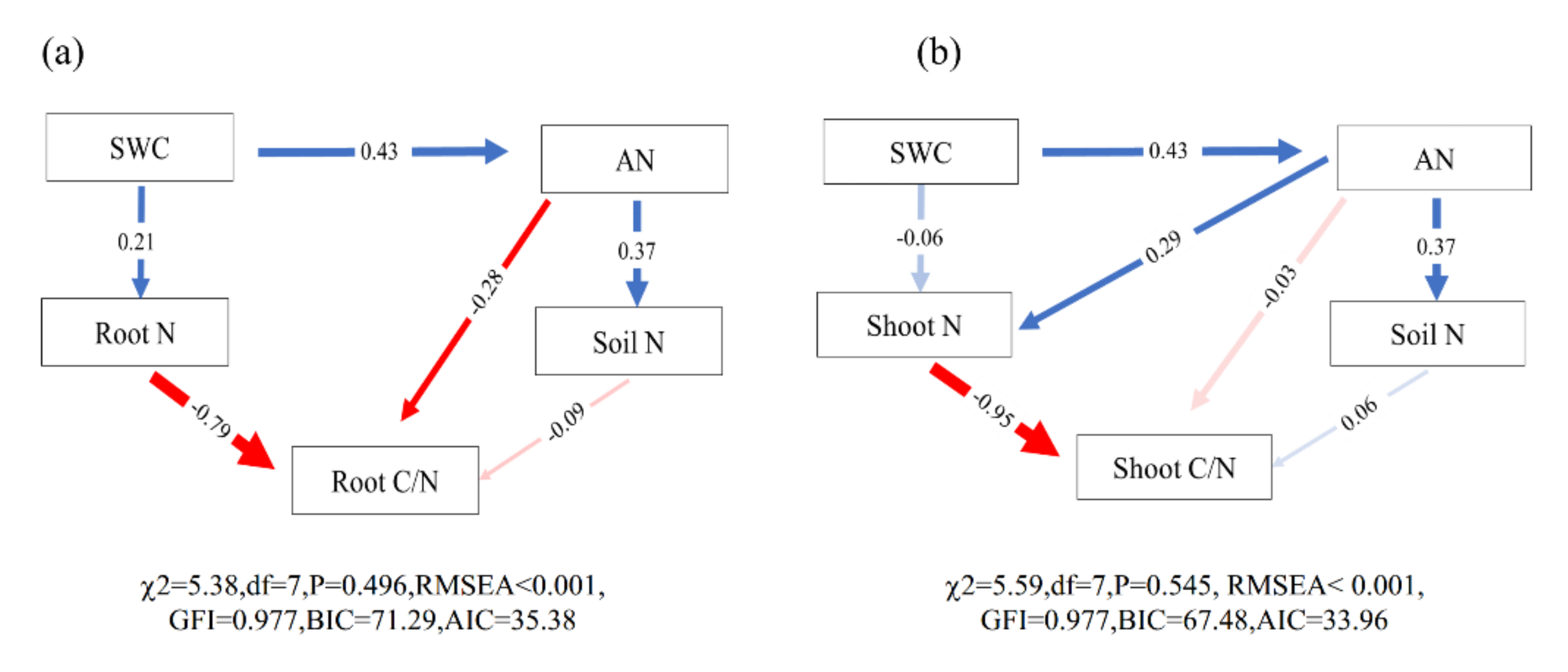
2.4. Relationship between the C/N Ratio of Three Plants and Environmental Factors
3. Discussion
3.1. Driving Factors of Variation in C/N Ratio in Three Plant Species across Different Land Use Types
3.2. Changes in Soil Nutrient Availability and Soil Moisture in Response to Grassland Management
4. Materials and Methods
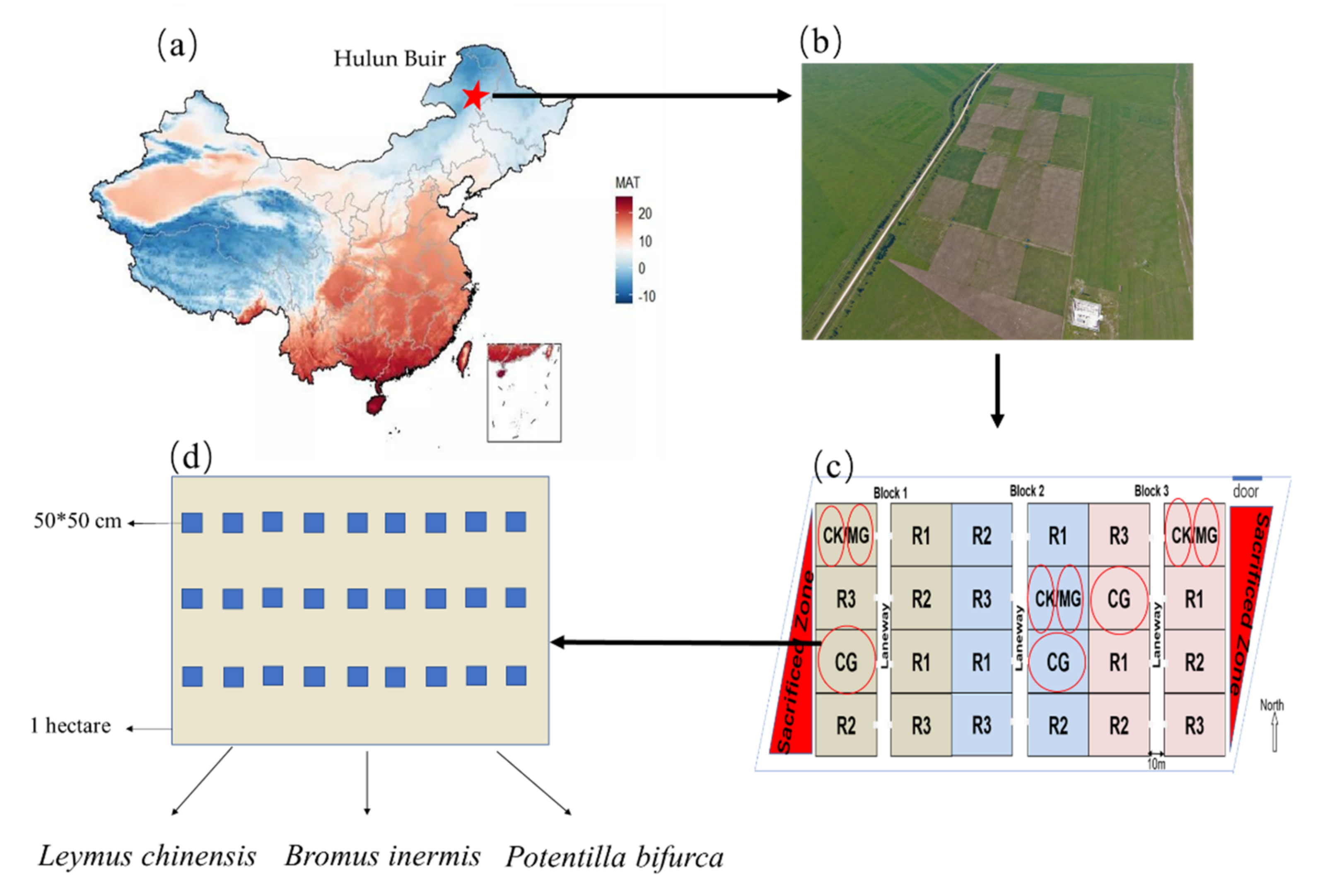
4.1. Study Site
4.2. Sampling and Chemical Measurement
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bai, Y.; Wu, J.; Clark, C.M.; Naeem, S.; Han, X.; Bai, Y.F.; Wu, J.G.; Clark, C.M.; Naeem, S.; Pan, Q.M.; et al. Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from Inner Mongolia grasslands. Glob. Chang. Biol. 2009, 16, 358–372. [Google Scholar] [CrossRef]
- Min, Q.; Liu, S.; Yang, X. Evaluation of the Ecosystem Services of the Inner Mongolia Steppe. Acta Agrestia Sin. 2004, 12, 165–169. [Google Scholar]
- Ning, J.; Jin, J.; Kuang, F.; Wan, X.; Zhang, C.; Guan, T. The Valuation of Grassland Ecosystem Services in Inner Mongolia of China and Its Spatial Differences. Sustainability 2019, 11, 7117. [Google Scholar] [CrossRef] [Green Version]
- Heyburn, J.; McKenzie, P.; Crawley, M.J.; Fornara, D.A. Effects of grassland management on plant C:N:P stoichiometry: Implications for soil element cycling and storage. Ecosphere 2017, 8, 1963. [Google Scholar] [CrossRef]
- Luo, G.P.; Han, Q.F.; Zhou, D.C.; Li, L.; Chen, X.; Li, Y.; Hu, Y.K.; Li, B.L. Moderate grazing can promote aboveground primary production of grassland under water stress. Ecol. Complex. 2012, 11, 126–136. [Google Scholar] [CrossRef]
- Pan, F.J.; Yang, L.Y.; Wang, C.L.; Yan, R.R.; Li, C.J.; Hu, Y.F.; Jiang, Y.; Cao, J.; Tan, H.Y.; Xin, X.P. Effects of mowing frequency on abundance, genus diversity and community traits of soil nematodes in a meadow steppe in northeast China. Plant Soil 2020. [Google Scholar] [CrossRef]
- Wiesmeier, M.; Steffens, M.; Kölbl, A.; Kögel-Knabner, I. Degradation and small-scale spatial homogenization of topsoils in intensively-grazed steppes of Northern China. Soil Tillage Res. 2009, 104, 299–310. [Google Scholar] [CrossRef]
- Han, D.; Wang, G.; Xue, B.; Liu, T.; Yinglan, A.; Xu, X. Evaluation of semiarid grassland degradation in North China from multiple perspectives. Ecol. Eng. 2018, 112, 41–50. [Google Scholar] [CrossRef]
- Li, S.; Verburg, P.H.; Lv, S.; Wu, J.; Li, X. Spatial analysis of the driving factors of grassland degradation under conditions of climate change and intensive use in Inner Mongolia, China. Reg. Environ. Chang 2012, 12, 461–474. [Google Scholar] [CrossRef]
- Hoogendoorn, C.J.; Newton, P.C.D.; Devantier, B.P.; Rolle, B.A.; Lloyd-West, C.M. Grazing intensity and micro-topographical effects on some nitrogen and carbon pools and fluxes in sheep-grazed hill country in New Zealand. Agric. Ecosyst. Environ. 2016, 217, 22–32. [Google Scholar] [CrossRef]
- Han, X.; Li, Y.; Du, X.; Li, Y.; Wang, Z.; Jiang, S.; Li, Q. Effect of grassland degradation on soil quality and soil biotic community in a semi-arid temperate steppe. Ecol. Processes 2020, 9, 1–11. [Google Scholar] [CrossRef]
- Mu, S.; Zhu, C.; Zhou, K.; Li, J. The Preventive Strategies of Degradation and the Approaches to Enhance Carbon Sequestration Ability in Inner Mongolia Grassland. Acta Agrestia Sin. 2017, 25, 217–225. [Google Scholar]
- Pan, W.; Song, Z.; Liu, H.; Mueller, K.; Yang, X.; Zhang, X.; Li, Z.; Liu, X.; Qiu, S.; Hao, Q.; et al. Impact of grassland degradation on soil phytolith carbon sequestration in Inner Mongolian steppe of China. Geoderma 2017, 308, 86–92. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Gallardol, A.; Bowker, M.A.; Wallenstein, M.D.; Luis Quero, J.; Ochoa, V.; Gozalo, B.; Garcia-Gomez, M.; Soliveres, S.; et al. Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature 2013, 502, 672–676. [Google Scholar] [CrossRef]
- Zhou, G.Y.; Zhou, X.H.; He, Y.H.; Shao, J.J.; Hu, Z.H.; Liu, R.Q.; Zhou, H.M.; Hosseinibai, S. Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: A meta-analysis. Glob. Chang Biol. 2017, 23, 1167–1179. [Google Scholar] [CrossRef]
- Lanuza, O.; Casanoves, F.; Delgado, D.; Van den Meersche, K. Leaf litter stoichiometry affects decomposition rates and nutrient dynamics in tropical forests under restoration in Costa Rica. Restor. Ecol. 2019, 27, 549–558. [Google Scholar] [CrossRef]
- Tang, B.; Man, J.; Bai, Y. Leaf nitrogen acquisition of Leymus chinensis varies with leaf age and land use change in a semiarid grassland. Environ. Exp. Bot. 2020, 175, 104051. [Google Scholar] [CrossRef]
- Liu, M.; Gong, J.R.; Pan, Y.; Luo, Q.P.; Zhai, Z.W.; Yang, L.L.; Xu, S. Response of dominant grassland species in the temperate steppe of Inner Mongolia to different land uses at leaf and ecosystem levels. Photosynthetica 2018, 56, 921–931. [Google Scholar] [CrossRef]
- Lu, X.-T.; Freschet, G.T.; Kazakou, E.; Wang, Z.-W.; Zhou, L.-S.; Han, X.-G. Contrasting responses in leaf nutrient-use strategies of two dominant grass species along a 30-yr temperate steppe grazing exclusion chronosequence. Plant Soil 2015, 387, 69–79. [Google Scholar] [CrossRef]
- Chi, Y.; Zhou, L.; Yang, Q.; Li, S.-p.; Zheng, S. Increased snowfall weakens complementarity of summer water use by different plant functional groups. Ecol. Evol. 2019, 9, 4264–4274. [Google Scholar] [CrossRef]
- Wedin, D.A. Nitrogen availability, plant soil feedbacks and grassland stability. In Proceedings of the VI International Rangeland Congress on People and Rangelands Building the Future, Townsville, QLD, Australia, July 19–23 1999; Volume 1, pp. 193–197. [Google Scholar]
- Medina-Roldán, E.; Paz-Ferreiro, J.; Bardgett, R.D. Grazing exclusion affects soil and plant communities, but has no impact on soil carbon storage in an upland grassland. Agric. Ecosyst. Environ. 2012, 149, 118–123. [Google Scholar] [CrossRef]
- Bai, Y.F.; Wu, J.G.; Clark, C.M.; Pan, Q.M.; Zhang, L.X.; Chen, S.P.; Wang, Q.B.; Han, X.G. Grazing alters ecosystem functioning and C:N:P stoichiometry of grasslands along a regional precipitation gradient. J. Appl. Ecol. 2012, 49, 1204–1215. [Google Scholar] [CrossRef]
- Lue, X.-T.; Han, X.-G. Nutrient resorption responses to water and nitrogen amendment in semi-arid grassland of Inner Mongolia, China. Plant Soil 2010, 327, 481–491. [Google Scholar] [CrossRef]
- Wang, C.; Wan, S.; Xing, X.; Zhang, L.; Han, X. Temperature and soil moisture interactively affected soil net N mineralization in temperate grassland in Northern China. Soil Biol. Biochem. 2006, 38, 1101–1110. [Google Scholar] [CrossRef]
- Huang, J.; Yu, H.; Zhang, F.; Li, M.; Lin, H. Water- and N-induced changes in soil C:N:P stoichiometry and its implications for N limitation of a desert steppe species, Glycyrrhiza uralensis. Pol. J. Ecol. 2016, 64, 241–254. [Google Scholar] [CrossRef]
- Breman, H.N.; Wit, C.T.D. Rangeland productivity and exploitation in the Sahel. Science 1983, 221, 1341–1347. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.S.A.; Kreyling, J.; Beierkuhnlein, C.; Jentsch, A. Ecotype-specific improvement of nitrogen status in European grasses after drought combined with rewetting. Acta Oecologica-Int. J. Ecol. 2016, 77, 118–127. [Google Scholar] [CrossRef]
- Xiao, H.; Yang, H.L.; Zhao, M.L.; Monaco, T.A.; Rong, Y.P.; Huang, D.; Song, Q.; Zhao, K.; Wang, D.P. Soil extracellular enzyme activities and the abundance of nitrogen-cycling functional genes responded more to N addition than P addition in an Inner Mongolian meadow steppe. Sci. Total Environ. 2021, 759, 143541. [Google Scholar] [CrossRef]
- Adams, K.J.; Valdis, K.; Eric, F.; Lora, B.; Van, D.P.W.H.; Vishal, S. Herbivory and Stoichiometric Feedbacks to Primary Production. PLoS ONE 2015, 10, e0129775. [Google Scholar]
- Wang, Z.; Lu, J.; Yang, H.; Zhang, X.; Luo, C.; Zhao, Y. Resorption of nitrogen, phosphorus and potassium from leaves of lucerne stands of different ages. Plant Soil 2014, 383, 301–312. [Google Scholar] [CrossRef]
- Turner, C.L.; Seastedt, T.R.; Dyer, M.I. Maximization of Aboveground Grassland Production: The Role of Defoliation Frequency, Intensity, and History. Ecol. Appl. 1993, 3, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Patterson, K.; Cakmak, T.; Cooper, A.; Lager, I.; Rasmusson, A.G.; Escobar, M.A. Distinct signalling pathways and transcriptome response signatures differentiate ammonium- and nitrate-supplied plants. Plant Cell Environ. 2010, 33, 1486–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, B.; Zheng, X.; Brüggemann, N.; Chen, W.; Dannenmann, M.; Han, X.; Sutton, M.A.; Wu, H.; Yao, Z.; Butterbach-Bahl, K. Grazing-induced reduction of natural nitrous oxide release from continental steppe. Nature 2010, 464, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Venterink, H.O.; Kardel, I.; Kotowski, W.; Peeters, W.; Wassen, M.J. Long-term effects of drainage and hay-removal on nutrient dynamics and limitation in the Biebrza mires, Poland. Biogeochemistry 2009, 93, 235–252. [Google Scholar] [CrossRef]
- Seagle, S.W.; McNaughton, S.J.; Ruess, R.W. Simulated effects of grazing on soil-nitrogen and mineralization in contrasting serengeti grasslands. Ecology 1992, 73, 1105–1123. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Leemans, D.K.; Cook, R.; Hobbs, P.J. Seasonality of the soil biota of grazed and ungrazed hill grasslands. Soil Biol. Biochem. 1997, 29, 1285–1294. [Google Scholar] [CrossRef]
- Wang, G.; Cao, F. Effects of soil water contents on nutrient uptake and allocation of baldcypress. For. Res. Beijing 2004, 17, 213–219. [Google Scholar]
- Zhou, J.; Cui, J.; Hu, F. Effect of soil water on the citrus root and leaf nutrition. China Fruits 2008, 5, 14–17. [Google Scholar]
- Chen, D.; Li, J.; Lan, Z.; Hu, S.; Bai, Y. Soil acidification exerts a greater control on soil respiration than soil nitrogen availability in grasslands subjected to long-term nitrogen enrichment. Funct. Ecol. 2016, 30, 658–669. [Google Scholar] [CrossRef]
- Zi, H.; Chen, Y.; Hu, L.; Wang, C. Effects of nitrogen addition on root dynamics in an alpine meadow, Northwestern Sichuan. Chin. J. Plant Ecol. 2018, 42, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Kirkby, C.A.; Richardson, A.E.; Wade, L.J.; Batten, G.D.; Kirkegaard, J.A. Carbon-nutrient stoichiometry to increase soil carbon sequestration. Soil Biol. Biochem. 2013, 60, 77–86. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Xu, H.; Zhang, H.; Zhang, Y. Grazing and Mowing Affect the Carbon-to-Nitrogen Ratio of Plants by Changing the Soil Available Nitrogen Content and Soil Moisture on the Meadow Steppe, China. Plants 2022, 11, 286. https://doi.org/10.3390/plants11030286
Wang L, Xu H, Zhang H, Zhang Y. Grazing and Mowing Affect the Carbon-to-Nitrogen Ratio of Plants by Changing the Soil Available Nitrogen Content and Soil Moisture on the Meadow Steppe, China. Plants. 2022; 11(3):286. https://doi.org/10.3390/plants11030286
Chicago/Turabian StyleWang, Le, Hengkang Xu, Hao Zhang, and Yingjun Zhang. 2022. "Grazing and Mowing Affect the Carbon-to-Nitrogen Ratio of Plants by Changing the Soil Available Nitrogen Content and Soil Moisture on the Meadow Steppe, China" Plants 11, no. 3: 286. https://doi.org/10.3390/plants11030286
APA StyleWang, L., Xu, H., Zhang, H., & Zhang, Y. (2022). Grazing and Mowing Affect the Carbon-to-Nitrogen Ratio of Plants by Changing the Soil Available Nitrogen Content and Soil Moisture on the Meadow Steppe, China. Plants, 11(3), 286. https://doi.org/10.3390/plants11030286