1. Introduction
In haploids or wide hybrids of plants, standard meiosis can sometimes be abbreviated to a single division, generating two products instead of four, but with somatic chromosome numbers. This phenomenon has been observed and described numerous times in haploids and wide hybrids of wheat,
Triticum aestivum L. [
1,
2,
3,
4,
5,
6]. It effectively doubles the chromosome number of progeny; a mechanism probably responsible in large part for the creation of allopolyploids [
7]. Several different terms have been proposed for such altered meiosis: “unreductional meiotic cell division”, “single division meiosis”, “haploid meiotic restitution” or “indeterminate meiotic restitution” [
5,
8,
9,
10,
11]. In some situations there appears to be no division at all with the resulting monad carrying twice the somatic chromosome number of the generating plant [
1]. For the sake of consistency with general terminology [
8], the term “first division restitution”, FDR, will be used here. In FDR, univalents line up in an orderly fashion on the metaphase plate of what should be the first meiotic division and separate sister chromatids in anaphase I; second division is abandoned. The final product of FDR is a dyad where each nucleus, in most cases, contains a somatic number of chromosomes. Occasional chiasmate pairing, or precocious migration of some univalents to the poles, alter the chromosome constitutions of the gametes and of resulting progeny [
12,
13,
14]. Apart from such deviations, the consequences of FDR are the same as of mitosis: two daughter nuclei containing the somatic number of chromosomes. Fusion of thus produced gametes with unreduced chromosome numbers produces progeny with doubled chromosome number, creating a fertile polyploid.
In standard meiosis, the basic unit of division in meiosis I is a bivalent, a structure composed of two homologues connected by a chiasma or chiasmata. Each homologue is fully replicated, so a bivalent consists of four chromatids: two sisters and two non-sisters. Dissolution of sister chromatid cohesion along the chromosome arms, but not in the centromeres, releases chiasmata, and the homologues are free to migrate to the opposite poles of the karyokinetic spindle. Separation of sister chromatids at the centromeres takes place in anaphase II. With four chromatids present, a bivalent contains four centromeres and care has to be taken to make sure the centromeres of sister chromatids (sister centromeres) co-orient to the same pole of the karyokinetic spindle. This is accomplished by different packaging of chromosomes for meiosis than for mitosis. In meiosis, sister chromatids are tightly bound by cohesins along their entire lengths, and sister centromeres are fused into single units. Hence, a bivalent presents only two centromeric units to the microtubules emanating from the poles [
15,
16,
17]. This meiosis-specific behavior of sister centromeres, which makes orderly separation first of bivalents in anaphase I and then sister chromatids in anaphase II, is accomplished by a step-wise control of meiotic kinases [
18]. However, in hexaploid wheat, univalents, which clearly are packaged in the same manner as chromosomes involved in chiasmate associations, sometimes remain on the metaphase plate and separate sister chromatids already in the first anaphase [
19]. The frequency of such precocious sister chromatid separation appears to be proportional to time spent on the metaphase plate as the proportion of univalents separating sister chromatids increases with the progression of anaphase I. Most often, however, univalents in hexaploid wheat show all evidence of monopolar interaction with the karyokinetic spindle, meandering between the poles as a consequence of transient microtubule stabilization, and when off the plate, do not separate sister chromatids. On the other hand, univalents in bipolar attachment to the karyokinetic spindle are always located on the metaphase plate and often undergo misdivision (breakage across the centromere) [
19].
Another striking difference in univalent behavior between standard meiosis and FDR is their orientation relative to the metaphase plate, or the axis of the karyokinetic spindle. In metaphase I of standard meiosis, univalents in a bipolar attachment to microtubules are almost always in a parallel orientation to the spindle axis (perpendicular to the metaphase plate) while those in a monopolar attachment are almost always V-shaped, probably as a consequence of friction with the cytoplasm during random meandering between the spindle poles, and the majority of them are located off the plate (on fixed preparations). Univalents left on the metaphase plate in anaphase I appear immobilized, are always oriented parallel to the plate and are straight. In FDR, univalents which congregate on the metaphase plate in MI are oriented parallel to it and are almost always straight.
In FDR, sister centromeres separate in what is the equivalent of the first meiotic division; it is an interesting question as to whether chromosomes, and specifically their centromeres, are pre-packaged for FDR differently than they are for standard meiosis, to facilitate orderly congregation on the metaphase plate, or is the packaging the same, and the conversion occurs during the division itself. Packaging similar to that for mitosis would easily explain the behavior of univalents on the FDR metaphase I plate. Mitotic-like organization, with sister centromeres on the opposite sides of replicated chromosomes, would greatly facilitate their bipolar attachment to the karyokinetic spindle, and hence, placement on the metaphase plate. However, if packaging of centromeres in FDR is the same as for normal meiosis, it is not clear how univalents with functionally single centromeres (fused sister centromeres), hence a strong propensity for monopolar interactions with the karyokinetic spindle and haphazard movements throughout the volume of a meiocyte, place themselves on the metaphase plate, attach amphitelically to the spindle, and do not appear to suffer from centric misdivision.
This study focused on the appearance of centromeres from the earliest identifiable stages of meiosis through anaphase I in wheat x rye (Secale cereale L.) F1 hybrids with FDR. The observations showed that while longitudinal differentiation along the centromeres can be observed in some chromosomes, prior to metaphase I there was no separation of sister centromeres. Such separation appeared to take place on the metaphase I plate itself. This brought about another question, so far unanswered, as to what mechanism delivers these univalents to the plate itself, and in parallel orientation to it. This parallel orientation implies that it may not be the centromeres, or not centromeres alone, that are responsible for the movement of chromosomes.
3. Discussion
There was nothing unusual about three different modes of meiotic division, in most cases side-by-side in individual anthers, observed in the material analyzed here; it has been observed in similar hybrids of wheat before [
1,
4,
6,
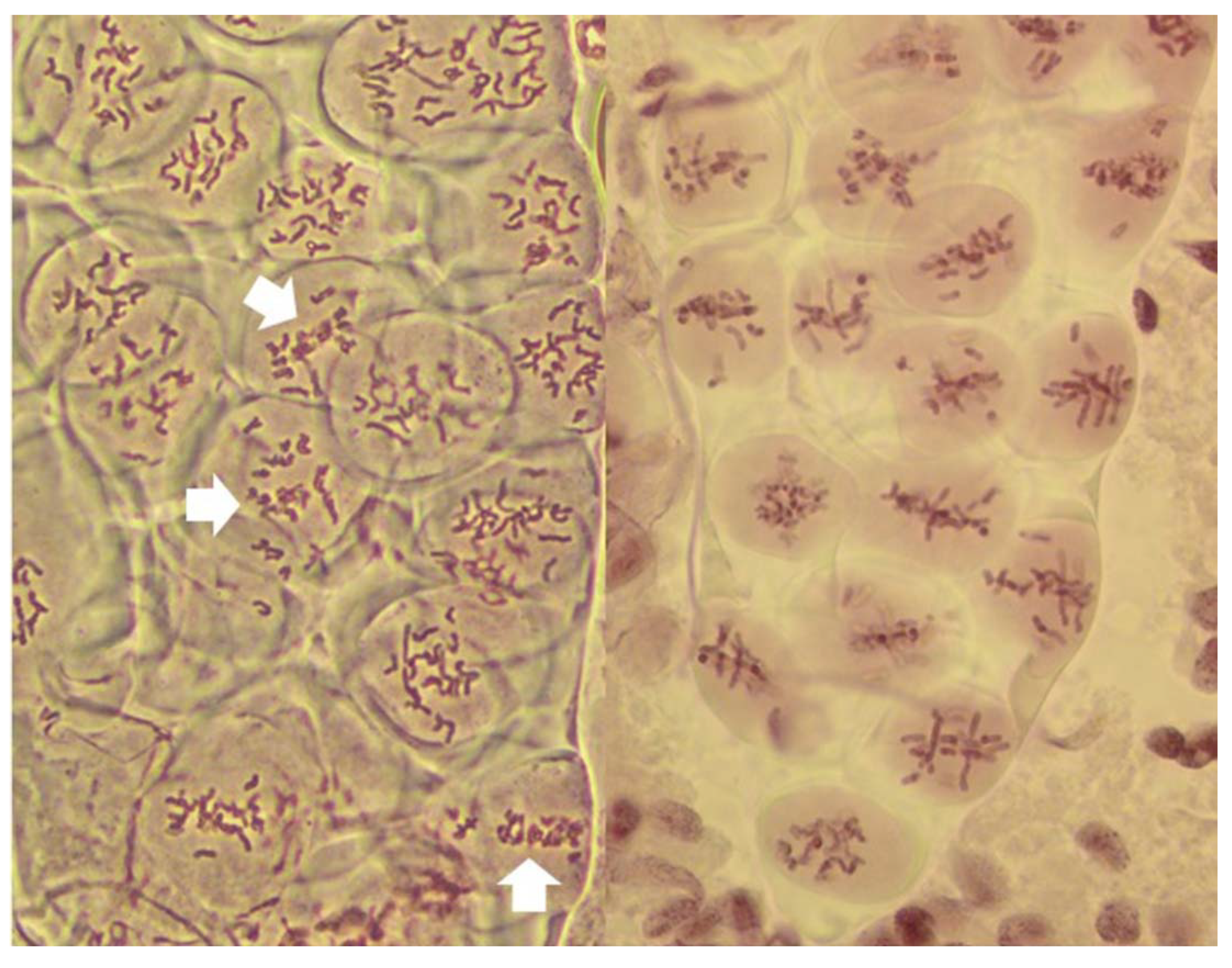
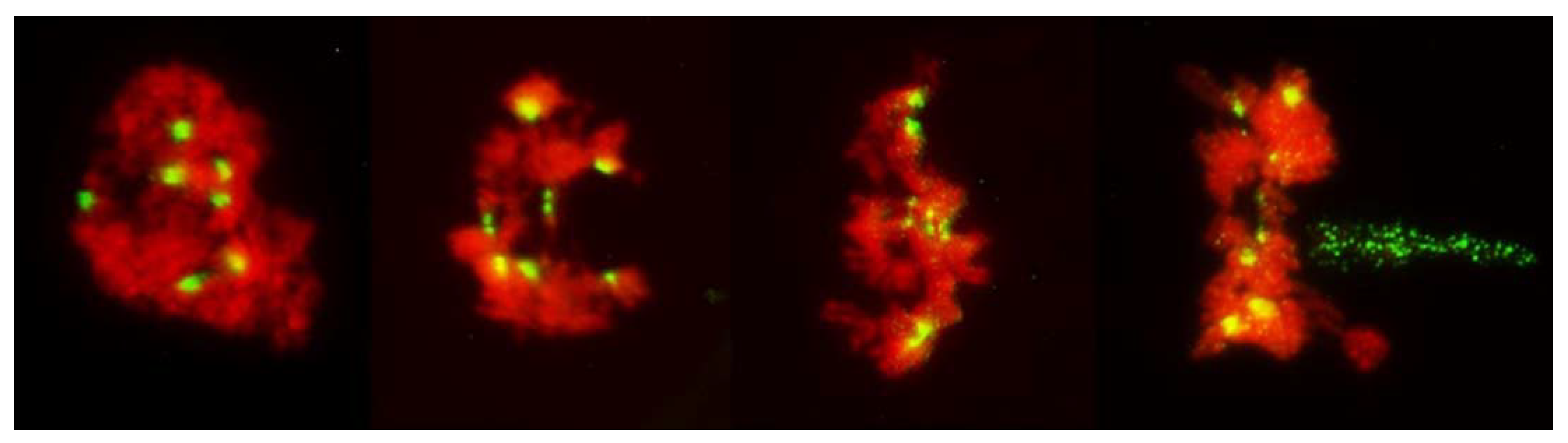
20]. Here, it served as built-in control for the FDR; the frequencies of normal reductional meiosis and of FDR were high in all tested plants guaranteeing that samples observed in the pre-MI stages contained PMCs destined for one of the three pathways. The no-division meiosis was the least frequent, and perhaps for that reason was observed in only three of the ten analyzed plants.
It has been proposed that FDR was delayed relative to normal meiosis [
21]. In standard reductional meiosis, univalents lingering on the metaphase plate frequently separate sister chromatids in AI, and this frequency increases with time spent on the plate [
19].). In this sense, more time spent on the metaphase plate in FDR would explain sister chromatid separation in AI. This notion is well supported by published images such as
Figure 1e in [
20] where a PMC undergoing FDR is surrounded by PMCs in MII-AII (that is, PMCs undergoing standard reductional meiosis). However, in this study, quite the opposite was observed, and with considerable frequency: in a column of PMCs in a typical MI stage both types of division, reductional and FDR, are present side-by-side (
Figure 1). As a matter of fact, because the FDR PMCs clearly show separation of sister chromatids, effectively they are already in the AI-like stage while PMCs with reductional division are still in MI, because bivalents are present. Therefore, the FDR PMCs appear to be ahead of the reductional division PMCs. On the other hand, the no-division PMCs clearly are delayed relative to reductional meiosis (and FDR) but the fault here appears to be in chromatin condensation; it is only partial, no metaphase-like chromosomes are ever formed, and there is no nuclear division, even if cytokinesis may proceed.
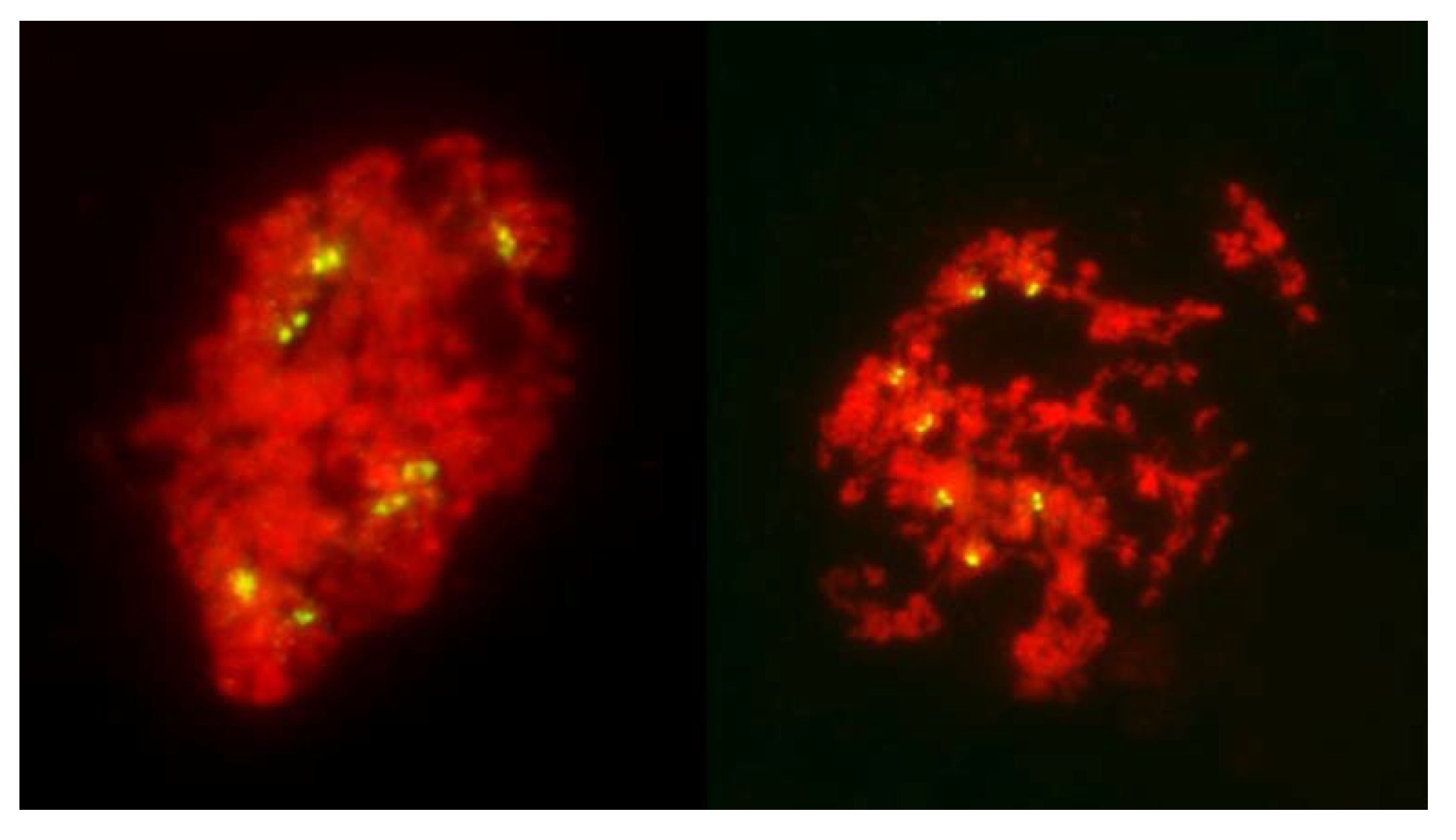
It is evident from the observations, on large numbers of PMCs and univalents, that packaging of centromeres was the same in all observed prophase I PMCs, regardless of their later mode of division: the FDR with essentially mitotic-like separation of sister chromatids, a standard reductional meiosis or no division at all. There is a clear difference between mitotic and meiotic packaging of chromosomes, including their centromeres, which makes separation of chromosomes from bivalents, and later of sister chromatids, possible [
22]. In yeast, the mono-orientation of sister kinetochores in meiosis involves a meiosis-specific cohesin subunit [
23] which makes the two-step dissolution of cohesion possible [
24]. This meiosis-specific cohesion subunit is protected in the first meiotic division, guaranteeing co-migration of sister chromatids to daughter nuclei, and dissolved only in the second division. Here, in all pre-MI PMCs observed, all sister centromeres were fused into single units from the earliest recognizable stages of the meiotic prophase until metaphase I (
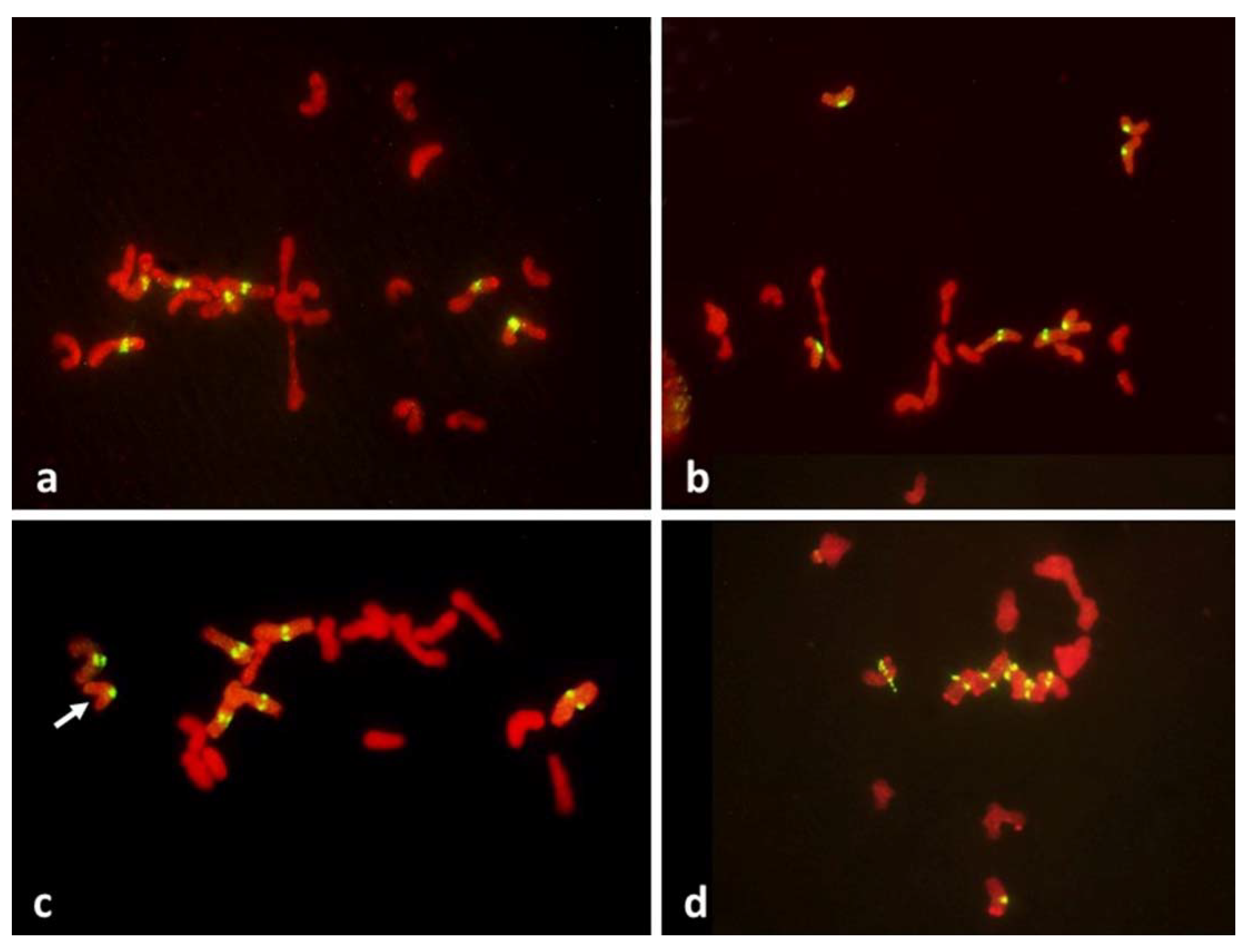
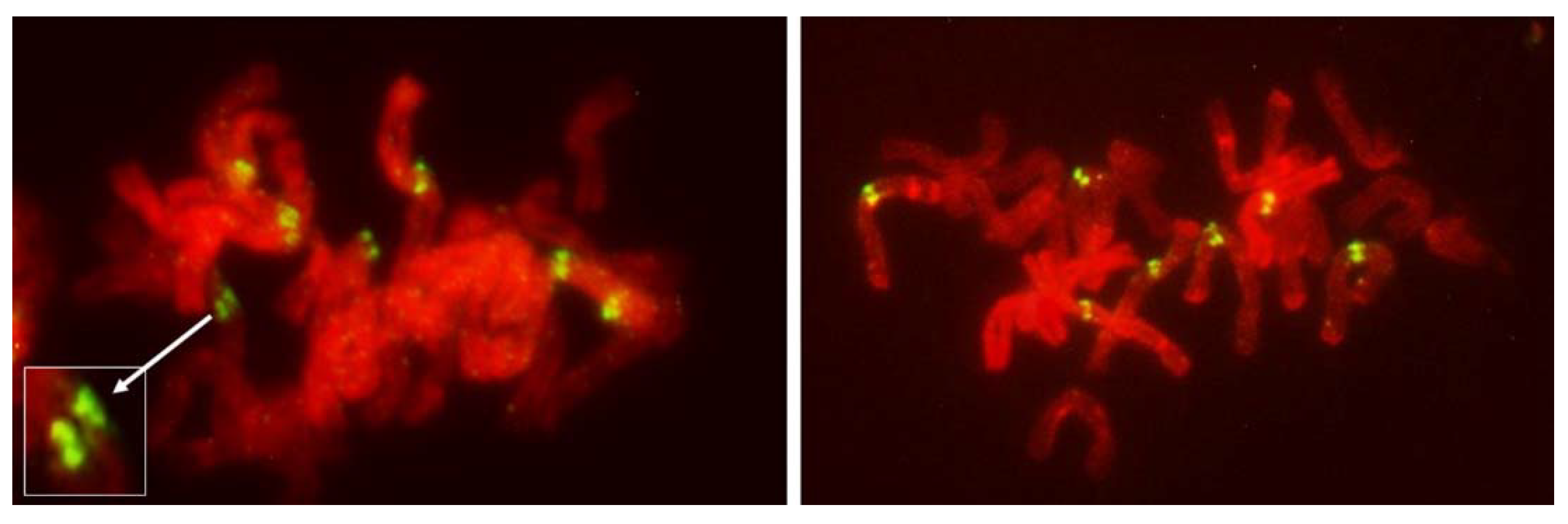
Figure 8) offering no hint of differences in packaging for different modes of meiosis. Reorganization of sister centromeres in univalents occurred only during MI (bivalents were still present) on the MI plate itself, and only in the PMCs undergoing FDR. This reorganization appeared to take place only in univalents located on the plate itself, or in its immediate vicinity, and never occurred in paired chromosomes. Univalents which precociously moved away from the plate were immune to change, and remained with their sister centromeres fused (
Figure 4 and
Figure 5). Very similar observations were described in [
25], in a similar material and using a similar approach: univalents with labeled centromeres approach the metaphase plate amphitelically oriented toward the spindle poles, and perpendicular to the spindle axis, and eventually separate sister chromatids. As in this study, figures in [
25] show no indication of sister centromere separation which would facilitate such an orderly migration.
In a standard nuclear division, be it mitosis or meiosis, placement of a chromosome on the metaphase plate is a consequence of a bipolar/amphitelic attachment of the centromeres, with their kinetochores, to the karyokinetic spindle. In mitosis, a replicated chromosome presents two centromeres to the karyokinetic spindle; these sister centromeres are on the opposite sides of a replicated chromosome, and in a constriction which further restricts the angle from which each one can be targeted by microtubule fibers. This promotes proper bipolar interaction with microtubule fibers which stabilizes the chromosome on the metaphase plate, with each centromere facing one of the spindle poles (
Figure 10). A bivalent in meiosis also presents two centromere units (each with fused sister centromeres) to the karyokinetic spindle and when properly attached, becomes stabilized on the metaphase plate, oriented perpendicular to it. For univalents, a bipolar attachment of fused sister centromeres also places them on the metaphase plate, and also orients them perpendicular to the plate. This often leads to centric misdivision (see
Figure 2 in [
19]). Only when such univalents resist misdivision and linger on the plate well into the anaphase, separation of sister centromeres takes place: they reorient parallel to the (former) metaphase plate, and eventually separate sister chromatids. Clearly, in wheat both meiotic steps of sister chromatid separation can take place in the first division [
19].
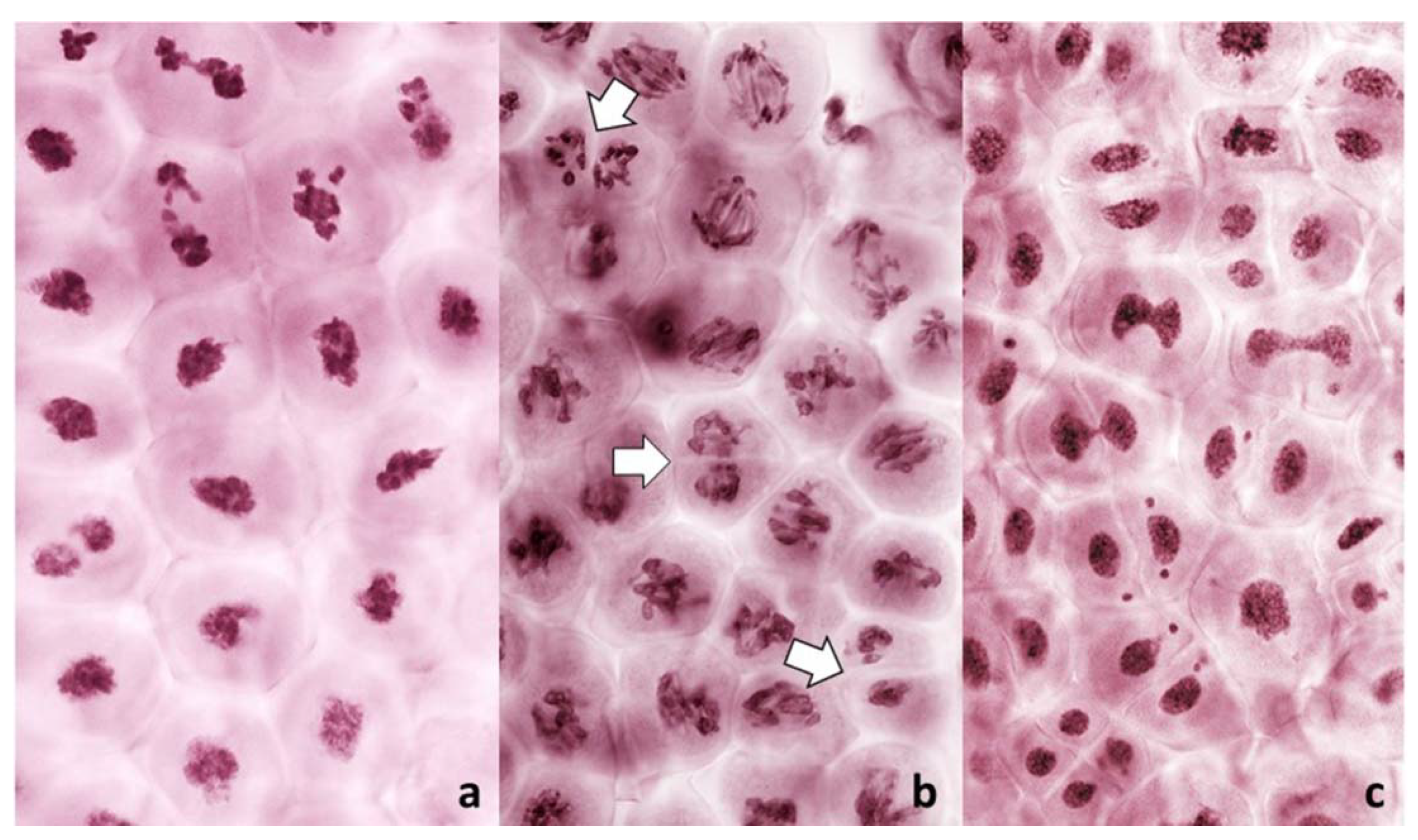
Occasionally, fixation may catch on the metaphase plate a meandering univalent with a monopolar attachment to the karyokinetic spindle, but these are fairly infrequent and easily recognizable events. Univalents in a monopolar attachment are almost always V-shaped (for two-armed chromosomes) and apart from extended centromeres, generally show no signs of tension. In this context, striking was the difference between the FDR and reductional meiosis in the disposition of univalents in a PMC. In MI of reductional meiosis (bivalents are present,
Figure 3), a majority of univalents congregated close to the poles of the karyokinetic spindle, none showed any sign of sister centromere separation and all had fused sister chromatids. Most were either V-shaped or lay parallel to the axis of the spindle. In the FDR meiocytes at MI (as indicated by the presence of bivalents), most univalents congregated on the metaphase plate or close to it, still with sister centromeres fused, but generally straight and perpendicular to the spindle axis (parallel to the metaphase plate). While on the plate itself, sister centromeres separated, at times when sister chromatids still appeared to be attached (
Figure 5). At the onset of anaphase, univalents away from the plate clearly showed separation of sister chromatids along the length of the arms while sister centromeres remained fused (
Figure 5a,c) while those on the plate were showing essentially the opposite.
Differences in univalent orientation and their centromere status imply that perhaps it is not the karyokinetic spindle that places them on the metaphase plate, or that it is not an interaction between the spindle and the centromeres. Cai et al. [
5] have shown that at this stage in the FDR, fully developed karyokinetic spindle is present. Here, even though the microtubules of the karyokinetic spindle were not visualized, it was evident that normally functional karyokinetic spindle also had to be present: bivalents were property positioned on the metaphase plate and were stretched; univalents away from the plate showed all signs of monopolar attachment to the spindle and of the movement dictated by such attachment. However, in the FDR PMCs, univalent behavior and orientation were different from that in standard reductional meiosis or in mitosis. It is not clear what mechanism/system may be responsible for the delivery of univalents to the metaphase plate in the FDR meiocytes in a fashion clearly different from the normal monopolar or bipolar centromere-spindle interaction. That some other mechanism of chromosome movement/transport may be at play is suggested by the appearance and behavior of chromatin in no-division PMCs: clearly, something attempts to separate that chromatin (mostly uncondensed or poorly condensed) and it is not by the attachment to the centromeres but to other parts of chromosomes (
Figure 6 and
Figure 7) as with some frequency the centromeres/centromere regions still provide cohesion of sister chromatids and appear to resist the separation (
Figure 7). Perhaps these are the abnormal karyokinetic spindles as described by [
20,
26,
27]. However, here this mechanism appears to operate simultaneously with a normally functional spindle apparatus, as indicated by perfectly normal positioning of bivalents, and univalents off the metaphase plate (V-shaped, clearly caught in transit through the cytoplasm). In large cells such as oocytes in amphibians, where the centromeres may be beyond the reach/range of microtubules there is an alternative system which delivers chromosomes to the spindle [
28]. Here, however, the situation is different: all univalents clearly are within the range of the spindle but most of them do not appear to interact with it in a standard way in FDR PMCs.
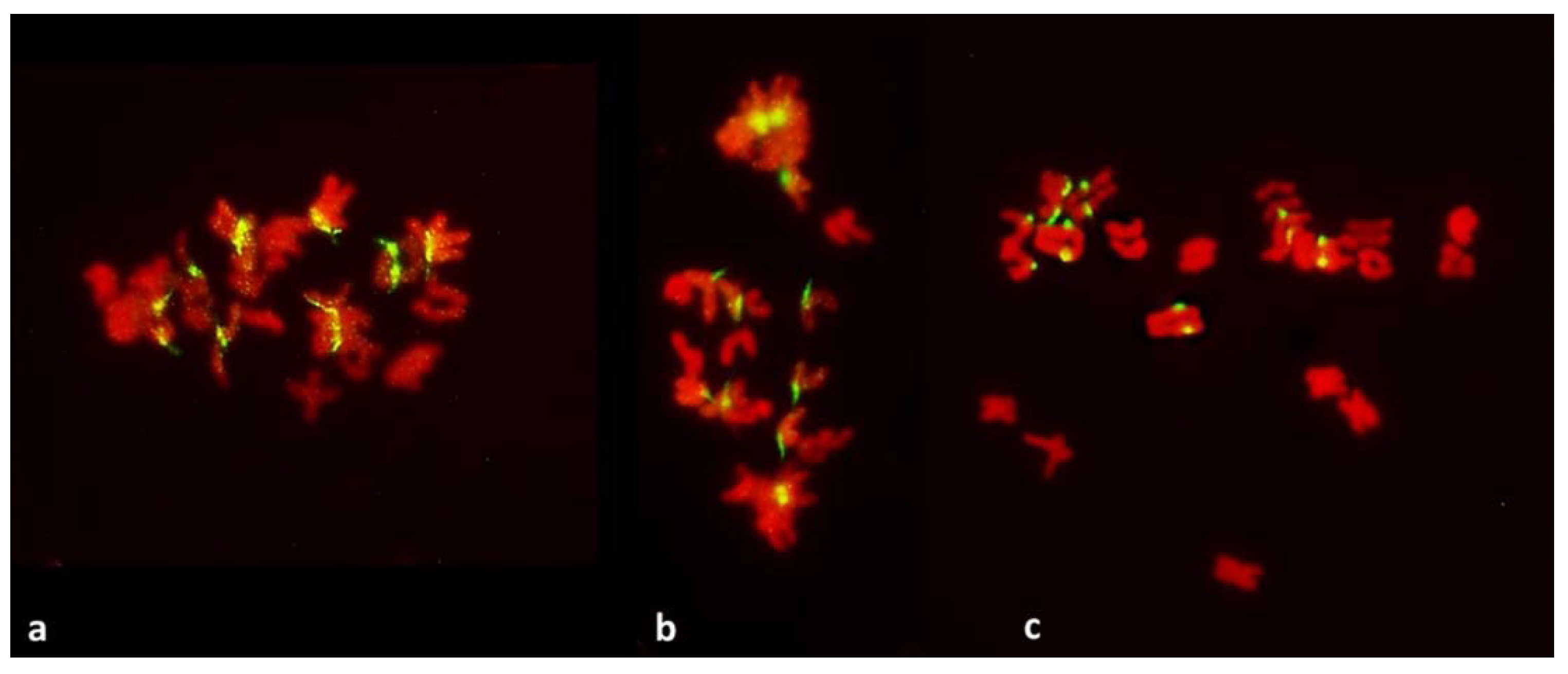
It could be argued that in the absence of pulling forces, separation of sister centromeres would be difficult to detect or be outright undetectable in the pre-MI stages. However, MI itself is informative, and so are several other configurations. In the MI stage of FDR, univalents congregate on the metaphase plate with their sister centromeres fused; these centromeres separate on the metaphase plate itself at which point their bipolar attachment, and the presence of pulling forces, is evident. At the same time and in the same PMCs, univalents beyond the plate still retain their centromere-fused configuration even when cohesion of sister chromatids away from the centromere is already released. This is in clear contrast to the mitotic configuration: in somatic cells of anthers with FDR undergoing standard mitosis (karyokinetic spindle is present) where all labeled centromeres on the metaphase plate are separated (
Figure 10). They are also separated in nuclei undergoing no division meiosis at the stage corresponding to tetrads, where no pulling forces appear to exist (
Figure 9).
4. Materials and Methods
All samples for this study were collected during the earlier study of the first meiotic restitution in an F
1 wheat–rye hybrid [
14]. The plant material consisted of ten F
1 hybrids of a tetraploid wheat line Do1 with a population rye
Secale cereale L. designated as MAD510. The Do1 line was selected by Dr. B. Lapinski, then at the Institute of Plant Genetics, Poznan, Poland, from among the hybrids of
Triticum turgidum var. dicoccoides ssp. spontaneonigrum x T. turgidum var. persicum. The selection criterion was its ability to produce fully fertile F
1 hybrids with rye, a characteristic later identified as a consequence of the restitution of the first meiotic division (FDR). MAD510 was created by an intercross of winter rye cultivars Motto, Amilo and Dankowskie Zlote, all originating from the Danko Plant Breeding, Choryn, Poland; the population is homozygous for an introgression of a segment of wheat chromosome 1D with the locus
Glu-D1 [
29].
The F1 hybrids were grown in a greenhouse at the University of California, Riverside CA campus. Tillers with spikes judged to be at meiosis were cut, spikes dissected and single anthers from individual spikelets were squashed in a drop of acetocarmine. If the desired stage of meiosis was present the remaining two anthers were fixed in a mixture of absolute ethyl alcohol and glacial acetic acid, in proportion 3:1, respectively, and stored at −20 °C for analyses.
Some observations were made on live-stained anthers during collection of the material. These observations were limited to a gross assessment of the proportions of pollen mother cells (PMCs) in various modes of meiosis. Observations of the centromeres were done on squash preparations from fixed anthers, probed in situ with DNA of the rye centromere sequence [
30] labeled with DIG-oxygenin using the DIG-Nick Translation Kit (Roche) according to manufacturer’s instructions, and hybridized to chromosome preparations according to the protocol in [
31]. Hybridization signals were detected with anti-DIG-FITC; herring DNA was used as a block, usually in about 50x excess relative to the probe. Counterstaining was done with 0.3% propidium iodide in the Vectashield antifade solution. Observations were made under a Zeiss Axioscope 20 equipped with epi-fluorescence, recorded with a SPOT RT Color digital camera (Diagnostic Instruments Inc., Sterling Heights, MI, USA ), and processed using the SPOT Advanced and Adobe Photoshop CS software. All images presented here were manipulated as needed to enhance contrast, remove debris if present, properly orient them on the N-S axis and tone down the background noise.5.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









