
War and Peas: Molecular Bases of Resistance to Powdery Mildew in Pea (Pisum sativum L.) and Other Legumes


Abstract
:1. Introduction
2. Know Your Enemy
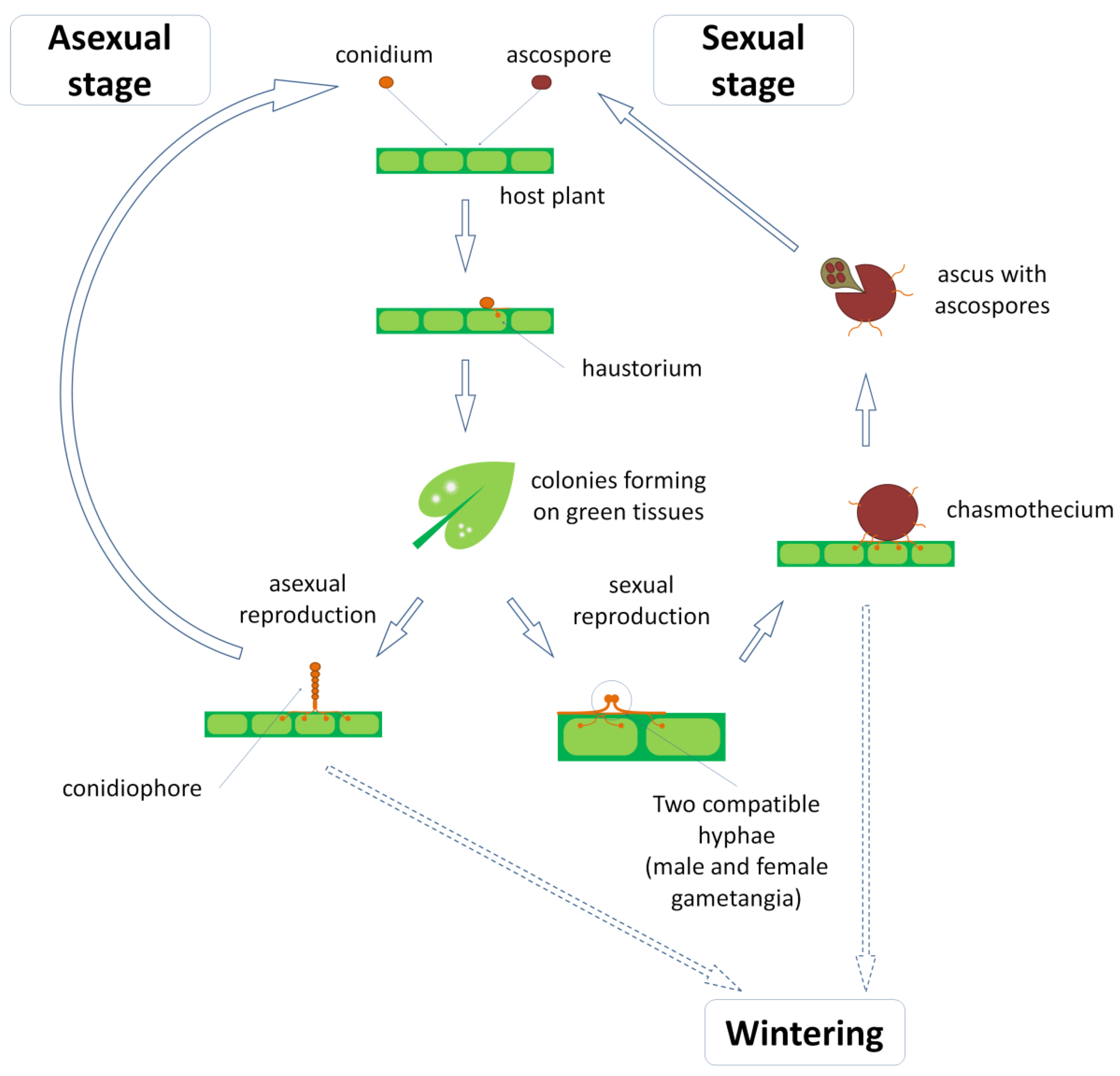
2.1. The Causative Agent of Powdery-Mildew Disease
2.2. Powdery Mildew in Legumes
3. The Art of War
3.1. Chemical Control of the Powdery Mildew
3.2. Biological Control of the Powdery Mildew
4. Close the Gate
5. Gathering Forces
6. Para Bellum: Future Perspectives and Challenges
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
| Name | Sequence, 5′–3′ | Melting t° |
|---|---|---|
| Er1_fw0 | GAA AGA AAA AAT GGC TGA AGA GG | 59.2 |
| Er1_fw1 | GAT AAG GGT CAA GTT GCA TTA G | 58.4 |
| Er1_fw3 | TTT CCA AAA GTA TAT AAG TAG ATC | 55.0 |
| Er1_rv2 | TAA GAA GGA AAA GCA CTG TGA AG | 59.2 |
| PsMLO7Fw | ATG CCA TGT CTC CTG TTC ACC | 61.2 |
| PsMLO5Rv | CTT TAT CTG CAA GAA TGT ACC | 55.4 |

References
- Tripathi, K.; Gore, P.G.; Singh, M.; Pamarthi, R.K.; Mehra, R.; Gayacharan, C. Legume Genetic Resources: Status and Opportunities for Sustainability. In Legume Crops-Prospects, Production and Uses; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Smýkal, P.; Coyne, C.J.; Ambrose, M.J.; Maxted, N.; Schaefer, H.; Blair, M.W.; Berger, J.; Greene, S.L.; Nelson, M.N.; Besharat, N.; et al. Legume Crops Phylogeny and Genetic Diversity for Science and Breeding. CRC. Crit. Rev. Plant Sci. 2015, 34, 43–104. [Google Scholar] [CrossRef] [Green Version]
- Collins, M.; Lang, D.J.; Kelling, K.A. Effects of Phosphorus, Potassium, and Sulfur on Alfalfa Nitrogen-Fixation under Field Conditions 2. Agron. J. 1986, 78, 959–963. [Google Scholar] [CrossRef]
- Snyder, C.S.; Bruulsema, T.W.; Jensen, T.L.; Fixen, P.E. Review of greenhouse gas emissions from crop production systems and fertilizer management effects. Agric. Ecosyst. Environ. 2009, 133, 247–266. [Google Scholar] [CrossRef]
- Hulse, J.H. Nature, composition, and utilization of food legumes. In Expanding the Production and Use of Cool Season Food Legumes; Springer: Dordrecht, The Netherlands, 1994; pp. 77–97. [Google Scholar]
- Upadhyaya, H.D.; Dwivedi, S.L.; Ambrose, M.; Ellis, N.; Berger, J.; Smýkal, P.; Debouck, D.; Duc, G.; Dumet, D.; Flavell, A.; et al. Legume genetic resources: Management, diversity assessment, and utilization in crop improvement. Euphytica 2011, 180, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Maxted, N.; Dulloo, E.; Ford-Lloyd, B.V.; Iriondo, J.M.; Jarvis, A. Gap analysis: A tool for complementary genetic conservation assessment. Divers. Distrib. 2008, 14, 1018–1030. [Google Scholar] [CrossRef]
- Rubiales, D.; Fondevilla, S.; Chen, W.; Gentzbittel, L.; Higgins, T.J.V.; Castillejo, M.A.; Singh, K.B.; Rispail, N. Achievements and Challenges in Legume Breeding for Pest and Disease Resistance. CRC. Crit. Rev. Plant Sci. 2015, 34, 195–236. [Google Scholar] [CrossRef] [Green Version]
- Martins, D.; de Sousa Araújo, S.; Rubiales, D.; Patto, M.C.V. Legume crops and biotrophic pathogen interactions: A continuous cross-talk of a multilayered array of defense mechanisms. Plants 2020, 9, 1460. [Google Scholar] [CrossRef]
- Rubiales, D.; Fernández-Aparicio, M.; Moral, A.; Barilli, E.; Sillero, J.C.; Fondevilla, S. Disease resistance in pea (Pisum sativum L.) types for autumn sowings in mediterranean environments. Czech J. Genet. Plant Breed. 2009, 45, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Sillero, J.C.; Fondevilla, S.; Davidson, J.; Vaz Patto, M.C.; Warkentin, T.D.; Thomas, J.; Rubiales, D. Screening techniques and sources of resistance to rusts and mildews in grain legumes. Euphytica 2006, 147, 255–272. [Google Scholar] [CrossRef]
- Glawe, D.A. The powdery mildews: A review of the world’s most familiar (yet poorly known) plant pathogens. Annu. Rev. Phytopathol. 2008, 46, 27–51. [Google Scholar] [CrossRef]
- Fondevilla, S.; Rubiales, D. Powdery mildew control in pea. A review. Agron. Sustain. Dev. 2012, 32, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Humphry, M.; Reinstädler, A.; Ivanov, S.; Bisseling, T.; Panstruga, R. Durable broad-spectrum powdery mildew resistance in pea er1 plants is conferred by natural loss-of-function mutations in PsMLO1. Mol. Plant Pathol. 2011, 12, 866–878. [Google Scholar] [CrossRef] [PubMed]
- Pavan, S.; Schiavulli, A.; Appiano, M.; Marcotrigiano, A.R.; Cillo, F.; Visser, R.G.F.; Bai, Y.; Lotti, C.; Ricciardi, L. Pea powdery mildew er1 resistance is associated to loss-of-function mutations at a MLO homologous locus. Theor. Appl. Genet. 2011, 123, 1425–1431. [Google Scholar] [CrossRef]
- Linde, M.; Shishkoff, N. Powdery Mildew. In Reference Module in Life Sciences; Taylor & Francis: Abingdon, UK, 2017; pp. 1–8. [Google Scholar] [CrossRef]
- Braun, U. The current systematics and taxonomy of the powdery mildews (Erysiphales): An overview. Mycoscience 2011, 52, 210–212. [Google Scholar] [CrossRef]
- Brown, J.K.M. Comparative genetics of avirulence and fungicide resistance in the powdery mildew fungi. In The Powdery Mildews: A Comprehensive Treatise; Bélanger, R.R., Bushnell, W.R., Dik, A.J., Carver, T.L.W., Eds.; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2002; pp. 56–65. [Google Scholar]
- Zhang, Z.; Henderson, C.; Perfect, E.; Carver, T.L.W.; Thomas, B.J.; Skamnioti, P.; Gurr, S.J. Of genes and genomes, needles and haystacks: Blumeria graminis and functionality. Mol. Plant Pathol. 2005, 6, 561–575. [Google Scholar] [CrossRef]
- Vogel, J.; Somerville, S. Powdery mildew of Arabidopsis: A model system for host-parasite interactions. In The Powdery Mildews: A Comprehensive Treatise; Bélanger, R.R., Bushnell, W.R., Dik, A.J., Carver, T.L.W., Eds.; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2002; pp. 161–168. [Google Scholar]
- Rispail, N.; Prats, E.; Rubiales, D. Medicago truncatula as a model to study powdery mildew resistance. In The Model Legume Medicago truncatula; de Bruijn, F.J., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 390–397. [Google Scholar] [CrossRef]
- Kunoh, H.; Itoh, O.; Kohno, M.; Ishizaki, H. Are primary germ tubes of conidia unique to Erysiphe graminis? Jpn. J. Phytopathol. 1979, 45, 675–682. [Google Scholar] [CrossRef] [Green Version]
- Hacquard, S. The genomics of powdery mildew fungi: Past achievements, present status and future prospects. Adv. Bot. Res. 2014, 70, 109–142. [Google Scholar] [CrossRef]
- Takamatsu, S. Phylogeny and evolution of the powdery mildew fungi (Erysiphales, Ascomycota) inferred from nuclear ribosomal DNA sequences. Mycoscience 2004, 45, 147–157. [Google Scholar] [CrossRef]
- Kiss, L.; Khosla, K.; Jankovics, T.; Niinomi, S.; Braun, U.; Takamatsu, S. A morphologically ill-founded powdery mildew species, Pleochaeta indica, is recognized as a phylogenetic species based on the analysis of the nuclear ribosomal DNA sequences. Mycol. Res. 2006, 110, 1301–1308. [Google Scholar] [CrossRef]
- Takamatsu, S.; Matsuda, S. Estimation of molecular clocks for ITS and 28S rDNA in Erysiphales. Mycoscience 2004, 45, 340–344. [Google Scholar] [CrossRef]
- Cook, R.T.A.; Inman, A.J.; Billings, C. Identification and classification of powdery mildew anamorphs using light and scanning electron microscopy and host range data. Mycol. Res. 1997, 101, 975–1002. [Google Scholar] [CrossRef]
- Takamatsu, S.; Braun, U.; Limkaisang, S. Phylogenetic relationships and generic affinity of Uncinula septata inferred from nuclear rDNA sequences. Mycoscience 2005, 46, 9–16. [Google Scholar] [CrossRef]
- To-anun, C.; Kom-un, S.; Sunawan, A.; Fangfuk, W.; Sato, Y.; Takamatsu, S. A new subgenus, Microidium, of Oidium (Erysiphaceae) on Phyllanthus spp. Mycoscience 2005, 46, 1–8. [Google Scholar] [CrossRef]
- Glawe, D.A. First Report of Powdery Mildew of Nandina domestica Caused by Microsphaera berberidis (Erysiphe berberidis) in the Pacific Northwest. Plant Health Prog. 2003, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, W.R.; Gubler, W.D.; Grove, G.G. Epidemiology of powdery mildews in agricultural pathosystems. In The Powdery Mildews: A Comprehensive Treatise; Bélanger, R.R., Bushnell, W.R., Dik, A.J., Carver, T.L.W., Eds.; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2002; pp. 169–199. [Google Scholar]
- Green, J.R.; Carver, T.L.W.; Gurr, S.J. The formation and function of infection and feeding structures. In The Powdery Mildews: A Comprehensive Treatise; Bélanger, R.R., Bushnell, W.R., Dik, A.J., Carver, T.L.W., Eds.; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2002; pp. 66–82. [Google Scholar]
- Carver, T.L.W.; Kunoh, H.; Thomas, B.J.; Nicholson, R.L. Release and visualization of the extracellular matrix of conidia of Blumeria graminis. Mycol. Res. 1999, 103, 547–560. [Google Scholar] [CrossRef]
- Mims, C.W.; Liljebjelke, K.A.; Richardson, E.A. Surface morphology, wall structure, and initial adhesion of conidia of the powdery mildew fungus Uncinuliella australiana. Phytopathology 1995, 85, 352–358. [Google Scholar] [CrossRef]
- Fondevilla, S.; Carver, T.L.W.; Moreno, M.T.; Rubiales, D. Identification and characterization of sources of resistance to Erysiphe pisi Syd. in Pisum spp. Plant Breed. 2007, 126, 113–119. [Google Scholar] [CrossRef]
- Gniwotta, F.; Vogg, G.; Gartmann, V.; Carver, T.L.W.; Riederer, M.; Jetter, R. What do microbes encounter at the plant surface? Chemical composition of pea leaf cuticular waxes. Plant Physiol. 2005, 139, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Niks, R.E.; Rubiales, D. Potentially durable resistance mechanisms in plants to specialised fungal pathogens. Euphytica 2002, 124, 201–216. [Google Scholar] [CrossRef]
- Consonni, C.; Humphry, M.E.; Hartmann, H.A.; Livaja, M.; Durner, J.; Westphal, L.; Vogel, J.; Lipka, V.; Kemmerling, B.; Schulze-Lefert, P.; et al. Conserved requirement for a plant host cell protein in powdery mildew pathogenesis. Nat. Genet. 2006, 38, 716–720. [Google Scholar] [CrossRef]
- Hückelhoven, R.; Panstruga, R. Cell biology of the plant-powdery mildew interaction. Curr. Opin. Plant Biol. 2011, 14, 738–746. [Google Scholar] [CrossRef]
- Pommerrenig, B.; Müdsam, C.; Kischka, D.; Ekkehard Neuhaus, H. Treat and trick: Common regulation and manipulation of sugar transporters during sink establishment by the plant and the pathogen. J. Exp. Bot. 2020, 71, 3930–3940. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Dubey, S.; Jain, D.; Chandran, D. The Medicago truncatula Sugar Transport Protein 13 and Its Lr67res-Like Variant Confer Powdery Mildew Resistance in Legumes via Defense Modulation. Plant Cell Physiol. 2021, 62, 650–667. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.A.J.; Kenton, P.; Lloyd, A.J.; Ougham, H.; Prats, E. The hypersensitive response; The centenary is upon us but how much do we know? J. Exp. Bot. 2008, 59, 501–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnathorst, W.C. Environmental relationships in the powdery mildews. Annu. Rev. Phytopathol. 1965, 3, 343–366. [Google Scholar] [CrossRef]
- Yarwood, C.E. Powdery mildews. Bot. Rev. 1957, 23, 235–301. [Google Scholar] [CrossRef]
- Plumb, R.T.; Turner, R.H. Scanning electron microscopy of Erysiphe graminis. Trans. Br. Mycol. Soc. 1972, 59, 149–150, IN20–IN21. [Google Scholar] [CrossRef]
- Grove, G.G. Meteorological factors affecting airborne conidia concentrations and the latent period of Podosphaera clandestina on sweet cherry. Plant Dis. 1998, 82, 741–746. [Google Scholar] [CrossRef] [Green Version]
- Adams, G.C., Jr.; Gottwald, T.R.; Leach, C.M. Environmental factors initiating liberation of conidia of powdery mildews. Phytopathology 1986, 76, 1239–1245. [Google Scholar] [CrossRef]
- Byrne, J.M.; Hausbeck, M.K.; Shaw, B.D. Factors affecting concentrations of airborne conidia of Oidium sp. among poinsettias in a greenhouse. Plant Dis. 2000, 84, 1089–1095. [Google Scholar] [CrossRef]
- Peries, O.S. Studies on strawberry mildew, caused by Sphaerotheca macularis (Wallr. ex Fries) Jaczewski* II. Host-parasite relationships on foliage of strawberry varieties. Ann. Appl. Biol. 1962, 50, 225–233. [Google Scholar] [CrossRef]
- Hermansen, J.E.; Torp, U.; Prahm, L.P. Studies of transport of live spores of cereal mildew and rust fungi across the North Sea. Grana 1978, 17, 41–46. [Google Scholar] [CrossRef]
- Braun, U.; Cook, R.T.A.; Inman, A.J.; Shin, H.D. The taxonomy of the powdery mildew fungi. In The Powdery Mildews: A Comprehensive Treatise; Bélanger, R.R., Bushnell, W.R., Dik, A.J., Carver, T.L.W., Eds.; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2002; pp. 13–55. [Google Scholar]
- Braun, U. A monograph of the Erysiphales (powdery mildews). Beih. Nova Hedwig. 1987, 89, 700. [Google Scholar]
- Schnathorst, W.C. Spread and life cycle of the lettuce powdery mildew fungus. Phytopathology 1959, 49, 464–468. [Google Scholar]
- Grove, G.G. Perennation of Uncinula necator in vineyards of Eastern Washington. Plant Dis. 2004, 88, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Rumbolz, J.; Gubler, W.D. Susceptibility of grapevine buds to infection by powdery mildew Erysiphe necator. Plant Pathol. 2005, 54, 535–548. [Google Scholar] [CrossRef]
- Takamatsu, S.; Ito, H.; Shiroya, Y.; Kiss, L.; Heluta, V. First comprehensive phylogenetic analysis of the genus Erysiphe (Erysiphales, Erysiphaceae) I. The Microsphaera lineage. Mycologia 2015, 107, 475–489. [Google Scholar] [CrossRef]
- Falloon, R.E.; Viljanen-Rollinson, S.L.H. Powdery mildew. In Compendium of Pea Diseases and Pests; Kraft, J.M., Pfleger, F.L., Eds.; American Phytopathological Society: St. Paul, MN, USA, 2001; pp. 28–29. [Google Scholar]
- Ondřej, M.; Dostálová, R.; Odstrčilová, L. Response of Pisum sativum germplasm resistant to Erysiphe pisi to inoculation with Erysiphe baeumleri, a new pathogen of pea. Plant Prot. Sci. 2005, 41, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Attanayake, R.N.; Glawe, D.A.; Dugan, F.M.; Chen, W. Erysiphe trifolii causing powdery mildew of lentil (Lens culinaris). Plant Dis. 2009, 93, 797–803. [Google Scholar] [CrossRef] [Green Version]
- Attanayake, R.N.; Glawe, D.A.; McPhee, K.E.; Dugan, F.M.; Chen, W. Erysiphe trifolii—A newly recognized powdery mildew pathogen of pea. Plant Pathol. 2010, 59, 712–720. [Google Scholar] [CrossRef]
- Takamatsu, S.; Shin, H.D.; Paksiri, U.; Limkaisang, S.; Taguchi, Y.; Binh, N.T.; Sato, Y. Two Erysiphe species associated with recent outbreak of soybean powdery mildew: Results of molecular phylogenetic analysis based on nuclear rDNA sequences. Mycoscience 2002, 43, 333–341. [Google Scholar] [CrossRef]
- Kasettranan, W.; Somta, P.; Srinives, P. Mapping of Quantitative Trait Loci Controlling Powdery Mildew Resistance in Mungbean (Vigna radiata (L.) Wilczek). J. Crop Sci. Biotechnol. 2010, 13, 155–161. [Google Scholar] [CrossRef]
- Jun, T.H.; Mian, M.A.R.; Kang, S.T.; Michel, A.P. Genetic mapping of the powdery mildew resistance gene in soybean PI 567301B. Theor. Appl. Genet. 2012, 125, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Dunleavy, J.M. Soybean Seed Yield Losses Caused by Powdery Mildew 1. Crop Sci. 1978, 18, 337–339. [Google Scholar] [CrossRef]
- Chaitieng, B.; Kaga, A.; Han, O.K.; Wang, X.W.; Wongkaew, S.; Laosuwan, P.; Tomooka, N.; Vaughan, D.A. Mapping a new source of resistance to powdery mildew in mungbean. Plant Breed. 2002, 121, 521–525. [Google Scholar] [CrossRef]
- Warkentin, T.D.; Rashid, K.Y.; Xue, A.G. Fungicidal control of powdery mildew in field pea. Can. J. Plant Sci. 1996, 76, 933–935. [Google Scholar] [CrossRef]
- Macleod, W.; Basandrai, A.; Basandrai, D.; Bakr, A.; Sandhu, J.S. Integrated foliar diseases management of legumes. In Proceedings of the International Conference on Grain Legumes: Quality Improvement, Value Addition and Trade, Kanpur, India, 14–16 February 2009; pp. 143–161. [Google Scholar]
- Lampson, M.A.; Kapoor, T.M. Unraveling cell division mechanisms with small-molecule inhibitors. Nat. Chem. Biol. 2006, 2, 19–27. [Google Scholar] [CrossRef]
- Richmond, D.V.; Phillips, A. The effect of benomyl and carbendazim on mitosis in hyphae of Botrytis cinerea Pers. ex Fr. and roots of Allium cepa L. Pestic. Biochem. Physiol. 1975, 5, 367–379. [Google Scholar] [CrossRef]
- Hippe, S.; Giesen, U. The effect of triadimenol on the cytology and growth of sensitive and resistant strains of Ustilago avenue. Ann. Appl. Biol. 1988, 112, 79–90. [Google Scholar] [CrossRef]
- Brown, I.F.; Taylor, H.M.; Hall, H.R. EL-222, a new fungicide with eradicative activity. Proc. Am. Phytopathol. Soc. 1975, 2, 31. [Google Scholar]
- Huggenberger, F.; Collins, M.A.; Skylakakis, G. Decreased sensitivity of Sphaerotheca fuliginea to fenarimol and other ergosterol-biosynthesis inhibitors. Crop Prot. 1984, 3, 137–149. [Google Scholar] [CrossRef]
- Baloch, R.I.; Mercer, E.I.; Wiggins, T.E.; Baldwin, B.C. Inhibition of ergosterol biosynthesis in Saccharomyces cerevisiae and Ustilago maydis by tridemorph, fenpropimorph and fenpropidin. Phytochemistry 1984, 23, 2219–2226. [Google Scholar] [CrossRef]
- Caux, P.Y.; Kent, R.A.; Fan, G.T.; Stephenson, G.L. Environmental fate and effects of chlorothalonil: A Canadian perspective. Crit. Rev. Environ. Sci. Technol. 1996, 26, 45–93. [Google Scholar] [CrossRef]
- Barak, E.; Edgington, L.V. Cross-resistance of botrytis cinerea to captan, thiram, chlorothalonil, and related fungicides. Can. J. Plant Pathol. 1984, 6, 318–320. [Google Scholar] [CrossRef]
- Bélanger, R.R.; Labbé, C. Control of powdery mildews without chemicals: Prophylactic and biological alternatives for horticultural crops. In The Powdery Mildews: A Comprehensive Treatise; Bélanger, R.R., Bushnell, W.R., Dik, A.J., Carver, T.L.W., Eds.; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2002; pp. 256–267. [Google Scholar]
- Pandey, M.K.; Sarma, B.K.; Singh, D.P.; Singh, U.P. Biochemical investigations of sclerotial exudates of Sclerotium rolfsii and their antifungal activity. J. Phytopathol. 2007, 155, 84–89. [Google Scholar] [CrossRef]
- Segarra, G.; Reis, M.; Casanova, E.; Trillas, M.I. Control of powdery mildew (Erysiphe polygoni) in tomato by foliar applications of compost tea. J. Plant Pathol. 2009, 683–689. [Google Scholar]
- Patil, N.B.; Zacharia, S.; Kumari, M. Eco-friendly management of downy mildew of Garden pea (Pisum sativum L.). Ecol. Environ. Conserv. 2016, 22, 691–694. [Google Scholar] [CrossRef] [Green Version]
- Maurya, S.; Singh, D.P.; Srivastava, J.S.; Singh, V.P. Effect of some plant extracts on pea powdery mildew (Erysiphe pisi). Ann. Plant Prot. Sci. 2004, 12, 288–291. [Google Scholar]
- Prithiviraj, B.; Singh, U.P.; Manickam, M.; Srivastava, J.S.; Ray, A.B. Antifungal activity of bergenin, a constituent of Flueggea microcarpa. Plant Pathol. 1997, 46, 224–228. [Google Scholar] [CrossRef]
- Prithiviraj, B.; Singh, U.P.; Singh, K.P.; Plank-Schumacher, K. Field evaluation of ajoene, a constituent of garlic (Allium sativum) and neemazal, a product of neem (Azadirachta indica) for the control of powdery mildew (Erysiphe pisi) of pea (Pisum sativum)/Bewertung von Ajoen, einem Inhaltsstoff des Knoblauchs (Allium sativum), und Neemazal, einem Produkt aus Niem (Azadirachta indica), zur Bekämpfung des Echten Mehltaus (Erysiphe pisi) an Erbsen (Pisum sativum) im Freiland. Z. Pflanzenkrankh. Pflanzenschutz J. Plant Dis. Prot. 1998, 105, 274–278. [Google Scholar]
- Vechet, L.; Martinkova, J.; Sindelarova, M.; Burketova, L. Compounds of natural origin inducing winter wheat resistance to powdery mildew (Blumeria graminis f. sp. tritici). Plant Soil Environ. 2005, 51, 468. [Google Scholar]
- Vimala, R.; Suriachandraselvan, M. Induced resistance in bhendi against powdery mildew by foliar application of salicylic acid. J. Biopestic. 2009, 2, 111–114. [Google Scholar]
- Faoro, F.; Maffi, D.; Cantu, D.; Iriti, M. Chemical-induced resistance against powdery mildew in barley: The effects of chitosan and benzothiadiazole. Biocontrol 2008, 53, 387–401. [Google Scholar] [CrossRef]
- Kiss, L. A review of fungal antagonists of powdery mildews and their potential as biocontrol agents. Pest Manag. Sci. 2003, 59, 475–483. [Google Scholar] [CrossRef]
- Paulitz, T.C.; Bélanger, R.R. Biological control in greenhouse systems. Annu. Rev. Phytopathol. 2001, 39, 103–133. [Google Scholar] [CrossRef] [PubMed]
- Elad, Y. Biological control of foliar pathogens by means of Trichoderma harzianum and potential modes of action. Crop Prot. 2000, 19, 709–714. [Google Scholar] [CrossRef]
- Duzan, H.M.; Mabood, F.; Zhou, X.; Souleimanov, A.; Smith, D.L. Nod factor induces soybean resistance to powdery mildew. Plant Physiol. Biochem. 2005, 43, 1022–1030. [Google Scholar] [CrossRef]
- Smigielski, L.; Laubach, E.M.; Pesch, L.; Glock, J.M.L.; Albrecht, F.; Slusarenko, A.; Panstruga, R.; Kuhn, H. Nodulation induces systemic resistance of Medicago truncatula and Pisum sativum against erysiphe pisi and primes for powdery mildew-triggered salicylic acid accumulation. Mol. Plant-Microbe Interact. 2019, 32, 1243–1255. [Google Scholar] [CrossRef]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Gianinazzi, S.; Gianinazzi-Pearson, V. Genes involved in resistance to powdery mildew in barley differentially modulate root colonization by the mycorrhizal fungus Glomus mosseae. Mycorrhiza 1999, 9, 237–240. [Google Scholar] [CrossRef]
- Mustafa, G.; Khong, N.G.; Tisserant, B.; Randoux, B.; Fontaine, J.; Magnin-Robert, M.; Reignault, P.; Sahraoui, A.L.H. Defence mechanisms associated with mycorrhiza-induced resistance in wheat against powdery milde. Funct. Plant Biol. 2017, 44, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.P.; Srivastava, J.S.; Bahadur, A.; Singh, U.P.; Singh, S.K. Arbuscular mycorrhizal fungi induced biochemical changes in pea (Pisum sativum) and their effect on powdery mildew (Erysiphe pisi)/Arbusculäre Mykorrhizapilze induzieren biochemische Veränderungen in Erbsen (Pisum sativum) und ihre Wirkung auf Echten Mehl. Z. Pflanzenkrankh. Pflanzenschutz J. Plant Dis. Prot. 2004, 111, 266–272. [Google Scholar]
- Stein, E.; Molitor, A.; Kogel, K.H.; Waller, F. Systemic resistance in Arabidopsis conferred by the mycorrhizal fungus Piriformospora indica requires jasmonic acid signaling and the cytoplasmic function of NPR1. Plant Cell Physiol. 2008, 49, 1747–1751. [Google Scholar] [CrossRef]
- Mustafa, G.; Randoux, B.; Tisserant, B.; Fontaine, J.; Magnin-Robert, M.; Lounès-Hadj Sahraoui, A.; Reignault, P. Phosphorus supply, arbuscular mycorrhizal fungal species, and plant genotype impact on the protective efficacy of mycorrhizal inoculation against wheat powdery mildew. Mycorrhiza 2016, 26, 685–697. [Google Scholar] [CrossRef]
- Larsen, J.; Yohalem, D. Interactions between mycorrhiza and powdery mildew of cucumber. Mycol. Prog. 2004, 3, 123–128. [Google Scholar] [CrossRef]
- Liu, Y.; Feng, X.; Gao, P.; Li, Y.; Christensen, M.J.; Duan, T. Arbuscular mycorrhiza fungi increased the susceptibility of Astragalus adsurgens to powdery mildew caused by Erysiphe pisi. Mycology 2018, 9, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Hafez, Y.M.; Attia, K.A.; Kamel, S.; Alamery, S.F.; El-Gendy, S.; Al-Doss, A.A.; Mehiar, F.; Ghazy, A.I.; Ibrahim, E.I.; Abdelaal, K.A.A. Bacillus subtilis as a bio-agent combined with nano molecules can control powdery mildew disease through histochemical and physiobiochemical changes in cucumber plants. Physiol. Mol. Plant Pathol. 2020, 111, 101489. [Google Scholar] [CrossRef]
- García-Gutiérrez, L.; Zeriouh, H.; Romero, D.; Cubero, J.; de Vicente, A.; Pérez-García, A. The antagonistic strain B acillus subtilis UMAF 6639 also confers protection to melon plants against cucurbit powdery mildew by activation of jasmonate-and salicylic acid-dependent defence responses. Microb. Biotechnol. 2013, 6, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Gong, Y.; Huo, Y.; Han, Q.; Kang, Z.; Huang, L. Endophytic Bacillus subtilis strain E1R-J is a promising biocontrol agent for wheat powdery mildew. Biomed Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Fukuda, M.; Amaki, Y.; Sakaguchi, T.; Inai, K.; Ishihara, A.; Nakajima, H. Importance of prumycin produced by Bacillus amyloliquefaciens SD-32 in biocontrol against cucumber powdery mildew disease. Pest Manag. Sci. 2017, 73, 2419–2428. [Google Scholar] [CrossRef]
- Hafez, Y.M.; El-Nagar, A.S.; Elzaawely, A.A.; Kamel, S.; Maswada, H.F. Biological control of Podosphaera xanthii the causal agent of squash powdery mildew disease by upregulation of defense-related enzymes. Egypt. J. Biol. Pest Control 2018, 28, 1–8. [Google Scholar] [CrossRef]
- Janousek, C.N.; Lorber, J.D.; Gubler, W.D. Combination and rotation of bacterial antagonists to control powdery mildew on pumpkin. J. Plant Dis. Prot. 2009, 116, 260–262. [Google Scholar] [CrossRef]
- Raguchander, T.; Prabakar, K.; Samiyappan, R. Held evaluation of Pseudomonas fluorescens and Bacillus subtius on the management of Cercospora leaf spot and powdery mildew in urdbean. Legum. Res. Int. J. 2005, 28, 137–139. [Google Scholar]
- Singh, U.P.; Prithiviraj, B.; Singh, K.P.; Sarma, B.K. Control of powdery mildew (Erysiphe pisi) of pea (Pisum sativum) by combined application of plant growth-promoting rhizobacteria and NeemazalTM. Z. Pflanzenkrankh. Pflanzenschutz 2000, 107, 59–66. [Google Scholar]
- Ghebrial, E.; El-Abeid, S.; Aref, E. Bio-Control of Chamomile Powdery Mildew Using Cyanobacteria and Some Antagonistic Microorganisms. Egypt. J. Phytopathol. 2019, 47, 53–77. [Google Scholar] [CrossRef]
- Whipps, J.M. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 2001, 52, 487–511. [Google Scholar] [CrossRef]
- Bélanger, R.R.; Labbé, C.; Lefebvre, F.; Teichmann, B. Mode of action of biocontrol agents: All that glitters is not gold. Can. J. Plant Pathol. 2012, 34, 469–478. [Google Scholar] [CrossRef]
- Laur, J.; Ramakrishnan, G.B.; Labbé, C.; Lefebvre, F.; Spanu, P.D.; Bélanger, R.R. Effectors involved in fungal–fungal interaction lead to a rare phenomenon of hyperbiotrophy in the tritrophic system biocontrol agent–powdery mildew–plant. New Phytol. 2018, 217, 713–725. [Google Scholar] [CrossRef] [Green Version]
- Considine, M.J.; Siddique, K.H.M.; Foyer, C.H. Nature’s pulse power: Legumes, food security and climate change. J. Exp. Bot. 2017, 68, 1815–1818. [Google Scholar] [CrossRef]
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. Agric. 2017, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Siddique, K.H.M.; Johansen, C.; Turner, N.C.; Jeuffroy, M.H.; Hashem, A.; Sakar, D.; Gan, Y.; Alghamdi, S.S. Innovations in agronomy for food legumes. A review. Agron. Sustain. Dev. 2012, 32, 45–64. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Lam, H.M.; Nguyen, H.T.; Siddique, K.H.M.; Varshney, R.K.; Colmer, T.D.; Cowling, W.; Bramley, H.; Mori, T.A.; Hodgson, J.M.; et al. Neglecting legumes has compromised human health and sustainable food production. Nat. Plants 2016, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, N.A. Plant disease susceptibility genes? Plant Cell 2002, 14, 1983–1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavan, S.; Jacobsen, E.; Visser, R.G.F.; Bai, Y. Loss of susceptibility as a novel breeding strategy for durable and broad-spectrum resistance. Mol. Breed. 2010, 25, 1. [Google Scholar] [CrossRef] [Green Version]
- De Almeida Engler, J.; Favery, B.; Engler, G.; Abad, P. Loss of susceptibility as an alternative for nematode resistance. Curr. Opin. Biotechnol. 2005, 16, 112–117. [Google Scholar] [CrossRef]
- Boyd, L.A.; Smith, P.H.; Foster, E.M.; Brown, J.K.M. The effects of allelic variation at the Mla resistance locus in barley on the early development of Erysiphe graminis f. sp. hordei and host responses. Plant J. 1995, 7, 959–968. [Google Scholar] [CrossRef]
- Jørgensen, J.H. Genetics of powdery mildew resistance in barley. Critical Reviews in. Plant Sci. 1994, 13, 97–119. [Google Scholar]
- Halterman, D.; Zhou, F.; Wei, F.; Wise, R.P.; Schulze-Lefert, P. The MLA6 coiled-coil, NBS-LRR protein confers AvrMla6-dependent resistance specificity to Blumeria graminis f. sp. hordei in barley and wheat. Plant J. 2001, 25, 335–348. [Google Scholar] [CrossRef] [Green Version]
- Halterman, D.A.; Wise, R.P. A single-amino acid substitution in the sixth leucine-rich repeat of barley MLA6 and MLA13 alleviates dependence on RAR1 for disease resistance signaling. Plant J. 2004, 38, 215–226. [Google Scholar] [CrossRef]
- Jørgensen, I.H. Discovery, characterization and exploitation of Mlo powdery mildew resistance in barley. Euphytica 1992, 63, 141–152. [Google Scholar] [CrossRef]
- Büschges, R.; Hollricher, K.; Panstruga, R.; Simons, G.; Wolter, M.; Frijters, A.; Van Daelen, R.; Van der Lee, T.; Diergaarde, P.; Groenendijk, J.; et al. The barley Mlo gene: A novel control element of plant pathogen resistance. Cell 1997, 88, 695–705. [Google Scholar] [CrossRef] [Green Version]
- Devoto, A.; Hartmann, H.A.; Piffanelli, P.; Elliott, C.; Simmons, C.; Taramino, G.; Goh, C.S.; Cohen, F.E.; Emerson, B.C.; Schulze-Lefert, P.; et al. Molecular phylogeny and evolution of the plant-specific seven-transmembrane MLO family. J. Mol. Evol. 2003, 56, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Devoto, A.; Piffanelli, P.; Nilsson, I.M.; Wallin, E.; Panstruga, R.; Von Heijne, G.; Schulze-Lefert, P. Topology, subcellular localization, and sequence diversity of the Mlo family in plants. J. Biol. Chem. 1999, 274, 34993–35004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Hartmann, H.A.; Wu, M.-J.; Friedman, E.J.; Chen, J.-G.; Pulley, M.; Schulze-Lefert, P.; Panstruga, R.; Jones, A.M. Expression analysis of the AtMLO gene family encoding plant-specific seven-transmembrane domain proteins. Plant Mol. Biol. 2006, 60, 583–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feechan, A.; Jermakow, A.M.; Torregrosa, L.; Panstruga, R.; Dry, I.B. Identification of grapevine MLO gene candidates involved in susceptibility to powdery mildew. Funct. Plant Biol. 2008, 35, 1255–1266. [Google Scholar] [CrossRef]
- Zhou, S.J.; Jing, Z.; Shi, J.L. Genome-wide identification, characterization, and expression analysis of the MLO gene family in Cucumis sativus. Genet. Mol. Res. 2013, 12, 6565–6578. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Y.; Zhang, H. Genome-wide analysis of the mildew resistance locus o (‘MLO’) gene family in tomato (‘Solanum lycopersicum’ L.). Plant Omics 2014, 7, 87–93. [Google Scholar]
- Iovieno, P.; Bracuto, V.; Pavan, S.; Lotti, C.; Ricciardi, L.; Andolfo, G. Identification and functional inference on the MLO-family in viridiplantae. J. Plant Pathol. 2016, 98, 587–594. [Google Scholar] [CrossRef]
- Rispail, N.; Rubiales, D. Genome-wide identification and comparison of legume MLO gene family. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wolter, M.; Hollricher, K.; Salamini, F.; Schulze-Lefert, P. The mlo resistance alleles to powdery mildew infection in barley trigger a developmentally controlled defence mimic phenotype. Mol. Gen. Genet. 1993, 239, 122–128. [Google Scholar] [CrossRef]
- Peterhänsel, C.; Freialdenhoven, A.; Kurth, J.; Kolsch, R.; Schulze-Lefert, P. Interaction analyses of genes required for resistance responses to powdery mildew in barley reveal distinct pathways leading to leaf cell death. Plant Cell 1997, 9, 1397–1409. [Google Scholar] [CrossRef]
- Kusch, S.; Panstruga, R. Mlo-based resistance: An apparently universal “weapon” to defeat powdery mildew disease. Mol. Plant-Microbe Interact. 2017, 30, 179–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.; Deng, D.; Duan, C.; Zong, X.; Xu, D.; He, Y.; Zhu, Z. Two novel er1 alleles conferring powdery mildew (Erysiphe pisi) resistance identified in a worldwide collection of pea (Pisum sativum L.) germplasms. Int. J. Mol. Sci. 2019, 20, 5071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.; Deng, D.; Wang, Z.; Duan, C.; Wu, X.; Wang, X.; Zong, X.; Zhu, Z. A novel er1 allele and the development and validation of its functional marker for breeding pea (Pisum sativum L.) resistance to powdery mildew. Theor. Appl. Genet. 2016, 129, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Fu, H.; Sun, S.; Duan, C.; Wu, X.; Yang, X.; Zhu, Z. Identification of powdery mildew resistance gene in pea line X9002. ACTA Agron. Sin. 2015, 41, 515–523. [Google Scholar] [CrossRef]
- Sudheesh, S.; Lombardi, M.; Leonforte, A.; Cogan, N.O.I.; Materne, M.; Forster, J.W.; Kaur, S. Consensus Genetic Map Construction for Field Pea (Pisum sativum L.), Trait Dissection of Biotic and Abiotic Stress Tolerance and Development of a Diagnostic Marker for the er1 Powdery Mildew Resistance Gene. Plant Mol. Biol. Report. 2015, 33, 1391–1403. [Google Scholar] [CrossRef]
- Ma, Y.; Coyne, C.J.; Main, D.; Pavan, S.; Sun, S.; Zhu, Z.; Zong, X.; Leitão, J.; McGee, R.J. Development and validation of breeder-friendly KASPar markers for er1, a powdery mildew resistance gene in pea (Pisum sativum L.). Mol. Breed. 2017, 37, 151. [Google Scholar] [CrossRef]
- Pavan, S.; Schiavulli, A.; Appiano, M.; Miacola, C.; Visser, R.G.F.; Bai, Y.; Lotti, C.; Ricciardi, L. Identification of a complete set of functional markers for the selection of er1 powdery mildew resistance in Pisum sativum L. Mol. Breed. 2013, 31, 247–253. [Google Scholar] [CrossRef]
- Ambrose, M.J. Screening for field resistance to Powdery Mildew (Eyrsiphe polygoni D. C.) in the JI Pisum Collection. Pisum Genet 2009, 41, 40–43. [Google Scholar]
- Santo, T.; Rashkova, M.; Alabaça, C.; Leitão, J. The ENU-induced powdery mildew resistant mutant pea (Pisum sativum L.) lines S(er1mut1) and F(er1mut2) harbour early stop codons in the PsMLO1 gene. Mol. Breed. 2013, 32, 723–727. [Google Scholar] [CrossRef]
- Fu, H.; Sun, S.; Zhu, Z.; Duan, C.; Yang, X. Phenotypic and genotypic identification of powdery mildew resistance in pea cultivars or lines from Canada. J. Plant Genet. Resour. 2014, 15, 1028–1033. [Google Scholar]
- Sun, S.; Fu, H.; Wang, Z.; Duan, C.; Zong, X.; Zhu, Z. Discovery of a novel er1 allele conferring powdery mildew resistance in Chinese Pea (Pisum sativum L.) landraces. PLoS ONE 2016, 11, e0147624. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Wang, Z.; Fu, H.; Duan, C.; Wang, X.; Zhu, Z. Resistance to powdery mildew in the pea cultivar Xucai 1 is conferred by the gene er1. Crop J. 2015, 3, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Binagwa, P.H.; Traore, S.M.; Egnin, M.; Bernard, G.C.; Ritte, I.; Mortley, D.; Kamfwa, K.; He, G.; Bonsi, C. Genome-Wide Identification of Powdery Mildew Resistance in Common Bean (Phaseolus vulgaris L.). Front. Genet. 2021, 12, 673069. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, R.; Singh, V.K.; Singh, B.D. Comparative phylogenetic analysis of genome-wide Mlo gene family members from Glycine max and Arabidopsis thaliana. Mol. Genet. Genom. 2014, 289, 345–359. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Zhao, J.; Du, C.; Xiang, Y.; Cao, J.; Qin, X. Genome-scale identification of MLO domain-containing genes in soybean (Glycine max L. Merr.). Genes Genet. Syst. 2012, 87, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Chankaew, S.; Somta, P.; Isemura, T.; Tomooka, N.; Kaga, A.; Vaughan, D.A.; Srinives, P. Quantitative trait locus mapping reveals conservation of major and minor loci for powdery mildew resistance in four sources of resistance in mungbean [Vigna radiata (L.) Wilczek]. Mol. Breed. 2013, 32, 121–130. [Google Scholar] [CrossRef]
- Parkhi, V.; Bhattacharya, A.; Char, B. Multiomics Technologies and Genetic Modification in Plants: Rationale, Opportunities and Reality. In Omics Technologies for Sustainable Agriculture and Global Food Security (Vol II); Kumar, A., Kumar, R., Shukla, P., Patel, H.K., Eds.; Springer: Singapore, 2021; Volume II, pp. 313–328. [Google Scholar]
- Prats, E.; Llamas, M.J.; Rubiales, D. Characterization of resistance mechanisms to Erysiphe pisi in Medicago truncatula. Phytopathology 2007, 97, 1049–1053. [Google Scholar] [CrossRef] [Green Version]
- Yaege, J.R.; Stuteville, D.L. Reactions of accessions in the annual Medicago core germ plasm collection to Erysiphe pisi. Plant Dis. 2002, 86, 312–315. [Google Scholar] [CrossRef] [Green Version]
- Ameline-Torregrosa, C.; Cazaux, M.; Danesh, D.; Chardon, F.; Cannon, S.B.; Esquerré-Tugayé, M.T.; Dumas, B.; Young, N.D.; Samac, D.A.; Huguet, T.; et al. Genetic dissection of resistance to anthracnose and powdery mildew in Medicago truncatula. Mol. Plant-Microbe Interact. 2008, 21, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Tang, F.; Caixeta, E.T.; Zhu, H. Epigenetic regulation of a powdery mildew resistance gene in medicago truncatula. Mol. Plant 2013, 6, 2000–2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohnes, D.G.; Wagner, R.E.; Bernard, R.L. Soybean genes Rj2, Rmd, and Rps2 in linkage group 19. J. Hered. 1993, 84, 109–111. [Google Scholar] [CrossRef]
- Polzin, K.M.; Lohnes, D.G.; Nickell, C.D.; Shoemaker, R.C. Integration of rps2, rmd, and rj2 into linkage group j of the soybean molecular map. J. Hered. 1994, 85, 300–303. [Google Scholar] [CrossRef]
- Lohnes, D.G.; Nickell, C.D. Effects of powdery mildew alleles Rmd-c, Rmd and rmd on yield and other characteristics in soybean. Plant Dis. 1994, 78, 299–301. [Google Scholar] [CrossRef]
- Kang, S.T.; Rouf Mian, M.A. Genetic map of the powdery mildew resistance gene in soybean PI 243540. Genome 2010, 53, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, R.C.; Schlueter, J.; Doyle, J.J. Paleopolyploidy and gene duplication in soybean and other legumes. Curr. Opin. Plant Biol. 2006, 9, 104–109. [Google Scholar] [CrossRef]
- Walling, J.G.; Shoemaker, R.; Young, N.; Mudge, J.; Jackson, S. Chromosome-level homeology in paleopolyploid soybean (Glycine max) revealed through integration of genetic and chromosome maps. Genetics 2006, 172, 1893–1900. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, R.; Singh, V.K.; Singh, B.D. Comparative analysis of genome-wide Mlo gene family in Cajanus cajan and Phaseolus vulgaris. Genetica 2016, 144, 229–241. [Google Scholar] [CrossRef]
- Polanco, C.; Sáenz De Miera, L.E.; Bett, K.; De La Vega, M.P. A genome-wide identification and comparative analysis of the lentil MLO genes. PLoS ONE 2018, 13, e0194945. [Google Scholar] [CrossRef]
- Yundaeng, C.; Somta, P.; Chen, J.; Yuan, X.; Chankaew, S.; Srinives, P.; Chen, X. Candidate gene mapping reveals VrMLO12 (MLO Clade II) is associated with powdery mildew resistance in mungbean (Vigna radiata [L.] Wilczek). Plant Sci. 2020, 298, 110594. [Google Scholar] [CrossRef]
- Traore, S.M.; Han, S.; Binagwa, P.; Xu, W.; Chen, X.; Liu, F.; He, G. Genome-wide identification of mlo genes in the cultivated peanut (Arachis hypogaea L.). Euphytica 2021, 217, 61. [Google Scholar] [CrossRef]
- Bolay, A.; Braun, U.; Delhey, R.; Kummer, V.; Piatek, M.; Wołczańska, A. Erysiphe deutziae—A new epidemic spread in Europe. Cryptogam. Mycol. 2005, 26, 293–298. [Google Scholar]
- Dynowska, M. A comparison of urban and suburban occurrence of Erysiphales with special emphasis on degree of host infection. Acta Soc. Bot. Pol. 1994, 63, 341–344. [Google Scholar] [CrossRef]
- Smith, J.M. A comment on the Red Queen. Am. Nat. 1976, 110, 325–330. [Google Scholar] [CrossRef]
- Cobos, M.J.; Satovic, Z.; Rubiales, D.; Fondevilla, S. Er3 gene, conferring resistance to powdery mildew in pea, is located in pea LGIV. Euphytica 2018, 214. [Google Scholar] [CrossRef]
- Jha, A.B.; Gali, K.K.; Alam, Z.; Lachagari, V.B.R.; Warkentin, T.D. Potential application of genomic technologies in breeding for fungal and oomycete disease resistance in pea. Agronomy 2021, 11, 1260. [Google Scholar] [CrossRef]
- Kreplak, J.; Madoui, M.A.; Cápal, P.; Novák, P.; Labadie, K.; Aubert, G.; Bayer, P.E.; Gali, K.K.; Syme, R.A.; Main, D.; et al. A reference genome for pea provides insight into legume genome evolution. Nat. Genet. 2019, 51, 1411–1422. [Google Scholar] [CrossRef]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol. Biol. 1985, 5, 69–76. [Google Scholar] [CrossRef]
- Sulima, A.S.; Zhukov, V.A.; Afonin, A.A.; Zhernakov, A.I.; Tikhonovich, I.A.; Lutova, L.A. Selection Signatures in the First Exon of Paralogous Receptor Kinase Genes from the Sym2 Region of the Pisum sativum L. Genome. Front. Plant Sci. 2017, 8, 1957. [Google Scholar] [CrossRef] [Green Version]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef]

| Psmlo1 Allele | Mutation | Cultivar/Landrace | Place of Origin |
|---|---|---|---|
| er1-1 (er1mut1) | C → G at position 680 of cDNA | Mexique 4/JI1559 [14] | Mexico |
| S [142] | Portugal (mutant obtained on the Solara cultivar) | ||
| Tara [143] | Canada (obtained on material from Argentine) | ||
| er1-2 | Large transposon insertion of unknown size in 14th exon (position 1262 of cDNA) leading to formation of various aberrant MLO1 transcripts | Stratagem/JI2302 [14] | Mexico |
| Franklin/PI 628275; Dorian; Nadir [15] | USA (on an unknown material) | ||
| X9002 [137,144]; Xucai 1 [145]; G0005576 [144] | China | ||
| er1-3 | G deletion at position 862 of cDNA | JI0210/Lucknow Boniya/PI 240515 [14] | India |
| er1-4 | A deletion at position 91 of cDNA | YI; JI1951 [14] | China |
| er1-5 | G → A at position 567 of cDNA | ROI3/02 [15] | Italy (mutant obtained on the Sprinter cultivar) |
| er1-6 | T → C at position 1121 of cDNA | Wandou G0001752, Baiwandou G0001763, Dabaiwandou G0001764, Fanwandou G0001767, Wandou G0001768, Dabaiwandou G0001778, Dabaiwandou G0001780, Wandou G0003824 [144] | China |
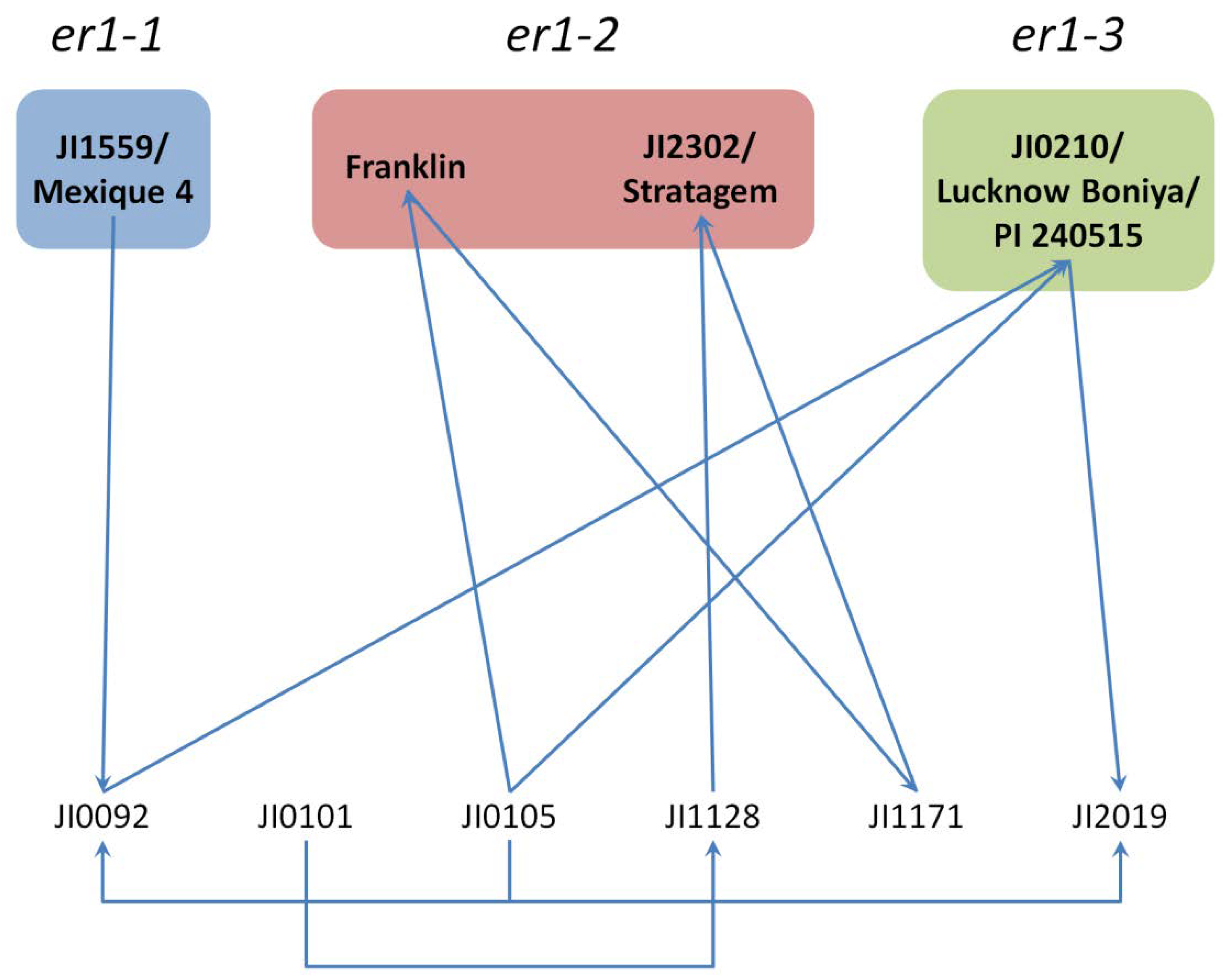
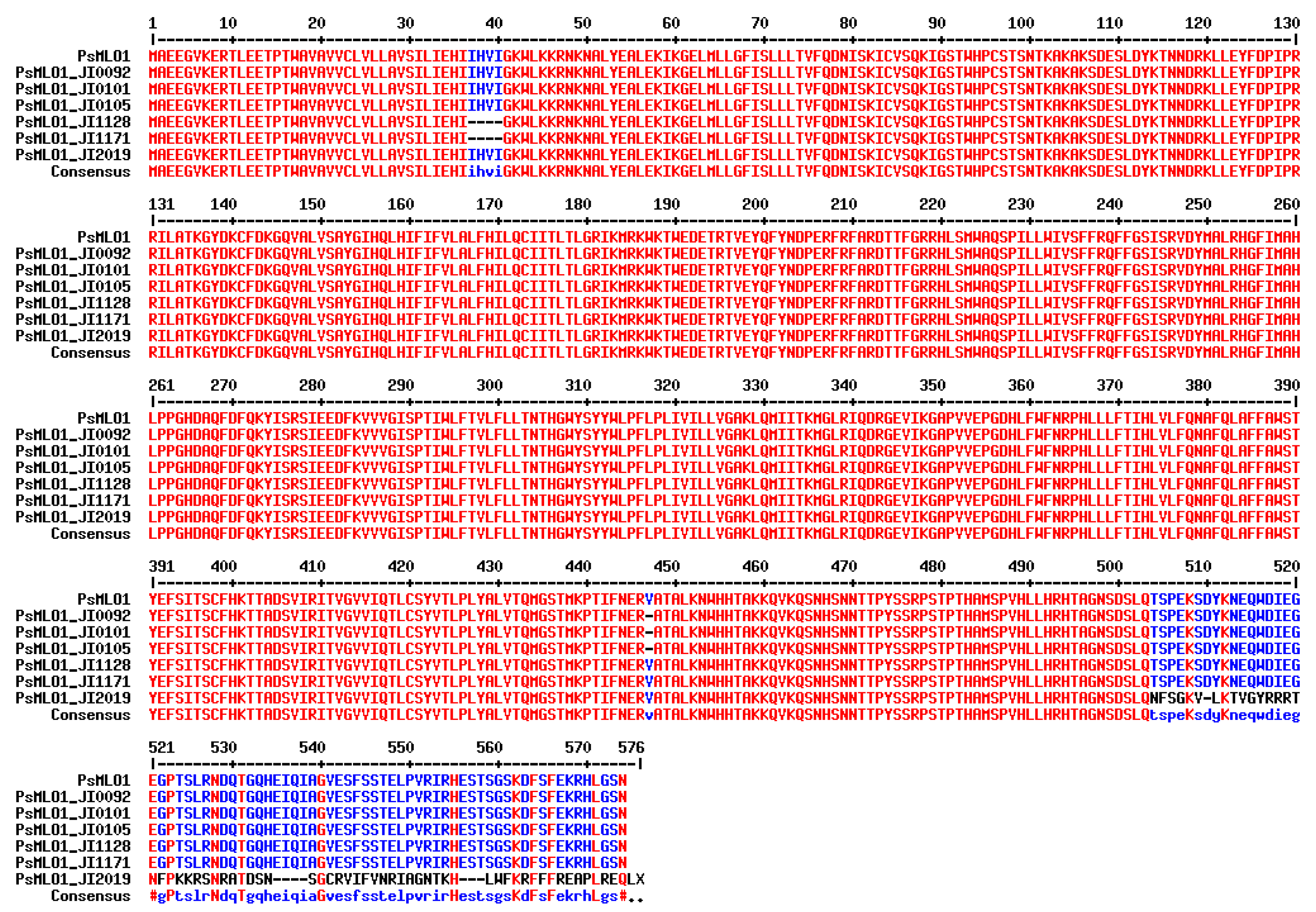
| er1-7 | TCATGTTATT deletion at position 111–120 of cDNA | DDR-11 [136] | India |
| JI1128 (see Appendix A of this article) | India | ||
| JI1171 (see Appendix A of this article) | India | ||
| er1-8 | GTG deletion at position 1339–1341 of cDNA | G0004839 [135] | Afghanistan |
| JI0092/PI 134271 (see Appendix A of this article) | Afghanistan | ||
| JI0101/PI 220175 (see Appendix A of this article) | Afghanistan | ||
| JI0105/PI 222070 (see Appendix A of this article) | Afghanistan | ||
| er1-9 | T deletion at position 928 of cDNA | G0004400 [135] | Australia |
| er1-10 (er1mut2) | G → A at position 939 of cDNA | F [142] | Portugal (mutant obtained on the Frilene cultivar) |
| er1-11 | GT insertion at position 2974–2975 of genomic DNA (intron 11) | Yarrum, ps1771 [138] | Australia |
| er1-12 | A insertion at position 1735 of cDNA | JI2019 (see Appendix A of this article) | India |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sulima, A.S.; Zhukov, V.A. War and Peas: Molecular Bases of Resistance to Powdery Mildew in Pea (Pisum sativum L.) and Other Legumes. Plants 2022, 11, 339. https://doi.org/10.3390/plants11030339
Sulima AS, Zhukov VA. War and Peas: Molecular Bases of Resistance to Powdery Mildew in Pea (Pisum sativum L.) and Other Legumes. Plants. 2022; 11(3):339. https://doi.org/10.3390/plants11030339
Chicago/Turabian StyleSulima, Anton S., and Vladimir A. Zhukov. 2022. "War and Peas: Molecular Bases of Resistance to Powdery Mildew in Pea (Pisum sativum L.) and Other Legumes" Plants 11, no. 3: 339. https://doi.org/10.3390/plants11030339
APA StyleSulima, A. S., & Zhukov, V. A. (2022). War and Peas: Molecular Bases of Resistance to Powdery Mildew in Pea (Pisum sativum L.) and Other Legumes. Plants, 11(3), 339. https://doi.org/10.3390/plants11030339