
Molecular Genetic Diversity and Combining Ability for Some Physiological and Agronomic Traits in Rice under Well-Watered and Water-Deficit Conditions

 ,
, 
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parental Genotypes and Hybridization
2.2. Experimental Site and Agronomic Practices
2.3. Measured Traits
2.3.1. Morphological and Physiological Traits
2.3.2. Grain Yield and Its Related Traits
2.4. Molecular Analysis
2.5. Statistical Analysis
3. Results
3.1. Analysis of Variance
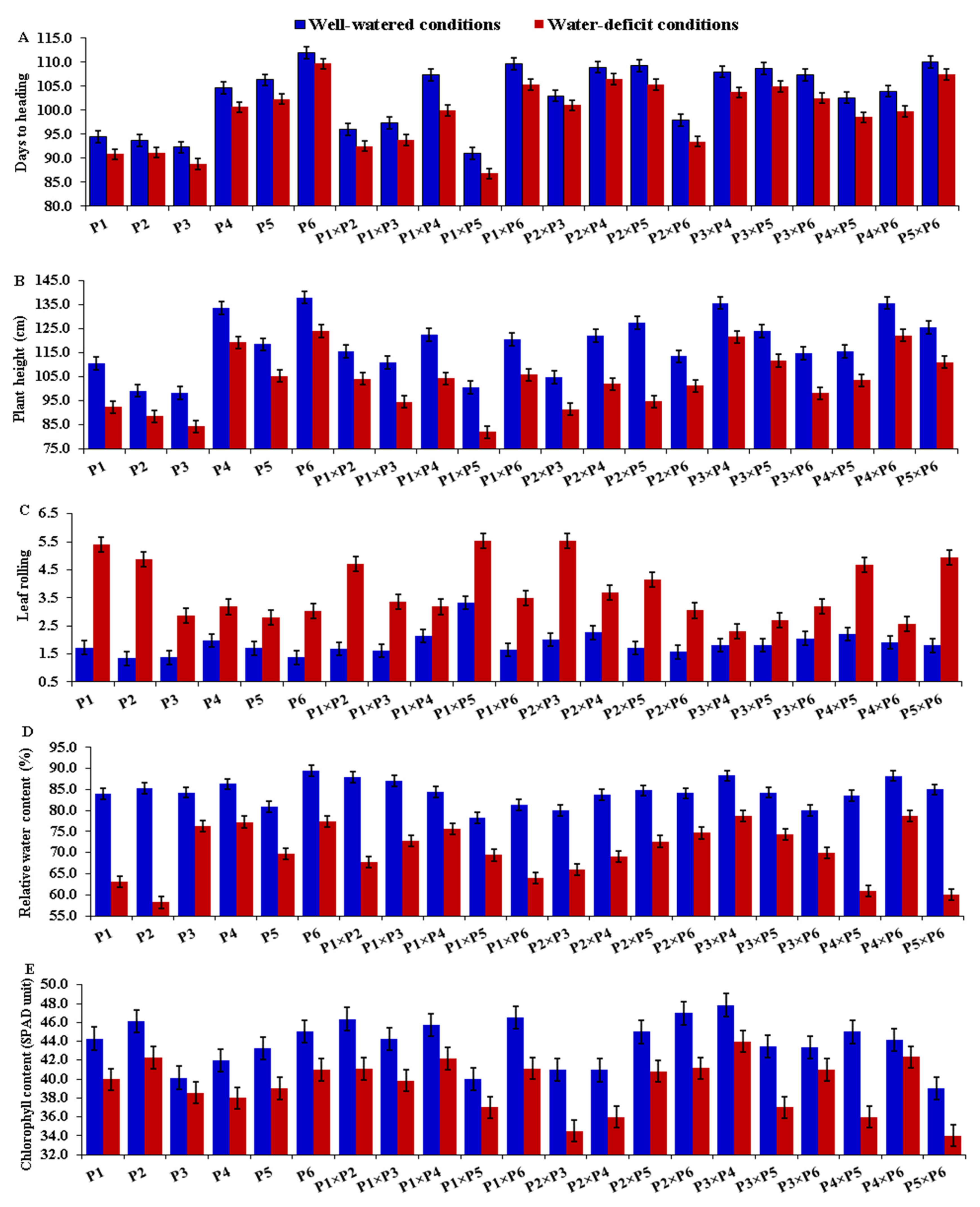
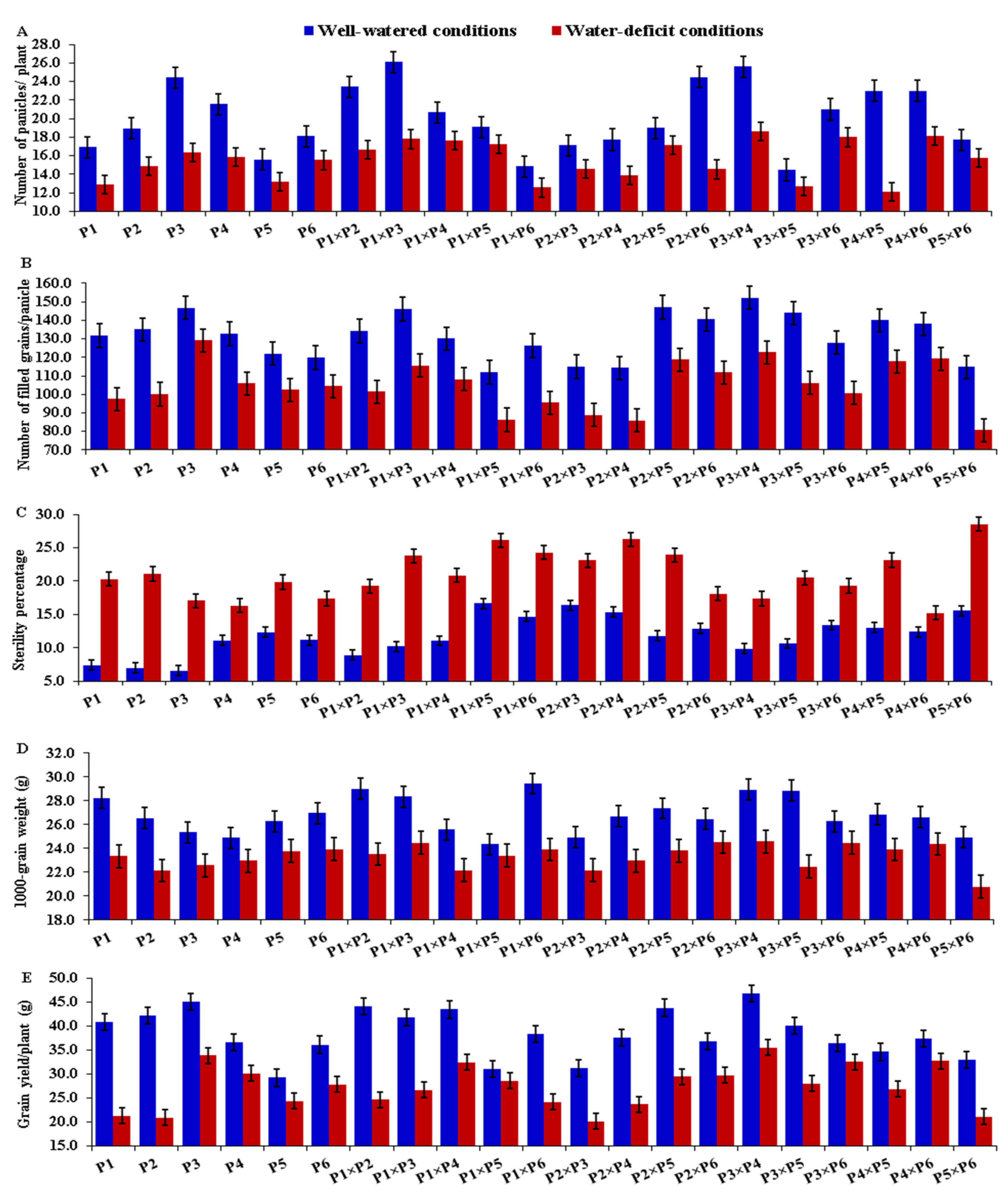
3.2. Mean Performance of Parents and F1 Hybrids
3.3. Genotypic Classification According to Drought Tolerance Indices
3.4. General Combining Ability (GCA) Effects
3.5. Specific Combining Ability (SCA) Effects
3.6. Heterosis Relative to Mid-Parent (MP) and Better-Parent (BP)
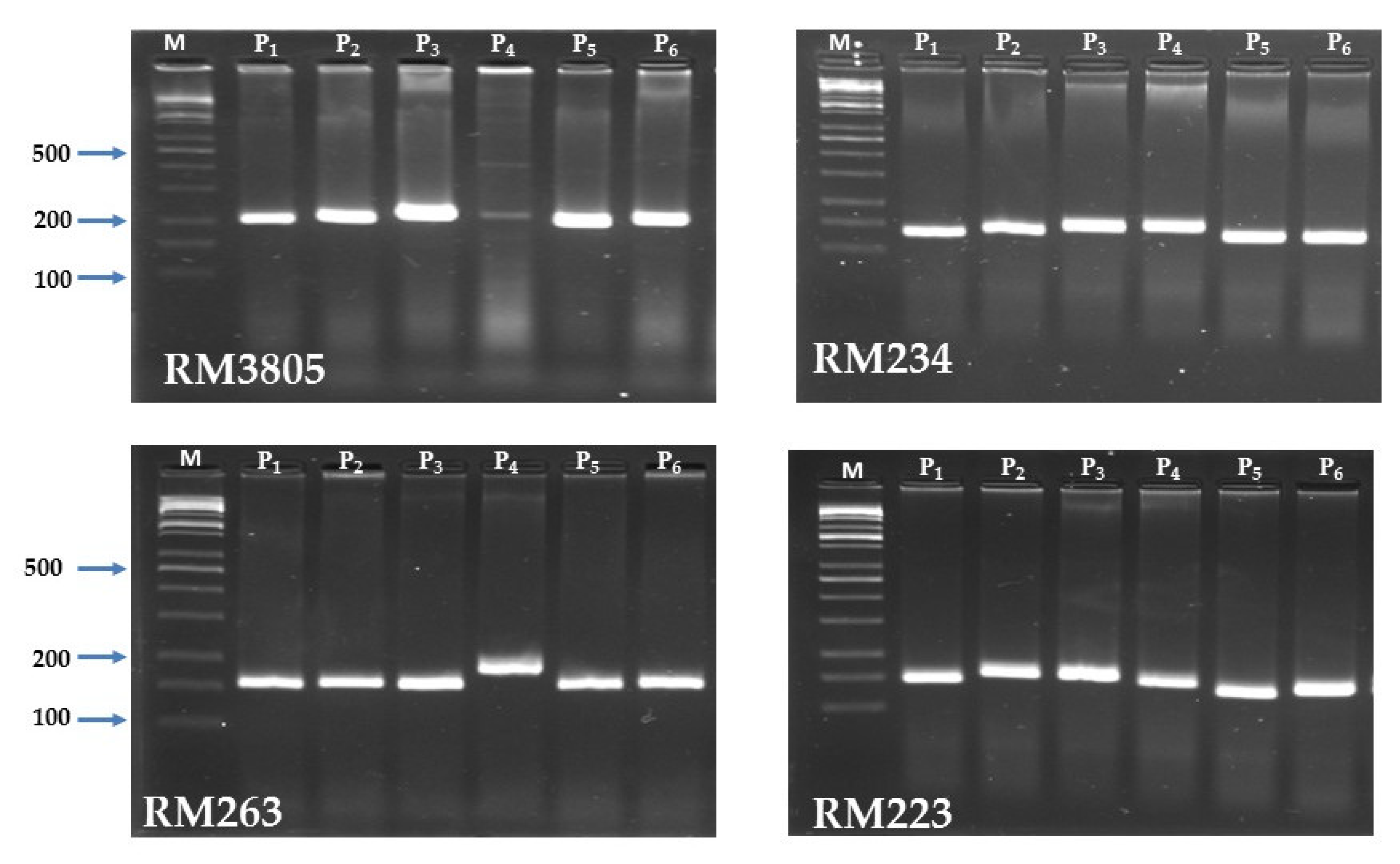
3.7. Microsatellites Based Polymorphism
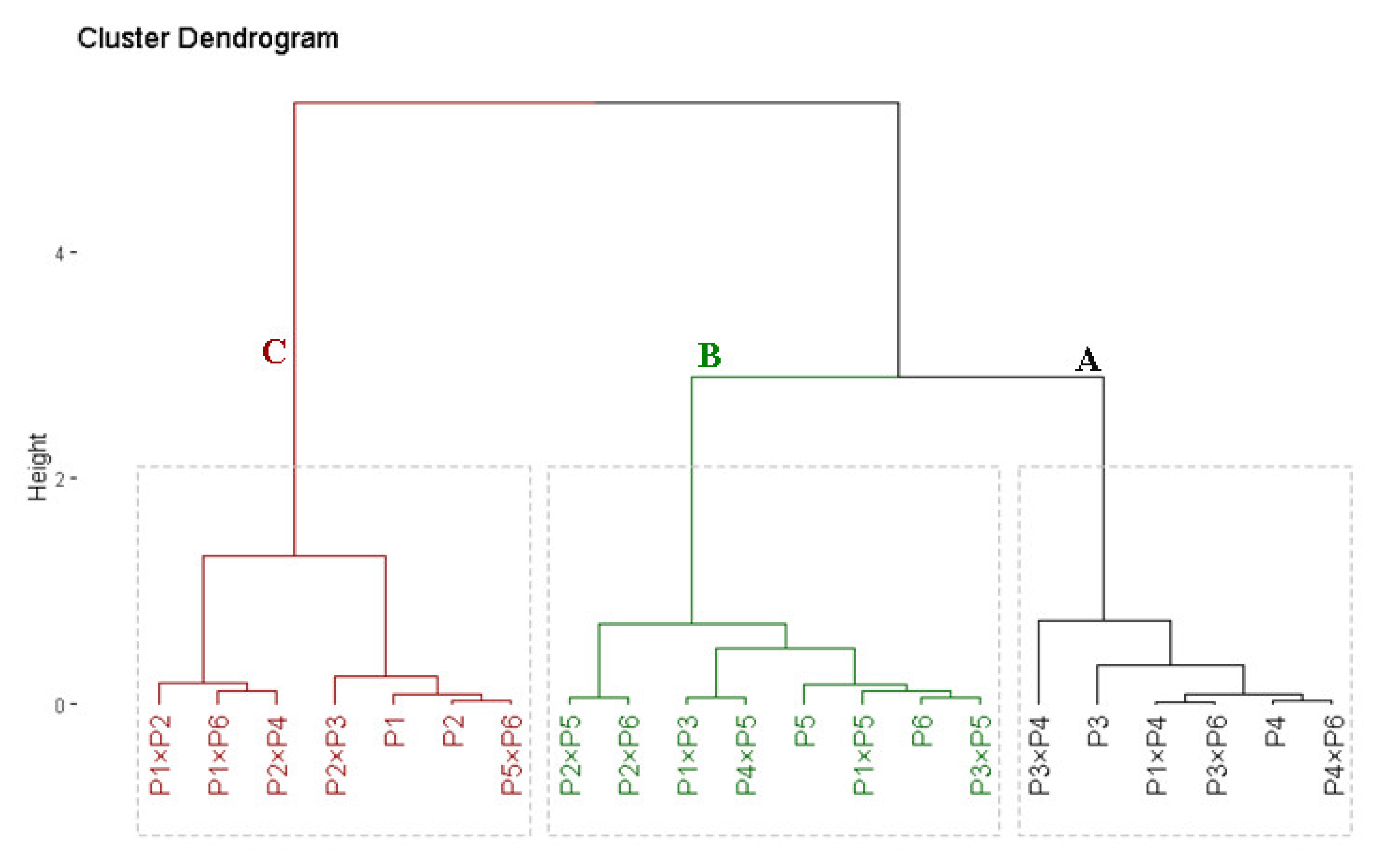
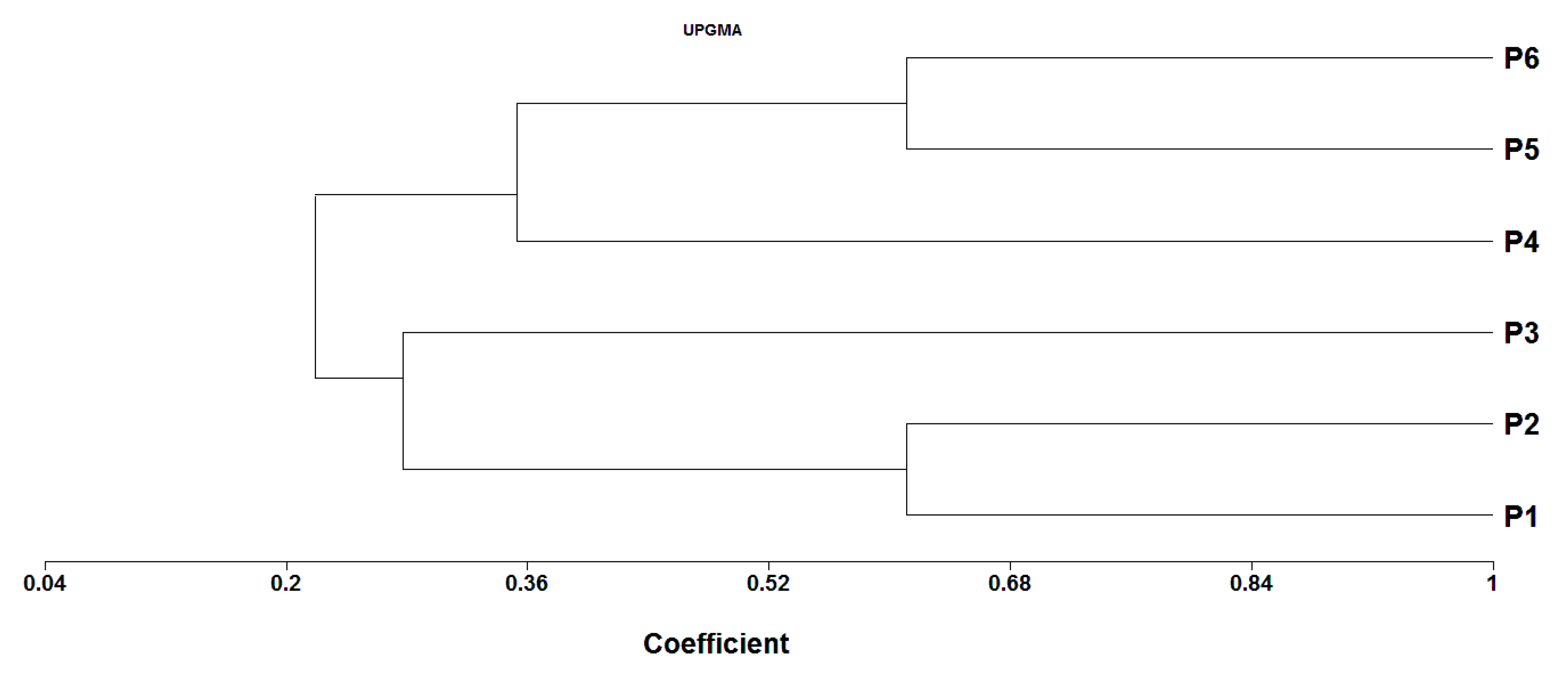
3.8. Genetic Distance (GD) and Cluster Analysis
3.9. Correlation between Parental GD, Hybrid Performance, SCA, and Heterosis
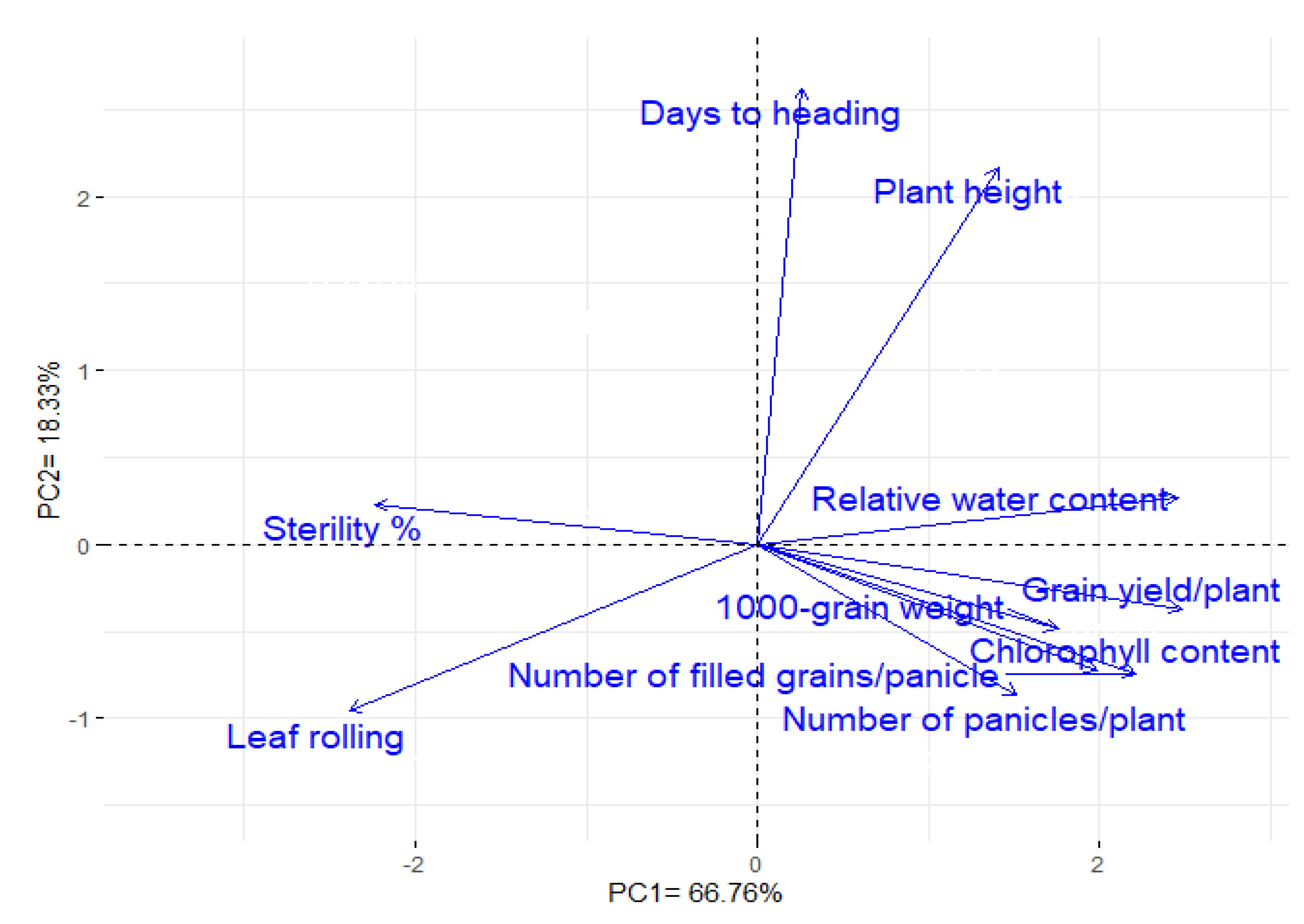
3.10. Interrelationship among the Studied Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khush, G.S. What it will take to feed 5.0 billion rice consumers in 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Naaz, S.; Singh, N.; Shukla, P.K.; Tripathi, R.; Yadav, H.K.; Shirke, P.A. Molecular, physiological and agronomic assessment of genetic diversity in rice varieties in relation to drought treatment. Curr. Plant Biol. 2022, 29, 100232. [Google Scholar] [CrossRef]
- El-Mowafi, H.F.; AlKahtani, M.D.; Abdallah, R.M.; Reda, A.M.; Attia, K.A.; El-Hity, M.A.; El-Dabaawy, H.E.; Husnain, L.A.; Al-Ateeq, T.K.; EL-Esawi, M.A. Combining ability and gene action for yield characteristics in novel aromatic cytoplasmic male sterile hybrid rice under water-stress conditions. Agriculture 2021, 11, 226. [Google Scholar] [CrossRef]
- Faostat. Food and Agriculture Organization of the United Nations. Statistical Database. 2022. Available online: http://www.fao.org/faostat/en/#data (accessed on 15 January 2022).
- Fageria, N. Yield physiology of rice. J. Plant Nutr. 2007, 30, 843–879. [Google Scholar] [CrossRef]
- Hafez, E.M.; Gowayed, S.M.; Nehela, Y.; Sakran, R.M.; Rady, A.; Awadalla, A.; Omara, A.E.-D.; Alowaiesh, B.F. Incorporated biochar-based soil amendment and exogenous glycine betaine foliar application ameliorate rice (Oryza sativa L.) tolerance and resilience to osmotic stress. Plants 2021, 10, 1930. [Google Scholar] [CrossRef]
- Gaballah, M.M.; Ghoneim, A.M.; Ghazy, M.I.; Mohammed, H.M.; Sakran, R.M.; Rehman, H.U.; Shamsudin, N.A.A. Root traits responses to irrigation intervals in rice (Oryza sativa). Int. J. Agric. Biol. 2021, 26, 22–30. [Google Scholar] [CrossRef]
- Yang, X.; Wang, B.; Chen, L.; Li, P.; Cao, C. The different influences of drought stress at the flowering stage on rice physiological traits, grain yield, and quality. Sci. Rep. 2019, 9, 3742. [Google Scholar] [CrossRef] [Green Version]
- Manickavelu, A.; Nadarajan, N.; Ganesh, S.; Gnanamalar, R.; Babu, R.C. Drought tolerance in rice: Morphological and molecular genetic consideration. Plant Growth Regul. 2006, 50, 121–138. [Google Scholar] [CrossRef]
- Mehana, M.; Abdelrahman, M.; Emadeldin, Y.; Rohila, J.S.; Karthikeyan, R. Impact of genetic improvements of rice on its water use and effects of climate variability in Egypt. Agriculture 2021, 11, 865. [Google Scholar] [CrossRef]
- Ghazy, M.I.; Salem, K.F.; Sallam, A. Utilization of genetic diversity and marker-trait to improve drought tolerance in rice (Oryza sativa L.). Mol. Biol. Rep. 2021, 48, 157–170. [Google Scholar] [CrossRef]
- Gracia, M.; Mansour, E.; Casas, A.; Lasa, J.; Medina, B.; Cano, J.L.M.; Moralejo, M.; López, A.; Fuster, P.L.; Escribano, J. Progress in the Spanish national barley breeding program. Span. J. Agric. Res. 2012, 10, 741–751. [Google Scholar] [CrossRef] [Green Version]
- Salem, T.; Rabie, H.; Mowafy, S.; Eissa, A.; Mansour, E. Combining ability and genetic components of egyptian cotton for earliness, yield, and fiber quality traits. SABRAO J. Breed. Genet. 2020, 52, 369–389. [Google Scholar]
- Kamara, M.M.; Ghazy, N.A.; Mansour, E.; Elsharkawy, M.M.; Kheir, A.; Ibrahim, K.M. Molecular genetic diversity and line× tester analysis for resistance to late wilt disease and grain yield in maize. Agronomy 2021, 11, 898. [Google Scholar] [CrossRef]
- Kamara, M.M.; Ibrahim, K.M.; Mansour, E.; Kheir, A.; Germoush, M.O.; El-Moneim, A.; Motawei, M.I.; Alhusays, A.Y.; Farid, M.A.; Rehan, M. Combining ability and gene action controlling grain yield and its related traits in bread wheat under heat stress and normal conditions. Agronomy 2021, 11, 1450. [Google Scholar] [CrossRef]
- Griffing, B. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci. 1956, 9, 463–493. [Google Scholar] [CrossRef]
- Anusha, G.; Rao, D.S.; Jaldhani, V.; Beulah, P.; Neeraja, C.; Gireesh, C.; Anantha, M.; Suneetha, K.; Santhosha, R.; Prasad, A.H. Grain Fe and Zn content, heterosis, combining ability and its association with grain yield in irrigated and aerobic rice. Sci. Rep. 2021, 11, 10579. [Google Scholar] [CrossRef] [PubMed]
- Suvi, W.T.; Shimelis, H.; Laing, M.; Mathew, I.; Shayanowako, A.I. Determining the combining ability and gene action for rice yellow mottle virus disease resistance and agronomic traits in rice (Oryza sativa L.). Agronomy 2021, 11, 12. [Google Scholar] [CrossRef]
- Verma, O.; Srivastava, H. Genetic component and combining ability analyses in relation to heterosis for yield and associated traits using three diverse rice-growing ecosystems. Field Crops Res. 2004, 88, 91–102. [Google Scholar] [CrossRef]
- Mackill, D.; Coffman, W.; Rutger, J. Pollen shedding and combining ability for high temperature tolerance in rice. Crop Sci. 1982, 22, 730–733. [Google Scholar] [CrossRef]
- Sakran, R.; El Shamey, E.; Anis, G. Diallel analysis of different rice genotypes under water deficiency conditions and assessing genetic diversity using ssr markers. J. Plant Prod. Sci. 2020, 11, 1319–1332. [Google Scholar] [CrossRef]
- Gramaje, L.V.; Caguiat, J.D.; Enriquez, J.O.S.; dela Cruz, Q.D.; Millas, R.A.; Carampatana, J.E.; Tabanao, D.A.A. Heterosis and combining ability analysis in CMS hybrid rice. Euphytica 2020, 216, 14. [Google Scholar] [CrossRef]
- Abaza, G.M.S.M.; Awaad, H.A.; Attia, Z.M.; Abdel-lateif, K.S.; Gomaa, M.A.; Abaza, S.M.S.M.; Mansour, E. Inducing potential mutants in bread wheat using different doses of certain physical and chemical mutagens. Plant Breed. Biotechnol. 2020, 8, 252–264. [Google Scholar] [CrossRef]
- Gharib, M.; Qabil, N.; Salem, A.; Ali, M.; Awaad, H.; Mansour, E. Characterization of wheat landraces and commercial cultivars based on morpho-phenological and agronomic traits. Cereal Res. Commun. 2021, 49, 149–159. [Google Scholar] [CrossRef]
- Palanga, K.K.; Traore, K.; Bimpong, K.; Jamshed, M.; Mkulama, M.A. Genetic diversity studies on selected rice varieties grown in Africa based on aroma, cooking and eating quality. Afr. J. Biotechnol. 2016, 15, 1136–1146. [Google Scholar]
- Xiao, J.; Li, J.; Yuan, L.; McCouch, S.; Tanksley, S. Genetic diversity and its relationship to hybrid performance and heterosis in rice as revealed by PCR-based markers. Theor. Appl. Genet. 1996, 92, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.J.; Septiningsih, E.M.; Suwardjo, F.; Santoso, T.J.; Silitonga, T.S.; McCouch, S.R. Genetic diversity analysis of traditional and improved Indonesian rice (Oryza sativa L.) germplasm using microsatellite markers. Theor. Appl. Genet. 2007, 114, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Sengar, R. DNA fingerprinting based decoding of indica rice (Oryza sativa L.) via molecular marker (SSR, ISSR, & RAPD) in aerobic condition. Adv. Crop Sci. Technol. 2015, 3, 167. [Google Scholar]
- Igartua, E.; Mansour, E.; Cantalapiedra, C.P.; Contreras-Moreira, B.; Gracia, M.P.; Fuster, P.; Escribano, J.; Molina-Cano, J.L.; Moralejo, M.; Thomas, W.T. Selection footprints in barley breeding lines detected by combining genotyping-by-sequencing with reference genome information. Mol. Breed. 2015, 35, 11. [Google Scholar] [CrossRef] [Green Version]
- Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogel, J.; Tingey, S.; Rafalski, A. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- Xangsayasane, P.; Xie, F.; Hernandez, J.E.; Boirromeo, T.H. Hybrid rice heterosis and genetic diversity of IRRI and Lao rice. Field Crops Res. 2010, 117, 18–23. [Google Scholar] [CrossRef]
- Jaikishan, I.; Rajendrakumar, P.; Ramesha, M.; Viraktamath, B.; Balachandran, S.; Neeraja, C.; Sujatha, K.; Srinivasa Rao, K.; Natarajkumar, P.; Hari, Y. Prediction of heterosis for grain yield in rice using ‘key’informative EST-SSR markers. Plant Breed. 2010, 129, 108–111. [Google Scholar] [CrossRef]
- Singh, V.K.; Upadhyay, P.; Sinha, P.; Mall, A.K.; Ellur, R.K.; Singh, A.; Jaiswal, S.K.; Biradar, S.; Ramakrishna, S.; Sundaram, R. Prediction of hybrid performance based on the genetic distance of parental lines in two-line rice (Oryza sativa L.) hybrids. J. Crop Sci. Biotechnol. 2011, 14, 1. [Google Scholar] [CrossRef]
- Yingheng, W.; Qiuhua, C.A.I.; Hongguang, X.; Fangxi, W.; Ling, L.; Wei, H.; Jianfu, Z. Determination of heterotic groups and heterosis analysis of yield performance in indica rice. Rice Sci. 2018, 25, 261–269. [Google Scholar] [CrossRef]
- Hua, J.; Xing, Y.; Xu, C.; Sun, X.; Yu, S.; Zhang, Q. Genetic dissection of an elite rice hybrid revealed that heterozygotes are not always advantageous for performance. Genetics 2002, 162, 1885–1895. [Google Scholar] [CrossRef] [PubMed]
- Jodon, N. Experiments on artificial hybridization of rice. Agron. J. 1938, 30, 294–305. [Google Scholar] [CrossRef]
- Butany, W. Mass emasculation in rice. Intern. Rice Com. Newsl. 1961, 9, 9–13. [Google Scholar]
- De Datta, S.; Malabuyoc, J.; Aragon, E. A field screening technique for evaluating rice germplasm for drought tolerance during the vegetative stage. Field Crops Res. 1988, 19, 123–134. [Google Scholar] [CrossRef]
- Barrs, H.; Weatherley, P. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J. DNA protocols for plants Molecular techniques in taxonomy. NATO ASI Ser. 1991, 57, 283–293. [Google Scholar]
- Vikram, P.; Swamy, B.; Dixit, S.; Ahmed, H.U.; Teresa Sta Cruz, M.; Singh, A.K.; Kumar, A. qDTY 1.1, a major QTL for rice grain yield under reproductive-stage drought stress with a consistent effect in multiple elite genetic backgrounds. BMC Genet. 2011, 12, 89. [Google Scholar] [CrossRef] [Green Version]
- Qu, Y.; Mu, P.; Zhang, H.; Chen, C.Y.; Gao, Y.; Tian, Y.; Wen, F.; Li, Z. Mapping QTLs of root morphological traits at different growth stages in rice. Genetica 2008, 133, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Venuprasad, R.; Bool, M.; Quiatchon, L.; Sta Cruz, M.; Amante, M.; Atlin, G. A large-effect QTL for rice grain yield under upland drought stress on chromosome. Mol. Breed. 2012, 30, 535–547. [Google Scholar] [CrossRef]
- Ramadan, E.; Elmoghazy, A.; El-Mowafi, H. Molecular markers based genetic diversity analysis for drought tolerance in rice (Oryza Sativa, L.) Using SSR Markers. Int. J. Sci. Res. Agric. Sci. 2015, 2, 1. [Google Scholar]
- Rosielle, A.; Hamblin, J. Theoretical aspects of selection for yield in stress and non-stress environment. Crop Sci. 1981, 21, 943–946. [Google Scholar] [CrossRef]
- Fernandez, G.C. Effective selection criteria for assessing plant stress tolerance. In Proceedings of the International Symposium on Adaptation of Vegetables and Other Food Crops in Temperature and Water Stress, Shanhua, Taiwan, 13–16 August 1992; pp. 257–270. [Google Scholar]
- Gavuzzi, P.; Rizza, F.; Palumbo, M.; Campanile, R.; Ricciardi, G.; Borghi, B. Evaluation of field and laboratory predictors of drought and heat tolerance in winter cereals. Can. J. Plant Sci. 1997, 77, 523–531. [Google Scholar] [CrossRef]
- Ward, J.H., Jr. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Venuprasad, R.; Lafitte, H.R.; Atlin, G.N. Response to direct selection for grain yield under drought stress in rice. Crop Sci. 2007, 47, 285–293. [Google Scholar] [CrossRef]
- Monkham, T.; Jongdee, B.; Pantuwan, G.; Sanitchon, J.; Mitchell, J.; Fukai, S. Genotypic variation in grain yield and flowering pattern in terminal and intermittent drought screening methods in rainfed lowland rice. Field Crops Res. 2015, 175, 26–36. [Google Scholar] [CrossRef]
- Mansour, E.; Mahgoub, H.A.; Mahgoub, S.A.; El-Sobky, E.-S.E.; Abdul-Hamid, M.I.; Kamara, M.M.; AbuQamar, S.F.; El-Tarabily, K.A.; Desoky, E.-S.M. Enhancement of drought tolerance in diverse Vicia faba cultivars by inoculation with plant growth-promoting rhizobacteria under newly reclaimed soil conditions. Sci. Rep. 2021, 11, 24142. [Google Scholar] [CrossRef]
- El-Sanatawy, A.M.; El-Kholy, A.S.; Ali, M.; Awad, M.F.; Mansour, E. Maize seedling establishment, grain yield and crop water productivity response to seed priming and irrigation management in a mediterranean arid environment. Agronomy 2021, 11, 756. [Google Scholar] [CrossRef]
- Desoky, E.-S.M.; Mansour, E.; El-Sobky, E.-S.E.; Abdul-Hamid, M.I.; Taha, T.F.; Elakkad, H.A.; Arnaout, S.M.; Eid, R.S.; El-Tarabily, K.A.; Yasin, M.A. Physio-biochemical and agronomic responses of faba beans to exogenously applied nano-silicon under drought stress conditions. Front. Plant Sci. 2021, 12, 637783. [Google Scholar] [CrossRef] [PubMed]
- Gaballah, M.M.; Metwally, A.M.; Skalicky, M.; Hassan, M.M.; Brestic, M.; El Sabagh, A.; Fayed, A.M. Genetic diversity of selected rice genotypes under water stress conditions. Plants 2021, 10, 27. [Google Scholar] [CrossRef]
- Sellammal, R.; Robin, S.; Raveendran, M. Association and heritability studies for drought resistance under varied moisture stress regimes in backcross inbred population of rice. Rice Sci. 2014, 21, 150–161. [Google Scholar] [CrossRef]
- Mansour, E.; Desoky, E.-S.M.; Ali, M.M.; Abdul-Hamid, M.I.; Ullah, H.; Attia, A.; Datta, A. Identifying drought-tolerant genotypes of faba bean and their agro-physiological responses to different water regimes in an arid Mediterranean environment. Agric. Water Manag. 2021, 247, 106754. [Google Scholar] [CrossRef]
- Ali, M.M.; Mansour, E.; Awaad, H.A. Drought tolerance in some field crops: State of the art review. In Mitigating Environmental Stresses for Agricultural Sustainability in Egypt; Springer: Berlin/Heidelberg, Germany, 2021; pp. 17–62. [Google Scholar]
- Kamarudin, Z.S.; Yusop, M.R.; Tengku Muda Mohamed, M.; Ismail, M.R.; Harun, A.R. Growth performance and antioxidant enzyme activities of advanced mutant rice genotypes under drought stress condition. Agronomy 2018, 8, 279. [Google Scholar] [CrossRef] [Green Version]
- Desoky, E.-S.M.; Mansour, E.; Ali, M.M.A.; Yasin, M.A.T.; Abdul-Hamid, M.I.E.; Rady, M.M.; Ali, E.F. Exogenously used 24-epibrassinolide promotes drought tolerance in maize hybrids by improving plant and water productivity in an arid environment. Plants 2021, 10, 354. [Google Scholar] [CrossRef]
- Shivakrishna, P.; Reddy, K.A.; Rao, D.M. Effect of PEG-6000 imposed drought stress on RNA content, relative water content (RWC), and chlorophyll content in peanut leaves and roots. Saudi J. Biol. Sci. 2018, 25, 285–289. [Google Scholar]
- Nahar, S.; Sahoo, L.; Tanti, B. Screening of drought tolerant rice through morpho-physiological and biochemical approaches. Biocatal. Agric. Biotechnol. 2018, 15, 150–159. [Google Scholar] [CrossRef]
- Desoky, E.-S.M.; Elrys, A.S.; Mansour, E.; Eid, R.S.; Selem, E.; Rady, M.M.; Ali, E.F.; Mersal, G.A.; Semida, W.M. Application of biostimulants promotes growth and productivity by fortifying the antioxidant machinery and suppressing oxidative stress in faba bean under various abiotic stresses. Sci. Hortic. 2021, 288, 110340. [Google Scholar] [CrossRef]
- Naroui Rad, M.R.; Kadir, M.A.; Yusop, M.R. Genetic behaviour for plant capacity to produce chlorophyll in wheat (Triticum aestivum) under drought stress. Aust. J. Crop Sci. 2012, 6, 415–420. [Google Scholar]
- Desoky, E.-S.M.; Merwad, A.-R.; Abo El-Maati, M.F.; Mansour, E.; Arnaout, S.M.; Awad, M.F.; Ramadan, M.F.; Ibrahim, S.A. Physiological and biochemical mechanisms of exogenously applied selenium for alleviating destructive impacts induced by salinity stress in bread wheat. Agronomy 2021, 11, 926. [Google Scholar] [CrossRef]
- Tabkhkar, N.; Rabiei, B.; Lahiji, H.S.; Chaleshtori, M.H. Genetic variation and association analysis of the SSR markers linked to the major drought-yield QTLs of rice. Biochem. Genet. 2018, 56, 356–374. [Google Scholar] [CrossRef]
- Kumar, S.; Dwivedi, S.K.; Basu, S.; Kumar, G.; Mishra, J.; Koley, T.K.; Rao, K.; Choudhary, A.; Mondal, S.; Kumar, S. Anatomical, agro-morphological and physiological changes in rice under cumulative and stage specific drought conditions prevailed in eastern region of India. Field Crops Res. 2020, 245, 107658. [Google Scholar] [CrossRef]
- Desoky, E.-S.M.; Mansour, E.; Yasin, M.A.; El Sobky, E.-S.E.; Rady, M.M. Improvement of drought tolerance in five different cultivars of Vicia faba with foliar application of ascorbic acid or silicon. Span. J. Agric. Res. 2020, 18, 16. [Google Scholar] [CrossRef]
- Elshayb, O.M.; Nada, A.M.; Ibrahim, H.M.; Amin, H.E.; Atta, A.M. Application of silica nanoparticles for improving growth, yield, and enzymatic antioxidant for the hybrid rice EHR1 growing under water regime Conditions. Materials 2021, 14, 1150. [Google Scholar] [CrossRef]
- Liu, J.; Liao, D.; Oane, R.; Estenor, L.; Yang, X.; Li, Z.; Bennett, J. Genetic variation in the sensitivity of anther dehiscence to drought stress in rice. Field Crops Res. 2006, 97, 87–100. [Google Scholar] [CrossRef]
- Mukamuhirwa, A.; Persson Hovmalm, H.; Bolinsson, H.; Ortiz, R.; Nyamangyoku, O.; Johansson, E. Concurrent drought and temperature stress in rice a possible result of the predicted climate change: Effects on yield attributes, eating characteristics, and health promoting compounds. Int. J. Environ. Res. 2019, 16, 1043. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, J.; Moya, T. Water deficits and yield in upland rice. Field Crops Res. 1981, 4, 247–259. [Google Scholar] [CrossRef]
- Muthuramu, S.; Jebaraj, S.; Ushakumari, R.; Gnanasekaran, M. Estimation of combining ability and heterosis for drought tolerance in different locations in rice (Oryza sativa L.). Electron. J. Plant Breed. 2010, 1, 1279–1285. [Google Scholar]
- Fasahat, P.; Rajabi, A.; Rad, J.M.; Derera, J. Principles and utilization of combining ability in plant breeding. Biom. Biostat. Int. J. 2016, 4, 1–24. [Google Scholar] [CrossRef]
- Zewdu, Z. Combining ability analysis of yield and yield components in selected rice (Oryza sativa L.) genotypes. Cogent Food agric. 2020, 6, 1811594. [Google Scholar] [CrossRef]
- Bassuony, N.N.; Zsembeli, J. Inheritance of some flag leaf and yield characteristics by half-diallel analysis in rice crops (Oryza Sativa, L.). Cereal Res. Commun. 2021, 49, 503–510. [Google Scholar] [CrossRef]
- Sharifi, P.; Dehghani, H.; Moumeni, A.; Moghaddam, M. Genetic main effect and genotype× environment interaction for cooking quality traits in a diallel set of Indica rice (Oryza sativa L.) varieties. Crop Pasture Sci. 2010, 61, 475–482. [Google Scholar] [CrossRef]
- Mau, Y.S.; Ndiwa, A.S.; Oematan, S.S.; Markus, J.E. Drought tolerance indices for selection of drought tolerant, high yielding upland rice genotypes. Aust. J. Crop Sci. 2019, 13, 170–178. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Barik, S.R.; Sahoo, A.; Mohapatra, S.; Nayak, D.K.; Mahender, A.; Meher, J.; Anandan, A.; Pandit, E. Population structure, genetic diversity and molecular marker-trait association analysis for high temperature stress tolerance in rice. PLoS ONE 2016, 11, e0160027. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Choudhury, D.R.; Tiwari, G.; Singh, A.K.; Kumar, S.; Srinivasan, K.; Tyagi, R.; Sharma, A.; Singh, N.; Singh, R. Genetic diversity trend in Indian rice varieties: An analysis using SSR markers. BMC Genet. 2016, 17, 127. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.M.; Rasaul, M.; Hossain, M.; Iftekharuddaula, K.; Hasegawa, H. Molecular characterization and genetic diversity analysis of rice (Oryza sativa L.) using SSR markers. J. Crop Improv. 2012, 26, 244–257. [Google Scholar] [CrossRef]
- Das, B.; Sengupta, S.; Parida, S.K.; Roy, B.; Ghosh, M.; Prasad, M.; Ghose, T.K. Genetic diversity and population structure of rice landraces from Eastern and North Eastern States of India. BMC Genet. 2013, 14, 71. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Li, J.; Li, X.; Liu, X.; Zhao, X.; Lu, Y. Population structure and genetic diversity in a rice core collection (Oryza sativa L.) investigated with SSR markers. PLoS ONE 2011, 6, e27565. [Google Scholar] [CrossRef] [Green Version]
- Salem, K.F.; Sallam, A. Analysis of population structure and genetic diversity of Egyptian and exotic rice (Oryza sativa L.) genotypes. Comptes Rendus Biol. 2016, 339, 1–9. [Google Scholar] [CrossRef]
- Chakravarthi, B.K.; Naravaneni, R. SSR marker based DNA fingerprinting and diversity study in rice (Oryza sativa. L). Afr. J. Biotechnol. 2006, 5, 684–688. [Google Scholar]
- Kanawapee, N.; Sanitchon, J.; Srihaban, P.; Theerakulpisut, P. Genetic diversity analysis of rice cultivars (Oryza sativa L.) differing in salinity tolerance based on RAPD and SSR markers. Electron. J. Biotechnol. 2011, 14, 2. [Google Scholar]
- Mukherjee, M.; Padhy, B.; Srinivasan, B.; Mahadani, P.; Baksh, S.Y.; Donde, R.; Singh, O.N.; Behera, L.; Swain, P.; Dash, S.K. Revealing genetic relationship and prospecting of novel donors among upland rice genotypes using qDTY-linked SSR markers. Rice Sci. 2018, 25, 308–319. [Google Scholar] [CrossRef]
- Sreewongchai, T.; Sripichitt, P.; Matthayatthaworn, W. Parental genetic distance and combining ability analyses in relation to heterosis in various rice origins. J. Crop Sci. Biotechnol. 2021, 24, 327–336. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhou, Z.; Yang, G.; Xu, C.; Liu, K.; Maroof, M.S. Molecular marker heterozygosity and hybrid performance in indica and japonica rice. Theor. Appl. Genet. 1996, 93, 1218–1224. [Google Scholar] [CrossRef]
- Habibullah, M.; Sarkar, S.; Islam, M.M.; Ahmed, K.U.; Rahman, M.; Awad, M.F.; ElSayed, A.I.; Mansour, E.; Hossain, M. Assessing the response of diverse sesame genotypes to waterlogging durations at different plant growth stages. Plants 2021, 10, 2294. [Google Scholar] [CrossRef]
- Mansour, E.; Moustafa, E.S.; Abdul-Hamid, M.I.; Ash-shormillesy, S.M.; Merwad, A.-R.M.; Wafa, H.A.; Igartua, E. Field responses of barley genotypes across a salinity gradient in an arid Mediterranean environment. Agric. Water Manag. 2021, 258, 107206. [Google Scholar] [CrossRef]
- Swailam, M.; Mowafy, S.; El-Naggar, N.; Mansour, E. Agronomic responses of diverse bread wheat genotypes to phosphorus levels and nitrogen forms in a semiarid environment. SABRAO J. Breed. Genet. 2021, 53, 592–608. [Google Scholar] [CrossRef]
- El-Sanatawy, A.M.; Ash-Shormillesy, S.M.; Qabil, N.; Awad, M.F.; Mansour, E. Seed halo-priming improves seedling vigor, grain yield, and water use efficiency of maize under varying irrigation regimes. Water 2021, 13, 2115. [Google Scholar] [CrossRef]
- Moustafa, E.S.; El-Sobky, E.-S.E.; Farag, H.I.; Yasin, M.A.; Attia, A.; Rady, M.O.; Awad, M.F.; Mansour, E. Sowing date and genotype influence on yield and quality of dual-purpose barley in a salt-affected arid region. Agronomy 2021, 11, 717. [Google Scholar] [CrossRef]
- Abd Allah, A.; Ammar, M.; Badawi, A. Screening rice genotypes for drought resistance in Egypt. J. Plant Breed. Crop Sci. 2010, 2, 205–215. [Google Scholar]
- Kumar, S.; Dwivedi, S.; Singh, S.; Jha, S.; Lekshmy, S.; Elanchezhian, R.; Singh, O.; Bhatt, B. Identification of drought tolerant rice genotypes by analysing drought tolerance indices and morpho-physiological traits. ABRAO J. Breed. Genet. 2014, 46, 217–230. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Name | Parentage | Origin | Drought Tolerance |
|---|---|---|---|---|
| P1 | Sakha-102 | GZ4096/Giza177 | Egypt | Sensitive |
| P2 | Sakha-105 | GZ5581-46-3/GZ 4316-7-1-1 | Egypt | Sensitive |
| P3 | Giza-179 | GZ6296/GZ1368 | Egypt | Tolerant |
| P4 | Dullar | Unknown | India | Tolerant |
| P5 | IRAT-112 | (IRAT 13/dourado precoce) | Ivory Coast | Tolerant |
| P6 | Moroberekan | IR 8-24-6-(M307 H5) | Guinea (West Africa) | Tolerant |
| Source of Variance | DF | DTH | PH | LR | RWC | CHLC | NP | NFG | SP | TGW | GYPP |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Years (Y) | 1 | 59.21 * | 670.29 * | 56.02 ** | 938.71 ** | 545.18 ** | 67.51 ** | 482.91 * | 55.81 ** | 15.37 * | 76.42 ** |
| Replication/Y | 4 | 6.23 | 60.96 | 0.4 | 4.06 | 9.5 | 1.12 | 41.18 | 0.89 | 1.22 | 2.97 |
| Irrigation (I) | 1 | 898.47 ** | 15047.07 ** | 231.23 ** | 12377.84 ** | 1250.43 ** | 1340.22 ** | 46572.20 ** | 5346.77 ** | 753.61 ** | 7674.90 ** |
| Y × I | 1 | 145.51 ** | 857.33 ** | 41.31 ** | 89.51 ** | 43.29* | 7.99 * | 681.84 * | 11.60 * | 6.45 * | 31.21 ** |
| Error a | 4 | 6.18 | 29.51 | 0.33 | 3.67 | 5.32 | 0.92 | 35.17 | 0.73 | 0.84 | 2.36 |
| Genotype (G) | 20 | 527.42 ** | 1693.12 ** | 4.81 ** | 194.00 ** | 74.93 ** | 77.73 ** | 1801.61 ** | 101.16 ** | 14.24 ** | 190.07 ** |
| G × Y | 20 | 6.29 ** | 15.49 ** | 1.88 ** | 10.26 ** | 3.52 ** | 8.12 ** | 169.64 ** | 5.77 ** | 2.67 ** | 25.64 ** |
| G × I | 20 | 3.89 ** | 66.62 ** | 2.93 ** | 102.63 ** | 8.47 ** | 24.06 ** | 112.19 ** | 28.74 ** | 5.87 ** | 82.93 ** |
| G × Y × I | 20 | 6.00 ** | 26.80 ** | 1.09 ** | 8.12 ** | 5.32 ** | 3.57 ** | 77.00 ** | 4.08 ** | 1.26 * | 19.53 ** |
| Pooled Error | 160 | 1.01 | 4.94 | 0.18 | 1.29 | 1.05 | 0.87 | 29.48 | 0.62 | 0.63 | 2.18 |
| GCA | 5 | 892.34 ** | 4309.89 ** | 6.97 ** | 284.41 ** | 57.28 ** | 87.79 ** | 1695.02 ** | 77.75 ** | 5.66 ** | 267.52 ** |
| SCA | 15 | 405.78 ** | 820.86 ** | 4.09 ** | 163.86 ** | 80.81 ** | 74.38 ** | 1837.15 ** | 108.96 ** | 17.10 ** | 164.25 ** |
| GCA × Y | 5 | 6.57 ** | 10.67 | 2.48 ** | 20.84 ** | 3.54 ** | 8.08 ** | 149.50 ** | 7.17 ** | 2.49 ** | 13.68 ** |
| SCA × Y | 15 | 6.20 ** | 17.09 ** | 1.68 ** | 6.73 ** | 3.51 ** | 8.13 ** | 176.35 ** | 5.30 ** | 2.72 ** | 29.63 ** |
| GCA × I | 5 | 4.44 ** | 20.53 ** | 5.94 ** | 128.00 ** | 4.75 ** | 17.43 ** | 120.99 ** | 53.53 ** | 3.67 ** | 188.29 ** |
| SCA × I | 15 | 3.71 ** | 81.99 ** | 1.93 ** | 94.17 ** | 9.71 ** | 26.27 ** | 109.26 ** | 20.48 ** | 6.60 ** | 47.81 ** |
| GCA × Y × I | 5 | 4.23 ** | 24.65 ** | 1.60 ** | 3.45 * | 3.59 ** | 1.15 | 10.52 | 3.61 ** | 0.84 | 13.17 ** |
| SCA × Y × I | 15 | 6.60 ** | 27.51 ** | 0.92 ** | 9.68 ** | 5.89 ** | 4.37 ** | 99.16 ** | 4.24 ** | 1.40 ** | 21.65 ** |
| GCA/SCA | 2.2 | 5.25 | 1.7 | 1.74 | 0.71 | 1.18 | 0.92 | 0.71 | 0.33 | 1.63 | |
| GCAxY/SCAxY | 1.06 | 0.62 | 1.47 | 3.1 | 1.01 | 0.99 | 0.85 | 1.35 | 0.92 | 0.46 | |
| GCAxI/SCAxI | 1.19 | 0.25 | 3.07 | 1.36 | 0.49 | 0.66 | 1.11 | 2.61 | 0.56 | 3.94 |
| Parent | Days to Heading | Plant Height | Leaf Rolling | Relative Water Content | Chlorophyll Content | |||||
| Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | |
| P1 | −3.90 ** | −4.36 ** | −4.73 ** | −5.66 ** | 0.11 * | 0.58 ** | −0.44 ** | −2.04 ** | 0.58 ** | 0.69 ** |
| P2 | −2.37 ** | −1.73 ** | −5.90 ** | −6.28 ** | −0.14 ** | 0.56 ** | 0.10 | −3.23 ** | 0.72 ** | 0.31 * |
| P3 | −1.58 ** | −1.42 ** | −5.33 ** | −4.34 ** | −0.12 * | −0.45 ** | −0.28 | 2.79 ** | −0.84 ** | −0.27 * |
| P4 | 2.34 ** | 1.84 ** | 8.71 ** | 8.96 ** | 0.16 ** | −0.45 ** | 1.31 ** | 3.17 ** | 0.11 | 0.11 |
| P5 | 1.59 ** | 1.57 ** | 0.13 | −0.90 ** | 0.16 ** | 0.15 ** | −1.59 ** | −1.93 ** | −0.98 ** | −1.60 ** |
| P6 | 3.92 ** | 4.09 ** | 7.13 ** | 8.22 ** | −0.16 ** | −0.39 ** | 0.90 ** | 1.24 ** | 0.41 ** | 0.75 ** |
| LSD (gi) 0.05 | 0.28 | 0.25 | 0.59 | 0.58 | 0.11 | 0.12 | 0.29 | 0.31 | 0.28 | 0.26 |
| LSD (gi) 0.01 | 0.37 | 0.33 | 0.78 | 0.76 | 0.14 | 0.15 | 0.38 | 0.41 | 0.37 | 0.35 |
| Parent | Number of Panicles/Plant | Number of Filled Grains/Panicle | Sterility Percentage | 1000-Grain Weight | Grain Yield/Plant | |||||
| Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | |
| P1 | −0.37 ** | −0.13 | −1.45 * | −3.91 ** | −0.82 ** | 0.95 ** | 0.69 ** | 0.09 | 1.44 ** | −1.54 ** |
| P2 | −0.16 | −0.26 * | −0.31 | −3.25 ** | −0.44 ** | 0.70 ** | −0.01 | −0.26 * | 1.12 ** | −2.76 ** |
| P3 | 1.53 ** | 0.71 ** | 6.90 ** | 7.44 ** | −1.14 ** | −1.13 ** | 0.05 | −0.01 | 2.22 ** | 2.38 ** |
| P4 | 1.53 ** | 0.43 ** | 2.09 ** | 4.07 ** | 0.14 | −1.47 ** | −0.40 ** | 0.06 | 0.53 ** | 2.49 ** |
| P5 | −2.06 ** | −0.92 ** | −2.73 ** | −2.33 ** | 1.20 ** | 1.82 ** | −0.34 ** | −0.18 | −3.50 ** | −1.08 ** |
| P6 | −0.47 ** | 0.17 | −4.50 ** | −2.03 ** | 1.05 ** | −0.88 ** | 0.01 | 0.31 ** | −1.82 ** | 0.52 ** |
| LSD (gi) 0.05 | 0.26 | 0.23 | 1.43 | 1.41 | 0.17 | 0.24 | 0.20 | 0.22 | 0.40 | 0.37 |
| LSD (gi) 0.01 | 0.35 | 0.30 | 1.90 | 1.88 | 0.22 | 0.32 | 0.27 | 0.29 | 0.53 | 0.50 |
| Cross | Days to Heading | Plant Height | Leaf Rolling | Relative Water Content | Chlorophyll Content | |||||
| Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | |
| P1 × P2 | −0.83 * | −0.66 | 7.70 ** | 13.07 ** | −0.15 | −0.21 | 3.95 ** | 2.62 ** | 1.24 ** | 0.72 * |
| P1 × P3 | −0.26 | 0.27 | 2.53 ** | 1.56 | −0.23 | −0.55 ** | 3.40 ** | 1.79 ** | 0.66 | 0.05 |
| P1 × P4 | 5.82 ** | 3.19 ** | 0.11 | −1.93 * | 0.01 | −0.73 ** | −0.82 * | 4.11 ** | 1.23 ** | 1.99 ** |
| P1 × P5 | −9.77 ** | −9.69 ** | −13.31 ** | −14.40 ** | 1.21 ** | 1.03 ** | −4.00 ** | 3.05 ** | −3.42 ** | −1.47 ** |
| P1 × P6 | 6.54 ** | 6.28 ** | −0.30 | 0.32 | −0.15 | −0.47 ** | −3.50 ** | −5.52 ** | 1.69 ** | 0.31 |
| P2 × P3 | 3.87 ** | 4.92 ** | −2.35 ** | −0.84 | 0.41 ** | 1.64 ** | −4.16 ** | −3.88 ** | −2.71 ** | −4.91 ** |
| P2 × P4 | 5.93 ** | 7.08 ** | 0.93 | −3.64 ** | 0.38* | −0.18 | −1.99 ** | −1.27 ** | −3.72 ** | −3.80 ** |
| P2 × P5 | 7.03 ** | 6.18 ** | 14.86 ** | −1.07 | −0.16 | −0.33 * | 1.92 ** | 7.50 ** | 1.44 ** | 2.74 ** |
| P2 × P6 | −6.65 ** | −8.17 ** | −6.14 ** | −3.73 ** | 0.01 | −0.89 ** | −1.23 ** | 6.34 ** | 2.03 ** | 0.71 |
| P3 × P4 | 4.15 ** | 4.03 ** | 13.86 ** | 14.05 ** | −0.09 | −0.58 ** | 2.92 ** | 2.49 ** | 4.72 ** | 4.79 ** |
| P3 × P5 | 5.58 ** | 5.49 ** | 10.70 ** | 13.94 ** | −0.09 | −0.78 ** | 1.81 ** | 3.19 ** | 1.43 ** | −0.50 |
| P3 × P6 | 1.91 ** | 0.57 | −5.53 ** | −8.65 ** | 0.48 ** | 0.26 | −4.96 ** | −4.34 ** | −0.06 | 1.15 ** |
| P4 × P5 | −4.36 ** | −4.18 ** | −11.75 ** | −7.52 ** | 0.04 | 1.22 ** | −0.47 | −10.56 ** | 2.04 ** | −1.89 ** |
| P4 × P6 | −5.37 ** | −5.47 ** | 1.45 | 2.12 ** | 0.05 | −0.35 * | 1.70 ** | 4.03 ** | −0.18 | 2.10 ** |
| P5 × P6 | 1.42 ** | 2.45 ** | −0.17 | 0.72 | −0.05 | 1.42 ** | 1.35 ** | −9.63 ** | −4.26 ** | −4.52 ** |
| LSD Sij 0.05 | 0.76 | 0.68 | 1.62 | 1.58 | 0.29 | 0.32 | 0.80 | 0.84 | 0.76 | 0.72 |
| LSD Sij 0.01 | 1.01 | 0.91 | 2.15 | 2.10 | 0.38 | 0.42 | 1.06 | 1.12 | 1.00 | 0.95 |
| Cross | Number of Panicles/Plant | Number of Filled Grains/Panicle | Sterility Percentage | 1000-Grain Weight | Grain Yield/Plant | |||||
| Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | Well- Watered | Water- Deficit | |
| P1 × P2 | 3.83 ** | 1.50 ** | 3.97 * | 3.98 * | −1.68 ** | −3.42 ** | 1.52 ** | 0.37 | 3.13 ** | 1.59 ** |
| P1 × P3 | 4.80 ** | 1.68 ** | 8.60 ** | 7.18 ** | 0.33 | 2.94 ** | 0.81 ** | 1.05 ** | −0.21 | −1.56 ** |
| P1 × P4 | −0.61 | 1.82 ** | −2.51 | 3.20 | −0.10 | 0.32 | −1.52 ** | −1.34 ** | 3.09 ** | 4.14 ** |
| P1 × P5 | 1.40 ** | 2.78 ** | −15.77 ** | −12.22 ** | 4.45 ** | 2.30 ** | −2.82 ** | 0.15 | −5.36 ** | 3.90 ** |
| P1 × P6 | −4.47 ** | −3.01 ** | 0.30 | −3.19 | 2.60 ** | 3.17 ** | 1.94 ** | 0.15 | 0.29 | −2.19 ** |
| P2 × P3 | −4.40 ** | −1.38 ** | −23.55 ** | −19.96 ** | 6.11 ** | 2.51 ** | −1.90 ** | −0.90 ** | −10.52 ** | −6.84 ** |
| P2 × P4 | −3.77 ** | −1.80 ** | −19.40 ** | −19.76 ** | 3.83 ** | 6.01 ** | 0.29 | −0.17 | −2.47 ** | −3.44 ** |
| P2 × P5 | 1.09 ** | 2.81 ** | 18.08 ** | 19.70 ** | −0.79 ** | 0.36 | 0.92 ** | 0.92 ** | 7.79 ** | 5.96 ** |
| P2 × P6 | 4.99 ** | −0.90 ** | 13.35 ** | 12.26 ** | 0.47 * | −2.74 ** | −0.34 | 1.12 ** | −0.88 | 4.59 ** |
| P3 × P4 | 2.43 ** | 1.96 ** | 11.31 ** | 6.52 ** | −0.94 ** | −1.07 ** | 2.48 ** | 1.19 ** | 5.71 ** | 3.30 ** |
| P3 × P5 | −5.13 ** | −2.66 ** | 7.88 ** | −3.57 | −1.23 ** | −1.25 ** | 2.34 ** | −0.68* | 2.90 ** | −0.60 |
| P3 × P6 | −0.20 | 1.59 ** | −6.35 ** | −9.38 ** | 1.66 ** | 0.26 | −0.60* | 0.83 ** | −2.32 ** | 2.23 ** |
| P4 × P5 | 3.40 ** | −2.89 ** | 8.69 ** | 11.29 ** | −0.15 | 1.75 ** | 0.79 ** | 0.69 * | −0.82 | −1.87 ** |
| P4 × P6 | 1.80 ** | 2.03 ** | 8.51 ** | 12.52 ** | −0.62 ** | −3.48 ** | 0.22 | 0.65 * | 0.28 | 2.36 ** |
| P5 × P6 | 0.09 | 0.96 ** | −9.89 ** | −19.79 ** | 1.48 ** | 6.54 ** | −1.53 ** | −2.70 ** | −0.12 | −5.74 ** |
| LSD Sij 0.05 | 0.72 | 0.63 | 3.94 | 3.88 | 0.46 | 0.66 | 0.55 | 0.59 | 1.10 | 1.03 |
| LSD Sij 0.01 | 0.95 | 0.83 | 5.22 | 5.15 | 0.60 | 0.87 | 0.73 | 0.79 | 1.46 | 1.36 |
| Cross | Days to Heading | Plant Height | Leaf Rolling | Relative Water Content (%) | Chlorophyll Content | |||||||||||||||
| M.P | B.P | M.P | B.P | M.P | B.P | M.P | B.P | M.P | B.P | |||||||||||
| W | S | W | S | W | S | W | S | W | S | W | S | W | S | W | S | W | S | W | S | |
| P1 × P2 | 1.99 ** | 1.67 ** | 2.47 ** | 1.83 ** | 10.14 ** | 15.12 ** | 16.51 ** | 17.61 ** | 9.47 | −8.53 | 25.44 | −3.62 | 3.93 ** | 11.47 ** | 3.12 ** | 7.05 ** | 2.55 * | −0.04 | 0.52 | −2.76 * |
| P1 × P3 | 4.21 ** | 4.39 ** | 5.47 ** | 5.60 ** | 6.23 ** | 7.07 ** | 12.95 ** | 12.20 ** | 3.66 | −18.76 ** | 16.99 | 16.78 | 3.46 ** | 4.47 ** | 3.26 ** | −4.47 ** | 4.77 ** | 1.50 | −0.14 | −0.30 |
| P1 × P4 | 7.77 ** | 4.39 ** | 13.55 ** | 10.01 ** | 0.36 | −1.42 | 10.78 ** | 13.00 ** | 15.34 | −26.02 ** | 23.75 | −0.26 | −0.86 | 7.58 ** | −2.21 ** | −2.20 * | 6.04 ** | 8.17 ** | 3.30 * | 5.52 ** |
| P1 × P5 | −9.39 ** | −10.12 ** | −3.74 ** | −4.46 ** | −12.26 ** | −17.03 ** | −9.12 ** | −11.19 ** | 94.74 ** | 31.61 ** | 96.46 ** | 98.44 ** | −4.96 ** | 4.43 ** | −6.71 ** | −0.42 | −8.61 ** | −6.28 ** | −9.69 ** | −7.41 ** |
| P1 × P6 | 6.17 ** | 5.03 ** | 15.98 ** | 15.88 ** | −3.05 ** | −2.15 * | 8.97 ** | 14.63 ** | 6.62 | −16.82 ** | 20.58 | 16.09 | −6.24 ** | −8.95 ** | −9.13 ** | −17.26 ** | 4.11 ** | 1.58 | 3.25* | 0.29 |
| P2 × P3 | 10.77 ** | 12.31 ** | 11.60 ** | 13.79 ** | 6.28 ** | 5.93 ** | 6.79 ** | 8.61 ** | 47.97 ** | 42.59 ** | 50.00 ** | 92.01 ** | −5.64 ** | −1.84 * | −6.20 ** | −13.47 ** | −4.93 ** | −14.62 ** | −11.10 ** | −18.38 ** |
| P2 × P4 | 9.89 ** | 11.03 ** | 16.35 ** | 16.81 ** | 5.01 ** | −1.84 | 23.24 ** | 15.25 ** | 36.35 ** | −8.19 | 69.08 ** | 16.23 * | −2.37 ** | 1.86 * | −2.94 ** | −10.70 ** | −7.08 ** | −10.30 ** | −11.24 ** | −14.83 ** |
| P2 × P5 | 9.34 ** | 8.87 ** | 16.72 ** | 15.54 ** | 17.18 ** | −2.28 * | 28.64 ** | 7.02 ** | 13.63 | 5.33 | 28.93 | 48.92 ** | 2.04 ** | 13.64 ** | −0.61 | 4.28 ** | 0.70 | 0.49 | −2.43 | −3.39 * |
| P2 × P6 | −4.72 ** | −6.89 ** | 4.61 ** | 2.56 ** | −4.27 ** | −4.78 ** | 14.52 ** | 14.31 ** | 16.08 | −22.70 ** | 17.46 | 1.16 | −3.74 ** | 10.16 ** | −5.99 ** | −3.47 ** | 3.07 ** | −1.19 | 1.86 | −2.67 |
| P3 × P4 | 9.65 ** | 9.51 ** | 17.01 ** | 16.81 ** | 17.09 ** | 19.50 ** | 38.19 ** | 44.37 ** | 7.81 | −4.34 | 31.55 | −20.14 * | 3.53 ** | 2.60 ** | 2.33 ** | 1.93 * | 16.46 ** | 14.97 ** | 13.87 ** | 14.15 ** |
| P3 × P5 | 9.43 ** | 9.80 ** | 17.74 ** | 18.15 ** | 14.38 ** | 17.80 ** | 26.23 ** | 32.54 ** | 17.70 | −8.22 | 31.55 | −3.11 | 2.07 ** | 1.90 * | 0.00 | −2.49 ** | 4.20 ** | −4.57 ** | 0.45 | −5.13 ** |
| P3 × P6 | 5.07 ** | 3.32 ** | 16.29 ** | 15.45 ** | −2.89 ** | −5.70 ** | 16.82 ** | 16.55 ** | 50.03 ** | 8.57 | 50.30 ** | 11.11 | −7.91 ** | −8.87 ** | −10.58 ** | −9.51 ** | 1.78 | 3.08 * | −3.76 ** | 0.00 |
| P4 × P5 | −2.70 ** | −2.91 ** | −1.94 ** | −2.11 ** | −8.35 ** | −7.86 ** | −2.53 * | −1.74 | 20.45 | 51.72 ** | 30.45 * | 68.42 ** | 0.02 | −16.98 ** | −3.13 ** | −21.06 ** | 5.57 ** | −6.49 ** | 4.04 ** | −7.69 ** |
| P4 × P6 | −4.03 ** | −5.13 ** | −0.67 | −0.88 | −0.06 | 0.48 | 1.62 | 2.45 * | 13.94 | −3.79 | 39.34 * | −14.32 | 0.42 | 1.87 * | −1.37 | 1.81 * | 1.49 | 7.18 ** | −1.94 | 3.26 * |
| P5 × P6 | 0.78 | 1.35 ** | 3.48 ** | 4.98 ** | −2.15* | −3.20 ** | 5.91 ** | 5.38 ** | 17.67 | 64.50 ** | 31.79 | 77.63 ** | −0.19 | −18.15 ** | −4.99 ** | −22.44 ** | −11.65 ** | −15.00 ** | −13.41 ** | −17.07 ** |
| Cross | Number of Panicles/Plant | Number of Filled Grains/Panicle | Sterility Percentage | 1000-Grain Weight | Grain Yield/Plant | |||||||||||||||
| M.P | B.P | M.P | B.P | M.P | B.P | M.P | B.P | M.P | B.P | |||||||||||
| W | S | W | S | W | S | W | S | W | S | W | S | W | S | W | S | W | S | W | S | |
| P1 × P2 | 30.69 ** | 19.76 ** | 23.77 ** | 11.74 ** | 0.55 | 2.80 | −0.70 | 1.43 | 23.76 ** | −6.82 ** | 27.28 ** | −4.99 | 5.92 ** | 3.51 | 2.76 | 0.89 | 6.34 ** | 16.71 ** | 4.69 * | 15.53 ** |
| P1 × P3 | 26.19 ** | 21.67 ** | 6.81 ** | 8.78 ** | 4.83 * | 1.87 | −0.54 | −10.65 ** | 46.05 ** | 27.40 ** | 54.96 ** | 39.39 ** | 5.88 ** | 6.60 ** | 0.45 | 4.86 * | −2.54 | −3.45 | −7.14 ** | −21.31 ** |
| P1 × P4 | 7.45 ** | 22.91 ** | −4.13 | 11.47 ** | −1.67 | 6.27 * | −2.06 | 1.99 | 19.57 ** | 13.85 ** | 49.62 ** | 27.59 ** | −3.64 * | −4.28 * | −9.35 ** | −5.04 * | 12.33 ** | 26.18 ** | 6.49 ** | 7.83 ** |
| P1 × P5 | 17.48 ** | 32.37 ** | 12.84 ** | 30.91 ** | −11.73 ** | −13.64 ** | −15.00 ** | −15.72 ** | 68.86 ** | 30.20 ** | 125.61 ** | 31.69 ** | −10.67 ** | −0.69 | −13.77 ** | −1.64 | −11.51 ** | 25.27 ** | −24.08 ** | 17.47 ** |
| P1 × P6 | −15.29 ** | −11.65 ** | −18.03 ** | −19.15 ** | 0.34 | −5.30 | −4.14 | −8.48 ** | 58.36 ** | 28.89 ** | 98.53 ** | 39.59 ** | 6.70 ** | 1.11 | 4.33 ** | −0.16 | −0.40 | −1.66 | −6.12 ** | −13.07 ** |
| P2 × P3 | −21.13 ** | −6.57 * | −30.00 ** | −10.77 ** | −18.41 ** | −22.41 ** | −21.66 ** | −31.14 ** | 141.16 ** | 21.17 ** | 148.58 ** | 35.41 ** | −3.85 * | −0.88 | −6.03 ** | −1.82 | −28.36 ** | −26.40 ** | −30.70 ** | −40.49 ** |
| P2 × P4 | −12.43 ** | −9.54 ** | −17.78 ** | −12.26 ** | −14.65 ** | −16.74 ** | −15.37 ** | −19.04 ** | 69.90 ** | 40.46 ** | 119.87 ** | 60.85 ** | 3.81 * | 1.84 | 0.55 | 0.04 | −4.48 * | −7.24 * | −10.78 ** | −21.40 ** |
| P2 × P5 | 10.07 ** | 22.21 ** | 0.35 | 15.21 ** | 14.35 ** | 17.39 ** | 8.80 ** | 16.10 ** | 22.15 ** | 16.94 ** | 69.02 ** | 20.63 ** | 3.65 * | 3.70 * | 3.12 | 0.13 | 22.82 ** | 30.22 ** | 4.01 | 20.95 ** |
| P2 × P6 | 32.33 ** | −4.42 | 29.40 ** | −6.40 | 10.15 ** | 9.21 ** | 3.99 | 6.94 * | 42.60 ** | −5.83 * | 84.90 ** | 4.16 | −1.09 | 6.40 ** | −1.87 | 2.43 | −5.87 ** | 22.07 ** | −12.57 ** | 6.96 * |
| P3 × P4 | 11.38 ** | 15.73 ** | 4.87 * | 13.89 ** | 8.90 ** | 4.39 | 3.72 | −4.99 * | 11.92 ** | 3.94 | 50.30 ** | 6.27 | 15.27 ** | 7.96 ** | 14.23 ** | 7.05 ** | 14.80 ** | 11.10 ** | 3.97 * | 4.91 * |
| P3 × P5 | −27.69 ** | −14.29 ** | −40.78 ** | −22.61 ** | 7.14 ** | −8.22 ** | −1.91 | −17.75 ** | 12.65 ** | 10.94 ** | 62.00 ** | 19.91 ** | 11.84 ** | −3.09 | 9.85 ** | −5.56 ** | 7.74 ** | −3.62 | −11.20 ** | −17.13 ** |
| P3 × P6 | −1.25 | 12.86 ** | −14.05 ** | 10.00 ** | −4.04 | −13.75 ** | −12.81 ** | −22.01 ** | 51.35 ** | 11.88 ** | 103.67 ** | 12.94 ** | 0.43 | 5.26 ** | −2.61 | 2.26 | −10.14 ** | 5.42 * | −19.06 ** | −4.05 |
| P4 × P5 | 23.78 ** | −16.30 ** | 6.61 * | −23.33 ** | 9.88 ** | 13.04 ** | 5.39 * | 11.11 ** | 11.09 ** | 27.95 ** | 17.36 ** | 41.62 ** | 5.02 ** | 2.32 | 2.25 | 0.55 | 5.22 * | −1.23 | −5.35 * | −10.61 ** |
| P4 × P6 | 15.96 ** | 15.75 ** | 6.61 * | 14.62 ** | 9.21 ** | 13.36 ** | 3.93 | 12.55 ** | 11.59 ** | −9.80 ** | 11.77 ** | −6.88 * | 2.72 | 3.95 * | −1.26 | 1.83 | 2.88 | 13.10 ** | 2.25 | 8.76 ** |
| P5 × P6 | 5.04 | 9.58 ** | −2.24 | 1.30 | −5.10 * | −22.08 ** | −5.88 * | −22.86 ** | 32.51 ** | 53.20 ** | 39.75 ** | 63.92 ** | −6.30 ** | −12.94 ** | −7.52 ** | −13.21 ** | 0.88 | −19.21 ** | −8.77 ** | −24.16 ** |
| Marker Name | Chromosome Number | Number of Alleles | Effective Number of Alleles | Major Allele Frequency | Gene Diversity | PIC |
|---|---|---|---|---|---|---|
| RM315 | 1 | 2 | 1.95 | 0.58 | 0.49 | 0.37 |
| RM543 | 1 | 2 | 1.80 | 0.67 | 0.44 | 0.35 |
| RM263 | 2 | 3 | 2.32 | 0.58 | 0.57 | 0.50 |
| RM279 | 2 | 4 | 3.79 | 0.33 | 0.74 | 0.69 |
| RM55 | 3 | 3 | 2.0 | 0.67 | 0.50 | 0.45 |
| RM518 | 4 | 4 | 3.60 | 0.33 | 0.72 | 0.67 |
| RM159 | 5 | 4 | 3.0 | 0.50 | 0.67 | 0.62 |
| RM3805 | 6 | 3 | 2.57 | 0.50 | 0.61 | 0.54 |
| RM70 | 7 | 3 | 2.57 | 0.50 | 0.61 | 0.54 |
| RM234 | 7 | 3 | 2.57 | 0.50 | 0.61 | 0.54 |
| RM72 | 8 | 4 | 3.60 | 0.33 | 0.72 | 0.67 |
| RM223 | 8 | 4 | 3.60 | 0.33 | 0.72 | 0.67 |
| RM160 | 9 | 3 | 2.57 | 0.50 | 0.61 | 0.54 |
| RM222 | 10 | 3 | 2.57 | 0.50 | 0.61 | 0.54 |
| RM332 | 11 | 3 | 2.0 | 0.67 | 0.50 | 0.45 |
| RM20A | 12 | 5. | 3.79 | 0.33 | 0.74 | 0.69 |
| Mean | 3.31 | 2.77 | 0.49 | 0.62 | 0.55 | |
| Parent | P1 | P2 | P3 | P4 | P5 | P6 |
|---|---|---|---|---|---|---|
| P1 | - | |||||
| P2 | 0.38 | - | ||||
| P3 | 0.78 | 0.67 | - | |||
| P4 | 0.88 | 0.82 | 0.77 | - | ||
| P5 | 0.89 | 0.72 | 0.78 | 0.59 | - | |
| P6 | 0.78 | 0.61 | 0.78 | 0.71 | 0.39 | - |
| Correlation | Irrig. | Days to Heading | Plant Height | Leaf Rolling | Relative Water Content | Chlorophyll Content | Number of Panicles/Plant | Number of Filled Grains/Panicle | Sterility Percentage | 1000-Grain Weight | Grain Yield/Plant |
|---|---|---|---|---|---|---|---|---|---|---|---|
| r (GD, F1) | W | 0.08 | −0.05 | 0.43 | −0.35 | 0.02 | −0.16 | 0.01 | 0.10 | −0.04 | 0.08 |
| S | −0.01 | −0.23 | −0.36 | 0.49 | 0.24 | 0.16 | 0.11 | −0.03 | 0.25 | 0.45 | |
| r (GD, SCA) | W | 0.19 | −0.06 | 0.38 | −0.39 | 0.07 | −0.33 | −0.12 | 0.26 | −0.07 | −0.06 |
| S | 0.12 | −0.29 | −0.33 | 0.34 | 0.27 | 0.07 | −0.02 | 0.08 | 0.23 | 0.31 | |
| r (GD, MP) | W | 0.22 | −0.03 | 0.34 | −0.29 | 0.19 | −0.33 | −0.13 | 0.14 | 0.03 | −0.03 |
| S | 0.15 | −0.18 | −0.42 | 0.25 | 0.35 | 0.13 | −0.01 | 0.04 | 0.24 | 0.26 | |
| r (GD, BP) | W | 0.40 | 0.09 | 0.38 | −0.15 | 0.13 | −0.35 | −0.22 | 0.44 | −0.05 | −0.14 |
| S | 0.34 | 0.07 | −0.14 | 0.20 | 0.34 | 0.15 | −0.11 | 0.05 | 0.29 | 0.09 | |
| r (F1, SCA) | W | 0.80 ** | 0.65 ** | 0.91 ** | 0.92 ** | 0.95 ** | 0.90 ** | 0.93 ** | 0.88 ** | 0.96 ** | 0.84 ** |
| S | 0.81 ** | 0.66 ** | 0.83 ** | 0.84 ** | 0.95 ** | 0.95 ** | 0.92 ** | 0.91 ** | 0.98 ** | 0.82 ** | |
| r (F1, MP) | V | 0.68 ** | 0.53 * | 0.88 ** | 0.86 ** | 0.89 ** | 0.84 ** | 0.91 ** | 0.68 ** | 0.91 ** | 0.77 ** |
| S | 0.69 ** | 0.52 * | 0.72 ** | 0.69 | 0.96 ** | 0.91 ** | 0.89 ** | 0.94 ** | 0.94 ** | 0.70 ** | |
| r (F1, BP) | W | 0.73 ** | 0.53 * | 0.89 ** | 0.83 ** | 0.92 ** | 0.77 ** | 0.84 ** | 0.66 ** | 0.87 ** | 0.80 ** |
| S | 0.75 ** | 0.55 * | 0.90 ** | 0.78 ** | 0.93 ** | 0.92 ** | 0.75 ** | 0.91 ** | 0.92 | 0.66 ** | |
| r (SCA, MP) | W | 0.95 ** | 0.95 ** | 0.99 ** | 0.97 ** | 0.96 ** | 0.98 ** | 0.99 ** | 0.92 ** | 0.98 ** | 0.97 ** |
| S | 0.96 ** | 0.97 ** | 0.95 ** | 0.95 ** | 0.95 ** | 0.97 ** | 0.98 ** | 0.98 ** | 0.97 ** | 0.97 ** | |
| r (SCA, BP) | W | 0.89 ** | 0.83 ** | 0.94 ** | 0.91 ** | 0.97 ** | 0.92 ** | 0.96 ** | 0.86 ** | 0.96 ** | 0.88 ** |
| S | 0.88 ** | 0.77 ** | 0.88 ** | 0.91 ** | 0.95 ** | 0.97 ** | 0.92 ** | 0.97 ** | 0.93 ** | 0.93 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakran, R.M.; Ghazy, M.I.; Rehan, M.; Alsohim, A.S.; Mansour, E. Molecular Genetic Diversity and Combining Ability for Some Physiological and Agronomic Traits in Rice under Well-Watered and Water-Deficit Conditions. Plants 2022, 11, 702. https://doi.org/10.3390/plants11050702
Sakran RM, Ghazy MI, Rehan M, Alsohim AS, Mansour E. Molecular Genetic Diversity and Combining Ability for Some Physiological and Agronomic Traits in Rice under Well-Watered and Water-Deficit Conditions. Plants. 2022; 11(5):702. https://doi.org/10.3390/plants11050702
Chicago/Turabian StyleSakran, Raghda M., Mohamed I. Ghazy, Medhat Rehan, Abdullah S. Alsohim, and Elsayed Mansour. 2022. "Molecular Genetic Diversity and Combining Ability for Some Physiological and Agronomic Traits in Rice under Well-Watered and Water-Deficit Conditions" Plants 11, no. 5: 702. https://doi.org/10.3390/plants11050702
APA StyleSakran, R. M., Ghazy, M. I., Rehan, M., Alsohim, A. S., & Mansour, E. (2022). Molecular Genetic Diversity and Combining Ability for Some Physiological and Agronomic Traits in Rice under Well-Watered and Water-Deficit Conditions. Plants, 11(5), 702. https://doi.org/10.3390/plants11050702