
Use of Different Organic Carbon Sources in Cynara cardunculus Cells: Effects on Biomass Productivity and Secondary Metabolites


 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
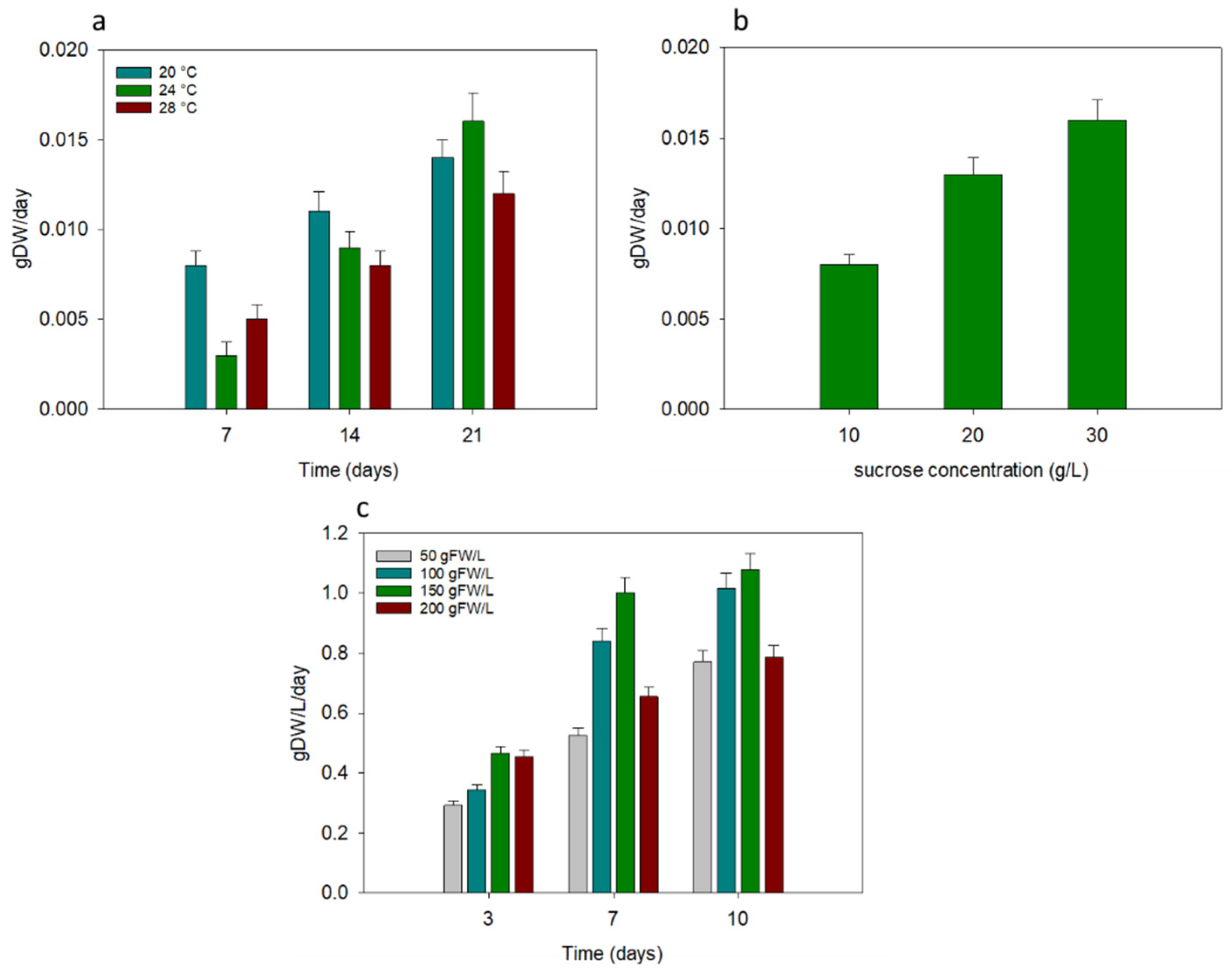
2.1. Optimization of Growth Conditions for Cynara cardunculus In Vitro Cell Cultures
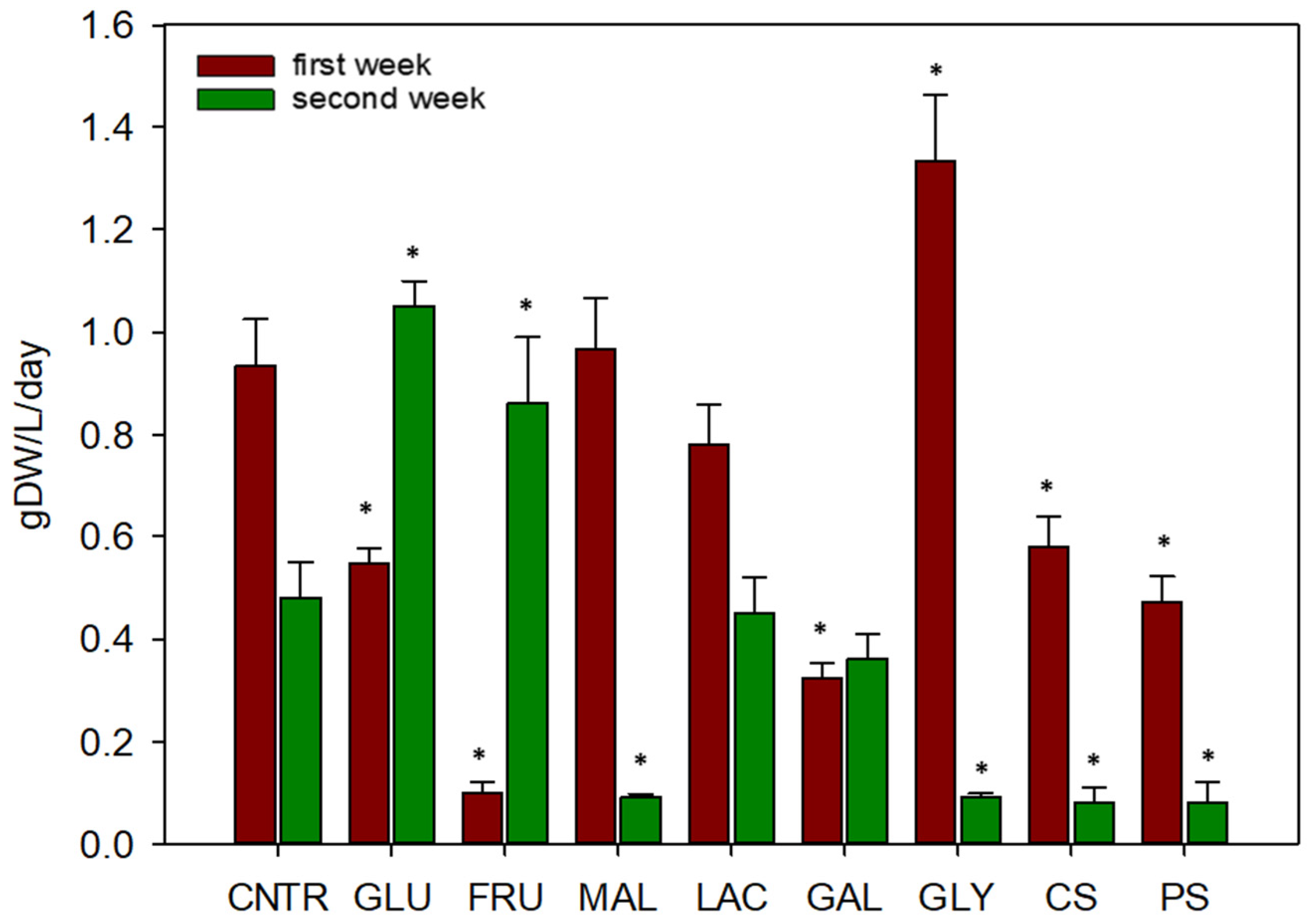
2.2. Utilization of Organic Carbon Sources by Cynara cardunculus Cellular Suspensions
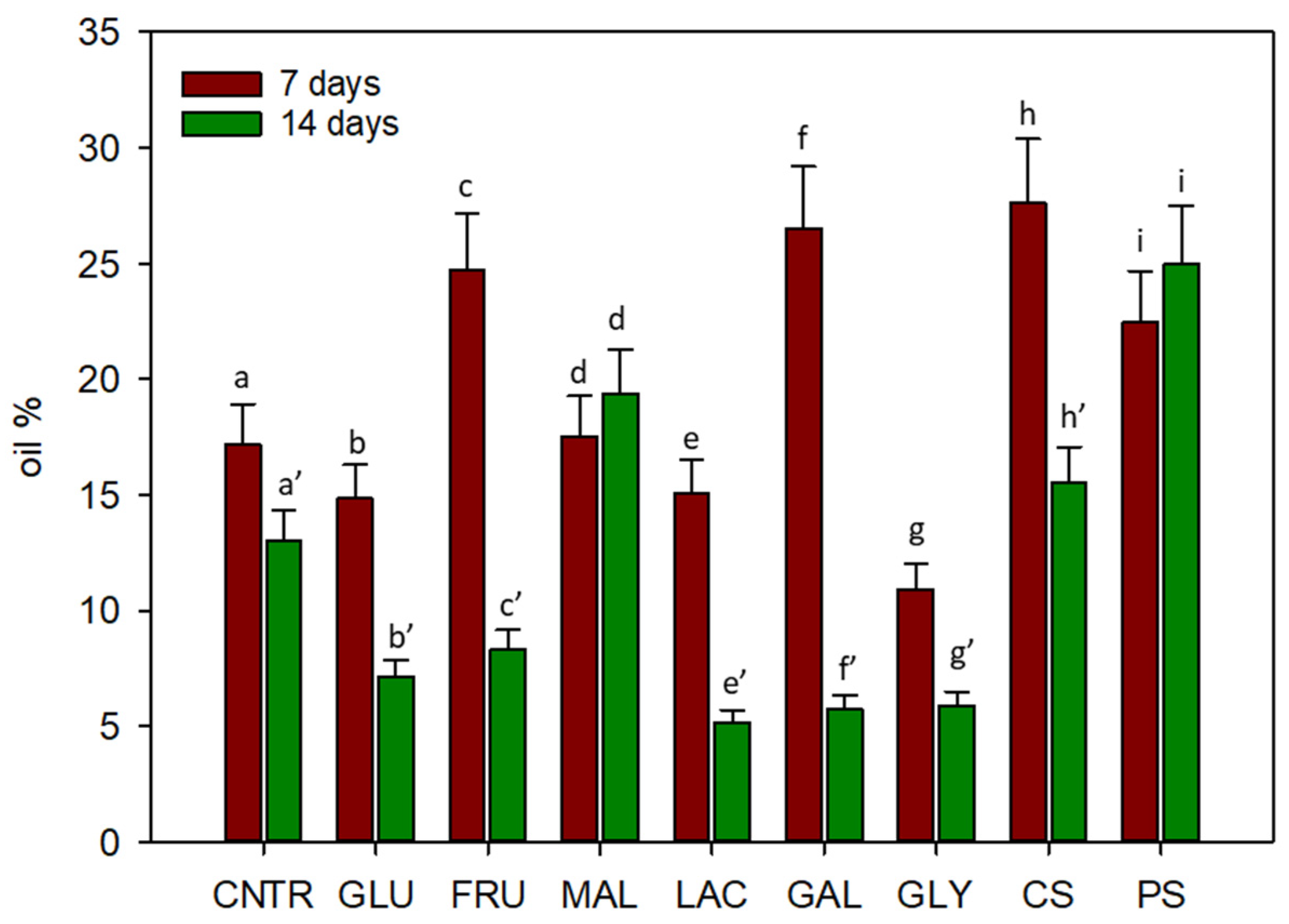
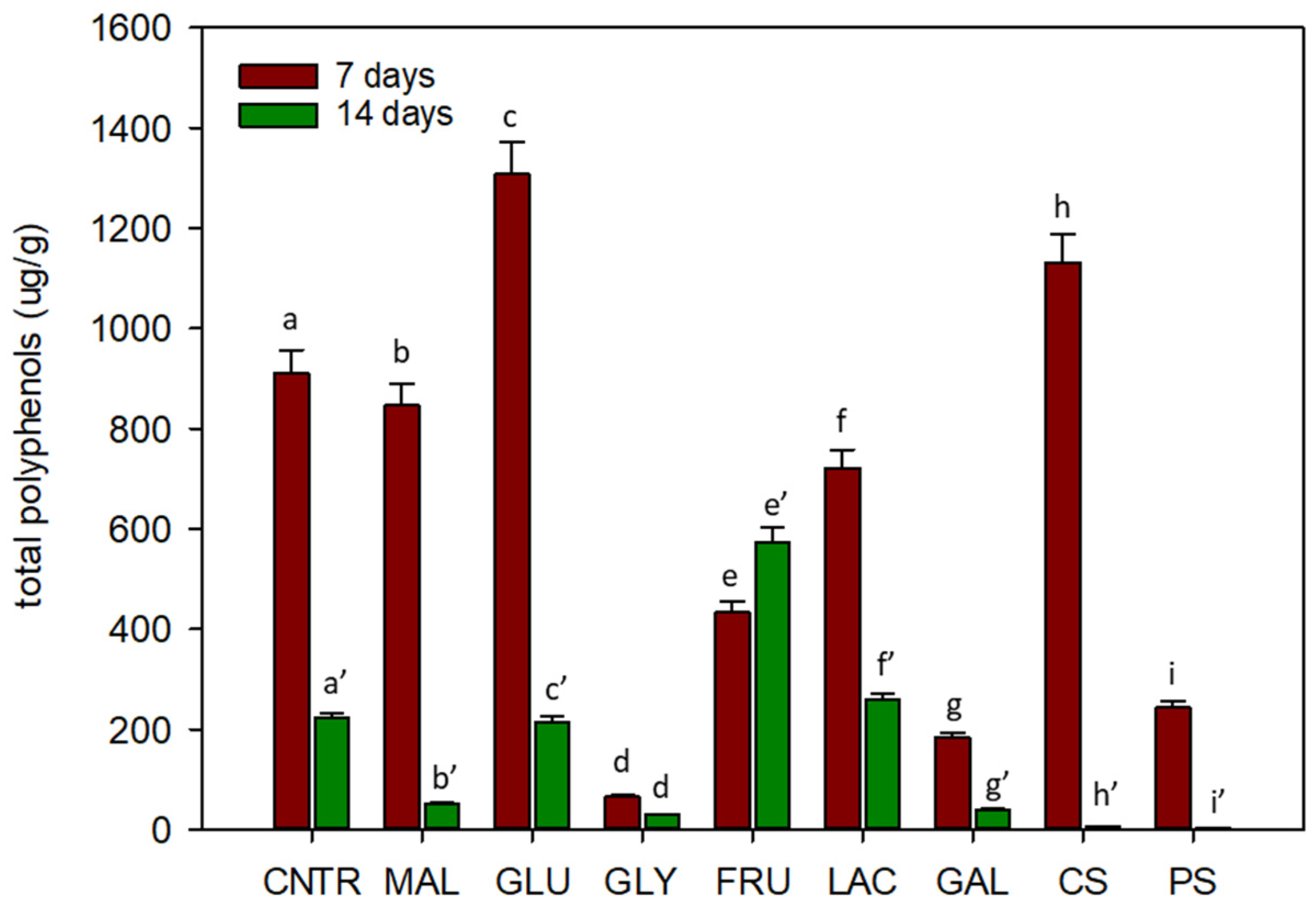
2.3. Chemical Analysis
2.3.1. Fatty Acids Analysis
2.3.2. Polyphenols Analysis
3. Discussion
4. Materials and Methods
4.1. In Vitro Cell Cultures of Cynara cardunculus: Optimization of Growth Conditions
4.2. Cynara cardunculus Cellular Suspensions: Growth Test on Organic Carbon Sources
4.3. Chemical Analysis
4.3.1. Lipid Extraction
4.3.2. Fatty Acids Analysis
4.3.3. Extraction of Polyphenols
4.3.4. UHPLC-HRMS Analysis of Polyphenols
4.3.5. Total Polyphenolic Content Determination
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Toscano, V.; Sollima, L.; Genovese, C.; Melilli, M.G.; Raccuia, S.A. Pilot plant system for biodiesel and pellet production from cardoon: Technical and economic feasibility. Acta Hortic. 2016, 1147, 429–442. [Google Scholar] [CrossRef]
- Fernández, J.; Curt, M.D.; Aguado, P.L. Industrial applications of Cynara cardunculus L. for energy and other uses. Ind. Crops Prod. 2006, 24, 222–229. [Google Scholar] [CrossRef]
- Borgognone, D.; Cardarelli, M.; Rea, E.; Lucini, L.; Colla, G. Salinity source-induced changes in yield, mineral composition, phenolic acids and flavonoids in leaves of artichoke and cardoon grown in floating system. J. Sci. Food Agric. 2014, 94, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.A.; Pereira, C.; Tzortzakis, N.; Barros, L.; Ferreira, I.C.F.R. Nutritional value and bioactive compounds characterization of plant parts from cynara cardunculus L. (asteraceae) cultivated in central Greece. Front. Plant. Sci. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curt, M.D.; Sánchez, G.; Fernández, J. The potential of Cynara cardunculus L. for seed oil production in a perennial cultivation system. Biomass Bioenergy 2002, 23, S0961–S9534. [Google Scholar] [CrossRef]
- Raccuia, S.A.; Melilli, M.G. Biomass and grain oil yields in Cynara cardunculus L. genotypes grown in a Mediterranean environment. Field Crop. Res. 2007, 101, 187–197. [Google Scholar] [CrossRef]
- Raccuia, S.A.; Piscioneri, I.; Sharma, N.; Melilli, M.G. Genetic variability in Cynara cardunculus L. domestic and wild types for grain oil production and fatty acids composition. Biomass Bioenergy 2011, 35, 3167–3173. [Google Scholar] [CrossRef]
- Hussain, M.S.; Fareed, S.; Ansari, S.; Rahman, M.A.; Ahmad, I.Z.; Saeed, M. Current approaches toward production of secondary plant metabolites. J. Pharm. Bioallied Sci. 2012, 4, 10–20. [Google Scholar] [CrossRef]
- Comino, C.; Moglia, A.; Repetto, A.; Tavazza, R. Globe Artichoke Tissue Culture and Its Biotechnological Application; Springer: Berlin/Heidelberg, Germany, 2019; pp. 41–64. [Google Scholar]
- Ikeuchi, M.; Sugimoto, K.; Iwase, A. Plant callus: Mechanisms of induction and repression. Plant. Cell 2013, 25, 3159–3173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ordas, R.J.; Tavazza, R.; Ancora, G. In vitro morphogenesis in the globe artichoke (Cynara scolymus L.). Plant Sci. 1990, 71, 233–237. [Google Scholar] [CrossRef]
- Ruta, C.; Tagarelli, A.; Campanelli, A.; De Mastro, G.; Morone-Fortunato, I. Callogenesis capability of artichoke (Cynara cardunculus var. scolymus L. Fiori). Acta Hortic. 2013, 983, 377–380. [Google Scholar] [CrossRef]
- Menin, B.; Moglia, A.; Comino, C.; Hakkert, J.C.; Lanteri, S.; Beekwilder, J. In vitro callus-induction in globe artichoke (Cynara cardunculus L. var. scolymus) as a system for the production of caffeoylquinic acids. J. Hortic. Sci. Biotechnol. 2013, 88, 537–542. [Google Scholar] [CrossRef]
- Kchouk, M.L.; Mliki, A.; Chatibi, A.; Ghorbel, A. In vitro organogenesis and transgenosis aspects in globe artichoke (Cynara scolymus L.). Cell. Mol. Biol. 1997, 43, 399–408. [Google Scholar]
- Moglia, A.; Menin, B.; Comino, C.; Lanteri, S.; Beekwilder, J. Globe artichoke callus as an alternative system for the production of dicaffeoylquinic acids. Acta Hortic. 2012, 961, 261–265. [Google Scholar] [CrossRef]
- Stepan-Sarkissian, G.; Fowler, M.W. Carbohydrates by Suspension Cultures. Plant Physiol. 1977, 59, 151–181. [Google Scholar]
- De Paiva Neto, V.B.; Otoni, W.C. Carbon sources and their osmotic potential in plant tissue culture: Does it matter? Sci. Hortic. 2003, 97, 193–202. [Google Scholar] [CrossRef]
- Mello, M.O.; Dias, C.T.S.; Amaral, A.F.C.; Melo, M. Growth of Bauhinia forficata Link, Curcuma zedoaria Roscoe and Phaseolus vulgaris L. cell suspension cultures with carbon sources. Sci. Agric. 2001, 58, 481–485. [Google Scholar] [CrossRef] [Green Version]
- Gertlowski, C.; Petersen, M. Influence of the carbon source on growth and rosmarinic acid production in suspension cultures of Coleus blumei. Plant. Cell. Tissue Organ. Cult. 1993, 34, 183–190. [Google Scholar] [CrossRef]
- Günter, E.A.; Ovodov, Y.S. Changes in cell wall polysaccharides of Silene vulgaris callus during culture. Phytochemistry 2002, 59, S0031–S9422. [Google Scholar] [CrossRef]
- Häkkinen, S.T.; Nygren, H.; Nohynek, L.; Puupponen-Pimiä, R.; Heiniö, R.L.; Maiorova, N.; Rischer, H.; Ritala, A. Plant cell cultures as food—Aspects of sustainability and safety. Plant. Cell Rep. 2020, 39, 1655–1668. [Google Scholar] [CrossRef]
- Clifford, M.N.; Wu, W.; Kirkpatrick, J.; Kuhnert, N. Profiling the Chlorogenic Acids and Other Caffeic Acid Derivatives of Herbal Chrysanthemum by LC−MS n. J. Agric. Food Chem. 2007, 55, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Wu, W.; Kirkpatrick, J.; Jaiswal, R.; Kuhnert, N. Profiling and characterisation by liquid chromatography/multi-stage mass spectrometry of the chlorogenic acids in Gardeniae Fructus. Rapid Commun. Mass Spectrom. 2010, 24, 3109–3120. [Google Scholar] [CrossRef] [PubMed]
- Limberg, M.; Cress, D.; Lark, K.G. Variants of Soybean Cells Which Can Grow in Suspension with Maltose as a Carbon-Energy Source. Plant. Physiol. 1979, 63, 718–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandino, G.; Meneghini, M.; Tavazza, R.; Lombardo, S.; Mauromicale, G. Phytochemicals accumulation and antioxidant activity in callus and suspension cultures of Cynara scolymus L. Plant. Cell. Tissue Organ. Cult. 2017, 128, 223–230. [Google Scholar] [CrossRef]
- Lattanzio, V.; Caretto, S.; Linsalata, V.; Colella, G.; Mita, G. Signal transduction in artichoke [Cynara cardunculus L. subsp. scolymus (L.) Hayek] callus and cell suspension cultures under nutritional stress. Plant. Physiol. Biochem. 2018, 127. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, N.R.; De Winter, W.; Van Iren, F.; Verpoorte, R. Initiation, growth and cryopreservation of plant cell suspension cultures. Nat. Protoc. 2011, 6, 715–742. [Google Scholar] [CrossRef]
- Schripsema, J.; Meijer, A.H.; van Iren, F.; ten Hoopen, H.J.G.; Verpoorte, R. Dissimilation curves as a simple method for the characterization of growth of plant cell suspension cultures. Plant. Cell. Tissue Organ. Cult. 1990, 22, 55–64. [Google Scholar] [CrossRef]
- Kieran, P.M.; MacLoughlin, P.F.; Malone, D.M. Plant cell suspension cultures: Some engineering considerations. J. Biotechnol. 1997, 59, 39–52. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, Y.; Wang, J.; Zhou, Y. Glycerol affects root development through regulation of multiple pathways in Arabidopsis. PLoS ONE 2014, 9, e86269. [Google Scholar] [CrossRef] [PubMed]
- Ben-Hayyim, G.; Neumann, H. Stimulatory Effect of Glycerol on Growth and Somatic Embryogenesis in Citrus Callus Cultures. Z. Pflanzenphysiol. 1983, 110, 331–337. [Google Scholar] [CrossRef]
- Ibrahim, H.A.; Abdellatif, Y.M.R. Effect of maltose and trehalose on growth, yield and some biochemical components of wheat plant under water stress. Ann. Agric. Sci. 2016, 61, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Chaubet, N.; Pareilleux, A. Characterization of β-Galactosidases of Medicago sativa Suspension-cultured Cells Growing on Lactose. Effect of the Growth Substrates on the Activities. Z. Pflanzenphysiol. 1982, 106, 401–407. [Google Scholar] [CrossRef]
- Callebaut, A.; Motte, J.-C. Growth of Cucumber Cells in Media with Lactose or Milk Whey as Carbon Source. Plant. Cell Rep. 1988, 7, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Krook, J.; Vreugdenhil, D.; Van Der Plas, L.H.W. Uptake and phosphorylation of glucose and fructose in Daucus carota cell suspensions are differently regulated. Plant. Physiol. Biochem. 2000, 38, S0981–S9428. [Google Scholar] [CrossRef]
- Botha, F.C.; O’Kennedy, M.M. Carbohydrate utilisation by cell suspension cultures of Phaseolus vulgaris. Physiol. Plant. 1998, 102, 429–436. [Google Scholar] [CrossRef]
- Cabasson, C.; Ollitrault, P.; Côte, F.-X.; Michaux-Ferriére, N.; Dambier, D.; Dalnic, R.; Teisson, C. Characteristics of Citrus cell cultures during undifferentiated growth on sucrose and somatic embryogenesis on galactose. Physiol. Plant. 1995, 93, 464–470. [Google Scholar] [CrossRef]
- Graziani, G.; Docimo, T.; De Palma, M.; Sparvoli, F.; Izzo, L.; Tucci, M.; Ritieni, A. Changes in phenolics and fatty acids composition and related gene expression during the development from seed to leaves of three cultivated cardoon genotypes. Antioxidants 2020, 9, 1096. [Google Scholar] [CrossRef]
- Lattanzio, V.; Linsalata, V.; Palmieri, S.; Van Sumere, C.F. The beneficial effect of citric and ascorbic acid on the phenolic browning reaction in stored artichoke (Cynara scolymus L.) heads. Food Chem. 1989, 33, 93–106. [Google Scholar] [CrossRef]
- Paolo, D.; Locatelli, F.; Cominelli, E.; Pirona, R.; Pozzo, S.; Graziani, G.; Ritieni, A.; De Palma, M.; Docimo, T.; Tucci, M.; et al. Towards a cardoon (Cynara cardunculus var. altilis)-based biorefinery: A case study of improved cell cultures via genetic modulation of the phenylpropanoid pathway. Int. J. Mol. Sci. 2021, 22, 1978. [Google Scholar] [CrossRef]
- Box, J.D. Investigation of the Folin-Ciocalteau phenol reagent for the determination of polyphenolic substances in natural waters. Water Res. 1983, 17, 511–525. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CNTR | MAL | GLU | GLY | FRU | LAC | GAL | CS | PS | |
|---|---|---|---|---|---|---|---|---|---|
| Palmitic (C16:0) | 46.2 ± 0.6 a | 52.1 ± 0.2 b | 38.9 ± 0.2 c | 47.8 ± 3.1 | 40 ± 0.5 d | 49.1 ± 0.2 | 40.8 ± 0.3 d | 45.1 ± 0.1 a | 36.4 ± 0.7 e |
| Stearic (C18:0) | 7.6 ± 0.1 a | 21.5 ± 0.2 b | 8.4 ± 0.1 c | 12.2 ± 0.8 d | 9.5 ± 0.1 e | 13 ± 0.1 d | 21.3 ± 0.1 b | 29.8 ± 0.1 f | 15.5 ± 0.3 g |
| Oleic (C18:1) | 6.7 ± 1.8 a | 1.5 ± 0.1 b | 12.3 ± 0.8 c | 17 ± 5.4 d | 12.6 ± 1.2 c | 4 ± 1.3 e | 11.3 ± 0.1 f | 12.2 ± 0.2 c | 16.3 ± 1.1 g |
| Linoleic (C18:2) | 27.3 ± 0.8 a | 16.9 ± 0.1 b | 29.5 ± 0.4 c | 21.7 ± 1.3 d | 27.3 ± 0.4 a | 21.8 ± 0.5 d | 19.9 ± 0.1 e | 11.0 ± 0.1 f | 31.1 ± 0.2 c |
| Linolenic (C18:3) | 9.8 ± 0.2 a | 6.9 ± 0.2 b | 9 ± 0.1 c | n.d. | 9.2 ± 0.2 c | 9.9 ± 0.6 a | 5.4 ± 0.3 d | n.d. | n.d. |
| Others | 2.4 ± 0.3 a | 0.9 ± 0.1 b | 1.9 ± 0.1 c | 1.3 ± 0.2 d | 1.3 ± 0.2 d | 2.1 ± 0.2 c | 1.2 ± 0.1 b | 0.9 ± 0.1 b | 0.6 ± 0.2 e |
| Peak ID | CNTR | MAL | GLU | GLY | FRU | LAC | GAL | CS | PS | |
|---|---|---|---|---|---|---|---|---|---|---|
| 3-CQA | 1 | 44.1 ± 3.1 a | 5.3 ± 0.4 b | 39.6 ± 3.2 c | 2.1 ± 0.1 d | 175.8 ± 10.5 e | 61.9 ± 4.9 f | 2.9 ± 0.2 d | 0.8 ± 0.1 g | 0.4 ± 0.1 h |
| p-Coumaric acid | 2 | 7 ± 0.5 a | 2.8 ± 0.2 b | 4.6 ± 0.4 c | 1.6 ± 0.1 d | 7.1 ± 0.5 a | 4.4 ± 0.4 c | 2.2 ± 0.2 d | 0.7 ± 0.1 e | 0.7 ± 0.1 e |
| 3-FQA | 3 | 6.4 ± 0.5 a | 6.3 ± 0.4 a | 7.1 ± 0.3 b | 7.8 ± 0.5 c | 5.6 ± 0.4 d | 8.4 ± 0.7 e | 3.3 ± 0.2 f | n.d. | n.d. |
| Ferulic acid | 4 | 6.6 ± 0.5 a | 1.3 ± 0.1 b | 7.1 ± 0.6 c | 1.7 ± 0.1 d | 16.1 ± 0.8 e | 2.7 ± 0.2 f | 1.8 ± 0.1 d | 0.5 ± 0.1 g | 0.3 ± 0.1 h |
| 5-FQA | 5 | 9.5 ± 0.7 a | 2.6 ± 0.2 b | 7.1 ± 0.6 c | 2.9 ± 0.2 d | 15.6 ± 1.1 e | 12.4 ± 0.7 f | 3.3 ± 0.2 g | n.d. | n.d. |
| 3,4-DiCQA | 6 | 9.1 ± 0.5 a | 5.4 ± 0.4 b | 20.5 ± 1.4 c | 7.3 ± 0.4 d | 17.8 ± 0.9 e | 8 ± 0.6 f | 15.6 ± 0.9 g | 0.5 ± 0.1 h | 0.2 ± 0.1 i |
| 1,5-DiCQA | 7 | 137.4 ± 8.2 a | 25.3 ± 2 b | 128 ± 7.7 c | 6.4 ± 0.4 d | 332.8 ± 23.3 e | 159.8 ± 12.8 f | 9.9 ± 0.9 g | 1.7 ± 0.1 h | 1.1 ± 0.1 i |
| 5-iFQA | 8 | 0.4 ± 0.1 a | 0.2 ± 0.1 b | 0.4 ± 0.1 a | 0.2 ± 0.1 b | 2.4 ± 0.2 c | 0.7 ± 0.1 d | n.d. | 0.2 ± 0.1 b | n.d. |
| Compounds | RT | Molecular Formula | Theoretical Mass [M-H]− | Experimental Mass [M-H]− | MS/MS Ions | Accuracy (Δppm) |
|---|---|---|---|---|---|---|
| 3-CQA | 8.30 | C16H17O9 | 353.08798 | 353.08783 | 191.05571–179.03461 | −0.42 |
| p-Coumaric acid | 9.58 | C9H8O3 | 163.03917 | 163.03912 | 119.04981 | −0.31 |
| 3-FQA | 9.75 | C17H19O9 | 367.10345 | 367.10335 | 193.04961 | −0.27 |
| Ferulic acid | 9.88 | C10H10O4 | 193.05063 | 193.05016 | 179.05556 | −2.43 |
| 5-FQA | 10.82 | C17H19O9 | 367.10345 | 367.10315 | 191.05501 | −0.82 |
| 3,4-DiCQA | 11.21 | C25H23O12 | 515.11950 | 515.11969 | 353.08755–191.05561 | 0.37 |
| 1,5-DiCQA | 11.64 | C25H23O12 | 515.11950 | 515.11957 | 353.08753–191.05550 | 0.14 |
| 5-iFQA | 11.86 | C17H19O9 | 367.10345 | 367.10309 | 193.04915 | −0.98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliviero, M.; Langellotti, A.L.; Russo, G.L.; Baselice, M.; Donadio, A.; Ritieni, A.; Graziani, G.; Masi, P. Use of Different Organic Carbon Sources in Cynara cardunculus Cells: Effects on Biomass Productivity and Secondary Metabolites. Plants 2022, 11, 701. https://doi.org/10.3390/plants11050701
Oliviero M, Langellotti AL, Russo GL, Baselice M, Donadio A, Ritieni A, Graziani G, Masi P. Use of Different Organic Carbon Sources in Cynara cardunculus Cells: Effects on Biomass Productivity and Secondary Metabolites. Plants. 2022; 11(5):701. https://doi.org/10.3390/plants11050701
Chicago/Turabian StyleOliviero, Maria, Antonio Luca Langellotti, Giovanni L. Russo, Marco Baselice, Andrea Donadio, Alberto Ritieni, Giulia Graziani, and Paolo Masi. 2022. "Use of Different Organic Carbon Sources in Cynara cardunculus Cells: Effects on Biomass Productivity and Secondary Metabolites" Plants 11, no. 5: 701. https://doi.org/10.3390/plants11050701
APA StyleOliviero, M., Langellotti, A. L., Russo, G. L., Baselice, M., Donadio, A., Ritieni, A., Graziani, G., & Masi, P. (2022). Use of Different Organic Carbon Sources in Cynara cardunculus Cells: Effects on Biomass Productivity and Secondary Metabolites. Plants, 11(5), 701. https://doi.org/10.3390/plants11050701