
Red Light and Glucose Enhance Cytokinin-Mediated Bud Initial Formation in Physcomitrium patens


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture and Growth Conditions
2.2. Light Treatment
2.3. Phytohormone Estimation
3. Results
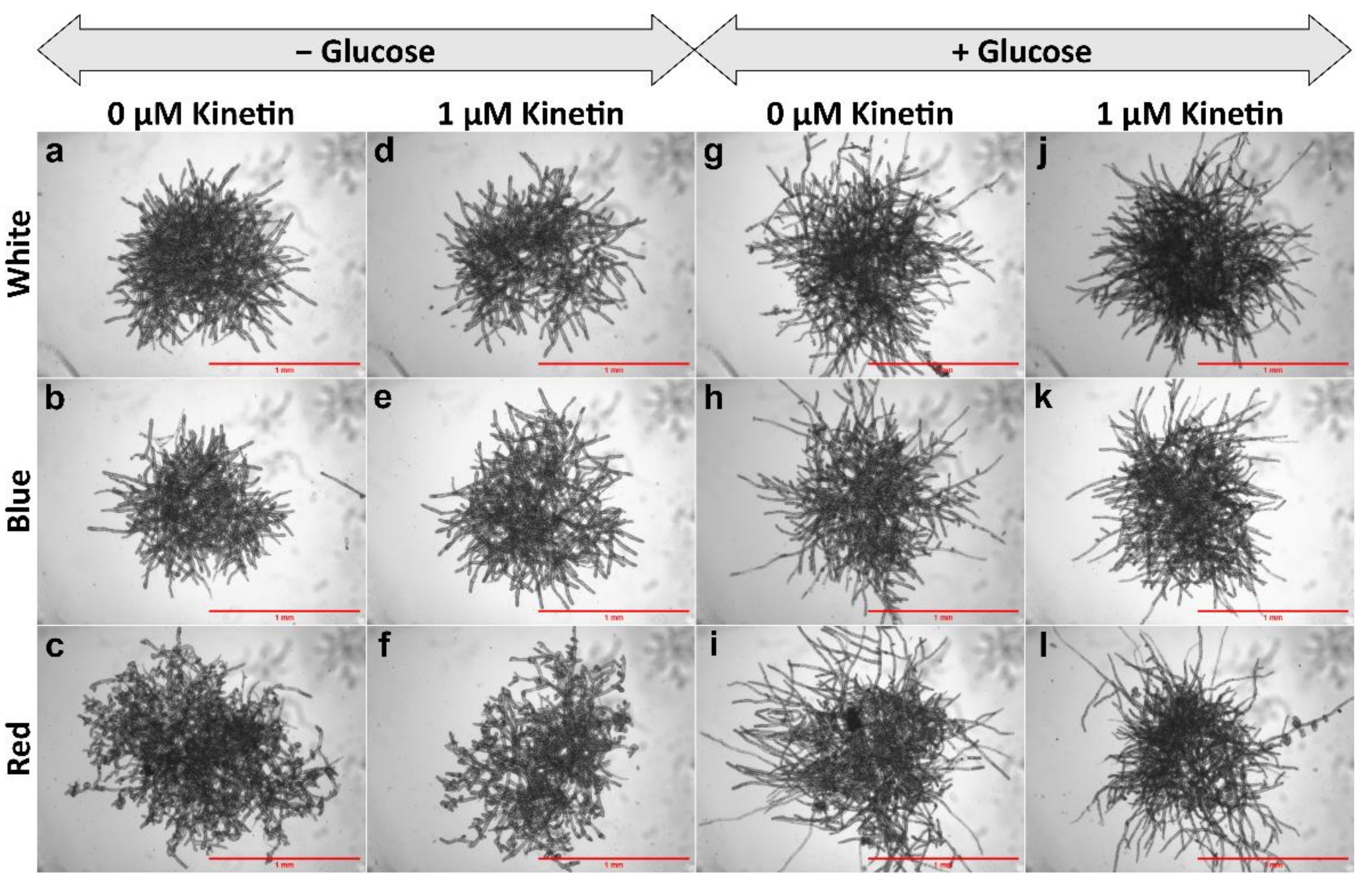
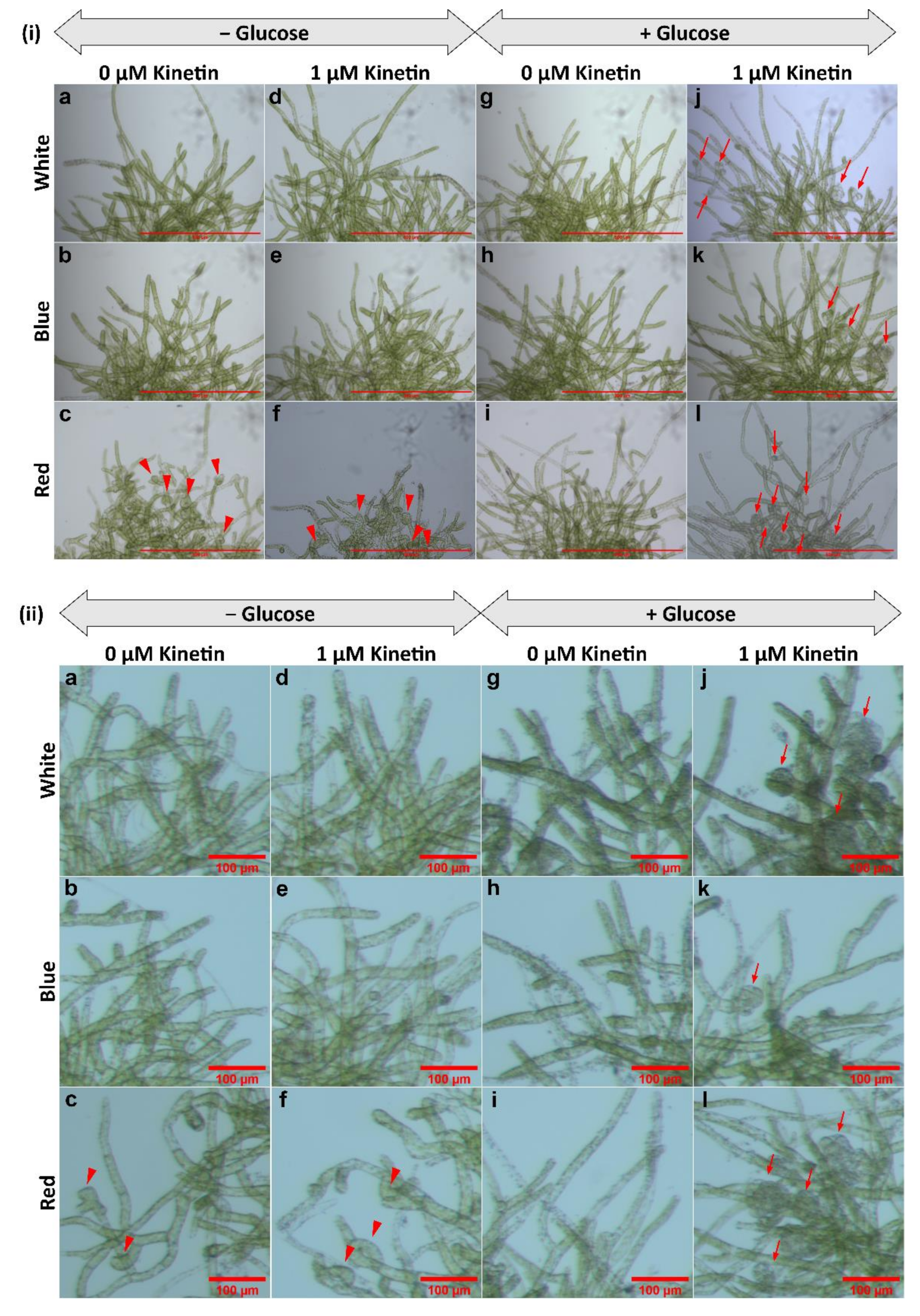
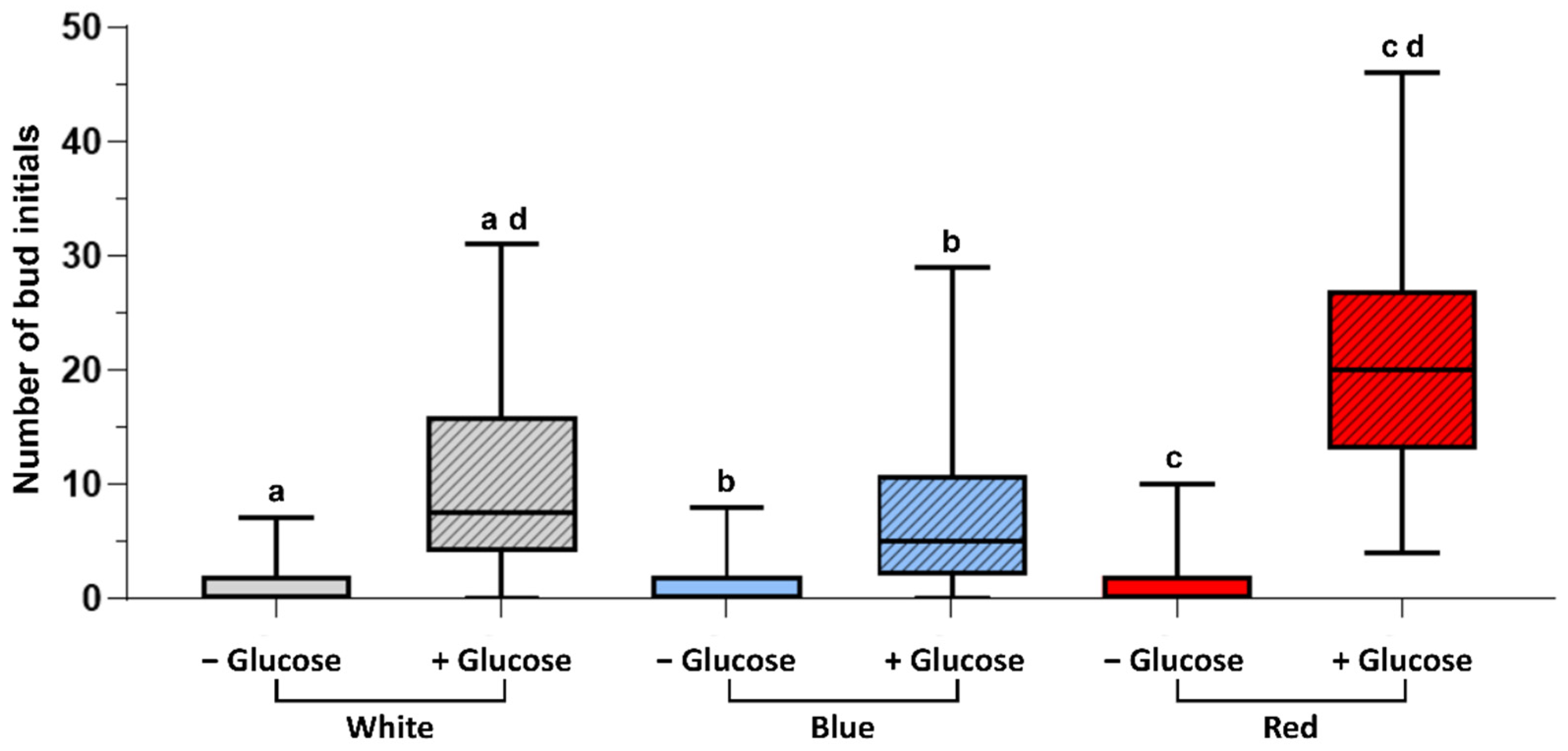
3.1. Gametophore Bud Formation Is Increased in Red Light and in Presence of Glucose
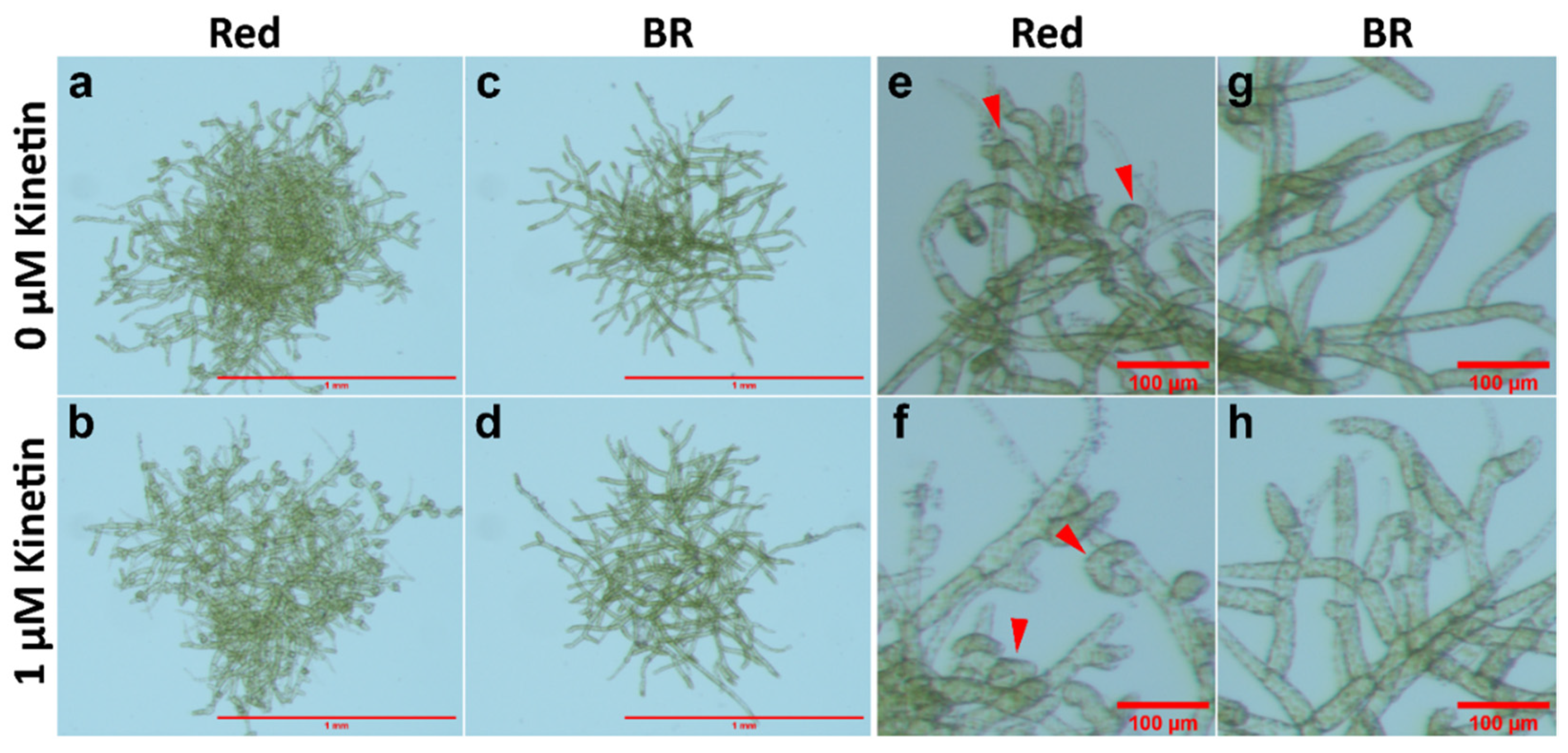
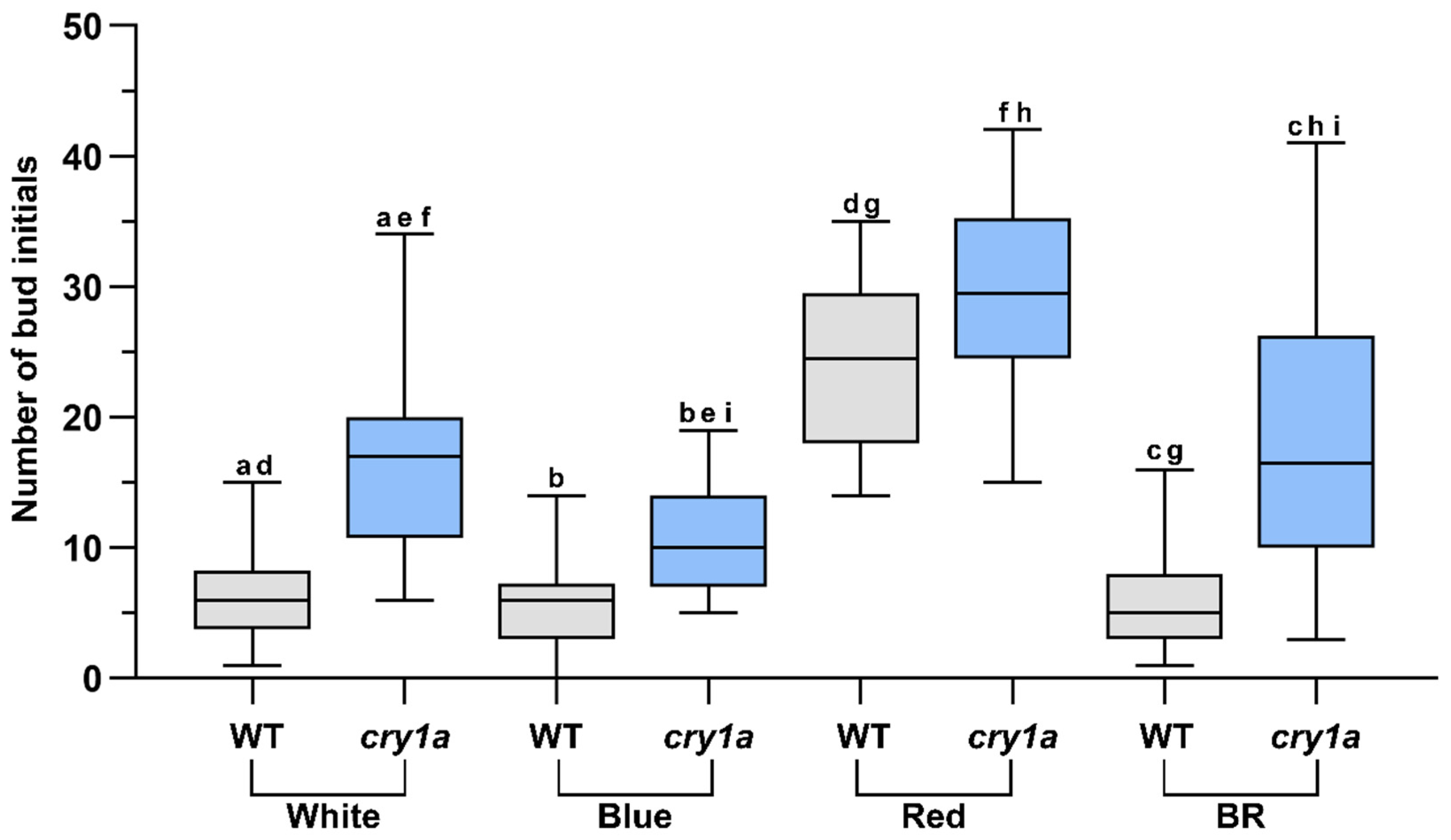
3.2. cry1a Mutants Produce More Number of Gametophore Buds Than WT
3.3. Gametophore Buds Are Not Formed under FR Light
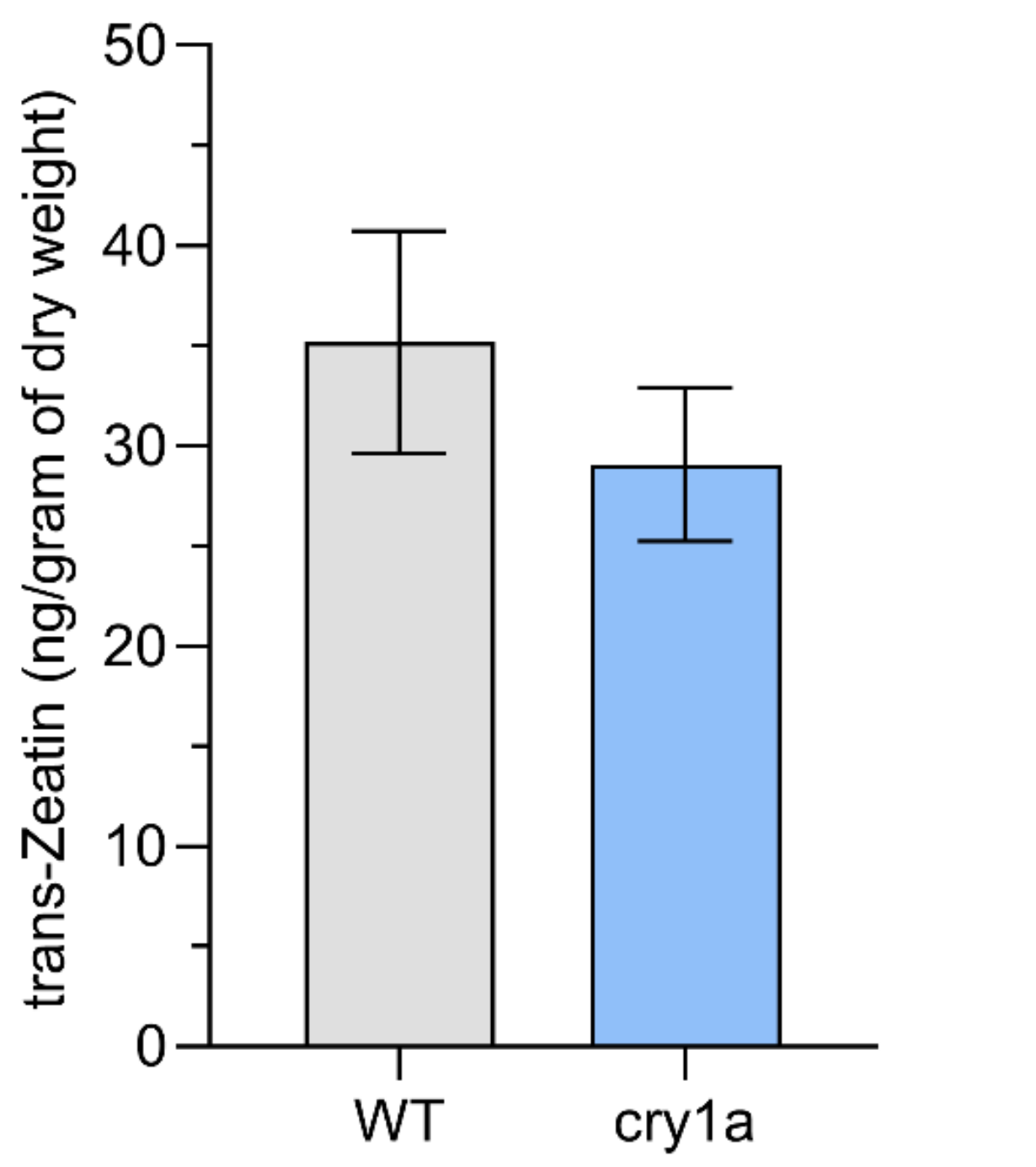
3.4. Phytohoromone Estimation
4. Discussion
4.1. Red Light and Glucose Enhance the Effect of Cytokinin
4.2. CRY1a Suppresses Bud Initial Development
4.3. FR Light Is Inhibitory for Bud Initial Development
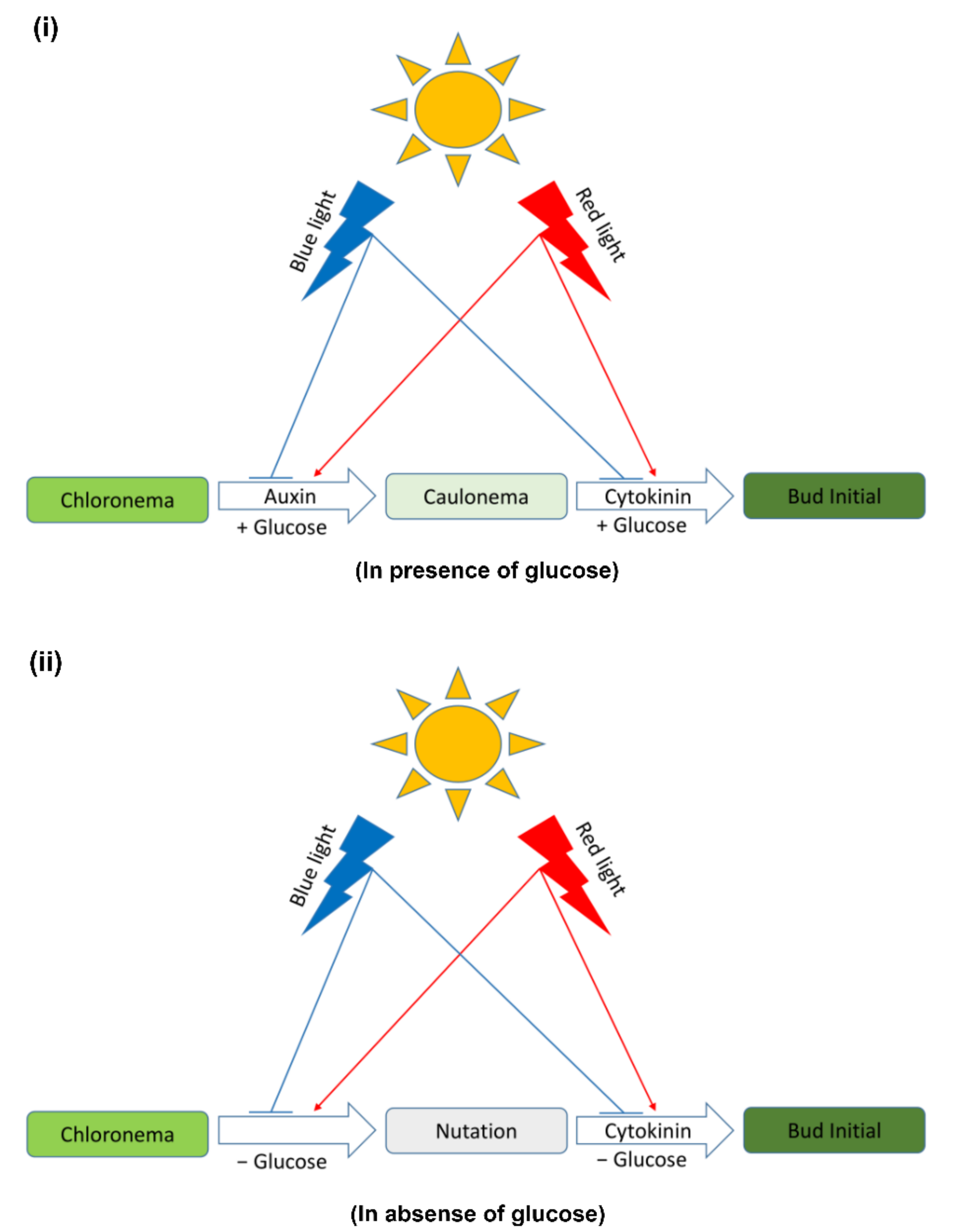
4.4. Light Quality and Carbon Supply May Influence the 2D–3D Transition in Physcomitrium
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rensing, S.A.; Goffinet, B.; Meyberg, R.; Wu, S.-Z.; Bezanilla, M. The moss Physcomitrium (Physcomitrella) patens: A model organism for non-seed plants. Plant Cell 2020, 32, 1361–1376. [Google Scholar] [CrossRef] [Green Version]
- Beike, A.K.; von Stackelberg, M.; Schallenberg-Rüdinger, M.; Hanke, S.T.; Follo, M.; Quandt, D.; McDaniel, S.F.; Reski, R.; Tan, B.C.; Rensing, S.A. Molecular evidence for convergent evolution and allopolyploid speciation within the Physcomitrium-Physcomitrella species complex. BMC Evol. Biol. 2014, 14, 158. [Google Scholar] [CrossRef] [Green Version]
- Medina, R.; Johnson, M.G.; Liu, Y.; Wickett, N.J.; Shaw, A.J.; Goffinet, B. Phylogenomic delineation of Physcomitrium (Bryophyta: Funariaceae) based on targeted sequencing of nuclear exons and their flanking regions rejects the retention of Physcomitrella, Physcomitridium and Aphanorrhegma. J. Syst. Evol. 2019, 57, 404–417. [Google Scholar] [CrossRef] [Green Version]
- Takezawa, D.; Komatsu, K.; Sakata, Y. ABA in bryophytes: How a universal growth regulator in life became a plant hormone? J. Plant Res. 2011, 124, 437–453. [Google Scholar] [CrossRef]
- Thelander, M.; Landberg, K.; Sundberg, E. Auxin-mediated developmental control in the moss Physcomitrella patens. J. Exp. Bot. 2018, 69, 277–290. [Google Scholar] [CrossRef]
- von Schwartzenberg, K.; Núñez, M.F.; Blaschke, H.; Dobrev, P.I.; Novák, O.; Motyka, V.; Strnad, M. Cytokinins in the bryophyte Physcomitrella patens: Analyses of activity, distribution, and cytokinin oxidase/dehydrogenase overexpression reveal the role of extracellular cytokinins. Plant Physiol. 2007, 145, 786–800. [Google Scholar] [CrossRef] [Green Version]
- Guillory, A.; Bonhomme, S. Phytohormone biosynthesis and signaling pathways of mosses. Plant Mol. Biol. 2021, 107, 245–277. [Google Scholar] [CrossRef]
- Hayashi, K.; Horie, K.; Hiwatashi, Y.; Kawaide, H.; Yamaguchi, S.; Hanada, A.; Nakashima, T.; Nakajima, M.; Mander, L.N.; Yamane, H.; et al. Endogenous diterpenes derived from ent-kaurene, a common gibberellin precursor, regulate protonema differentiation of the moss Physcomitrella patens. Plant Physiol. 2010, 153, 1085–1097. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Kawaide, H.; Notomi, M.; Sakigi, Y.; Matsuo, A.; Nozaki, H. Identification and functional analysis of bifunctional ent-kaurene synthase from the moss Physcomitrella patens. FEBS Lett. 2006, 580, 6175–6181. [Google Scholar] [CrossRef] [Green Version]
- Coudert, Y.; Palubicki, W.; Ljung, K.; Novak, O.; Leyser, O.; Harrison, C.J. Three ancient hormonal cues co-ordinate shoot branching in a moss. Elife 2015, 4, e06808. [Google Scholar] [CrossRef] [Green Version]
- Yasumura, Y.; Pierik, R.; Fricker, M.D.; Voesenek, L.A.C.J.; Harberd, N.P. Studies of Physcomitrella patens reveal that ethylene-mediated submergence responses arose relatively early in land-plant evolution. Plant J. 2012, 72, 947–959. [Google Scholar] [CrossRef]
- Johri, M.M.; Desai, S. Auxin regulation of caulonema formation in moss protonema. Nat. New Biol. 1973, 245, 223–224. [Google Scholar] [CrossRef]
- Ashton, N.W.; Grimsley, N.H.; Cove, D.J. Analysis of gametophytic development in the moss, Physcomitrella patens, using auxin and cytokinin resistant mutants. Planta 1979, 144, 427–435. [Google Scholar] [CrossRef]
- Bhatla, S.C.; Bopp, M. The hormonal regulation of protonema development in mosses III. Auxin-resistant mutants of the moss Funaria hygrometrica Hedw. J. Plant Physiol. 1985, 120, 233–243. [Google Scholar] [CrossRef]
- Bhatla, S.C.; Kapoor, S.; Khurana, J.P. Involvement of calcium in auxin-induced cell differentiation in the protonema of the wild strain and auxin mutants of the moss Funaria hygrometrica. J. Plant Physiol. 1996, 147, 547–552. [Google Scholar] [CrossRef]
- Johri, M.M. Caulonema differentiation in Funaria protonema. Int. J. Dev. Biol. 2020, 64, 21–28. [Google Scholar] [CrossRef]
- Hoffmann, B.; Proust, H.; Belcram, K.; Labrune, C.; Boyer, F.D.; Rameau, C.; Bonhomme, S. Strigolactones inhibit caulonema elongation and cell division in the moss Physcomitrella patens. PLoS ONE 2014, 9, e99206. [Google Scholar] [CrossRef]
- Mitra, G.C.; Allsopp, A. Effects of kinetin, gibberellic acid and certain auxins on the development of shoot buds on the protonema of Pohlia nutans. Nature 1959, 183, 974–975. [Google Scholar] [CrossRef]
- Szweykowska, A. Kinetin-induced formation of gametophores in dark cultures of Ceratodon purpureus. J. Exp. Bot. 1963, 14, 137–141. [Google Scholar] [CrossRef]
- Brandes, H.; Kende, H. Studies on cytokinin-controlled bud formation in moss protonemata. Plant Physiol. 1968, 43, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Szweykowska, A.; Guzowska, I.; Gallas, J. Studies on the activity of kinetin in cultures of Funaria hygrometrica II. Effect of kinetin on isolated protonema cells. Acta Soc. Bot. Pol. 1968, 37, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Chopra, R.N.; Bhatla, S.C. Regulation of gametangial formation in bryophytes. Bot. Rev. 1983, 49, 29–63. [Google Scholar] [CrossRef]
- Chopra, R.N.; Dhingra-Babbar, S. Studies on bud induction in the moss Trematodon brevicalyx Dixon. New Phytol. 1984, 97, 613–620. [Google Scholar] [CrossRef]
- Bopp, M.; Jacob, H.J. Cytokinin effect on branching and bud formation in Funaria. Planta 1986, 169, 462–464. [Google Scholar] [CrossRef]
- Arif, M.A.; Hiss, M.; Tomek, M.; Busch, H.; Meyberg, R.; Tintelnot, S.; Reski, R.; Rensing, S.A.; Frank, W. ABA-induced vegetative diaspore formation in Physcomitrella patens. Front. Plant Sci. 2019, 10, 315. [Google Scholar] [CrossRef] [Green Version]
- Yasumura, Y.; Pierik, R.; Kelly, S.; Sakuta, M.; Voesenek, L.A.C.J.; Harberd, N.P. An ancestral role for CONSTITUTIVE TRIPLE RESPONSE1 proteins in both ethylene and abscisic acid signaling. Plant Physiol. 2015, 169, 283–298. [Google Scholar] [CrossRef]
- Biswal, D.P.; Panigrahi, K.C.S. Light- and hormone-mediated development in non-flowering plants: An overview. Planta 2020, 253, 1. [Google Scholar] [CrossRef]
- de Wit, M.; Galvão, V.C.; Fankhauser, C. Light-mediated hormonal regulation of plant growth and development. Annu. Rev. Plant Biol. 2016, 67, 513–537. [Google Scholar] [CrossRef]
- Galvão, V.C.; Fankhauser, C. Sensing the light environment in plants: Photoreceptors and early signaling steps. Curr. Opin. Neurobiol. 2015, 34, 46–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathews, S. Phytochrome-mediated development in land plants: Red light sensing evolves to meet the challenges of changing light environments. Mol. Ecol. 2006, 15, 3483–3503. [Google Scholar] [CrossRef]
- Han, Y.J.; Song, P.S.; Kim, J.I. Phytochrome-mediated photomorphogenesis in plants. J. Plant Biol. 2007, 50, 230–240. [Google Scholar] [CrossRef]
- Wang, Q.; Lin, C. Mechanisms of cryptochrome-mediated photoresponses in plants. Annu. Rev. Plant Biol. 2020, 71, 103–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, T.; Doi, M.; Suetsugu, N.; Kagawa, T.; Wada, M.; Shimazaki, K. phot1 and phot2 mediate blue light regulation of stomatal opening. Nature 2001, 414, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Rizzini, L.; Favory, J.J.; Cloix, C.; Faggionato, D.; O’Hara, A.; Kaiserli, E.; Baumeister, R.; Schäfer, E.; Nagy, F.; Jenkins, G.I.; et al. Perception of UV-B by the Arabidopsis UVR8 protein. Science 2011, 332, 103–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somers, D.E.; Schultz, T.F.; Milnamow, M.; Kay, S.A. ZEITLUPE encodes a novel clock-associated PAS protein from Arabidopsis. Cell 2000, 101, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Paik, I.; Huq, E. Plant photoreceptors: Multi-functional sensory proteins and their signaling networks. Semin. Cell Dev. Biol. 2019, 92, 114–121. [Google Scholar] [CrossRef]
- van Gelderen, K.; Kang, C.; Pierik, R. Light signaling, root development, and plasticity. Plant Physiol. 2018, 176, 1049–1060. [Google Scholar] [CrossRef] [Green Version]
- Kong, S.G.; Okajima, K. Diverse photoreceptors and light responses in plants. J. Plant Res. 2016, 129, 111–114. [Google Scholar] [CrossRef] [Green Version]
- Li, F.-W.; Melkonian, M.; Rothfels, C.J.; Villarreal, J.C.; Stevenson, D.W.; Graham, S.W.; Wong, G.K.S.; Pryer, K.M.; Mathews, S. Phytochrome diversity in green plants and the origin of canonical plant phytochromes. Nat. Commun. 2015, 6, 7852. [Google Scholar] [CrossRef] [Green Version]
- Imaizumi, T.; Kadota, A.; Hasebe, M.; Wada, M. Cryptochrome light signals control development to suppress auxin sensitivity in the moss Physcomitrella patens. Plant Cell 2002, 14, 373–386. [Google Scholar] [CrossRef] [Green Version]
- Kasahara, M.; Kagawa, T.; Sato, Y.; Kiyosue, T.; Wada, M. Phototropins mediate blue and red light-induced chloroplast movements in Physcomitrella patens. Plant Physiol. 2004, 135, 1388–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soriano, G.; Cloix, C.; Heilmann, M.; Núñez-Olivera, E.; Martínez-Abaigar, J.; Jenkins, G.I. Evolutionary conservation of structure and function of the UVR8 photoreceptor from the liverwort Marchantia polymorpha and the moss Physcomitrella patens. New Phytol. 2018, 217, 151–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linde, A.-M.; Eklund, D.M.; Kubota, A.; Pederson, E.R.A.; Holm, K.; Gyllenstrand, N.; Nishihama, R.; Cronberg, N.; Muranaka, T.; Oyama, T.; et al. Early evolution of the land plant circadian clock. New Phytol. 2017, 216, 576–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cove, D.J.; Schild, A.; Ashton, N.W.; Hartmann, E. Genetic and physiological studies of the effect of light on the development of the moss, Physcomitrella patens. Photochem. Photobiol. 1978, 27, 249–254. [Google Scholar] [CrossRef]
- Possart, A.; Fleck, C.; Hiltbrunner, A. Shedding (far-red) light on phytochrome mechanisms and responses in land plants. Plant Sci. 2014, 217–218, 36–46. [Google Scholar] [CrossRef]
- Mittmann, F.; Brücker, G.; Zeidler, M.; Repp, A.; Abts, T.; Hartmann, E.; Hughes, J. Targeted knockout in Physcomitrella reveals direct actions of phytochrome in the cytoplasm. Proc. Natl. Acad. Sci. USA 2004, 101, 13939–13944. [Google Scholar] [CrossRef] [Green Version]
- Bao, L.; Yamamoto, K.T.; Fujita, T. Phototropism in gametophytic shoots of the moss Physcomitrella patens. Plant Signal. Behav. 2015, 10, e1010900. [Google Scholar] [CrossRef] [Green Version]
- Lymperopoulos, P.; Msanne, J.; Rabara, R. Phytochrome and phytohormones: Working in tandem for plant growth and development. Front. Plant Sci. 2018, 9, 1037. [Google Scholar] [CrossRef] [Green Version]
- Thelander, M.; Olsson, T.; Ronne, H. Effect of the energy supply on filamentous growth and development in Physcomitrella patens. J. Exp. Bot. 2005, 56, 653–662. [Google Scholar] [CrossRef] [Green Version]
- Mitra, G.C.; Misra, L.P.; Prabha, C. Interaction of red and blue light on the development of the protonema and bud formation in Pohlia nutans. Planta 1965, 65, 42–48. [Google Scholar] [CrossRef]
- Mitra, G.C.; Misra, L.P.; Kaul, K.N. Effects of coconut milk and kinetin on the development of the protonema and bud formation in Pohlia nutans. Nature 1962, 195, 1219–1220. [Google Scholar] [CrossRef]
- Simon, P.E.; Naef, J.B. Light dependency of the cytokinin-induced bud initiation in protonemata of the moss Funaria hygrometrica. Physiol. Plant. 1981, 53, 13–18. [Google Scholar] [CrossRef]
- Chopra, R.N.; Gupta, U. Dark-induction of buds in Funaria hygrometrica Hedw. Bryologist 1967, 70, 102–104. [Google Scholar] [CrossRef]
- Nebel, B.J.; Naylor, A.W. Light, temperature and carbohydrate requirements for shoot-bud initiation from protonemata in the moss Physcomitrium turbinatum. Am. J. Bot. 1968, 55, 38–44. [Google Scholar] [CrossRef]
- Zavala, A.M.; Pérez, N.M.; Becerra, A.A.; López, M.A.V. Glucose inhibits the shoot bud formation in the moss Bryum billarderi. Cent. Eur. J. Biol. 2012, 7, 648–654. [Google Scholar] [CrossRef] [Green Version]
- Sabovljevic, A.; Sabovljevic, M.; Grubisic, D.; Konjevic, R. The effect of sugars on development of two moss species (Bryum argentum and Atrichum undulatum) during in vitro culture. Belg. J. Bot. 2005, 138, 79–84. [Google Scholar]
- Bopp, M.; Brandes, H. Versuche zur analyse der protonema entwicklung der laubmoose: II. Über den zusammenhang zwischen protonema differenzierung und kinetin wirkung bei der bildung von moos knospen. Planta 1964, 62, 116–136. [Google Scholar] [CrossRef]
- Reski, R.; Abel, W.O. Induction of budding on chloronemata and caulonemata of the moss, Physcomitrella patens, using isopentenyladenine. Planta 1985, 165, 354–358. [Google Scholar] [CrossRef]
- Ashton, N.W.; Cove, D.J. The isolation and preliminary characterisation of auxotrophic and analogue resistant mutants of the moss, Physcomitrella patens. Mol. Gen. Genet. 1977, 154, 87–95. [Google Scholar] [CrossRef]
- Šimura, J.; Antoniadi, I.; Široká, J.; Tarkowská, D.; Strnad, M.; Ljung, K.; Novák, O. Plant hormonomics: Multiple phytohormone profiling by targeted metabolomics. Plant Physiol. 2018, 177, 476–489. [Google Scholar] [CrossRef] [Green Version]
- Szweykowska, A.; Szweykowska, A.; Ratajczak, W.; Schneider, J. Studies on the activity of kinetin in cultures of Funaria hygrometrica I. Bud-inducing activity of kinetin and protein synthesis. Acta Soc. Bot. Pol. 1968, 37, 145–155. [Google Scholar] [CrossRef]
- Moody, L.A. The 2D to 3D growth transition in the moss Physcomitrella patens. Curr. Opin. Plant Biol. 2019, 47, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Kieber, J.J.; Schaller, G.E. Cytokinin signaling in plant development. Development 2018, 145, dev149344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindner, A.C.; Lang, D.; Seifert, M.; Podlešáková, K.; Novák, O.; Strnad, M.; Reski, R.; von Schwartzenberg, K. Isopentenyltransferase-1 (IPT1) knockout in Physcomitrella together with phylogenetic analyses of IPTs provide insights into evolution of plant cytokinin biosynthesis. J. Exp. Bot. 2014, 65, 2533–2543. [Google Scholar] [CrossRef] [Green Version]
- von Schwartzenberg, K.; Lindner, A.C.; Gruhn, N.; Šimura, J.; Novák, O.; Strnad, M.; Gonneau, M.; Nogué, F.; Heyl, A. CHASE domain-containing receptors play an essential role in the cytokinin response of the moss Physcomitrella patens. J. Exp. Bot. 2016, 67, 667–679. [Google Scholar] [CrossRef] [Green Version]
- Hyoung, S.; Cho, S.H.; Chung, J.H.; So, W.M.; Cui, M.H.; Shin, J.S. Cytokinin oxidase PpCKX1 plays regulatory roles in development and enhances dehydration and salt tolerance in Physcomitrella patens. Plant Cell Rep. 2020, 39, 419–430. [Google Scholar] [CrossRef]
- Moody, L.A.; Kelly, S.; Rabbinowitsch, E.; Langdale, J.A. Genetic regulation of the 2D to 3D growth transition in the moss Physcomitrella patens. Curr. Biol. 2018, 28, 473–478.e5. [Google Scholar] [CrossRef] [Green Version]
- Demko, V.; Perroud, P.F.; Johansen, W.; Delwiche, C.F.; Cooper, E.D.; Remme, P.; Ako, A.E.; Kugler, K.G.; Mayer, K.F.X.; Quatrano, R.; et al. Genetic analysis of DEFECTIVE KERNEL1 loop function in three-dimensional body patterning in Physcomitrella patens. Plant Physiol. 2014, 166, 903–919. [Google Scholar] [CrossRef] [Green Version]
- Whitewoods, C.D.; Cammarata, J.; Nemec Venza, Z.; Sang, S.; Crook, A.D.; Aoyama, T.; Wang, X.Y.; Waller, M.; Kamisugi, Y.; Cuming, A.C.; et al. CLAVATA was a genetic novelty for the morphological innovation of 3D Growth in land plants. Curr. Biol. 2018, 28, 2365–2376.e5. [Google Scholar] [CrossRef] [Green Version]
- Moody, L.A.; Kelly, S.; Clayton, R.; Weeks, Z.; Emms, D.M.; Langdale, J.A. NO GAMETOPHORES 2 is a novel regulator of the 2D to 3D growth transition in the moss Physcomitrella patens. Curr. Biol. 2021, 31, 555–563.e4. [Google Scholar] [CrossRef]
- Goss, C.A.; Brockmann, D.J.; Bushoven, J.T.; Roberts, A.W. A CELLULOSE SYNTHASE (CESA) gene essential for gametophore morphogenesis in the moss Physcomitrella patens. Planta 2012, 235, 1355–1367. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, T.; Hiwatashi, Y.; Shigyo, M.; Kofuji, R.; Kubo, M.; Ito, M.; Hasebe, M. AP2-type transcription factors determine stem cell identity in the moss Physcomitrella patens. Development 2012, 139, 3120–3129. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biswal, D.P.; Panigrahi, K.C.S. Red Light and Glucose Enhance Cytokinin-Mediated Bud Initial Formation in Physcomitrium patens. Plants 2022, 11, 707. https://doi.org/10.3390/plants11050707
Biswal DP, Panigrahi KCS. Red Light and Glucose Enhance Cytokinin-Mediated Bud Initial Formation in Physcomitrium patens. Plants. 2022; 11(5):707. https://doi.org/10.3390/plants11050707
Chicago/Turabian StyleBiswal, Durga Prasad, and Kishore Chandra Sekhar Panigrahi. 2022. "Red Light and Glucose Enhance Cytokinin-Mediated Bud Initial Formation in Physcomitrium patens" Plants 11, no. 5: 707. https://doi.org/10.3390/plants11050707
APA StyleBiswal, D. P., & Panigrahi, K. C. S. (2022). Red Light and Glucose Enhance Cytokinin-Mediated Bud Initial Formation in Physcomitrium patens. Plants, 11(5), 707. https://doi.org/10.3390/plants11050707