
The Virome of ‘Lamon Bean’: Application of MinION Sequencing to Investigate the Virus Population Associated with Symptomatic Beans in the Lamon Area, Italy



Abstract
:1. Introduction
2. Results
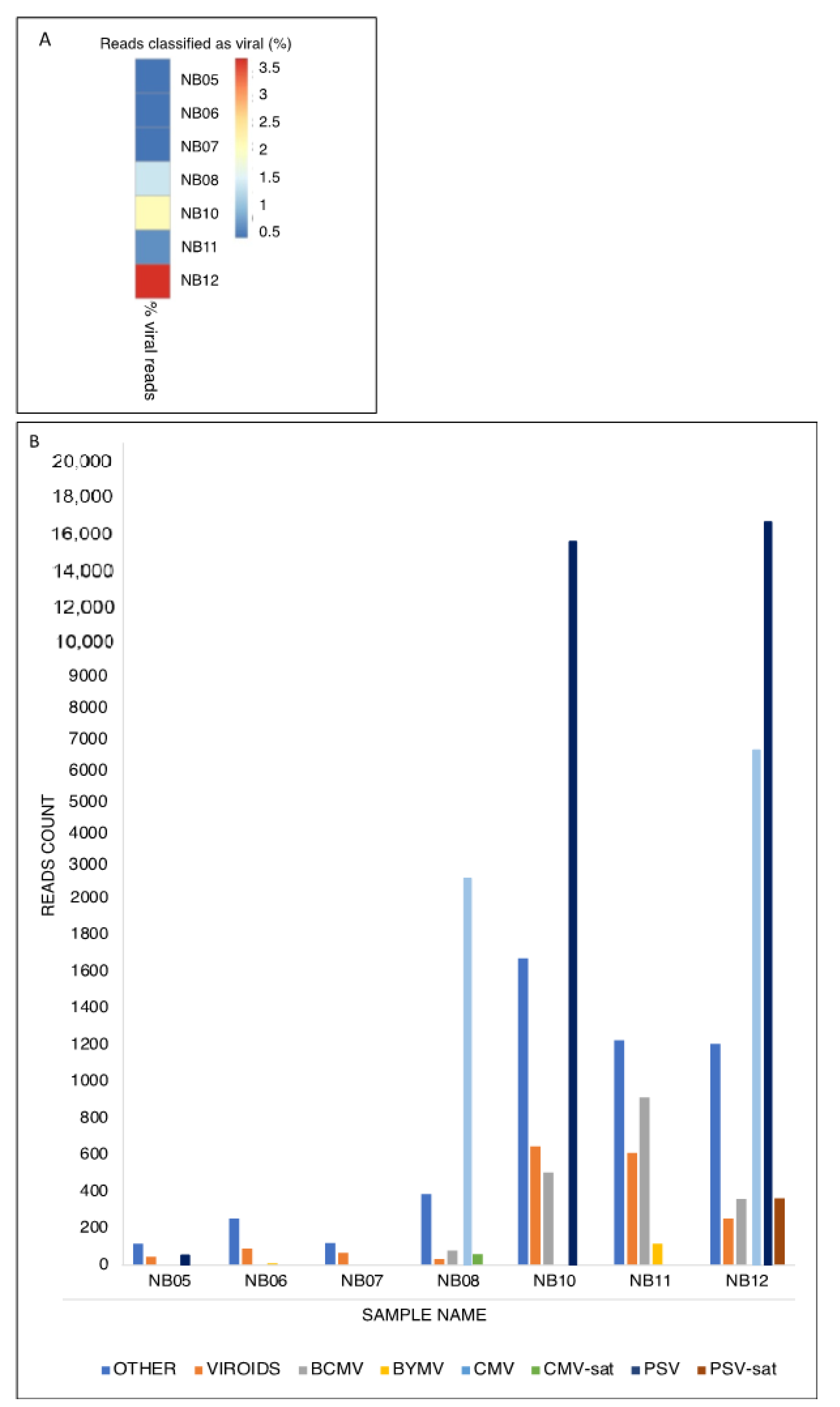
2.1. Virome Determination
2.2. RT-PCR and ELISA Detection
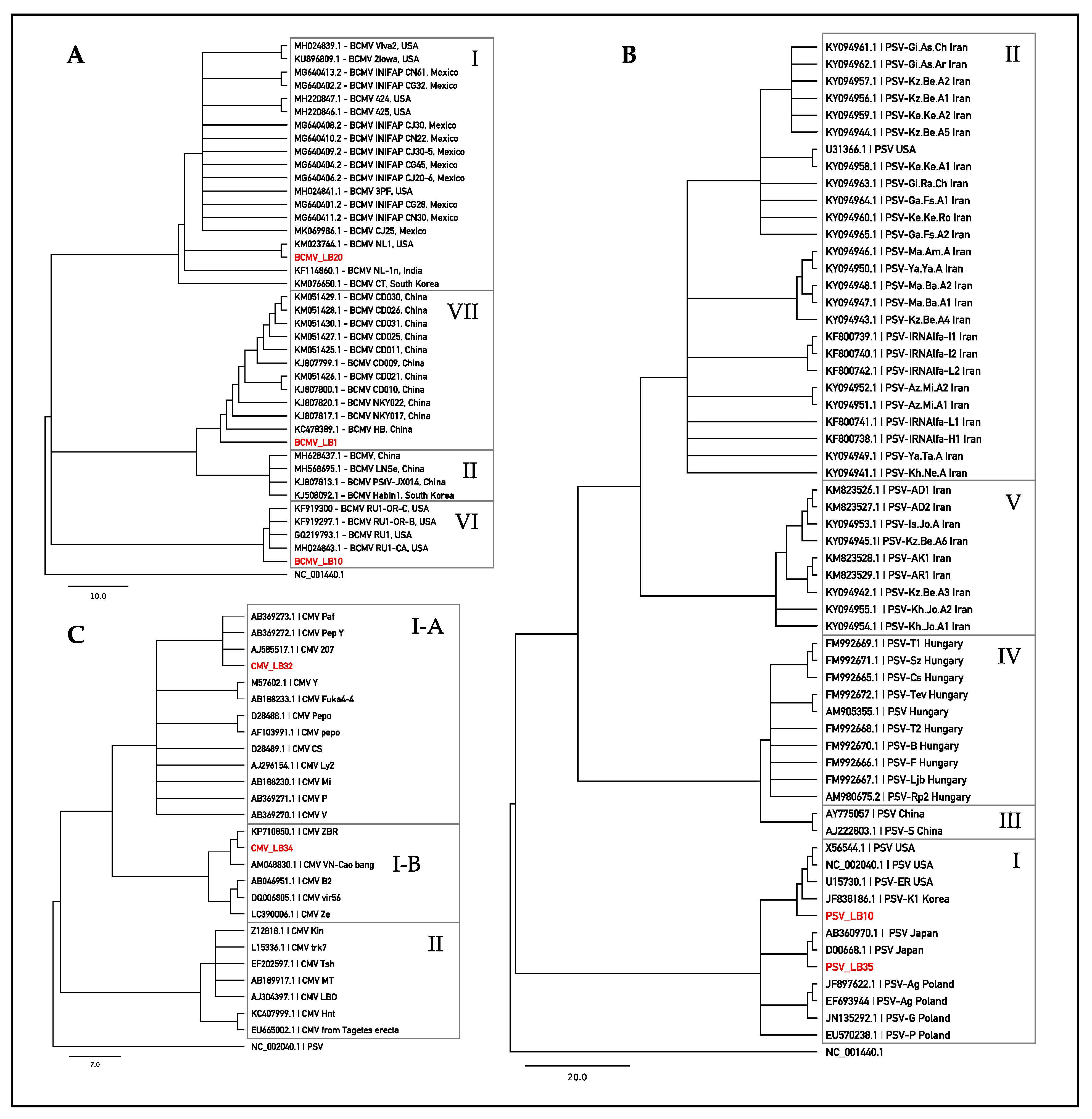
2.3. BLAST Analysis of Sanger Sequences and Phylogenetic Investigation
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. RNA Extraction and cDNA Synthesis
4.3. cDNA Library Preparation
4.4. MinION Sequencing
4.5. Data Acquisition
4.6. Validation of Sequencing Data by RT-PCR
4.7. BLAST Analyses of Sanger Sequences and Phylogenetic Investigation
4.8. Serological Detection
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rojas, M.R.; Gilbertson, R.L. Emerging plant viruses: A diversity of mechanisms and opportunities. In Plant Virus Evolution; Roossinck, M.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 27–51. ISBN 978-3-540-75763-4. [Google Scholar]
- Lecoq, H.; Moury, B.; Desbiez, C.; Palloix, A.; Pitrat, M. Durable virus resistance in plants through conventional approaches: A challenge. Virus Res. 2004, 100, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Worrall, E.A.; Wamonje, F.O.; Mukeshimana, G.; Harvey, J.J.; Carr, J.P.; Mitter, N. Bean Common Mosaic Virus and Bean Common Mosaic Necrosis Virus. Adv. Virus Res. 2015, 93, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Fang, G.; Allison, R.; Zambolim, E.; Maxwell, D.; Gilbertson, R. The Complete Nucleotide Sequence and Genome Organization of Bean Common Mosaic Virus (NL3 Strain). Virus Res. 1995, 39, 13–23. [Google Scholar] [CrossRef]
- Drijfhout, E.; Silbernagel, M.J.; Burke, D.W. Differentiation of strains of bean common mosaic virus. Eur. J. Plant Pathol. 1978, 84, 13–26. [Google Scholar] [CrossRef]
- Larsen, R.C.; Miklas, P.N.; Druffel, K.L.; Wyatt, S.D. NL-3 K Strain Is a Stable and Naturally Occurring Interspecific Recombinant Derived from Bean common mosaic necrosis virus and Bean common mosaic virus. Phytopathology 2005, 95, 1037–1042. [Google Scholar] [CrossRef] [Green Version]
- Palukaitis, P.; Roossinck, M.J.; Dietzgen, R.G.; Francki, R.I.B. Cucumber Mosaic Virus. Adv. Virus Res. 1992, 41, 281–348. [Google Scholar]
- Gildow, F.E.; Shah, D.; Sackett, W.M.; Butzler, T.; Nault, B.A.; Fleischer, S.J. Transmission Efficiency of Cucumber mosaic virus by Aphids Associated with Virus Epidemics in Snap Bean. Phytopathology 2008, 98, 1233–1241. [Google Scholar] [CrossRef] [Green Version]
- Roossinck, M.J. Cucumber mosaic virus, a model for RNA virus evolution. Mol. Plant Pathol. 2001, 2, 59–63. [Google Scholar] [CrossRef]
- Roossinck, M.J.; Zhang, L.; Hellwald, K.-H. Rearrangements in the 5 J Nontranslated Region and Phylogenetic Analyses of Cucumber Mosaic Virus RNA 3 Indicate Radial Evolution of Three Subgroups. J. Virol. 1999, 73, 7. [Google Scholar] [CrossRef] [Green Version]
- Wylie, S.J.; Jones, R.A.C. Role of Recombination in the Evolution of Host Specialization Within Bean yellow mosaic virus. Phytopathology 2009, 99, 512–518. [Google Scholar] [CrossRef] [Green Version]
- Riechmann, J.L.; Lain, S.; Garcia, J.A. Highlights and prospects of potyvirus molecular biology. J. Gen. Virol. 1992, 73, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Urcuqui-Inchima, S.; Haenni, A.-L.; Bernardi, F. Potyvirus proteins: A wealth of functions. Virus Res. 2001, 74, 157–175. [Google Scholar] [CrossRef]
- Wylie, S.J.; Coutts, B.A.; Jones, M.G.K.; Jones, R.A.C. Phylogenetic Analysis of Bean yellow mosaic virus Isolates from Four Continents: Relationship Between the Seven Groups Found and Their Hosts and Origins. Plant Dis. 2008, 92, 1596–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovisolo, O.; Hull, R.; Rösler, O. Coevolution of Viruses with Hosts and Vectors and Possible Paleontology. Adv. Appl. Microbiol. 2003, 62, 325–379. [Google Scholar] [CrossRef]
- Xu, Z.; Barnett, O.; Gibson, P. Characterization of Peanut Stunt Virus strains by host reactions, serology, and RNA patterns. Phytopathology 1986, 76, 390–395. [Google Scholar] [CrossRef]
- Roossinck, M.J.; Sleat, D.; Palukaitis, P. Satellite RNAs of plant viruses: Structures and biological effects. Microbiol. Rev. 1992, 56, 265–279. [Google Scholar] [CrossRef]
- Mink, G.I.; Silbernagel, M.J.; Saksena, K.N. Host range, purification, and properties of the western strain of peanut stunt virus. Phytopathol. 1969, 59, 1625–1631. [Google Scholar]
- Diaz-Ruiz, J.; Kaper, J. Nucleotide sequence relationships among thirty Peanut Stunt Virus isolates determined by competition hybridization. Arch. Virol. 1983, 75, 277–281. [Google Scholar] [CrossRef]
- Chalupowicz, L.; Dombrovsky, A.; Gaba, V.; Luria, N.; Reuven, M.; Beerman, A.; Lachman, O.; Dror, O.; Nissan, G.; Manulis-Sasson, S. Diagnosis of plant diseases using the Nanopore sequencing platform. Plant Pathol. 2018, 68, 229–238. [Google Scholar] [CrossRef]
- Adams, I.P.; Glover, R.H.; Monger, W.A.; Mumford, R.; Jackeviciene, E.; Navalinskiene, M.; Samuitiene, M.; Boonham, N. Next-generation sequencing and metagenomic analysis: A universal diagnostic tool in plant virology. Mol. Plant Pathol. 2009, 10, 537–545. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Daubert, S.; Golino, D.; Islas, C.; Rowhani, A. Comparison of Next-Generation Sequencing Versus Biological Indexing for the Optimal Detection of Viral Pathogens in Grapevine. Phytopathology 2015, 105, 758–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Ding, S.-W.; Zhang, Y.; Zhu, S. Identification of Viruses and Viroids by Next-Generation Sequencing and Homology-Dependent and Homology-Independent Algorithms. Annu. Rev. Phytopathol. 2015, 53, 425–444. [Google Scholar] [CrossRef] [PubMed]
- Roossinck, M.J. Deep sequencing for discovery and evolutionary analysis of plant viruses. Virus Res. 2017, 239, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.; Choi, H.; Kim, S.-M.; Kim, S.-L.; Lee, B.C.; Cho, W.K. The pepper virome: Natural co-infection of diverse viruses and their quasispecies. BMC Genom. 2017, 18, 453. [Google Scholar] [CrossRef] [Green Version]
- Czotter, N.; Molnar, J.; Szabó, E.; Demian, E.; Kontra, L.; Baksa, I.; Szittya, G.; Kocsis, L.; Deák, T.; Bisztray, G.; et al. NGS of Virus-Derived Small RNAs as a Diagnostic Method Used to Determine Viromes of Hungarian Vineyards. Front. Microbiol. 2018, 9, 122. [Google Scholar] [CrossRef] [Green Version]
- Abadkhah, M.; Hajizadeh, M.; Koolivand, D. Global population genetic structure ofBean common mosaic virus. Arch. Phytopathol. Plant Prot. 2020, 53, 266–281. [Google Scholar] [CrossRef]
- Wang, D.F.; Wang, J.R.; Cui, L.Y.; Wang, S.T.; Niu, Y. Molecular identification and phylogeny of cucumber mosaic virus and zucchini yellow mosaic virus co-infecting Luffa cylindrica L. in Shanxi, China. J. Plant Pathol. 2020, 102, 477–487. [Google Scholar] [CrossRef]
- Amid-Motlagh, M.H.; Massumi, H.; Heydarnejad, J.; Mehrvar, M.; Hajimorad, M.R. Nucleotide sequence analyses of coat protein gene of peanut stunt virus isolates from alfalfa and different hosts show a new tentative subgroup from Iran. Virus Dis. 2017, 28, 295–302. [Google Scholar] [CrossRef]
- Bravo, E.; Calvert, L.A.; Morales, F.J. The complete nucleotide sequence of the genomic RNA of Bean common mosaic virus strain NL4. Genetica 2008, 32, 37–46. [Google Scholar]
- Martin, K.; Hill, J.H.; Cannon, S. Occurrence and Characterization of Bean common mosaic virus Strain NL1 in Iowa. Plant Dis. 2014, 98, 1593. [Google Scholar] [CrossRef]
- Feng, X.; Poplawsky, A.R.; Nikolaeva, O.V.; Myers, J.R.; Karasev, A. Recombinants of Bean common mosaic virus (BCMV) and Genetic Determinants of BCMV Involved in Overcoming Resistance in Common Bean. Phytopathology 2014, 104, 786–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; Orellana, G.E.; Myers, J.R.; Karasev, A.V. Recessive Resistance to Bean common mosaic virus Conferred by the bc-1 and bc-2 Genes in Common Bean (Phaseolus vulgaris) Affects Long-Distance Movement of the Virus. Phytopathology 2018, 108, 1011–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.C.; Naidu, R.A.; Aboul-Ata, A.E.; Ghabrial, S.A. Evidence for the occurrence of two distinct subgroups of peanut stunt cucumovirus strains: Molecular characterization of RNA3. J. Gen. Virol. 1997, 78, 929–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraile, A.; Alonso-Prados, J.L.; Aranda, M.A.; Bernal, J.J.; Malpica, J.M.; García-Arenal, F. Genetic exchange by recombination or reassortment is infrequent in natural populations of a tripartite RNA plant virus. J. Virol. 1997, 71, 934–940. [Google Scholar] [CrossRef] [Green Version]
- Roossinck, M.J. Evolutionary History of Cucumber Mosaic Virus Deduced by Phylogenetic Analyses. J. Virol. 2002, 76, 3382–3387. [Google Scholar] [CrossRef] [Green Version]
- Pooggin, M.M. Small RNA-Omics for Plant Virus Identification, Virome Reconstruction, and Antiviral Defense Characterization. Front. Microbiol. 2018, 9, 2779. [Google Scholar] [CrossRef]
- Wang, Y.; Gaba, V.; Yang, J.; Palukaitis, P.; Gal-On, A. Characterization of Synergy Between Cucumber mosaic virus and Potyviruses in Cucurbit Hosts. Phytopathology 2002, 92, 51–58. [Google Scholar] [CrossRef]
- Chiquito-Almanza, E.; Acosta-Gallegos, J.A.; García-Álvarez, N.C.; Garrido-Ramírez, E.R.; Montero-Tavera, V.; Guevara-Olvera, L.; Anaya-López, J.L. Simultaneous Detection of both RNA and DNA Viruses Infecting Dry Bean and Occurrence of Mixed Infections by BGYMV, BCMV and BCMNV in the Central-West Region of Mexico. Viruses 2017, 9, 63. [Google Scholar] [CrossRef] [Green Version]
- Moreno, A.B.; Lopez-Moya, J.J. When viruses play team sports: Mixed infections in plants. Phytopathology 2020, 110, 29–48. [Google Scholar] [CrossRef]
- Minicka, J.; Zarzyńska-Nowak, A.; Budzyńska, D.; Borodynko-Filas, N.; Hasiów-Jaroszewska, B. High-Throughput Sequencing Facilitates Discovery of New Plant Viruses in Poland. Plants 2020, 9, 820. [Google Scholar] [CrossRef]
- Singh, S.P.; Schwartz, H.F. Breeding Common Bean for Resistance to Diseases: A Review. Crop Sci. 2010, 50, 2199–2223. [Google Scholar] [CrossRef]
- Bonnet, J.; Fraile, A.; Sacristán, S.; Malpica, J.M.; García-Arenal, F. Role of recombination in the evolution of natural populations of Cucumber mosaic virus, a tripartite RNA plant virus. Virology 2005, 332, 359–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.-C.; Hsu, Y.-H.; Lin, N.-S. Satellite RNAs and Satellite Viruses of Plants. Viruses 2009, 1, 1325–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collmer, C.W.; Howell, S.H. Role of Satellite RNA in the Expression of Symptoms Caused by Plant Viruses. Annu. Rev. Phytopathol. 1992, 30, 419–442. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.; Tousignant, M. Cucumber Mosaic Virus-Associated RNA 5: I. Role of Host Plant and Helper Strain in Determining Amount of Associated RNA 5 with Virions. Virology 1977, 80, 186–195. [Google Scholar] [CrossRef]
- Kaper, J.; Collmer, C. Modulation of Viral Plant Diseases by Secondary RNA Agents. In RNA Genetics; Volume III. Variability of RNA Genomes; CRC Press Inc.: Boca Raton, FL, USA, 1988; pp. 171–194. [Google Scholar]
- Kaper, J. Rapid synthesis of double-stranded cucumber mosaic virus-associated RNA 5: Mechanism controlling viral pathogenesis? Biochem. Biophys. Res. Commun. 1982, 105, 1014–1022. [Google Scholar] [CrossRef]
- García-Arenal, F.; Palukaitis, P. Structure and Functional Relationships of Satellite RNAs of Cucumber Mosaic Virus. Protein Secret. Export. Bact. 1999, 239, 37–63. [Google Scholar] [CrossRef]
- Kouadio, K.T.; De Clerck, C.; Agneroh, T.A.; Parisi, O.; Lepoivre, P.; Jijakli, H. Role of Satellite RNAs in Cucumber Mosaic Virus-Host Plant Interactions: A Review. Biotechnol. Agron. Soc. Environ. 2013, 17, 644–650. [Google Scholar]
- Garcia-Arenal, F.; Zaitlin, M.; Palukaitis, P. Nucleotide sequence analysis of six satellite RNAs of cucumber mosaic virus: Primary sequence and secondary structure alterations do not correlate with differences in pathogenicity. Virology 1987, 158, 339–347. [Google Scholar] [CrossRef]
- Moriones, E.; Diaz, I.; Rodriguez-Cerezo, E.; Fraile, A.; Garcia-Arenal, F. Differential interactions among strains of tomato aspermy virus and satellite RNAs of cucumber mosaic virus. Virology 1992, 186, 475–480. [Google Scholar] [CrossRef]
- Fisher, J.R. Identification of Three Distinct Classes of Satellite RNAs Associated with Two Cucumber mosaic virus Serotypes from the Ornamental Groundcover Vinca minor. Plant Health Prog. 2012, 13, 18. [Google Scholar] [CrossRef] [Green Version]
- Ferreiro, C.; Ostrówka, K.; Lopez-Moya, J.J.; Díaz-Ruíz, J.R. Nucleotide sequence and symptom modulating analysis of a Peanut stunt virus-associated satellite RNA from Poland: High level of sequence identity with the American PSV satellites. Eur. J. Plant Pathol. 1996, 102, 779–786. [Google Scholar] [CrossRef]
- Militão, V.; Rodríguez-Cerezo, E.; Moreno, I.; Garcia-Arenal, F. Differential interactions among isolates of peanut stunt cucumovirus and its satellite RNA. J. Gen. Virol. 1998, 79, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Obrępalska-Stęplowska, A.; Wieczorek, P.; Budziszewska, M.; Jeszke, A.; Renaut, J. How can plant virus satellite RNAs alter the effects of plant virus infection? A study of the changes in the Nicotiana benthamiana proteome after infection by Peanut stunt virus in the presence or absence of its satellite RNA. Proteomics 2013, 13, 2162–2175. [Google Scholar] [CrossRef] [PubMed]
- Uga, H.; Tsuda, S. A One-Step Reverse Transcription-Polymerase Chain Reaction System for the Simultaneous Detection and Identification of Multiple Tospovirus Infections. Phytopathology 2005, 95, 166–171. [Google Scholar] [CrossRef]
- Van Dijk, E.L.; Jaszczyszyn, Y.; Naquin, D.; Thermes, C. The Third Revolution in Sequencing Technology. Trends Genet. 2018, 34, 666–681. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Holt, K.E. Performance of neural network basecalling tools for Oxford Nanopore sequencing. Genome Biol. 2019, 20, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Pruss, G.; Ge, X.; Shi, X.M.; Carrington, J.; Vance, V.B. Plant viral synergism: The potyviral genome encodes a broad-range pathogenicity enhancer that transactivates replication of heterologous viruses. Plant Cell 1997, 9, 859–868. [Google Scholar] [CrossRef]
- Syller, J.; Grupa, A. Antagonistic within-host interactions between plant viruses: Molecular basis and impact on viral and host fitness: Antagonistic Interactions between Plant Viruses. Mol. Plant Pathol. 2016, 17, 769–782. [Google Scholar] [CrossRef] [Green Version]
- De Coster, W.; D’Hert, S.; Schultz, D.T.; Cruts, M.; Van Broeckhoven, C. NanoPack: Visualizing and Processing Long-Read Sequencing Data. Bioinformatics 2018, 34, 2666–2669. [Google Scholar] [CrossRef]
- Tange, O. Gnu Parallel-the Command-Line Power Tool. USENIX Mag. 2011, 36, 42–47. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamberlain, S.; Szöcs, E. Taxize: Taxonomic search and retrieval in R. F1000Research 2013, 2, 191. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Programming with ggplot2. In ggplot2; Springer: Cham, Switzerland, 2016; pp. 241–253. [Google Scholar]
- Marcolungo, L.; Passera, A.; Maestri, S.; Segala, E.; Alfano, M.; Gaffuri, F.; Marturano, G.; Casati, P.; Bianco, P.A.; Delledonne, M. Real-Time On-Site Diagnosis of Quarantine Pathogens in Plant Tissues by Nanopore-Based Sequencing. Pathogens 2022, 11, 199. [Google Scholar] [CrossRef]
- Tarquini, G.; Ermacora, P.; Bianchi, G.L.; De Amicis, F.; Pagliari, L.; Martini, M.; Loschi, A.; Saldarelli, P.; Loi, N.; Musetti, R. Localization and subcellular association of Grapevine Pinot Gris Virus in grapevine leaf tissues. Protoplasma 2018, 255, 923–935. [Google Scholar] [CrossRef] [Green Version]
- Tarquini, G.; Zaina, G.; Ermacora, P.; De Amicis, F.; Franco-Orozco, B.; Loi, N.; Martini, M.; Bianchi, G.L.; Pagliari, L.; Firrao, G.; et al. Agroinoculation of Grapevine Pinot Gris Virus in tobacco and grapevine provides insights on viral pathogenesis. PLoS ONE 2019, 14, e0214010. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Family | Species | Variety | Sample ID | Site | RT-PCR | |||
|---|---|---|---|---|---|---|---|---|
| BCMV | CMV | PSV | BYMV | |||||
| Leguminosae | P. vulgaris | Unknown | LB-8 | Fonzaso | - | - | + | - |
| Calonega | LB-2 | Feltre | + | - | + | - | ||
| LB-10 | Feltre | + | - | + | - | |||
| LB-11 | Trichiana | - | - | - | - | |||
| LB-15 | Trichiana | + | - | - | - | |||
| LB-19 | Feltre (NB05) | - | - | + | - | |||
| LB-25 | Feltre | - | - | - | - | |||
| LB-30 | Sovramonte | + | - | - | - | |||
| LB-35 | Sovramonte | + | + | + | - | |||
| LB-36 | Sovramonte | + | + | - | - | |||
| Canalino | LB-1 | Feltre | + | - | - | - | ||
| LB-7 | Fonzaso | + | - | - | - | |||
| LB-18 | Feltre (NB10) | + | - | + | - | |||
| LB-24 | Fonzaso (NB12) | + | + | + | - | |||
| LB-26 | Sovramonte | + | + | + | - | |||
| LB-27 | Sovramonte | + | - | - | - | |||
| Spagnol | LB-32 | Lamon | - | + | + | - | ||
| LB-9 | Feltre | + | - | - | - | |||
| LB-34 | Sovramonte | + | + | - | - | |||
| Spagnolet | LB-3 | Feltre | + | - | - | - | ||
| LB-12 | Trichiana | - | - | + | - | |||
| LB-13 | Trichiana | + | - | - | - | |||
| LB-14 | Trichiana | + | - | - | - | |||
| LB-16 | Feltre | + | - | - | - | |||
| LB-17 | Feltre | + | - | - | - | |||
| LB-22 | Feltre (NB07) | - | - | - | - | |||
| LB-23 | Feltre | - | - | + | - | |||
| LB-33 | Feltre | + | - | + | - | |||
| LB-4 | Feltre (NB11) | + | - | - | - | |||
| LB-5 | Feltre | + | - | - | - | |||
| LB-6 | Feltre (NB08) | + | + | - | - | |||
| LB-20 | Feltre | + | - | - | - | |||
| LB-21 | Feltre (NB06) | - | - | - | - | |||
| Trifolium pratense L. | - | HO-21 | Feltre | - | + | + | - | |
| HO-25 | Feltre | - | + | + | - | |||
| HO-34 | Fonzaso | - | - | + | - | |||
| HO-35 | Fonzaso | - | - | + | - | |||
| HO-36 | Fonzaso | - | - | - | - | |||
| HO-37 | Fonzaso | - | - | - | - | |||
| HO-48 | Sovramonte | - | + | - | - | |||
| Medicago sativa L. | - | HO-24 | Feltre | - | - | - | - | |
| Amaranthaceae | Chenopodium album L. | - | HO-2 | Trichiana | - | - | - | - |
| HO-39 | Fonzaso | - | - | - | - | |||
| Amaranthus retroflexus L. | - | HO-38 | Fonzaso | - | - | - | - | |
| Achillea millefolium L. | - | HO-41 | Lamon | - | - | - | - | |
| Apiaceae | Daucus carota L. | - | HO-47 | Sovramonte | - | - | - | - |
| Solanaceae | Solanum tuberosum L. | - | HO-23 | Feltre | - | - | + | - |
| HO-46 | Lamon | - | - | + | - | |||
| Asteraceae | Taraxacum officinale L. | - | HO-12 | Feltre | - | - | - | - |
| HO-13 | Feltre | - | - | - | - | |||
| HO-14 | Feltre | - | - | - | - | |||
| HO-15 | Feltre | - | - | - | - | |||
| Asteraceae | Taraxacum officinale L. | - | HO-49 | Sovramonte | - | - | - | - |
| HO-50 | Sovramonte | - | - | - | - | |||
| Asteraceae | Weber and Chrysanthemum sp. | - | HO-22 | Feltre | - | - | - | - |
| HO-42 | Lamon | - | - | - | - | |||
| HO-43 | Lamon | - | - | - | - | |||
| HO-44 | Lamon | - | - | + | - | |||
| HO-45 | Lamon | - | - | - | - | |||
| Municipality | N° of Samples | ELISA Based Incidence (%) | |||||
|---|---|---|---|---|---|---|---|
| BCMV | CMV | PSV | Mixed Infections Rate | ||||
| BCMV and CMV | BCMV and PSV | BCMV, CMV and PSV | |||||
| Belluno | 20 | 40 | 0 | 30 | 0 | 30 | 0 |
| Feltre | 36 | 72 | 0 | 0 | 0 | 0 | 0 |
| Fonzaso | 4 | 100 | 50 | 0 | 50 | 0 | 0 |
| Lamon | 32 | 81 | 0 | 0 | 0 | 0 | 0 |
| Sovramonte | 16 | 100 | 62 | 25 | 62 | 25 | 12.5 |
| Trichiana | 4 | 100 | 0 | 0 | 0 | 0 | 0 |
| Sample ID | Variety | Sequence Name | Accession | Strain * | Sequence Similarity (%) | Pathotype or Subgroup |
|---|---|---|---|---|---|---|
| LB-1 | Canalino | BCMV_LB1 | OL874991 | NL4 | 99.0 | VII |
| LB-2 | Calonega | BCMV_LB2 | OL874987 | NL4 | 98.8 | VII |
| PSV_LB2 | OL875028 | PSV-ER | 97.8 | IA | ||
| LB-3 | Spagnolet | BCMV_LB3 | OL875007 | NL4 | 99.2 | VII |
| LB-4 | Spagnolet | BCMV_LB4 | OL874992 | NL4 | 99.0 | VII |
| LB-5 | Spagnolet | BCMV_LB5 | OL874997 | NL4 | 99.2 | VII |
| LB-6 | Spagnolet | BCMV_LB6 | OL874986 | NL4 | 98.7 | VII |
| CMV_LB6 | OL875010 | Fny-CMV | 99.2 | IA | ||
| LB-7 | Canalino | BCMV_LB7 | OL874998 | NL4 | 99.2 | VII |
| LB-9 | Spagnol | BCMV_LB9 | OL874999 | NL1 | 99.2 | I |
| LB-10 | Calonega | BCMV_LB10 | OL874988 | RU-1 | 99.2 | VI |
| PSV_LB10 | OL875020 | PSV-ER | 92.7 | IA | ||
| LB-12 | Spagnolet | PSV_LB12 | OL875026 | PSV-ER | 97.8 | IA |
| LB-13 | Spagnolet | BCMV_LB13 | OL875000 | NL4 | 99.0 | VII |
| LB-14 | Spagnolet | BCMV_LB14 | OL874995 | NL4 | 99.0 | VII |
| LB-15 | Calonega | BCMV_LB15 | OL874990 | NL4 | 99.0 | VII |
| LB-16 | Spagnolet | BCMV_LB16 | OL875001 | NL1 | 99.2 | I |
| LB-17 | Spagnolet | BCMV_LB17 | OL874994 | NL4 | 97.7 | VII |
| LB-18 | Canalino | BCMV_LB18 | OL875002 | NL4 | 98.8 | VII |
| PSV_LB18 | OL875019 | PSV-ER | 99.3 | IA | ||
| LB-19 | Calonega | PSV_LB19 | OL875023 | PSV-ER | 97.8 | IA |
| LB-20 | Spagnolet | BCMV_LB20 | OL875009 | NL1 | 99.6 | I |
| LB-23 | Spagnolet | PSV_LB23 | OL875027 | PSV-ER | 97.9 | IA |
| LB-24 | Canalino | BCMV_LB24 | OL875003 | NL4 | 98.8 | VII |
| CMV_LB24 | OL875014 | Fny-CMV | 99.3 | IA | ||
| PSV_LB24 | OL875021 | PSV-ER | 98.9 | IA | ||
| LB-26 | Canalino | BCMV_LB26 | OL874996 | RU1-CA | 95.6 | VI |
| CMV_LB26 | OL875015 | Fny-CMV | 99.3 | IA | ||
| PSV_LB26 | OL875017 | PSV-ER | 98.9 | IA | ||
| LB-27 | Canalino | BCMV_LB27 | OL875008 | NL4 | 99.2 | VII |
| LB-30 | Calonega | BCMV_LB30 | OL875004 | NL4 | 99.2 | VII |
| LB-32 | Spagnol | CMV_LB32 | OL875012 | Fny-CMV | 99.2 | IA |
| PSV_LB32 | OL875024 | PSV-ER | 98.7 | IA | ||
| LB-33 | Spagnolet | BCMV_LB33 | OL875005 | NL4 | 99.2 | VII |
| PSV_LB33 | OL875022 | PSV-ER | 98.7 | IA | ||
| LB-34 | Spagnol | BCMV_LB34 | OL874993 | NL4 | 99.2 | VII |
| CMV_LB34 | OL875011 | As-CMV | 96.6 | IB | ||
| LB-35 | Calonega | BCMV_LB35 | OL875006 | NL4 | 99.2 | VII |
| CMV_LB35 | OL875013 | As-CMV | 95.3 | IB | ||
| PSV_LB35 | OL875025 | PSV-ER | 99.2 | IA | ||
| LB-36 | Calonega | BCMV_LB36 | OL874989 | NL4 | 98.8 | VII |
| CMV_LB36 | OL875016 | As-CMV | 96.4 | IB |
| Virus | Primer Sequence (5′ → 3′) | Tm | Size | Reference |
|---|---|---|---|---|
| Bean common mosaic virus (BCMV) | For: ACCACGCTGCAGCTAAAGAGAACA Rev: AATCTAGATGATATCATACTCTCTA | 55 °C | 657 bp | Xu and Hampton (1996) |
| Cucumber mosaic virus (CMV) | For: CAGGTGGTTAACGGTACTTT Rev: CGGTAGAATCAAATTTCGGC | 60 °C | 748 bp | This study |
| Peanut stunt virus (PSV) | For: AGCCGTCGATATACCTTTTG Rev: CTCTTCACAATCACCAGGAG G | 55 °C | 1033 bp | This study |
| Bean yellow mosaic virus (BYMV) | For: CAGTTTATTATGCAGCGG Rev: GTTACCATCAATCTTCCTGCC | 55 °C | 644 bp | Uga et al., 2005 |
| Bean yellow mosaic virus (BYMV) | For: TGAAGGGCATTTTGTCAACA Rev: TTAATGAGCTTGCCGTCAAA | 55 °C | 669 bp | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarquini, G.; Martini, M.; Maestri, S.; Firrao, G.; Ermacora, P. The Virome of ‘Lamon Bean’: Application of MinION Sequencing to Investigate the Virus Population Associated with Symptomatic Beans in the Lamon Area, Italy. Plants 2022, 11, 779. https://doi.org/10.3390/plants11060779
Tarquini G, Martini M, Maestri S, Firrao G, Ermacora P. The Virome of ‘Lamon Bean’: Application of MinION Sequencing to Investigate the Virus Population Associated with Symptomatic Beans in the Lamon Area, Italy. Plants. 2022; 11(6):779. https://doi.org/10.3390/plants11060779
Chicago/Turabian StyleTarquini, Giulia, Marta Martini, Simone Maestri, Giuseppe Firrao, and Paolo Ermacora. 2022. "The Virome of ‘Lamon Bean’: Application of MinION Sequencing to Investigate the Virus Population Associated with Symptomatic Beans in the Lamon Area, Italy" Plants 11, no. 6: 779. https://doi.org/10.3390/plants11060779
APA StyleTarquini, G., Martini, M., Maestri, S., Firrao, G., & Ermacora, P. (2022). The Virome of ‘Lamon Bean’: Application of MinION Sequencing to Investigate the Virus Population Associated with Symptomatic Beans in the Lamon Area, Italy. Plants, 11(6), 779. https://doi.org/10.3390/plants11060779