
Brief Pollination Assessment of a Critically Endangered Food-Deceptive Orchid (Cypripedium guttatum) Using a Network Approach


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Flowering Phenology
2.3. Effective Pollinators
2.4. Network Sampling
2.5. Network Analysis
3. Results
3.1. Flowering Phenology
3.2. Effective Pollinators
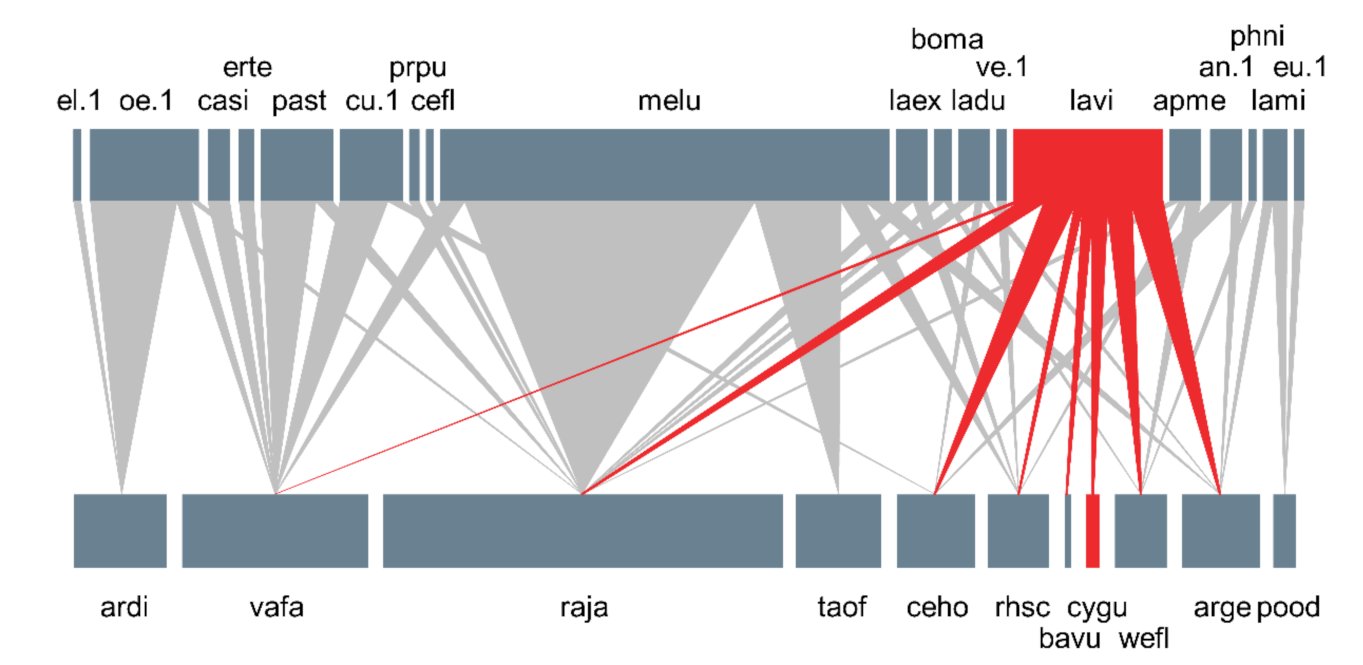
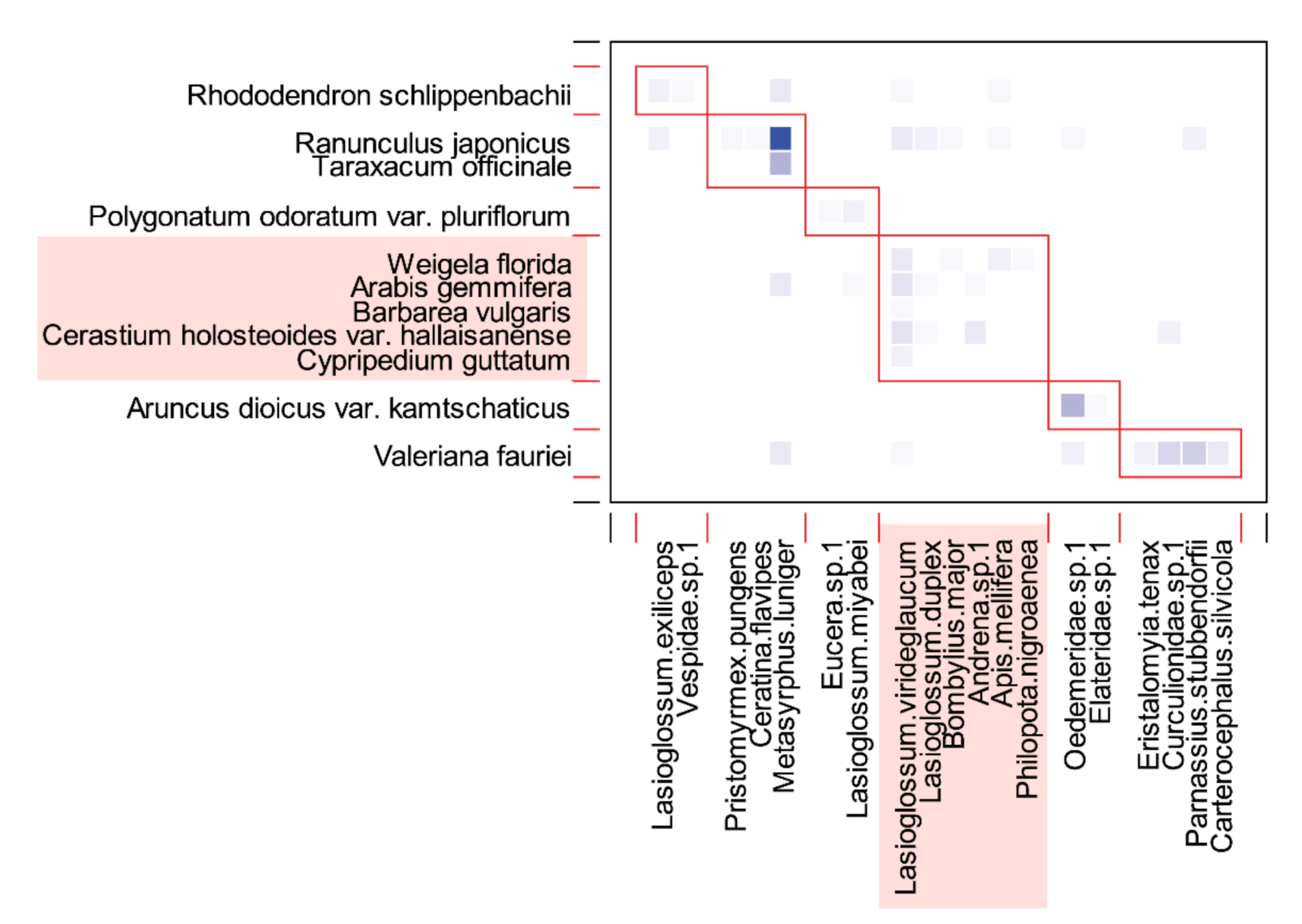
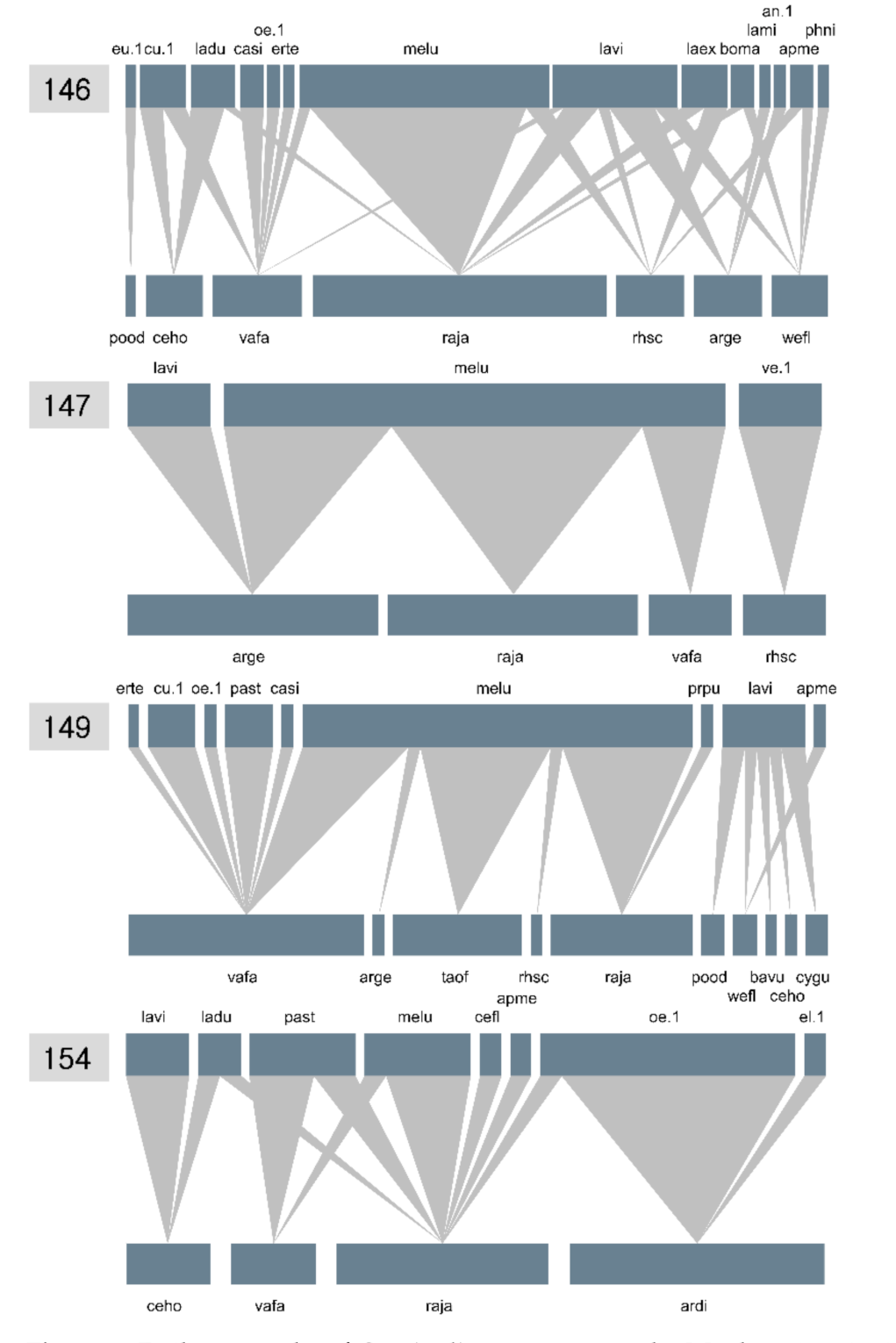
3.3. Plant–Pollinator Network
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef] [Green Version]
- IUCN. The IUCN Red List of Threatened Species. 2020. Available online: https://www.iucnredlist.org/ (accessed on 22 January 2020).
- Swarts, N.D.; Dixon, K.W. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, N.; Whitfield, J.; Pollard, G.; Bedggood, W.; Argall, M.; Dixon, K.; Davis, B.; Swarts, N. Orchid re-introductions: An evaluation of success and ecological considerations using key comparative studies from Australia. Plant Ecol. 2016, 217, 81–95. [Google Scholar] [CrossRef]
- Bleho, B.; Borkowsky, C.L.; Grantham, M.A.; Hamel, C.D. A 20 y analysis of weather and management effects on a small white Lady’s-slipper (Cypripedium candidum) population in Manitoba. Am. Midl. Nat. 2021, 185, 32–48. [Google Scholar]
- Fay, M.F. Orchid conservation: How can we meet the challenges in the twenty-first century? Bot. Stud. 2018, 59, 16. [Google Scholar] [CrossRef] [Green Version]
- Reiter, N.; Vlcek, K.; O’Brien, N.; Gibson, M.; Pitts, D.; Brown, G.R.; Bower, C.C.; Phillips, R.D. Pollinator rarity limits reintroduction sites in an endangered sexually deceptive orchid (Caladenia hastata): Implications for plants with specialized pollination systems. Bot. J. Linn. Soc. 2017, 184, 122–136. [Google Scholar] [CrossRef] [Green Version]
- Bell, S.A.J. Successful recruitment following translocation of a threatened terrestrial orchid (Diuris tricolor) into mining rehabilitation in the Hunter Valley of NSW. Ecol. Manag. Restor. 2021, 22, 204–207. [Google Scholar] [CrossRef]
- IUCN/SSC. Guidelines for Reintroductions and Other Conservation Translocations; Version 1.0; IUCN Species Survival Commission: Gland, Switzerland, 2013; p. viiii-57. [Google Scholar]
- LaBar, T.; Campbell, C.; Yang, S.; Albert, R.; Shea, K. Restoration of plant-pollinator interaction networks via species translocation. Theor. Ecol. 2014, 7, 209–220. [Google Scholar] [CrossRef]
- Fay, M.F.; Rankou, H. Slipper orchids on the IUCN Red List. In 2015 Annual Report to the Environment Agency—Abu Dhabi. Framework Support for Implementing the Strategic Plan of the IUCN Species Survival Commission; Stuart, S., Ed.; IUCN Species Survival Commission: Gland, Switzerland, 2016; pp. 106–111. Available online: http://cmsdata.iucn.org/downloads/2015_ead_report_final_web.pdf (accessed on 8 February 2022).
- Pemberton, R.W. Pollination of slipper orchids (Cypripedioideae): A review. Lankesteriana 2013, 13, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, R.L. Trends in the pollination ecology of the Orchidaceae: Evolution and systematics. Can. J. Bot. 1992, 70, 642–650. [Google Scholar] [CrossRef]
- Bernhardt, P.; Edens-Meier, R.; Westhus, E.; Vance, N. Bee-mediated pollen transfer in two populations of Cypripedium montanum Douglas ex Lindley. J. Pollinat. Ecol. 2014, 13, 188–202. [Google Scholar] [CrossRef]
- Edens-Meier, R.; Luo, Y.; Pemberton, R.; Bernhardt, P. Pollination and floral evolution of Slipper orchids (subfamily Cypripedioideae). In Darwin’s Orchids Then and Now; Edens-Meier, R., Bernhardt, P., Eds.; The University of Chicago Press: Chicago, IL, USA, 2014; pp. 265–287. [Google Scholar]
- Jersáková, J.; Johnson, S.D.; Kindlmann, P. Mechanisms and evolution of deceptive pollination in orchids. Biol. Rev. Camb. Philos. Soc. 2006, 81, 219–235. [Google Scholar] [CrossRef]
- Walsh, R.; Michaels, H.J. When it pays to cheat: Examining how generalized food deception increases male and female fitness in a terrestrial orchid. PLoS ONE 2017, 12, e0171286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, S.D.; Schiestl, F.P. Floral Mimicry; Oxford University Press: Oxford, UK, 2016; pp. 33–52. [Google Scholar]
- Aguiar, J.M.R.B.V.; Giurfa, M.; Sazima, M. A cognitive analysis of deceptive pollination: Associative mechanisms underlying pollinators’ choices in non-rewarding colour polymorphic scenarios. Sci. Rep. 2020, 10, 9476. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, L.A. Anthecological studies of the lady’s slipper Cypripedium calceolus (Orchidaceae). Bot. Not. 1979, 132, 329–347. [Google Scholar]
- Bänziger, H.; Sun, H.; Luo, Y.B. Pollination of a slippery lady slipper orchid in south-west China: Cypripedium guttatum (Orchidaceae). Bot. J. Linn. Soc. 2005, 148, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Luo, Y.B.; Bernhardt, P.; Yang, X.Q.; Kou, Y. Deceptive pollination of the Lady’s Slipper Cypripedium tibeticum (Orchidaeceae). Plant Syst. Evol. 2006, 262, 53–63. [Google Scholar] [CrossRef]
- Kull, T. Fruit-set and recruitment in populations of Cypripedium calceolus L. in Estonia. Bot. J. Linn. Soc. 2008, 126, 27–38. [Google Scholar] [CrossRef]
- Antonelli, A.; Johan Dahlberg, C.; Carlgren, K.H.; Appelqvist, T. Pollination of the Lady’s slipper orchid (Cypripedium calceolus) in Scandinavia-taxonomic and conservational aspects. Nord. J. Bot. 2009, 27, 266–273. [Google Scholar] [CrossRef]
- Suetsugu, K.; Fukushima, S. Pollination biology of the endangered orchid Cypripedium japonicum in a fragmented forest of Japan. Plant Species Biol. 2014, 29, 294–299. [Google Scholar] [CrossRef]
- Fritz, A.L.; Nilsson, L.A. How pollinator-mediated mating varies with population size in plants. Oecologia 1994, 100, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, P.; Edens-Meier, R. What we think we know vs. what we need to know about orchid pollination and conservation: Cypripedium L. as a model lineage. Bot. Rev. 2010, 76, 204–219. [Google Scholar] [CrossRef]
- Phillips, R.D.; Reiter, N.; Peakall, R. Orchid conservation: From theory to practice. Ann. Bot. 2020, 126, 345–362. [Google Scholar] [CrossRef]
- Bänziger, H.; Sun, H.Q.; Luo, Y.B. Pollination of wild lady slipper orchids Cypripedium yunnanense and C. flavum (Orchidaceae) in south-west China: Why are there no hybrids? Bot. J. Linn. Soc. 2008, 156, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Landi, P.; Minoarivelo, H.; Brännström, Ȧ.; Hui, C.; Dieckmann, U. Complexity and stability of ecological networks: A review of the theory. Popul. Ecol. 2018, 60, 319–345. [Google Scholar] [CrossRef]
- Bascompte, J.; Jordano, P. Mutualistic Networks; Princeton University Press: Princeton, NJ, USA, 2013. [Google Scholar]
- Heinen, J.H.; Rahbek, C.; Borregaard, M.K. Conservation of species interactions to achieve self-sustaining ecosystems. Ecography 2020, 43, 1603–1611. [Google Scholar] [CrossRef]
- Bascompte, J.; Jordano, P.; Melián, C.J.; Olesen, J.M. The nested assembly of plant-animal mutualistic networks. Proc. Natl. Acad. Sci. USA 2003, 100, 9383–9387. [Google Scholar] [CrossRef] [Green Version]
- Chacoff, N.P.; Resasco, J.; Vázquez, D.P. Interaction frequency, network position, and the temporal persistence of interactions in a plant-pollinator network. Ecology 2018, 99, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Martín González, A.M.; Dalsgaard, B.; Olesen, J.M. Centrality measures and importance of generalist species in pollination networks. Ecol. Complex. 2010, 7, 36–43. [Google Scholar] [CrossRef]
- Blüthgen, N.; Menzel, F.; Blüthgen, N. Measuring specialization in species interaction networks. BMC Ecol. 2006, 6, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olesen, J.M.; Bascompte, J.; Dupont, Y.L.; Jordano, P. The modularity of pollination networks. Proc. Natl. Acad. Sci. USA 2007, 104, 19891–19896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pocock, M.J.O.; Johnson, O.; Wasiuk, D. Succinctly assessing the topological importance of species in flower-pollinator networks. Ecol. Complex. 2011, 8, 265–272. [Google Scholar] [CrossRef]
- Hegland, S.J.; Dunne, J.; Nielsen, A.; Memmott, J. How to monitor ecological communities cost-efficiently: The example of plant-pollinator networks. Biol. Conserv. 2010, 143, 2092–2101. [Google Scholar] [CrossRef]
- Song, C.; Rohr, R.; Saavedra, S. Why are some plant-pollinator networks more nested than others? J. Anim. Ecol. 2017, 86, 1417–1424. [Google Scholar] [CrossRef]
- Joffard, N.; Massol, F.; Grenié, M.; Montgelard, C.; Schatz, B. Effect of pollination strategy, phylogeny and distribution on pollination niches of Euro-Mediterranean orchids. J. Ecol. 2019, 107, 478–490. [Google Scholar] [CrossRef] [Green Version]
- Argue, C.L. The Pollination Biology of North American Orchids: Volume 1, North of Florida and Mexico; Springer: New York, NY, USA, 2012. [Google Scholar]
- Gibson, R.H.; Knott, B.; Eberlein, T.; Memmott, J. Sampling method influences the structure of plant-pollinator networks. Oikos 2011, 120, 822–831. [Google Scholar] [CrossRef]
- Carvalheiro, L.G.; Barbosa, E.R.M.; Memmott, J. Pollinator network, alien species and the conservation of rare plants: Trinia glauca as a case study. J. Appl. Ecol. 2008, 45, 1419–1427. [Google Scholar] [CrossRef]
- Almeida-Neto, M.; Guimarães, P.; Guimarães, P.R., Jr.; Loyola, R.D.; Urlich, W. A consistent metric for nestedness analysis in ecological systems: Reconciling concept and measurement. Oikos 2008, 117, 1227–1239. [Google Scholar] [CrossRef]
- Dormann, C.F.; Strauss, R. A method for detecting modules in quantitative bipartite networks. Methods Ecol. Evol. 2014, 5, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Patefield, W.M. Algorithm AS159. An efficient method of generating r × c tables with given row and column totals. J. R. Stat. Soc. Ser. C Appl. Stat. 1981, 30, 91–97. [Google Scholar]
- Dormann, C.F.; Gruber, B.; Fründ, J. Introducing the bipartite package: Analysing ecological networks. R News 2008, 8, 8–11. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 2 April 2021).
- Hoeppke, C.; Simmons, B. Maxnodf: An R package for fair and fast comparisons of nestedness between networks. Methods Ecol. Evol. 2021, 12, 580–585. [Google Scholar] [CrossRef]
- Fründ, J.; McCann, K.S.; Williams, N.M. Sampling bias is a challenge for quantifying specialization and network structure: Lessons from a quantitative niche model. Oikos 2015, 125, 502–513. [Google Scholar] [CrossRef]
- Young, J.G.; Valdovinos, F.S.; Newman, M.E.J. Reconstruction of plant-pollinator networks from observational data. Nat. Commun. 2021, 12, 3911. [Google Scholar] [CrossRef] [PubMed]
- Murao, R.; Tadauchi, O.; Lee, H.S. Synopsis of Lasioglossum (Dialictus) Robertson, 1902 (Hymenoptera, Apoidea, Halictidae) in Japan, the Korean Peninsula and Taiwan. Eur. J. Taxon. 2015, 137, 1–50. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Family Name | Scientific Name |
|---|---|---|
| 1 | Liliaceae | Polygonatum odoratum var. pluriflorum |
| 2 | Ranunculaceae | Ranunculus japonicus |
| 3 | Violaceae | Viola mandshurica |
| 4 | Ericaceae | Rhododendron schlippenbachii |
| 5 | Orchidaceae | Cypripedium guttatum |
| 6 | Rosaceae | Aruncus dioicus var. kamtschaticus |
| 7 | Caprifoliaceae | Weigela florida |
| 8 | Brassicaceae | Arabis gemmifera |
| 9 | Asteraceae | Taraxacum officinale |
| 10 | Brassicaceae | Barbarea vulgaris |
| 11 | Valerianaceae | Valeriana fauriei |
| 12 | Caryophyllaceae | Cerastium holosteoides var. hallaisanensis |
| No. | Entering Time | Escaping Time | Visitation Duration (s) |
|---|---|---|---|
| 1 | 13:53:09 | 13:54:05 | 55.0 |
| 2 | 14:35:37 | 14:36:45 | 68.0 |
| 3 | 14:37:40 | 14:38:40 | 60.0 |
| 4 | 14:38:42 | 14:40:00 | 78.0 |
| 5 | 12:58:55 | 13:00:02 | 67.0 |
| 6 | 12:50:05 | 12:51:16 | 71.0 |
| 7 | 13:59:05 | 14:00:02 | 57.0 |
| 8 | 14:20:40 | 14:21:55 | 45.0 |
| Mean | 62.0 | ||
| Standard deviation | 10.4 | ||
| Date | Time of Day | ||||||
|---|---|---|---|---|---|---|---|
| 0900–1000 h | 1000–1100 h | 1100–1200 h | 1200–1300 h | 1300–1400 h | 1400–1500 h | 1500–1600 h | |
| 28 May 2018 | 0 | 2 | 2 | 2 | 1 | 3 | 0 |
| 31 May 2018 | 0 | 0 | 2 | 1 | 2 | 3 | 0 |
| 4 June 2018 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Higher Trophic Level (i.e., Pollinator) | Unweighted Degree | Betweenness Centrality | Closeness Centrality |
|---|---|---|---|
| Eucera sp. 1 | 1 | 0.000 | 0.030 |
| Pristomyrmex pungens | 1 | 0.000 | 0.055 |
| Ceratina flavipes | 1 | 0.000 | 0.055 |
| Lasioglossum exiliceps | 2 | 0.016 | 0.057 |
| Lasioglossum virideglaucum | 8 | 0.259 | 0.070 |
| Lasioglossum duplex | 3 | 0.083 | 0.062 |
| Lasioglossum miyabei | 2 | 0.153 | 0.047 |
| Eristalomyia tenax | 1 | 0.000 | 0.049 |
| Metasyrphus luniger | 5 | 0.176 | 0.068 |
| Curculionidae sp. 1 | 2 | 0.016 | 0.053 |
| Oedemeridae sp. 1 | 3 | 0.188 | 0.063 |
| Elateridae sp. 1 | 1 | 0.000 | 0.035 |
| Vespidae sp 1 | 1 | 0.000 | 0.044 |
| Parnassius stubbendorfii | 2 | 0.035 | 0.061 |
| Bombylius major | 2 | 0.024 | 0.057 |
| Carterocephalus silvicola | 1 | 0.000 | 0.049 |
| Andrena sp 1 | 2 | 0.005 | 0.047 |
| Apis mellifera | 3 | 0.044 | 0.059 |
| Philopota nigroaenea | 1 | 0.000 | 0.042 |
| Lower Trophic Level (i.e., Plant) | Unweighted Degree | Betweenness Centrality | Closeness Centrality |
|---|---|---|---|
| Polygonatum odoratum var. pluriflorum | 2 | 0.000 | 0.059 |
| Ranunculus japonicus | 10 | 0.238 | 0.106 |
| Viola mandshurica | 0 | - | - |
| Rhododendron schlippenbachii | 5 | 0.048 | 0.100 |
| Cypripedium guttatum | 1 | 0.000 | 0.095 |
| Aruncus dioicus var. kamtschaticus | 2 | 0.000 | 0.065 |
| Weigela florida | 4 | 0.000 | 0.095 |
| Arabis gemmifera | 5 | 0.476 | 0.106 |
| Taraxacum officinale | 1 | 0.000 | 0.078 |
| Barbarea vulgaris | 1 | 0.000 | 0.095 |
| Valeriana fauriei | 7 | 0.238 | 0.106 |
| Cerastium holosteoides var. hallaisanensis | 4 | 0.000 | 0.095 |
| Survey Date (Julian) | ||||
|---|---|---|---|---|
| 146 | 147 | 149 | 154 | |
| Network size (plants × animals) | 8 × 14 | 4 × 3 | 10 × 9 | 4 × 8 |
| Total number of links (binary) | 25 | 5 | 17 | 12 |
| Raw NODF | 0.239 | 0.333 | 0.216 | 0.353 |
| Max NODF | 0.598 | 0.889 | 0.469 | 0.647 |
| NODFc | 1.571 | 1.668 | 2.495 | 1.933 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Lee, H.-S.; Bae, K.-H. Brief Pollination Assessment of a Critically Endangered Food-Deceptive Orchid (Cypripedium guttatum) Using a Network Approach. Plants 2022, 11, 798. https://doi.org/10.3390/plants11060798
Lee H, Lee H-S, Bae K-H. Brief Pollination Assessment of a Critically Endangered Food-Deceptive Orchid (Cypripedium guttatum) Using a Network Approach. Plants. 2022; 11(6):798. https://doi.org/10.3390/plants11060798
Chicago/Turabian StyleLee, Hakbong, Heung-Sik Lee, and Kee-Hwa Bae. 2022. "Brief Pollination Assessment of a Critically Endangered Food-Deceptive Orchid (Cypripedium guttatum) Using a Network Approach" Plants 11, no. 6: 798. https://doi.org/10.3390/plants11060798
APA StyleLee, H., Lee, H. -S., & Bae, K. -H. (2022). Brief Pollination Assessment of a Critically Endangered Food-Deceptive Orchid (Cypripedium guttatum) Using a Network Approach. Plants, 11(6), 798. https://doi.org/10.3390/plants11060798