
Endogenous Hormone Profile and Sugars Display Differential Distribution in Leaves and Pseudobulbs of Laelia anceps Plants Induced and Non-Induced to Flowering by Exogenous Gibberellic Acid

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
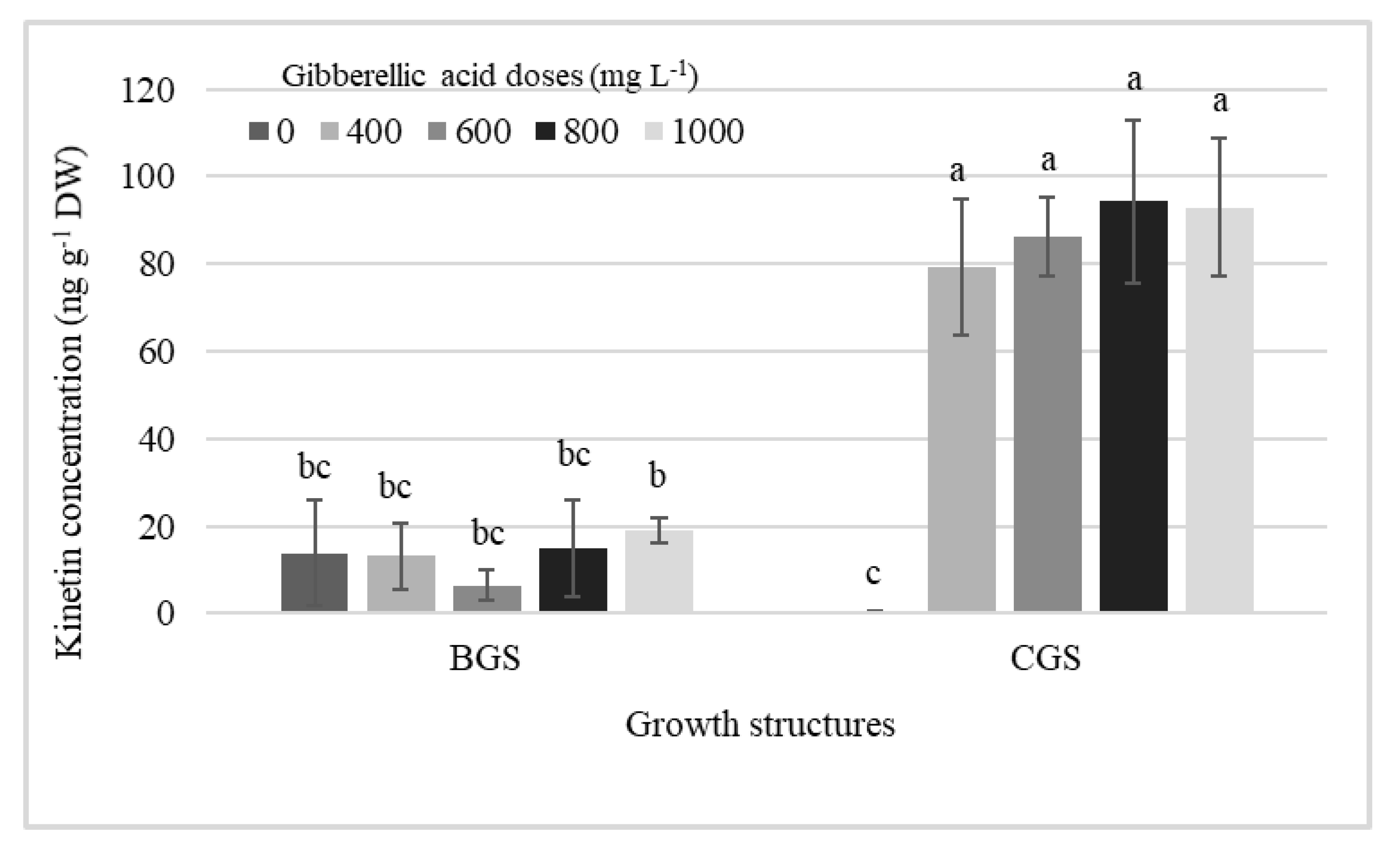
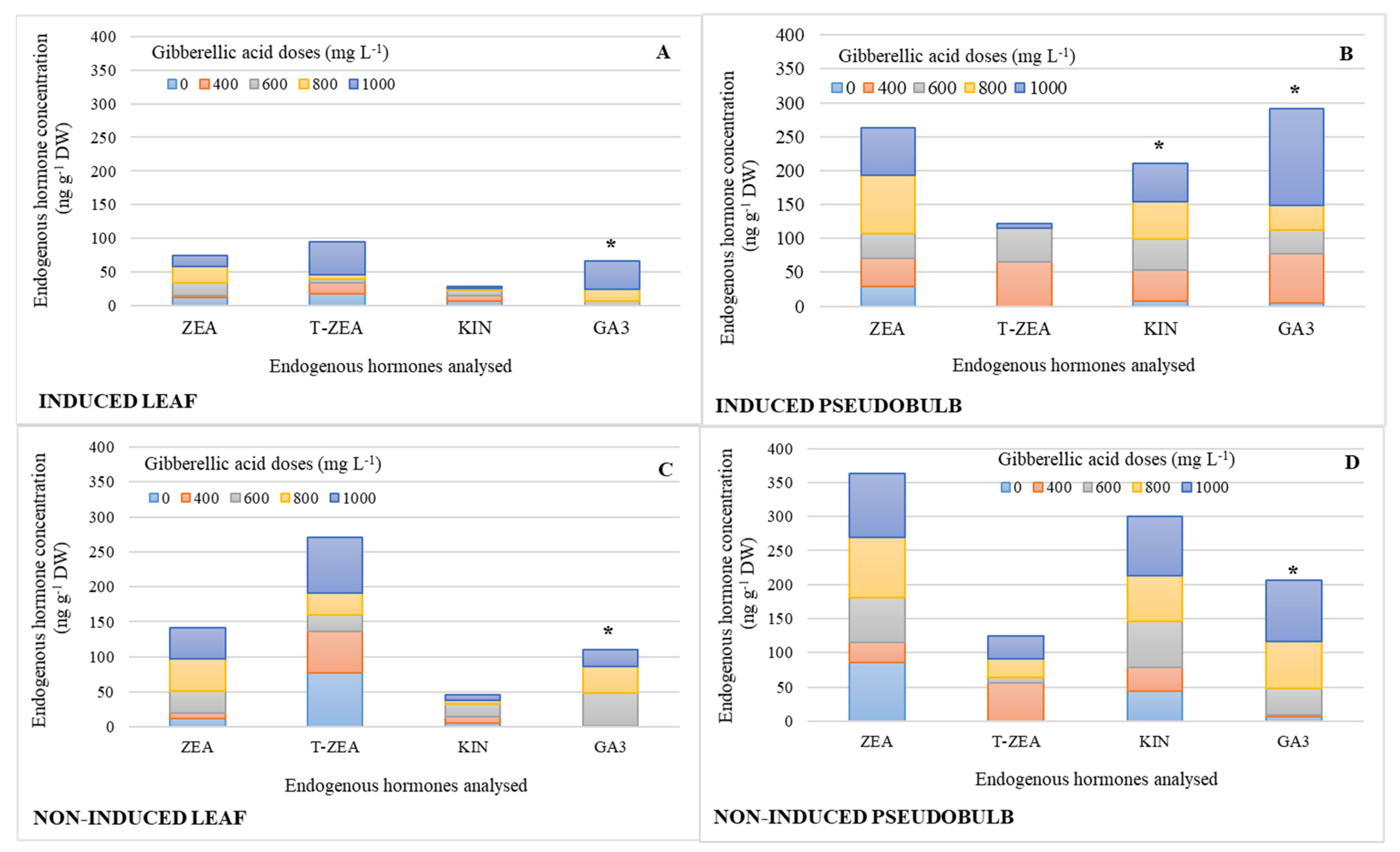
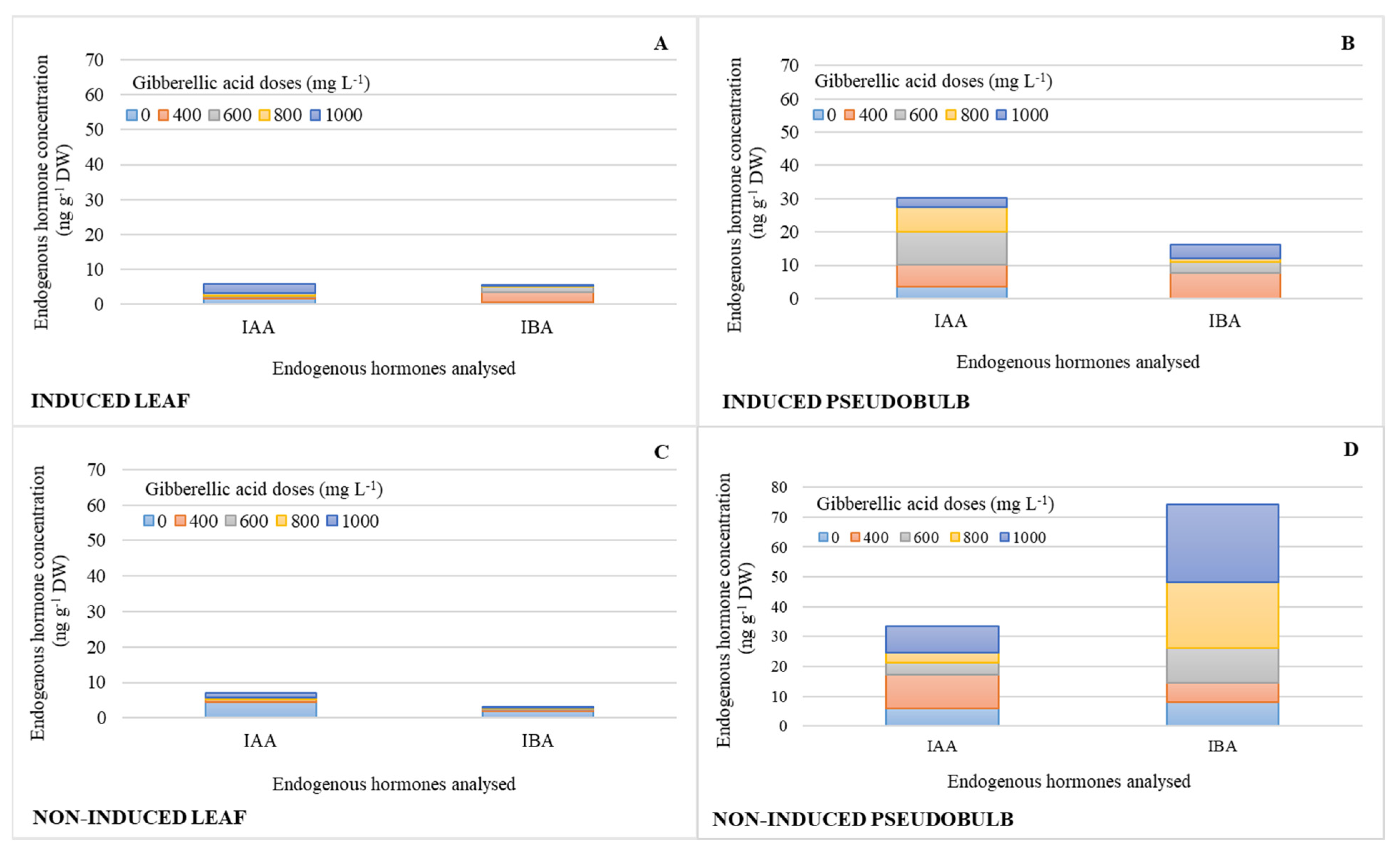
2.1. Endogenous Hormonal Concentration by Treatment Effect
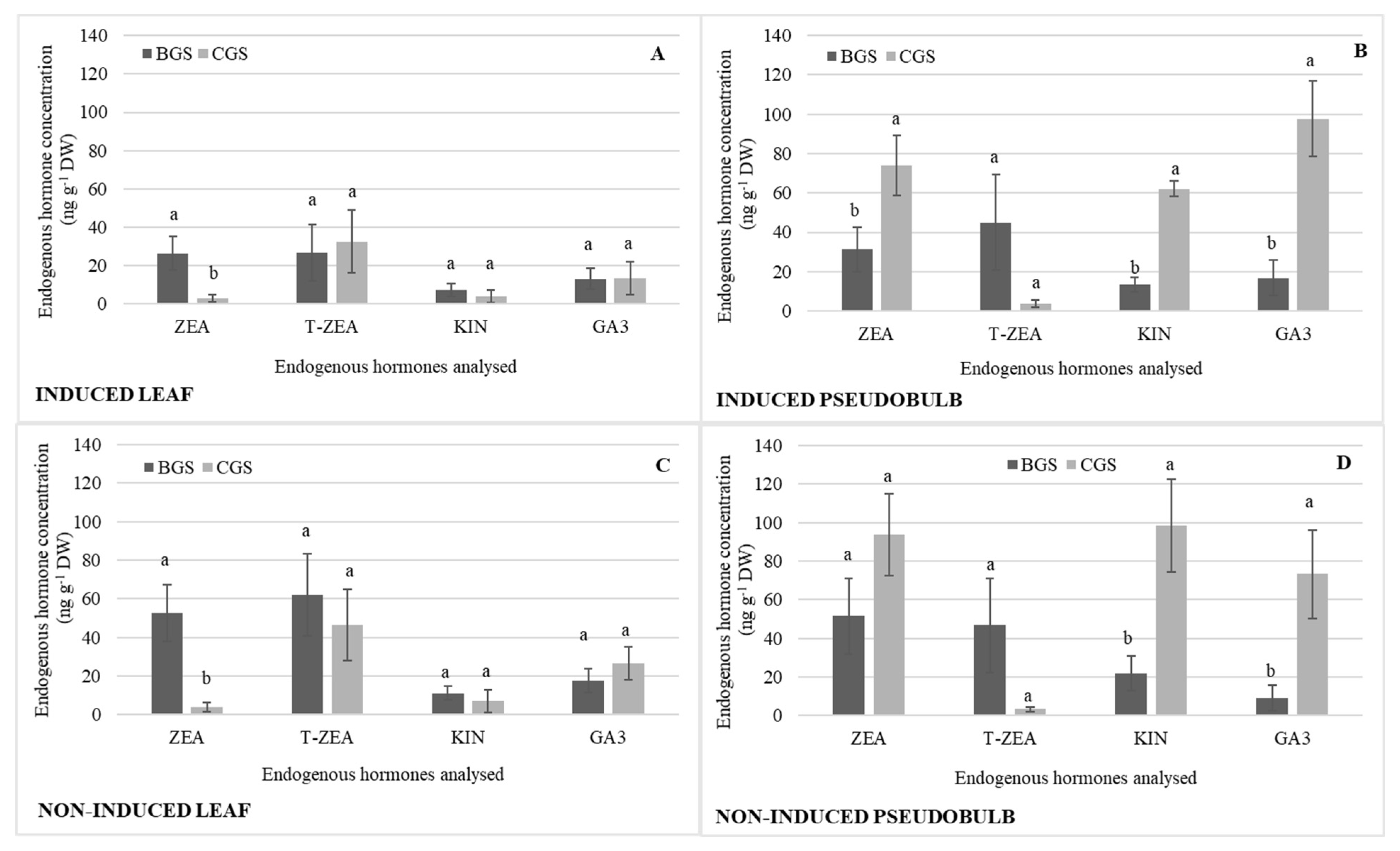
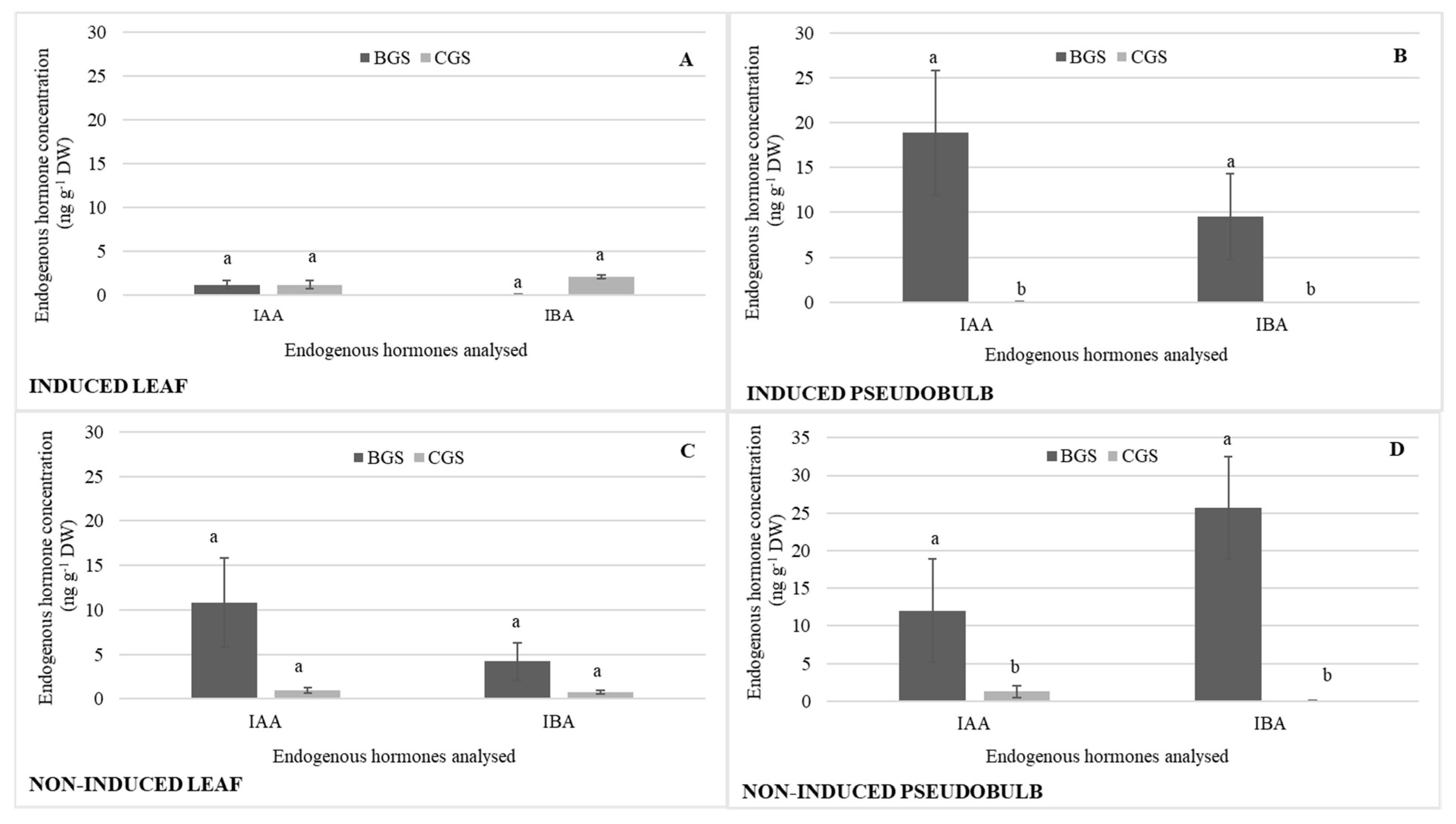
2.2. Endogenous Hormonal Concentration by Distribution in Growth Structure
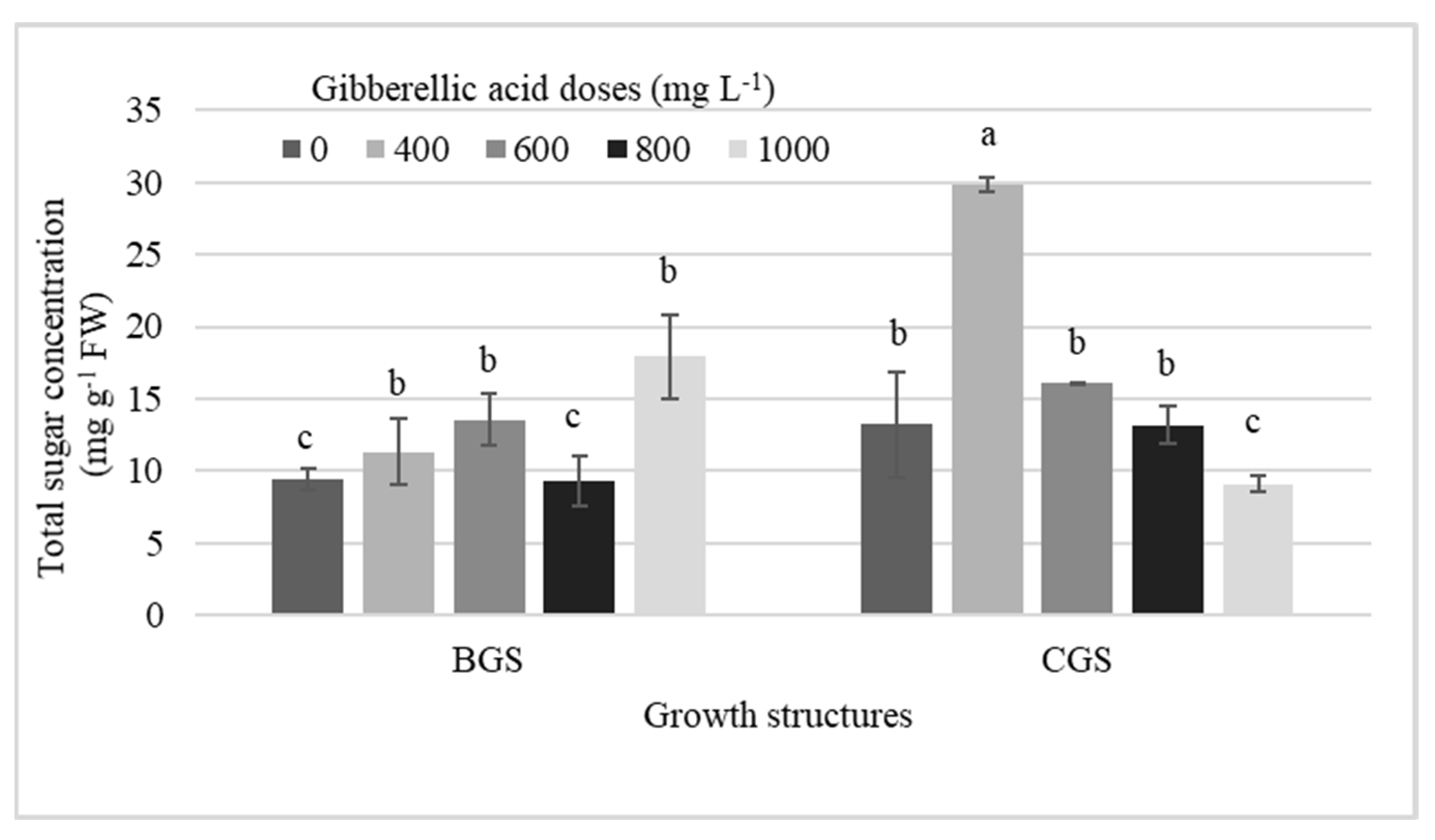
2.3. Concentration of Total Sugars
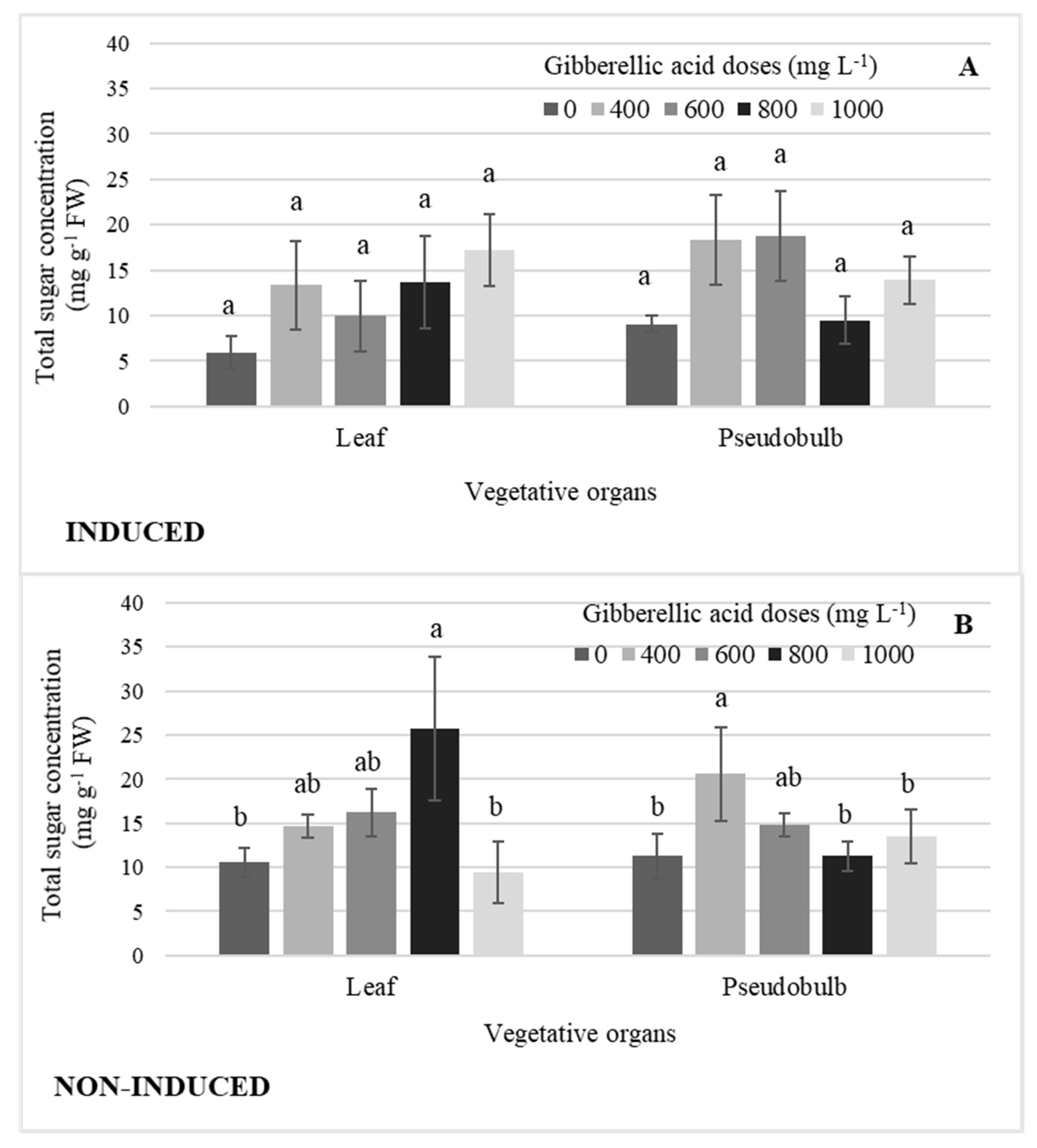
2.4. Total Sugars by Exogenous GA3 Dose Effect
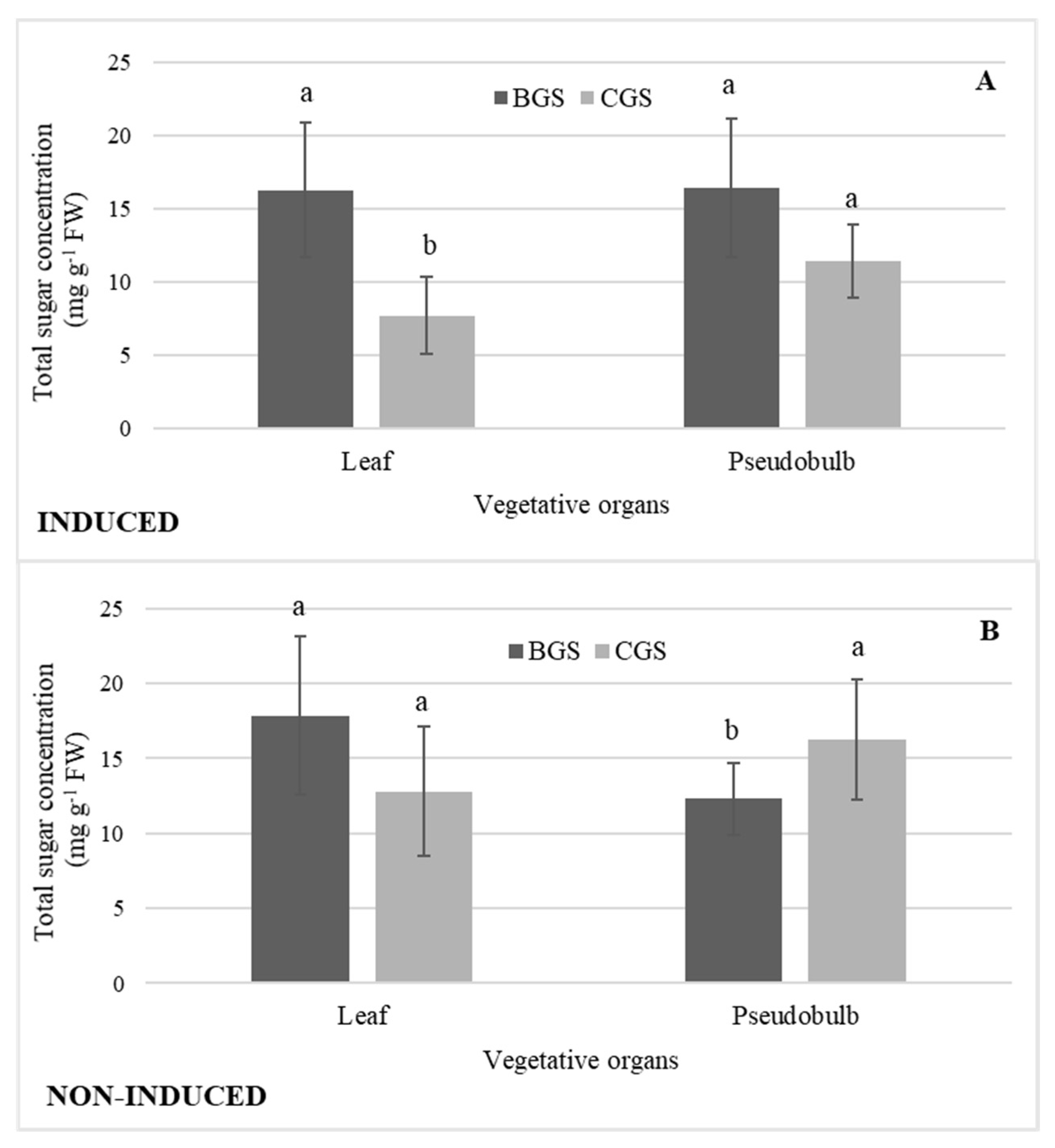
2.5. Concentration of Total Sugars by Distribution in Growth Structures
3. Discussion
3.1. Endogenous Hormone
3.2. Total Sugars
4. Materials and Methods
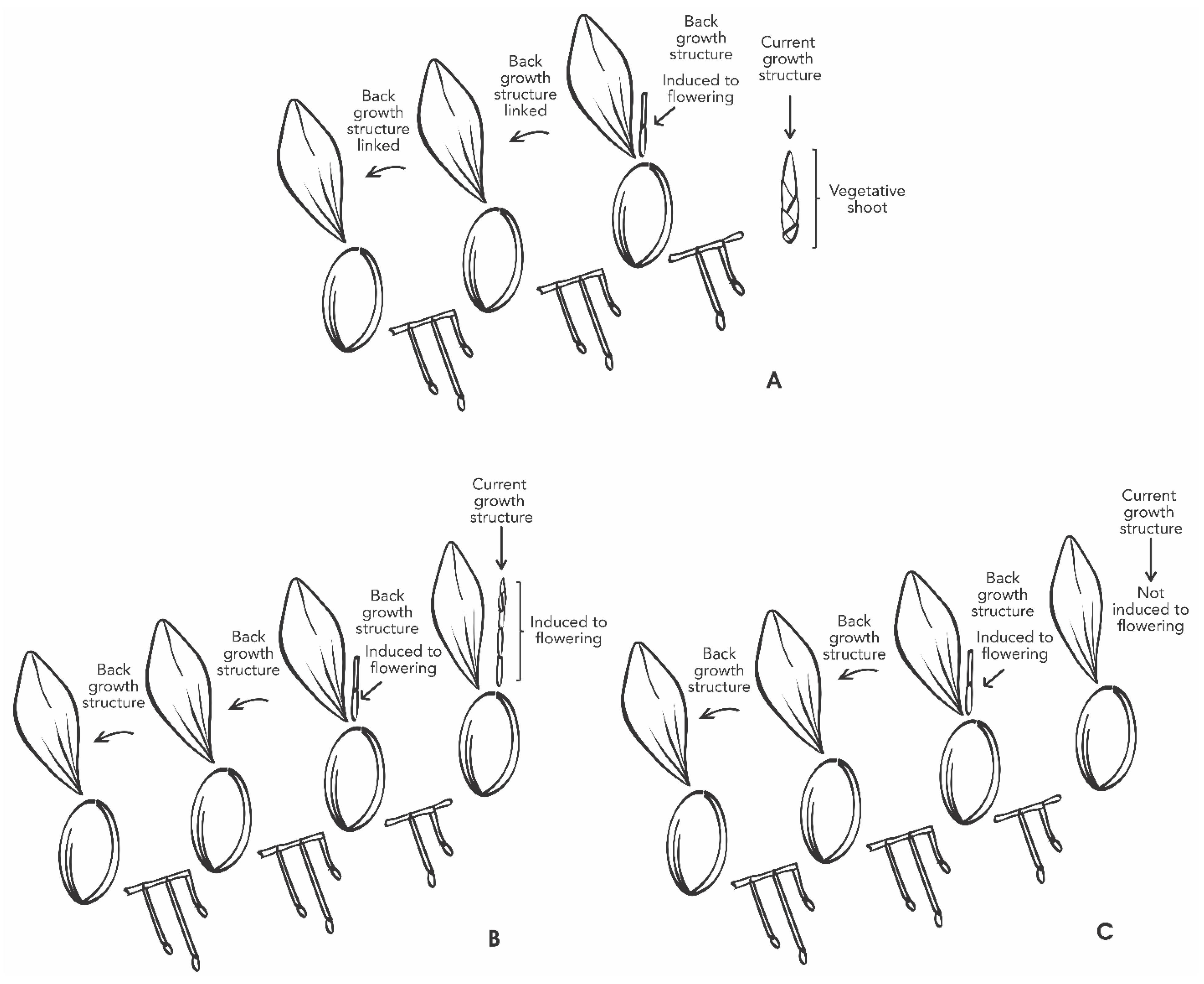
4.1. Plant Material
4.2. Inductive Treatments and Experimental Design
4.3. Growth Conditions
4.4. Data Collection
4.5. Determination of Endogenous Plant Hormones
4.6. Determination of Total Sugars
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Atwood, J.T. The size of the Orchidaceae and the systematic distribution of epiphytic orchids. Selbyana 1986, 9, 171–186. [Google Scholar]
- Dressler, R.L. How many orchid species? Selbyana 2005, 26, 155–158. [Google Scholar]
- Zhang, S.; Yang, Y.; Li, J.; Qin, J.; Zhang, W.; Huang, W.; Hu, H. Physiological diversity of orchids. Plant Divers. 2018, 40, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-L.; Viswanath, K.K.; Tong, C.-G.; An, H.R.; Jang, S.; Chen, F.-C. Floral induction and flower development of orchids. Front. Plant Sci. 2019, 10, 1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.-M.; Tong, C.-G.; Jang, S. Current progress in orchid flowering/flower development research. Plant Signal. Behav. 2017, 12, e1322245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, Y.Y.; Dean, C. The transition to flowering. Plant Cell 1998, 10, 1973–1989. [Google Scholar] [CrossRef] [PubMed]
- Eveland, A.L.; Jackson, D.P. Sugars, signalling, and plant development. J. Exp. Bot. 2012, 63, 3367–3377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciereszko, I. Regulatory roles of sugars in plant growth and development. Acta Soc. Bot. Pol. 2018, 87, 3583. [Google Scholar] [CrossRef] [Green Version]
- Corbesier, L.; Lejeune, P.; Bernier, G. The role of carbohydrates in the induction of flowering in Arabidopsis thaliana: Comparison between the wild type and a starchless mutant. Planta 1998, 206, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Parcy, F. Flowering: A time for integration. Int. J. Dev. Biol. 2004, 49, 585–593. [Google Scholar] [CrossRef] [Green Version]
- Amasino, R.M.; Michaels, S.D. The timing of flowering. Plant Physiol. 2010, 154, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Mouradov, A.; Cremer, F.; Coupland, G. Control of flowering time: Interacting pathways as a basis for diversity. Plant Cell 2002, 14, S111–S130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zhang, D.; Fan, S.; Du, L.; Shen, Y.; Xing, L.; Li, Y.; Ma, J.; Han, M. Effect of exogenous GA3 and its inhibitor paclobutrazol on floral formation, endogenous hormones, and flowering-associated genes in ‘Fuji’apple (Malus domestica Borkh.). Plant Physiol. Biochem. 2016, 107, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Hew, C.; Clifford, P. Plant growth regulators and the orchid cut-flower industry. Plant Growth Regul. 1993, 13, 231–239. [Google Scholar] [CrossRef]
- Rademacher, W. Plant growth regulators: Backgrounds and uses in plant production. J. Plant Growth Regul. 2015, 34, 845–872. [Google Scholar] [CrossRef]
- Meijón, M.; Cañal, M.J.; Fernández, H.; Rodríguez, A.; Fernández, B.; Rodríguez, R.; Feito, I. Hormonal profile in vegetative and floral buds of azalea: Levels of polyamines, gibberellins, and cytokinins. J. Plant Growth Regul. 2011, 30, 74–82. [Google Scholar] [CrossRef]
- Guan, Y.-R.; Xue, J.-Q.; Xue, Y.-Q.; Yang, R.-W.; Wang, S.-L.; Zhang, X.-X. Effect of exogenous GA3 on flowering quality, endogenous hormones, and hormone-and flowering-associated gene expression in forcing-cultured tree peony (Paeonia suffruticosa). J. Integr. Agric. 2019, 18, 1295–1311. [Google Scholar] [CrossRef]
- Su, W.-R.; Chen, W.-S.; Koshioka, M.; Mander, L.N.; Hung, L.-S.; Chen, W.-H.; Fu, Y.-M.; Huang, K.-L. Changes in gibberellin levels in the flowering shoot of Phalaenopsis hybrida under high temperature conditions when flower development is blocked. Plant Physiol. Biochem. 2001, 39, 45–50. [Google Scholar] [CrossRef]
- Peres, L.E.; Majerowicz, N.; Kerbauy, G.B. Dry matter partitioning differences between shoots and roots in two contrasting genotypes of orchids and their relationship with endogenous levels of auxins, cytokinins and abscisic acid. Rev. Bras. Fisiol. Veg. 2001, 13, 185–195. [Google Scholar] [CrossRef]
- Tejeda-Sartorius, O.; Vaquera-Huerta, H.; Trejo-Téllez, L.I.; Soto-Hernández, R.M.; Sánchez-Escudero, J. Gibberellic acid and 6-benzyladenine reduce time to flowering and improve flower quality of. Folia Hortic. 2021, 33, 121–133. [Google Scholar] [CrossRef]
- Barciszewski, J.; Rattan, S.I.; Siboska, G.; Clark, B.F. Kinetin—45 years on. Plant Sci. 1999, 148, 37–45. [Google Scholar] [CrossRef]
- Ge, L.; Yong, J.W.H.; Goh, N.K.; Chia, L.S.; Tan, S.N.; Ong, E.S. Identification of kinetin and kinetin riboside in coconut (Cocos nucifera L.) water using a combined approach of liquid chromatography–tandem mass spectrometry, high performance liquid chromatography and capillary electrophoresis. J. Chromatogr. B 2005, 829, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Moneruzzaman, K.; Hossain, A.B.; Normaniza, O.; Saifuddin, M.; Sani, W.; Nasrulhaq-Boyce, A. Effects of Removal of Young Leaves and Kinetin on Inflorescence Development and Bract Enlargement of ‘Bougainvillea glabra’ var. ‘Elizabeth Angus’. Aust. J. Crop Sci. 2010, 4, 467–473. [Google Scholar]
- Wu, P.-H.; Chang, D.C. Cytokinin treatment and flower quality in Phalaenopsis orchids: Comparing N-6-benzyladenine, kinetin and 2-isopentenyl adenine. Afr. J. Biotechnol. 2012, 11, 1592–1596. [Google Scholar]
- Singh, M.; Tiwari, N. Thidiazuron outpaces 6-benzylamino purine and kinetin in delaying flower senescence in Gladiolus grandiflora by alleviating physiological and biochemical responses. J. Appl. Biol. Biotechnol. 2021, 9, 56–62. [Google Scholar]
- Chen, W.-S.; Liao, L.-J.; Chen, C.-Y.; Huang, K.-L. Kinetin, gibberellic acid and sucrose affect vase life in Oncidium spp. Acta Bot. Gall. 2001, 148, 177–181. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, M.G.; Runkle, E.S. Benzyladenine promotes flowering in Doritaenopsis and Phalaenopsis orchids. J. Plant Growth Regul. 2008, 27, 141–150. [Google Scholar] [CrossRef]
- Wen, Z.; Guo, W.; Li, J.; Lin, H.; He, C.; Liu, Y.; Zhang, Q.; Liu, W. Comparative transcriptomic analysis of vernalization-and cytokinin-induced floral transition in Dendrobium nobile. Sci. Rep. 2017, 7, 45748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phengphachanh, B.; Naphrom, D.; Bundithya, W.; Potapohn, N. Effects of day-length and gibberellic acid (GA3) on flowering and endogenous hormone levels in Rhynchostylis gigantea (Lindl.) Ridl. J. Agric. Sci. 2012, 4, 217. [Google Scholar] [CrossRef]
- Ulger, S.; Sonmez, S.; Karkacier, M.; Ertoy, N.; Akdesir, O.; Aksu, M. Determination of endogenous hormones, sugars and mineral nutrition levels during the induction, initiation and differentiation stage and their effects on flower formation in olive. Plant Growth Regul. 2004, 42, 89–95. [Google Scholar] [CrossRef]
- Corbesier, L.; Prinsen, E.; Jacqmard, A.; Lejeune, P.; Van Onckelen, H.; Périlleux, C.; Bernier, G. Cytokinin levels in leaves, leaf exudate and shoot apical meristem of Arabidopsis thaliana during floral transition. J. Exp. Bot. 2003, 54, 2511–2517. [Google Scholar] [CrossRef]
- Pallardy, S.G. Physiology of Woody Plants; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Zhang, D.; Ren, L.; Yue, J.-h.; Wang, L.; Zhuo, L.-h.; Shen, X.-h. GA4 and IAA were involved in the morphogenesis and development of flowers in Agapanthus praecox ssp. orientalis. J. Plant Physiol. 2014, 171, 966–976. [Google Scholar] [CrossRef]
- Epstein, E.; Ludwig-Müller, J. Indole-3-butyric acid in plants: Occurrence, synthesis, metabolism and transport. Physiol. Plant. 1993, 88, 382–389. [Google Scholar] [CrossRef]
- Strader, L.C.; Wheeler, D.L.; Christensen, S.E.; Berens, J.C.; Cohen, J.D.; Rampey, R.A.; Bartel, B. Multiple facets of Arabidopsis seedling development require indole-3-butyric acid–derived auxin. Plant Cell 2011, 23, 984–999. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, A.; Liu, X.; Sun, J.; Guo, W.; Zhang, J.; Lyu, Y. Changes in the morphology of the bud meristem and the levels of endogenous hormones after low temperature treatment of different Phalaenopsis cultivars. S. Afr. J. Bot. 2019, 125, 499–504. [Google Scholar] [CrossRef]
- de Melo Ferreira, W.; Kerbauy, G.B.; Kraus, J.E.; Pescador, R.; Suzuki, R.M. Thidiazuron influences the endogenous levels of cytokinins and IAA during the flowering of isolated shoots of Dendrobium. J. Plant Physiol. 2006, 163, 1126–1134. [Google Scholar] [CrossRef]
- Novak, S.D.; Luna, L.J.; Gamage, R.N. Role of auxin in orchid development. Plant Signal. Behav. 2014, 9, e972277. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, J.K. Role of pseudobulbs in growth and flowering of Catasetum viridiflavum (Orchidaceae). Am. J. Bot. 1990, 77, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Zotz, G. What are backshoots good for? Seasonal changes in mineral, carbohydrate and water content of different organs of the epiphytic orchid, Dimerandra emarginata. Ann. Bot. 1999, 84, 791–798. [Google Scholar] [CrossRef] [Green Version]
- Ng, C.K.Y.; Hew, C.S. Orchid pseudobulbs–false’bulbs with a genuine importance in orchid growth and survival! Sci. Hortic. 2000, 83, 165–172. [Google Scholar] [CrossRef]
- Chen, J.; Zhu, F.; Liu, L.; Yi, L.; Dai, Y.; Chen, S.; He, A.; Xia, S. Integrative analyses of transcriptome and metabolome shed light on the regulation of secondary metabolites in pseudobulbs of two Bletilla striata (Thunb.) Reichb. f. varieties. J. Appl. Res. Med. Aromat. Plants 2021, 20, 100280. [Google Scholar] [CrossRef]
- Iqbal, N.; Nazar, R.; Khan, M.I.R.; Masood, A.; Khan, N.A. Role of gibberellins in regulation of source–sink relations under optimal and limiting environmental conditions. Curr. Sci. 2011, 100, 998–1007. [Google Scholar]
- Yong, J.; Hew, C. The importance of photoassimilate contribution from the current shoot and connected back shoots to inflorescence size in the thin-leaved sympodial orchid Oncidium goldiana. Int. J. Plant Sci. 1995, 156, 450–459. [Google Scholar] [CrossRef]
- Hsiao, Y.-Y.; Pan, Z.-J.; Hsu, C.-C.; Yang, Y.-P.; Hsu, Y.-C.; Chuang, Y.-C.; Shih, H.-H.; Chen, W.-H.; Tsai, W.-C.; Chen, H.-H. Research on orchid biology and biotechnology. Plant Cell Physiol. 2011, 52, 1467–1486. [Google Scholar] [CrossRef] [Green Version]
- Kozłowska, M.; Rybus-Zając, M.; Stachowiak, J.; Janowska, B. Changes in carbohydrate contents of Zantedeschia leaves under gibberellin-stimulated flowering. Acta Physiol. Plant. 2007, 29, 27–32. [Google Scholar] [CrossRef]
- Mornya, P.; Cheng, F.; Li, H. Chronological changes in plant hormone and sugar contents in cv. Ao-Shuang autumn flowering tree peony. Hortic. Sci. 2011, 38, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Welti, R.; Wang, X. Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography–mass spectrometry. Nat. Protoc. 2010, 5, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Southgate, D.A.T. Determination of Food Carbohydrates; Applied Science Publishers: London, UK, 1976. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Induced Leaf | Induced Pseudobulb | |||
|---|---|---|---|---|---|
| ZEA | ZEA | KIN | IBA | GA3 | |
| GA3 | 0.7217 | 0.3456 | 0.0164 * | 0.3643 | 0.0456 * |
| GS | 0.0260 * | 0.0443 * | <0.0001 ** | 0.0145 * | 0.0097 ** |
| GA3xGS | 0.5921 | 0.6476 | 0.0171 * | 0.3643 | 0.3012 |
| Factors | Non-Induced Leaf | Non-Induced Pseudobulb | |||
| ZEA | KIN | IAA | IBA | GA3 | |
| GA3 | 0.4072 | 0.6835 | 0.6969 | 0.5244 | 0.0209 * |
| GS | 0.0049 ** | 0.0059 ** | 0.0157 * | 0.0238 * | 0.0017 ** |
| GA3xGS | 0.5475 | 0.5809 | 0.9663 | 0.5210 | 0.0593 |
| Induced Leaf | Non-Induced Leaf | Induced Pseudobulb | Non-Induced Pseudobulb | ||
|---|---|---|---|---|---|
| GA3 DOSES (mg L−1) | GA | GA | KIN | GA | GA |
| 0 | 0.0 b | 0 b | 7.0 b | 5.5 b | 6.0 b |
| 400 | 0.0 b | 1.8 b | 46.3 ab | 71.8 ab | 2.7 b |
| 600 | 7.3 b | 47.1 a | 46.4 ab | 35.5 ab | 39.1 ab |
| 800 | 17.6 ab | 37.0 ab | 54.6 a | 36.2 ab | 68.7 ab |
| 1000 | 41.9 a | 24.6 ab | 55.9 a | 143.0 a | 89.3 a |
| Factors | Induced | Non-Induced | ||
|---|---|---|---|---|
| Leaf | Pseudobulb | Leaf | Pseudobulb | |
| GA3 | 0.1120 | 0.0734 | 0.0301 * | 0.0021 ** |
| GS | 0.0037 ** | 0.0726 | 0.1250 | 0.0093 ** |
| GA3xGS | 0.7213 | 0.9021 | 0.8213 | 0.0001 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tejeda-Sartorius, O.; Soto-Hernández, R.M.; San Miguel-Chávez, R.; Trejo-Téllez, L.I.; Caamal-Velázquez, H. Endogenous Hormone Profile and Sugars Display Differential Distribution in Leaves and Pseudobulbs of Laelia anceps Plants Induced and Non-Induced to Flowering by Exogenous Gibberellic Acid. Plants 2022, 11, 845. https://doi.org/10.3390/plants11070845
Tejeda-Sartorius O, Soto-Hernández RM, San Miguel-Chávez R, Trejo-Téllez LI, Caamal-Velázquez H. Endogenous Hormone Profile and Sugars Display Differential Distribution in Leaves and Pseudobulbs of Laelia anceps Plants Induced and Non-Induced to Flowering by Exogenous Gibberellic Acid. Plants. 2022; 11(7):845. https://doi.org/10.3390/plants11070845
Chicago/Turabian StyleTejeda-Sartorius, Olga, Ramón Marcos Soto-Hernández, Rubén San Miguel-Chávez, Libia Iris Trejo-Téllez, and Humberto Caamal-Velázquez. 2022. "Endogenous Hormone Profile and Sugars Display Differential Distribution in Leaves and Pseudobulbs of Laelia anceps Plants Induced and Non-Induced to Flowering by Exogenous Gibberellic Acid" Plants 11, no. 7: 845. https://doi.org/10.3390/plants11070845
APA StyleTejeda-Sartorius, O., Soto-Hernández, R. M., San Miguel-Chávez, R., Trejo-Téllez, L. I., & Caamal-Velázquez, H. (2022). Endogenous Hormone Profile and Sugars Display Differential Distribution in Leaves and Pseudobulbs of Laelia anceps Plants Induced and Non-Induced to Flowering by Exogenous Gibberellic Acid. Plants, 11(7), 845. https://doi.org/10.3390/plants11070845