
Biochemistry and Molecular Basis of Intracellular Flavonoid Transport in Plants



Abstract
:1. Introduction
1.1. Biological Relevance of Specialized Metabolites
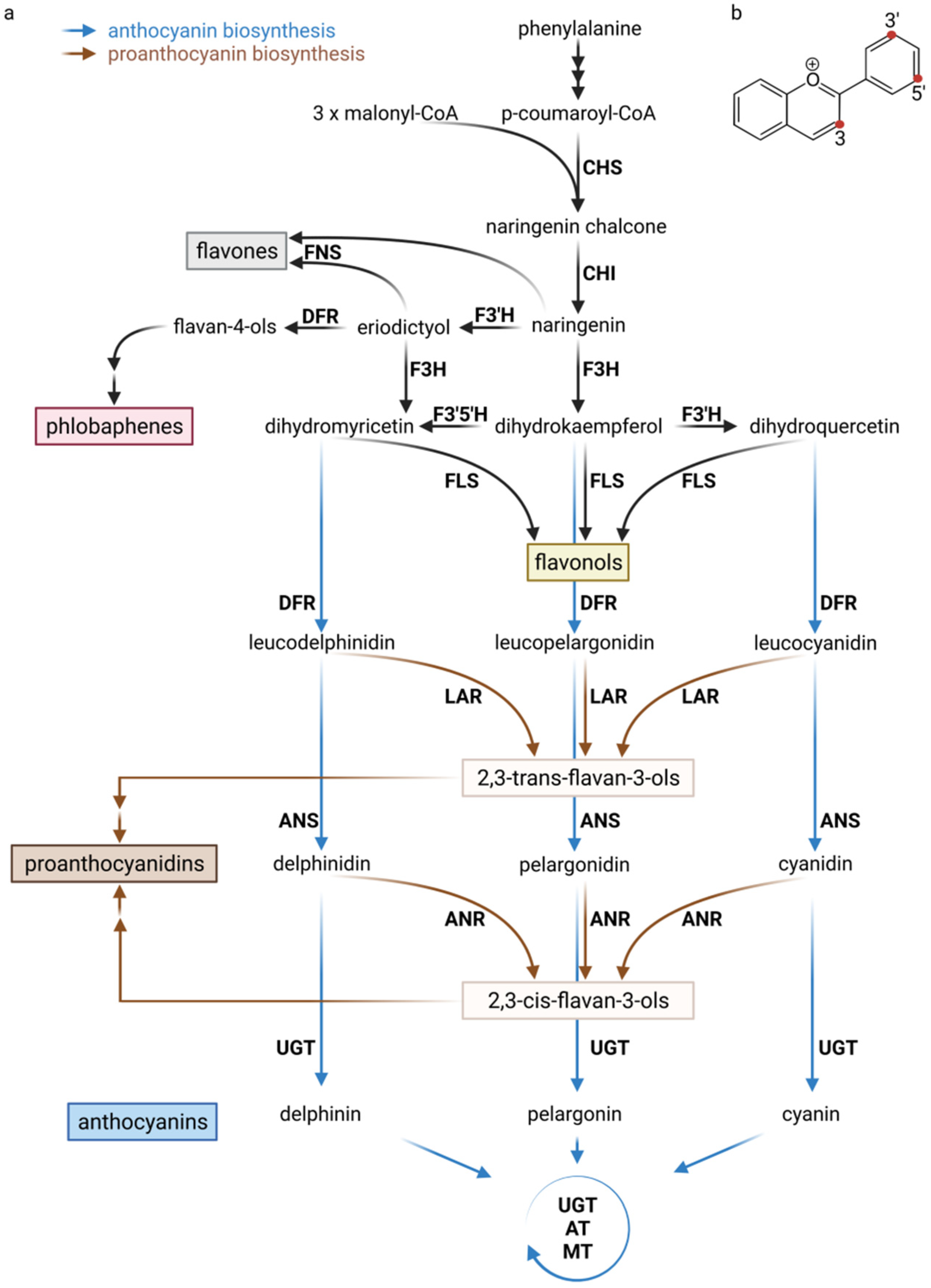
1.2. Biosynthesis of Flavonoids
1.3. Transport and Subcellular Localizaiton of Flavonoids
1.4. Membrane Permeability of Specialized Metabolites
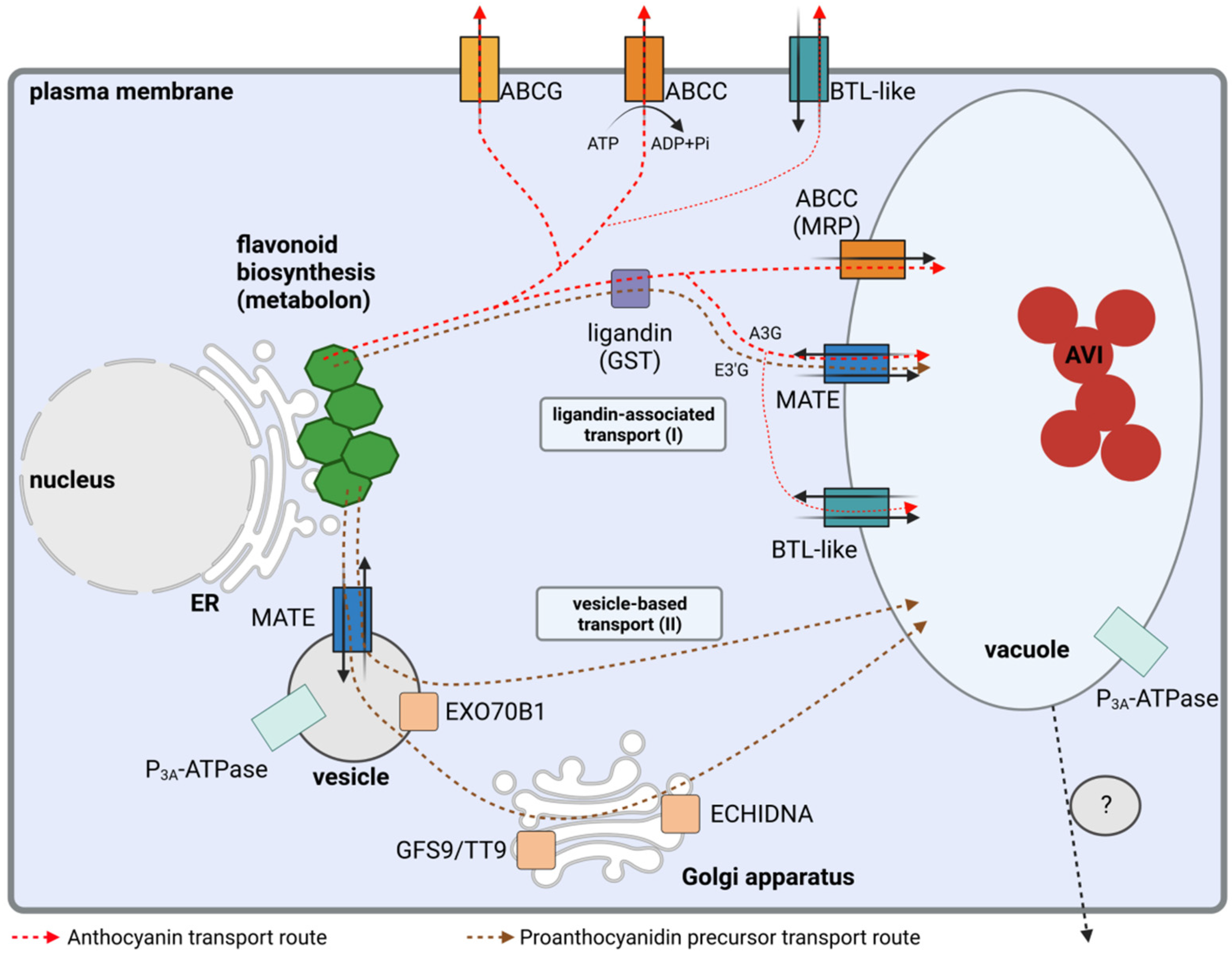
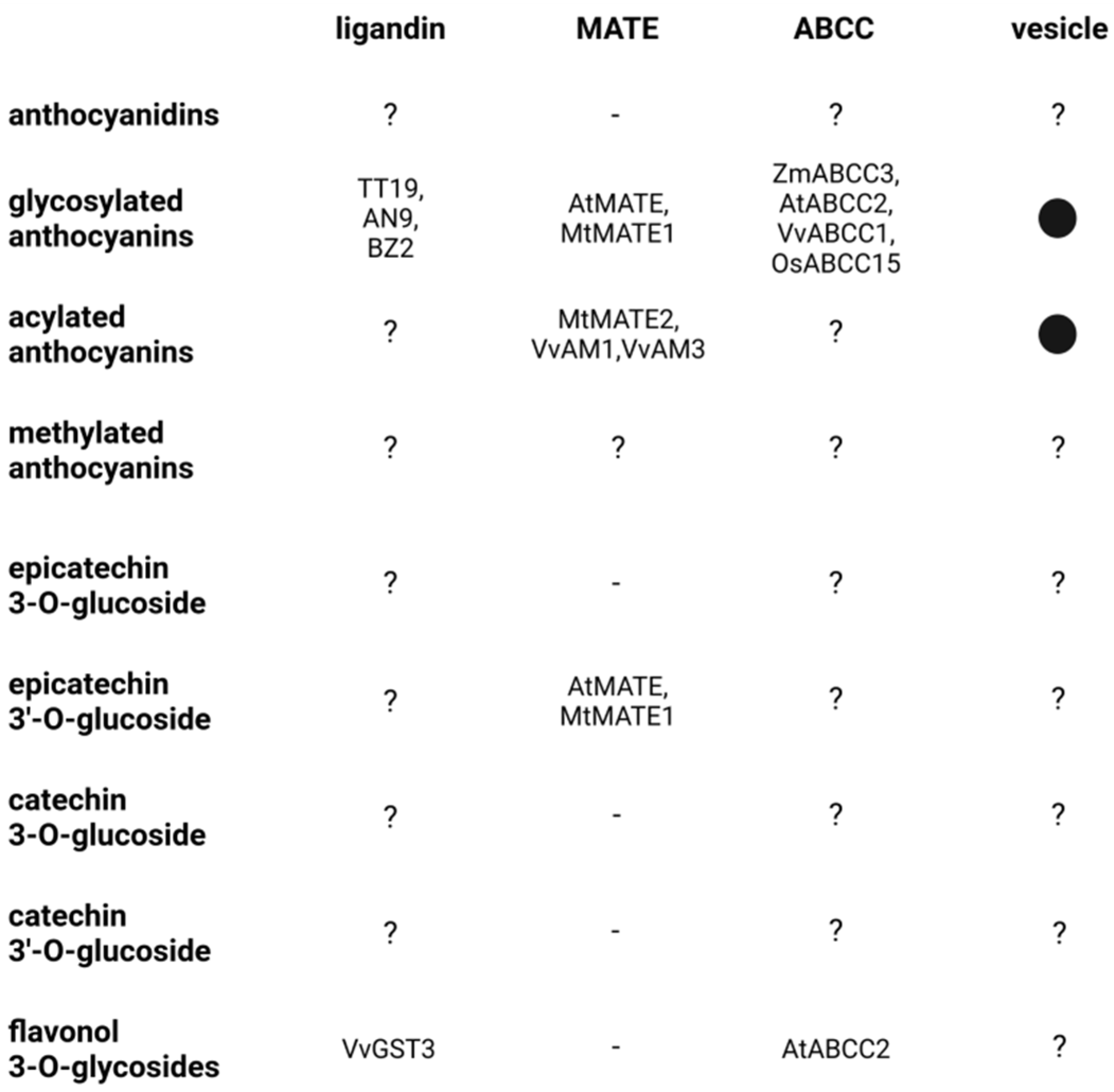
2. Ligandin and Transporter-Associated Sequestration of Flavonoids into the Vacuole

{kind=link}
{kind=link}
{kind=link}
| Function | Gene | Species | Reference |
|---|---|---|---|
| Ligandin (GST) | AN9 | Petunia hybrid (petunia) | [97] |
| TT19 | Arabidopsis thaliana | [98] | |
| BZ2 | Zea mays (maize) | [96] | |
| VvGST1, VvGST4 | Vitis vinifera (grape vine) | [100,108] | |
| PstGST1 | Perilla frutescens (deulkkae) | [99] | |
| PpGST1/Riant | Prunus persica (peach) | [116] | |
| BnGSTF6, BnGSTF12 | Brassica napus (rapeseed) | [117] | |
| AcGST1 | Actinidia chinensis (kiwifruit) | [107] | |
| BRACT1 | Euphorbia pulcherima (poinsettia) | [118] | |
| RsGST1 | Raphanus sativus (radish) | [119] | |
| RAP | Fragaria vesca (strawberry) | [103] | |
| MdGSTF6 | Malus domestica (apple) | [104] | |
| LcGST4 | Litchi chinensis (lychee) | [120] | |
| IbGSTF4 | Ipomoea batatas (sweet potato) | [106] | |
| CkmGST3 | Cyclamen spec. | [121] | |
| FL3/DcGSTF2 | Dianthus caryophyllus (carnation) | [122] | |
| PcGST1 | Petroselium crispum (parsley) | [123] | |
| CmGST1 | Chrysanthemum spec. | [124] | |
| CsGSTF1 | Camelia sinensis (tea) | [125] | |
| DcGST1 | Daucus carota (carrot) | [126,127] | |
| GmGST26A/GmHsp26A | Glycine max (soybean) | [97] | |
| MATE | TT12, FFT | Arabidopsis thaliana | [113,128] |
| MtMATE1, MtMATE2 | Medicago truncatula (barrelclover) | [52,129] | |
| VvAM1, VvAM3 | Vitis vinifera (grape vine) | [130,131,132] | |
| LhDTX35 | Lilium spp. | [133] | |
| MdMATE1, MdMATE2 | Malus domestica (apple) | [134] | |
| BnTT12 | Brassica napus (rapeseed) | [135] | |
| RsMATE9 | Raphanus sativus (radish) | [136] | |
| SlMTP77 | Solanum lycopersicum (tomato) | [137] | |
| VcMATE, 2, 3, 5, 7, 8, 9 | Vaccinium corymbosum (blueberry) | [138] | |
| GmMATE1 | Glycine max (soybean) | [139] | |
| FaTT12-1 | Fragaria vesca (strawberry) | [140] | |
| GhTT12 | Gossypium hirsutum (cotton) | [141] | |
| DcMATE1 | Daucus carota (carrot) | [127] | |
| DkMATE1 | Diospyros kaki (kaki persimmon) | [142] | |
| ABCC (MRP) | ZmMRP3(ZmABCC3), ZmMRP4(ZmABCC4) | Zea mays (maize) | [143] |
| AtABCC2 | Arabidopsis thaliana | [144] | |
| VvABCC1 | Vitis vinifera (grape vine) | [145] | |
| OsMRP15 | Oryza sativa (rice) | [146] | |
| RsABC | Raphanus sativus (radish) | [147] | |
| P3A-ATPase | AHA10/TT13 | Arabidopsis thaliana | [148,149] |
| PH5 | Petunia hybrid (petunia) | [150] | |
| GmPH5 | Glycine max (soybean) | [151] | |
| H+-PPase | VHP1 | Arabidopsis thaliana | [152] |
| BTL-like | -1 | Vitis vinifera (grape vine) | [115] |
| -1 | Dianthus caryophyllus (carnation) | [114] | |
| Vesicle trafficking | GFS9/TT9 | Arabidopsis thaliana | [153] |
| ECHIDNA | Arabidopsis thaliana | [154] | |
| EXO70B1 | Arabidopsis thaliana | [155] |
2.1. Anthocyanin Transport
2.2. Proanthocyanin Transport
2.3. Transport of Other Flavonoids
3. Flavonoid Transport in Vesicles
3.1. Anthocyanin Transport
3.2. PA Transport
3.3. Flavonol Transport
4. Secretion of Flavonoids and Long Distance Transport
5. Conclusions and Open Questions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weng, J.-K. The Evolutionary Paths towards Complexity: A Metabolic Perspective. New Phytol. 2014, 201, 1141–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, C.; Fernie, A.R.; Luo, J. Exploring the Diversity of Plant Metabolism. Trends Plant Sci. 2019, 24, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Afendi, F.M.; Okada, T.; Yamazaki, M.; Hirai-Morita, A.; Nakamura, Y.; Nakamura, K.; Ikeda, S.; Takahashi, H.; Altaf-Ul-Amin, M.; Darusman, L.K.; et al. KNApSAcK Family Databases: Integrated Metabolite-Plant Species Databases for Multifaceted Plant Research. Plant Cell Physiol. 2012, 53, e1. [Google Scholar] [CrossRef] [Green Version]
- Fernie, A.R.; Trethewey, R.N.; Krotzky, A.J.; Willmitzer, L. Metabolite Profiling: From Diagnostics to Systems Biology. Nat. Rev. Mol. Cell Biol. 2004, 5, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. Biosynthesis of Flavonoids and Effects of Stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Jaakola, L.; Määttä-Riihinen, K.; Kärenlampi, S.; Hohtola, A. Activation of Flavonoid Biosynthesis by Solar Radiation in Bilberry (Vaccinium Myrtillus L.) Leaves. Planta 2004, 218, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Fini, A.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Tattini, M. Stress-Induced Flavonoid Biosynthesis and the Antioxidant Machinery of Plants. Plant Signal. Behav. 2011, 6, 709–711. [Google Scholar] [CrossRef]
- Petrussa, E.; Braidot, E.; Zancani, M.; Peresson, C.; Bertolini, A.; Patui, S.; Vianello, A. Plant Flavonoids—Biosynthesis, Transport and Involvement in Stress Responses. Int. J. Mol. Sci. 2013, 14, 14950–14973. [Google Scholar] [CrossRef]
- Albert, N.W.; Thrimawithana, A.H.; McGhie, T.K.; Clayton, W.A.; Deroles, S.C.; Schwinn, K.E.; Bowman, J.L.; Jordan, B.R.; Davies, K.M. Genetic Analysis of the Liverwort Marchantia Polymorpha Reveals That R2R3MYB Activation of Flavonoid Production in Response to Abiotic Stress Is an Ancient Character in Land Plants. New Phytol. 2018, 218, 554–566. [Google Scholar] [CrossRef] [Green Version]
- Bian, X.-H.; Li, W.; Niu, C.-F.; Wei, W.; Hu, Y.; Han, J.-Q.; Lu, X.; Tao, J.-J.; Jin, M.; Qin, H.; et al. A Class B Heat Shock Factor Selected for during Soybean Domestication Contributes to Salt Tolerance by Promoting Flavonoid Biosynthesis. New Phytol. 2020, 225, 268–283. [Google Scholar] [CrossRef] [Green Version]
- Liang, T.; Shi, C.; Peng, Y.; Tan, H.; Xin, P.; Yang, Y.; Wang, F.; Li, X.; Chu, J.; Huang, J.; et al. Brassinosteroid-Activated BRI1-EMS-SUPPRESSOR 1 Inhibits Flavonoid Biosynthesis and Coordinates Growth and UV-B Stress Responses in Plants. Plant Cell 2020, 32, 3224–3239. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grotewold, E. The Genetics and Biochemistry of Floral Pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef] [PubMed]
- Tohge, T.; de Souza, L.P.; Fernie, A.R. Current Understanding of the Pathways of Flavonoid Biosynthesis in Model and Crop Plants. J. Exp. Bot. 2017, 68, 4013–4028. [Google Scholar] [CrossRef] [PubMed]
- Pucker, B.; Reiher, F.; Schilbert, H.M. Automatic Identification of Players in the Flavonoid Biosynthesis with Application on the Biomedicinal Plant Croton Tiglium. Plants 2020, 9, 1103. [Google Scholar] [CrossRef]
- Kovinich, N.; Kayanja, G.; Chanoca, A.; Riedl, K.; Otegui, M.S.; Grotewold, E. Not All Anthocyanins Are Born Equal: Distinct Patterns Induced by Stress in Arabidopsis. Planta 2014, 240, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Landi, M.; Tattini, M.; Gould, K.S. Multiple Functional Roles of Anthocyanins in Plant-Environment Interactions. Environ. Exp. Bot. 2015, 119, 4–17. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Higashi, Y.; Nakabayashi, R. The Origin and Evolution of Plant Flavonoid Metabolism. Front. Plant Sci. 2019, 10, 943. [Google Scholar] [CrossRef] [Green Version]
- Pollastri, S.; Tattini, M. Flavonols: Old Compounds for Old Roles. Ann. Bot. 2011, 108, 1225–1233. [Google Scholar] [CrossRef] [Green Version]
- Emiliani, J.; Grotewold, E.; Falcone Ferreyra, M.L.; Casati, P. Flavonols Protect Arabidopsis Plants against UV-B Deleterious Effects. Mol. Plant 2013, 6, 1376–1379. [Google Scholar] [CrossRef] [Green Version]
- Lepiniec, L.; Debeaujon, I.; Routaboul, J.-M.; Baudry, A.; Pourcel, L.; Nesi, N.; Caboche, M. Genetics and Biochemistry of Seed Flavonoids. Annu. Rev. Plant Biol. 2006, 57, 405–430. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Sarnala, S. Proanthocyanidin Biosynthesis—A Matter of Protection. Plant Physiol. 2020, 184, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, J.-L.; Jez, J.M.; Bowman, M.E.; Dixon, R.A.; Noel, J.P. Structure of Chalcone Synthase and the Molecular Basis of Plant Polyketide Biosynthesis. Nat. Struct. Mol. Biol. 1999, 6, 775–784. [Google Scholar] [CrossRef]
- Jez, J.M.; Bowman, M.E.; Dixon, R.A.; Noel, J.P. Structure and Mechanism of the Evolutionarily Unique Plant Enzyme Chalcone Isomerase. Nat. Struct. Mol. Biol. 2000, 7, 786–791. [Google Scholar] [CrossRef]
- Forkmann, G.; Heller, W.; Grisebach, H. Anthocyanin Biosynthesis in Flowers of Matthiola Incana Flavanone 3-and Flavonoid 3′-Hydroxylases. Z. Nat. C 1980, 35, 691–695. [Google Scholar] [CrossRef]
- Martens, S.; Forkmann, G. Cloning and Expression of Flavone Synthase II from Gerbera Hybrids. Plant J. 1999, 20, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Martens, S.; Forkmann, G.; Britsch, L.; Wellmann, F.; Matern, U.; Lukačin, R. Divergent Evolution of Flavonoid 2-Oxoglutarate-Dependent Dioxygenases in Parsley 1. FEBS Lett. 2003, 544, 93–98. [Google Scholar] [CrossRef] [Green Version]
- de Vetten, N.; ter Horst, J.; van Schaik, H.-P.; de Boer, A.; Mol, J.; Koes, R. A Cytochrome B5 Is Required for Full Activity of Flavonoid 3′,5′-Hydroxylase, a Cytochrome P450 Involved in the Formation of Blue Flower Colors. Proc. Natl. Acad. Sci. USA 1999, 96, 778–783. [Google Scholar] [CrossRef] [Green Version]
- Brugliera, F.; Barri-Rewell, G.; Holton, T.A.; Mason, J.G. Isolation and Characterization of a Flavonoid 3′-Hydroxylase CDNA Clone Corresponding to the Ht1 Locus of Petunia Hybrida. Plant J. 2002, 19, 441–451. [Google Scholar] [CrossRef]
- Holton, T.A.; Brugliera, F.; Tanaka, Y. Cloning and Expression of Flavonol Synthase from Petunia Hybrida. Plant J. 1993, 4, 1003–1010. [Google Scholar] [CrossRef]
- Shirley, B.W.; Hanley, S.; Goodman, H.M. Effects of Ionizing Radiation on a Plant Genome: Analysis of Two Arabidopsis Transparent Testa Mutations. Plant Cell 1992, 4, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, M.K.; Murrell, J.R.; Shirley, B.W. Characterization of Flavonol Synthase and Leucoanthocyanidin Dioxygenase Genes in Arabidopsis. Further Evidence for Differential Regulation of “Early” and “Late” Genes. Plant Physiol. 1997, 113, 1437–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrahams, S.; Lee, E.; Walker, A.R.; Tanner, G.J.; Larkin, P.J.; Ashton, A.R. The Arabidopsis TDS4 Gene Encodes Leucoanthocyanidin Dioxygenase (LDOX) and Is Essential for Proanthocyanidin Synthesis and Vacuole Development. Plant J. 2003, 35, 624–636. [Google Scholar] [CrossRef] [PubMed]
- Tanner, G.J.; Francki, K.T.; Abrahams, S.; Watson, J.M.; Larkin, P.J.; Ashton, A.R. Proanthocyanidin Biosynthesis in Plants: Purification of Legume LEUCOANTHOCYANIDIN REDUCTASE and Molecular Cloning of its cDNA. J. Biol. Chem. 2003, 278, 31647–31656. [Google Scholar] [CrossRef] [Green Version]
- Xie, D.-Y.; Sharma, S.B.; Paiva, N.L.; Ferreira, D.; Dixon, R.A. Role of Anthocyanidin Reductase, Encoded by BANYULS in Plant Flavonoid Biosynthesis. Science 2003, 299, 396–399. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Nakayama, T.; Yonekura-Sakakibara, K.; Fukui, Y.; Nakamura, N.; Yamaguchi, M.; Tanaka, Y.; Kusumi, T.; Nishino, T. CDNA Cloning, Heterologous Expressions, and Functional Characterization of Malonyl-Coenzyme A: Anthocyanidin 3-O-Glucoside-6"-O-Malonyltransferase from Dahlia Flowers. Plant Physiol. 2002, 130, 2142–2151. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, N.; Nishizaki, Y.; Ozeki, Y.; Miyahara, T. The Role of Acyl-Glucose in Anthocyanin Modifications. Molecules 2014, 19, 18747–18766. [Google Scholar] [CrossRef]
- Du, H.; Wu, J.; Ji, K.-X.; Zeng, Q.-Y.; Bhuiya, M.-W.; Su, S.; Shu, Q.-Y.; Ren, H.-X.; Liu, Z.-A.; Wang, L.-S. Methylation Mediated by an Anthocyanin, O-Methyltransferase, Is Involved in Purple Flower Coloration in Paeonia. J. Exp. Bot. 2015, 66, 6563–6577. [Google Scholar] [CrossRef] [Green Version]
- Stafford, H.A. The Metabolism of Aromatic Compounds. Annu. Rev. Plant Physiol. 1974, 25, 459–486. [Google Scholar] [CrossRef]
- Winkel, B.S.J. Metabolic Channeling in Plants. Annu. Rev. Plant Biol. 2004, 55, 85–107. [Google Scholar] [CrossRef]
- Nakayama, T.; Takahashi, S.; Waki, T. Formation of Flavonoid Metabolons: Functional Significance of Protein-Protein Interactions and Impact on Flavonoid Chemodiversity. Front. Plant Sci. 2019, 10, 821. [Google Scholar] [CrossRef] [PubMed]
- Tohge, T.; Nishiyama, Y.; Hirai, M.Y.; Yano, M.; Nakajima, J.; Awazuhara, M.; Inoue, E.; Takahashi, H.; Goodenowe, D.B.; Kitayama, M.; et al. Functional Genomics by Integrated Analysis of Metabolome and Transcriptome of Arabidopsis Plants Over-Expressing an MYB Transcription Factor. Plant J. 2005, 42, 218–235. [Google Scholar] [CrossRef] [PubMed]
- Stintzing, F.C.; Carle, R. Functional Properties of Anthocyanins and Betalains in Plants, Food, and in Human Nutrition. Trends Food Sci. Technol. 2004, 15, 19–38. [Google Scholar] [CrossRef]
- Luo, J.; Nishiyama, Y.; Fuell, C.; Taguchi, G.; Elliott, K.; Hill, L.; Tanaka, Y.; Kitayama, M.; Yamazaki, M.; Bailey, P.; et al. Convergent Evolution in the BAHD Family of Acyl Transferases: Identification and Characterization of Anthocyanin Acyl Transferases from Arabidopsis thaliana. Plant J. 2007, 50, 678–695. [Google Scholar] [CrossRef]
- Offen, W.; Martinez-Fleites, C.; Yang, M.; Kiat-Lim, E.; Davis, B.G.; Tarling, C.A.; Ford, C.M.; Bowles, D.J.; Davies, G.J. Structure of a Flavonoid Glucosyltransferase Reveals the Basis for Plant Natural Product Modification. EMBO J. 2006, 25, 1396–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, X.; Zhuang, J.; Wu, Y.; Wang, P.; Zhao, G.; Liu, Y.; Jiang, X.; Gao, L.; Xia, T. Identification of a Flavonoid Glucosyltransferase Involved in 7-OH Site Glycosylation in Tea Plants (Camellia sinensis). Sci. Rep. 2017, 7, 5926. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, S.; Xu, Z.; Li, M.; Chen, K.; Zhang, Y.; Hu, Z.; Zhang, M.; Zhang, Z.; Qiao, X.; et al. Highly Promiscuous Flavonoid 3-O-Glycosyltransferase from Scutellaria baicalensis. Org. Lett. 2019, 21, 2241–2245. [Google Scholar] [CrossRef]
- Alseekh, S.; Perez de Souza, L.; Benina, M.; Fernie, A.R. The Style and Substance of Plant Flavonoid Decoration; towards Defining Both Structure and Function. Phytochemistry 2020, 174, 112347. [Google Scholar] [CrossRef]
- Poustka, F.; Irani, N.G.; Feller, A.; Lu, Y.; Pourcel, L.; Frame, K.; Grotewold, E. A Trafficking Pathway for Anthocyanins Overlaps with the Endoplasmic Reticulum-to-Vacuole Protein-Sorting Route in Arabidopsis and Contributes to the Formation of Vacuolar Inclusions. Plant Physiol. 2007, 145, 1323–1335. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Dixon, R.A. The “ins” and “Outs” of Flavonoid Transport. Trends Plant Sci. 2010, 15, 72–80. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Li, H.; Huang, J.-R. Arabidopsis TT19 Functions as a Carrier to Transport Anthocyanin from the Cytosol to Tonoplasts. Mol. Plant 2012, 5, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Huhman, D.; Shadle, G.; He, X.-Z.; Sumner, L.W.; Tang, Y.; Dixon, R.A. MATE2 Mediates Vacuolar Sequestration of Flavonoid Glycosides and Glycoside Malonates in Medicago truncatula. Plant Cell 2011, 23, 1536–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grotewold, E.; Davies, K. Trafficking and Sequestration of Anthocyanins. Nat. Prod. Commun. 2008, 3. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J. Flavonoid Transport Mechanisms: How to Go, and with Whom. Trends Plant Sci. 2015, 20, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Marinova, K.; Kleinschmidt, K.; Weissenböck, G.; Klein, M. Flavonoid Biosynthesis in Barley Primary Leaves Requires the Presence of the Vacuole and Controls the Activity of Vacuolar Flavonoid Transport. Plant Physiol. 2007, 144, 432–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oettmeier, W.; Heupel, A. Identification of Flavonoids and Cinnamic Acid Derivatives from Spinach Chloroplast Preparations. Z. Nat. B 1972, 27, 177–183. [Google Scholar] [CrossRef]
- Hutzler, P.; Fischbach, R.; Heller, W.; Jungblut, T.P.; Reuber, S.; Schmitz, R.; Veit, M.; Weissenböck, G.; Schnitzler, J.-P. Tissue Localization of Phenolic Compounds in Plants by Confocal Laser Scanning Microscopy. J. Exp. Bot. 1998, 49, 953–965. [Google Scholar] [CrossRef]
- Peer, W.A.; Brown, D.E.; Tague, B.W.; Muday, G.K.; Taiz, L.; Murphy, A.S. Flavonoid Accumulation Patterns of Transparent Testa Mutants of Arabidopsis. Plant Physiol. 2001, 126, 536–548. [Google Scholar] [CrossRef] [Green Version]
- Buer, C.S.; Muday, G.K. The Transparent Testa4 Mutation Prevents Flavonoid Synthesis and Alters Auxin Transport and the Response of Arabidopsis Roots to Gravity and Light. Plant Cell 2004, 16, 1191–1205. [Google Scholar] [CrossRef] [Green Version]
- Feucht, W.; Treutter, D.; Polster, J. Flavanol Binding of Nuclei from Tree Species. Plant Cell Rep. 2004, 22, 430–436. [Google Scholar] [CrossRef]
- Agati, G.; Matteini, P.; Goti, A.; Tattini, M. Chloroplast-Located Flavonoids Can Scavenge Singlet Oxygen. New Phytol. 2007, 174, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Böttner, L.; Grabe, V.; Gablenz, S.; Böhme, N.; Appenroth, K.J.; Gershenzon, J.; Huber, M. Differential Localization of Flavonoid Glucosides in an Aquatic Plant Implicates Different Functions under Abiotic Stress. Plant Cell Environ. 2021, 44, 900–914. [Google Scholar] [CrossRef] [PubMed]
- Sarma, A.D.; Sharma, R. Anthocyanin-DNA Copigmentation Complex: Mutual Protection against Oxidative Damage. Phytochemistry 1999, 52, 1313–1318. [Google Scholar] [CrossRef]
- Naoumkina, M.; Dixon, R.A. Subcellular Localization of Flavonoid Natural Products: A Signaling Function? Plant Signal. Behav. 2008, 3, 573–575. [Google Scholar] [CrossRef] [Green Version]
- Saslowsky, D.E.; Warek, U.; Winkel, B.S.J. Nuclear Localization of Flavonoid Enzymes in Arabidopsis. J. Biol. Chem. 2005, 280, 23735–23740. [Google Scholar] [CrossRef] [Green Version]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as Antioxidants in Plants: Location and Functional Significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Tegeder, M.; Rentsch, D. Uptake and Partitioning of Amino Acids and Peptides. Mol. Plant 2010, 3, 997–1011. [Google Scholar] [CrossRef]
- Chen, L.-Q.; Cheung, L.S.; Feng, L.; Tanner, W.; Frommer, W.B. Transport of Sugars. Annu. Rev. Biochem. 2015, 84, 865–894. [Google Scholar] [CrossRef]
- Hedrich, R.; Sauer, N.; Neuhaus, H.E. Sugar Transport across the Plant Vacuolar Membrane: Nature and Regulation of Carrier Proteins. Curr. Opin. Plant Biol. 2015, 25, 63–70. [Google Scholar] [CrossRef]
- Forde, B.G. Nitrate Transporters in Plants: Structure, Function and Regulation. Biochim. Biophys. Acta BBA Biomembr. 2000, 1465, 219–235. [Google Scholar] [CrossRef]
- Buchner, P.; Takahashi, H.; Hawkesford, M.J. Plant Sulphate Transporters: Co-Ordination of Uptake, Intracellular and Long-Distance Transport. J. Exp. Bot. 2004, 55, 1765–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, T.; Nishizawa, N.K. Iron Uptake, Translocation, and Regulation in Higher Plants. Annu. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selmar, D.; Radwan, A.; Hijazin, T.; Abouzeid, S.; Yahyazadeh, M.; Lewerenz, L.; Kleinwächter, M.; Nowak, M. Horizontal Natural Product Transfer: Intriguing Insights into a Newly Discovered Phenomenon. J. Agric. Food Chem. 2019, 67, 8740–8745. [Google Scholar] [CrossRef] [PubMed]
- Trapp, S. Plant Uptake and Transport Models for Neutral and Ionic Chemicals. Environ. Sci. Pollut. Res. 2004, 11, 33. [Google Scholar] [CrossRef]
- Collins, C.D.; Martin, I.; Doucette, W. Plant Uptake of Xenobiotics. In Organic Xenobiotics and Plants: From Mode of Action to Ecophysiology; Schröder, P., Collins, C.D., Eds.; Plant Ecophysiology; Springer: Dordrecht, The Netherlands, 2011; pp. 3–16. ISBN 978-90-481-9852-8. [Google Scholar]
- Inoue, J.; Chamberlain, K.; Bromilow, R.H. Physicochemical Factors Affecting the Uptake by Roots and Translocation to Shoots of Amine Bases in Barley. Pestic. Sci. 1998, 54, 8–21. [Google Scholar] [CrossRef]
- Nwoko, C.O. Trends in Phytoremediation of Toxic Elemental and Organic Pollutants. Afr. J. Biotechnol. 2010, 9, 6010–6016. [Google Scholar] [CrossRef]
- Sibout, R.; Höfte, H. Plant Cell Biology: The ABC of Monolignol Transport. Curr. Biol. 2012, 22, R533–R535. [Google Scholar] [CrossRef] [Green Version]
- Cronin, M.T.; Livingstone, D.J. Calculation of Physicochemical Properties. In Predicting Chemical Toxicity and Fate, 1st ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 31–40. [Google Scholar]
- Trapp, S. Modelling Uptake into Roots and Subsequent Translocation of Neutral and Ionisable Organic Compounds. Pest Manag. Sci. 2000, 56, 767–778. [Google Scholar] [CrossRef]
- Trapp, S. Bioaccumulation of Polar and Ionizable Compounds in Plants. In Ecotoxicology Modeling; Springer: New York, NY, USA, 2009; pp. 299–353. [Google Scholar]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Settings. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Pollastri, M.P. Overview on the Rule of Five. Curr. Protoc. Pharmacol. 2010, 9, 12. [Google Scholar] [CrossRef]
- Yahyazadeh, M.; Nowak, M.; Kima, H.; Selmar, D. Horizontal Natural Product Transfer: A Potential Source of Alkaloidal Contaminants in Phytopharmaceuticals. Phytomedicine 2017, 34, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Hijazin, T.; Radwan, A.; Abouzeid, S.; Dräger, G.; Selmar, D. Uptake and Modification of Umbelliferone by Various Seedlings. Phytochemistry 2019, 157, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Shitan, N.; Sawada, K.; Van Montagu, M.C.; Inzé, D.; Rischer, H.; Goossens, A.; Oksman-Caldentey, K.-M.; Moriyama, Y.; Yazaki, K. Vacuolar Transport of Nicotine Is Mediated by a Multidrug and Toxic Compound Extrusion (MATE) Transporter in Nicotiana tabacum. Proc. Natl. Acad. Sci. USA 2009, 106, 2447–2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pakdeechanuan, P.; Shoji, T.; Hashimoto, T. Root-to-Shoot Translocation of Alkaloids Is Dominantly Suppressed in Nicotiana Alata. Plant Cell Physiol. 2012, 53, 1247–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shitan, N.; Hayashida, M.; Yazaki, K. Translocation and Accumulation of Nicotine via Distinct Spatio-Temporal Regulation of Nicotine Transporters in Nicotiana Tabacum. Plant Signal. Behav. 2015, 10, e1035852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, M.; Selmar, D. Cellular Distribution of Alkaloids and Their Translocation via Phloem and Xylem: The Importance of Compartment PH. Plant Biol. 2016, 18, 879–882. [Google Scholar] [CrossRef]
- Hijazin, T.; Radwan, A.; Lewerenz, L.; Abouzeid, S.; Selmar, D. The Uptake of Alkaloids by Plants from the Soil Is Determined by Rhizosphere PH. Rhizosphere 2020, 15, 100234. [Google Scholar] [CrossRef]
- Matile, P. Localization of Alkaloids and Mechanism of Their Accumulation in Vacuoles of Chelidonium Majus Laticifers. In Nova Acta Leopoldina. Supplementum 1976, 7, 139–156. [Google Scholar]
- Braidot, E.; Zancani, M.; Petrussa, E.; Peresson, C.; Bertolini, A.; Patui, S.; Macrì, F.; Vianello, A. Transport and Accumulation of Flavonoids in Grapevine (Vitis vinifera L.). Plant Signal. Behav. 2008, 3, 626–632. [Google Scholar] [CrossRef] [Green Version]
- Edwards, R.; Dixon, D.P.; Walbot, V. Plant Glutathione S-Transferases: Enzymes with Multiple Functions in Sickness and in Health. Trends Plant Sci. 2000, 5, 193–198. [Google Scholar] [CrossRef]
- Nozue, M.; Yamada, K.; Nakamura, T.; Kubo, H.; Kondo, M.; Nishimura, M. Expression of a Vacuolar Protein (VP24) in Anthocyanin-Producing Cells of Sweet Potato in Suspension Culture. Plant Physiol. 1997, 115, 1065–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, S. Transport of Flavonoids: From Cytosolic Synthesis to Vacuolar Accumulation. In The Science of Flavonoids; Grotewold, E., Ed.; Springer: New York, NY, USA, 2006; pp. 123–146. ISBN 978-0-387-28822-2. [Google Scholar]
- Marrs, K.A.; Alfenito, M.R.; Lloyd, A.M.; Walbot, V. A Glutathione S-Transferase Involved in Vacuolar Transfer Encoded by the Maize Gene Bronze-2. Nature 1995, 375, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Alfenito, M.R.; Souer, E.; Goodman, C.D.; Buell, R.; Mol, J.; Koes, R.; Walbot, V. Functional Complementation of Anthocyanin Sequestration in the Vacuole by Widely Divergent Glutathione S-Transferases. Plant Cell 1998, 10, 1135–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, S.; Shikazono, N.; Tanaka, A. TRANSPARENT TESTA 19 Is Involved in the Accumulation of Both Anthocyanins and Proanthocyanidins in Arabidopsis. Plant J. 2004, 37, 104–114. [Google Scholar] [CrossRef]
- Yamazaki, M.; Shibata, M.; Nishiyama, Y.; Springob, K.; Kitayama, M.; Shimada, N.; Aoki, T.; Ayabe, S.-I.; Saito, K. Differential Gene Expression Profiles of Red and Green Forms of Perilla frutescens Leading to Comprehensive Identification of Anthocyanin Biosynthetic Genes. FEBS J. 2008, 275, 3494–3502. [Google Scholar] [CrossRef]
- Conn, S.; Curtin, C.; Bézier, A.; Franco, C.; Zhang, W. Purification, Molecular Cloning, and Characterization of Glutathione S-Transferases (GSTs) from Pigmented Vitis Vinifera L. Cell Suspension Cultures as Putative Anthocyanin Transport Proteins. J. Exp. Bot. 2008, 59, 3621–3634. [Google Scholar] [CrossRef] [Green Version]
- Marrs, K.A. The Functions and Regulation of Glutathione S-Transferase in Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 127–158. [Google Scholar] [CrossRef]
- Mueller, L.A.; Goodman, C.D.; Silady, R.A.; Walbot, V. AN9, a Petunia Glutathione S-Transferase Required for Anthocyanin Sequestration, Is a Flavonoid-Binding Protein. Plant Physiol. 2000, 123, 1561–1570. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Dai, C.; Li, Y.; Feng, J.; Liu, Z.; Kang, C. Reduced Anthocyanins in Petioles Codes for a GST Anthocyanin Transporter That Is Essential for the Foliage and Fruit Coloration in Strawberry. J. Exp. Bot. 2018, 69, 2595–2608. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Chen, M.; He, N.; Chen, X.; Wang, N.; Sun, Q.; Zhang, T.; Xu, H.; Fang, H.; Wang, Y.; et al. MdGSTF6, Activated by MdMYB1, Plays an Essential Role in Anthocyanin Accumulation in Apple. Hortic. Res. 2019, 6, 40. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Dong, W.; Zhu, Y.; Allan, A.C.; Lin-Wang, K.; Xu, C. PpGST1, an Anthocyanin-Related Glutathione S-Transferase Gene, Is Essential for Fruit Coloration in Peach. Plant Biotechnol. J. 2020, 18, 1284–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kou, M.; Liu, Y.; Li, Z.; Zhang, Y.; Tang, W.; Yan, H.; Wang, X.; Chen, X.; Su, Z.; Arisha, M.H.; et al. A Novel Glutathione S-Transferase Gene from Sweetpotato, IbGSTF4, Is Involved in Anthocyanin Sequestration. Plant Physiol. Biochem. 2019, 135, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qi, Y.; Zhang, A.; Wu, H.; Liu, Z.; Ren, X. Molecular Cloning and Functional Characterization of AcGST1, an Anthocyanin-Related Glutathione S-Transferase Gene in Kiwifruit (Actinidia chinensis). Plant Mol. Biol. 2019, 100, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Díaz, R.; Madrid-Espinoza, J.; Salinas-Cornejo, J.; González-Villanueva, E.; Ruiz-Lara, S. Differential Roles for VviGST1, VviGST3, and VviGST4 in Proanthocyanidin and Anthocyanin Transport in Vitis vinífera. Front. Plant Sci. 2016, 7, 1166. [Google Scholar] [CrossRef] [Green Version]
- Klein, M.; Weissenböck, G.; Dufaud, A.; Gaillard, C.; Kreuz, K.; Martinoia, E. Different Energization Mechanisms Drive the Vacuolar Uptake of a Flavonoid Glucoside and a Herbicide Glucoside. J. Biol. Chem. 1996, 271, 29666–29671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazaki, K. Transporters of Secondary Metabolites. Curr. Opin. Plant Biol. 2005, 8, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Martinoia, E.; Klein, M.; Geisler, M.; Bovet, L.; Forestier, C.; Kolukisaoglu, Ü.; Müller-Röber, B.; Schulz, B. Multifunctionality of Plant ABC Transporters—More than Just Detoxifiers. Planta 2002, 214, 345–355. [Google Scholar] [CrossRef]
- Klein, M.; Burla, B.; Martinoia, E. The Multidrug Resistance-Associated Protein (MRP/ABCC) Subfamily of ATP-Binding Cassette Transporters in Plants. FEBS Lett. 2006, 580, 1112–1122. [Google Scholar] [CrossRef] [Green Version]
- Marinova, K.; Pourcel, L.; Weder, B.; Schwarz, M.; Barron, D.; Routaboul, J.-M.; Debeaujon, I.; Klein, M. The Arabidopsis MATE Transporter TT12 Acts as a Vacuolar Flavonoid/H+ -Antiporter Active in Proanthocyanidin-Accumulating Cells of the Seed Coat. Plant Cell 2007, 19, 2023–2038. [Google Scholar] [CrossRef] [Green Version]
- Passamonti, S.; Cocolo, A.; Braidot, E.; Petrussa, E.; Peresson, C.; Medic, N.; Macri, F.; Vianello, A. Characterization of Electrogenic Bromosulfophthalein Transport in Carnation Petal Microsomes and Its Inhibition by Antibodies against Bilitranslocase. FEBS J. 2005, 272, 3282–3296. [Google Scholar] [CrossRef] [Green Version]
- Braidot, E.; Petrussa, E.; Bertolini, A.; Peresson, C.; Ermacora, P.; Loi, N.; Terdoslavich, M.; Passamonti, S.; Macrì, F.; Vianello, A. Evidence for a Putative Flavonoid Translocator Similar to Mammalian Bilitranslocase in Grape Berries (Vitis Vinifera L.) during Ripening. Planta 2008, 228, 203. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Liao, L.; Zhou, H.; Gu, C.; Wang, L.; Han, Y. A Small Indel Mutation in an Anthocyanin Transporter Causes Variegated Colouration of Peach Flowers. J. Exp. Bot. 2015, 66, 7227–7239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, L.; Zhu, Y.; Liu, R.; Zhang, A.; Zhu, M.; Xu, W.; Lin, A.; Lu, K.; Li, J. Genome Wide Identification and Comparative Analysis of Glutathione Transferases (GST) Family Genes in Brassica Napus. Sci. Rep. 2019, 9, 9196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilperte, V.; Boehm, R.; Debener, T. A Highly Mutable GST Is Essential for Bract Colouration in Euphorbia Pulcherrima Willd. Ex Klotsch. BMC Genom. 2021, 22, 208. [Google Scholar] [CrossRef] [PubMed]
- Lai, B.; You, Y.; Zhang, L.; Wang, Q.; Chen, F.; Luo, G.; Du, L.; Wang, H. Identification and Functional Characterization of RsGST1, an Anthocyanin-Related Glutathione S-Transferase Gene in Radish. J. Plant Physiol. 2021, 263, 153468. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Zhao, J.; Lai, B.; Qin, Y.; Wang, H.; Hu, G. LcGST4 Is an Anthocyanin-Related Glutathione S-Transferase Gene in Litchi Chinensis Sonn. Plant Cell Rep. 2016, 35, 831–843. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.; Akita, Y.; Ishizaka, H.; Narumi, I.; Tanaka, A. Molecular Characterization of an Anthocyanin-Related Glutathione S-Transferase Gene in Cyclamen. J. Plant Physiol. 2012, 169, 636–642. [Google Scholar] [CrossRef]
- Sasaki, N.; Nishizaki, Y.; Uchida, Y.; Wakamatsu, E.; Umemoto, N.; Momose, M.; Okamura, M.; Yoshida, H.; Yamaguchi, M.; Nakayama, M.; et al. Identification of the Glutathione S-Transferase Gene Responsible for Flower Color Intensity in Carnations. Plant Biotechnol. 2012, 29, 223–227. [Google Scholar] [CrossRef] [Green Version]
- Loyall, L.; Uchida, K.; Braun, S.; Furuya, M.; Frohnmeyer, H. Glutathione and a UV Light–Induced Glutathione S-Transferase Are Involved in Signaling to Chalcone Synthase in Cell Cultures. Plant Cell 2000, 12, 1939–1950. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, X.; Li, F.; Xiang, L.; Chen, K. The Isolation and Identification of Anthocyanin-Related GSTs in Chrysanthemum. Horticulturae 2021, 7, 231. [Google Scholar] [CrossRef]
- Wei, K.; Wang, L.; Zhang, Y.; Ruan, L.; Li, H.; Wu, L.; Xu, L.; Zhang, C.; Zhou, X.; Cheng, H.; et al. A Coupled Role for CsMYB75 and CsGSTF1 in Anthocyanin Hyperaccumulation in Purple Tea. Plant J. 2019, 97, 825–840. [Google Scholar] [CrossRef] [PubMed]
- Meng, G.; Clausen, S.K.; Rasmussen, S.K. Transcriptome Analysis Reveals Candidate Genes Related to Anthocyanin Biosynthesis in Different Carrot Genotypes and Tissues. Plants 2020, 9, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iorizzo, M.; Curaba, J.; Pottorff, M.; Ferruzzi, M.G.; Simon, P.; Cavagnaro, P.F. Carrot Anthocyanins Genetics and Genomics: Status and Perspectives to Improve Its Application for the Food Colorant Industry. Genes 2020, 11, 906. [Google Scholar] [CrossRef] [PubMed]
- Thompson, E.P.; Wilkins, C.; Demidchik, V.; Davies, J.M.; Glover, B.J. An Arabidopsis Flavonoid Transporter Is Required for Anther Dehiscence and Pollen Development. J. Exp. Bot. 2010, 61, 439–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Dixon, R.A. MATE Transporters Facilitate Vacuolar Uptake of Epicatechin 3′-O-Glucoside for Proanthocyanidin Biosynthesis in Medicago Truncatula and Arabidopsis. Plant Cell 2009, 21, 2323–2340. [Google Scholar] [CrossRef] [Green Version]
- Gomez, C.; Terrier, N.; Torregrosa, L.; Vialet, S.; Fournier-Level, A.; Verriès, C.; Souquet, J.-M.; Mazauric, J.-P.; Klein, M.; Cheynier, V.; et al. Grapevine MATE-Type Proteins Act as Vacuolar H+-Dependent Acylated Anthocyanin Transporters. Plant Physiol. 2009, 150, 402–415. [Google Scholar] [CrossRef] [Green Version]
- Gomez, C.; Conejero, G.; Torregrosa, L.; Cheynier, V.; Terrier, N.; Ageorges, A. In Vivo Grapevine Anthocyanin Transport Involves Vesicle-Mediated Trafficking and the Contribution of AnthoMATE Transporters and GST. Plant J. 2011, 67, 960–970. [Google Scholar] [CrossRef]
- Pérez-Díaz, R.; Ryngajllo, M.; Pérez-Díaz, J.; Peña-Cortés, H.; Casaretto, J.A.; González-Villanueva, E.; Ruiz-Lara, S. VvMATE1 and VvMATE2 Encode Putative Proanthocyanidin Transporters Expressed during Berry Development in Vitis Vinifera L. Plant Cell Rep. 2014, 33, 1147–1159. [Google Scholar] [CrossRef]
- Xu, H.; Yang, P.; Cao, Y.; Tang, Y.; He, G.; Xu, L.; Ming, J. Cloning and Functional Characterization of a Flavonoid Transport-Related MATE Gene in Asiatic Hybrid Lilies (Lilium spp.). Genes 2020, 11, 418. [Google Scholar] [CrossRef]
- Frank, S.; Keck, M.; Sagasser, M.; Niehaus, K.; Weisshaar, B.; Stracke, R. Two Differentially Expressed MATE Factor Genes from Apple Complement the Arabidopsis Transparent Testa12 Mutant. Plant Biol. 2011, 13, 42–50. [Google Scholar] [CrossRef]
- Chai, Y.-R.; Lei, B.; Huang, H.-L.; Li, J.-N.; Yin, J.-M.; Tang, Z.-L.; Wang, R.; Chen, L. TRANSPARENT TESTA 12 Genes from Brassica Napus and Parental Species: Cloning, Evolution, and Differential Involvement in Yellow Seed Trait. Mol. Genet. Genom. 2009, 281, 109–123. [Google Scholar] [CrossRef] [PubMed]
- M’mbone, M.E.; Cheng, W.; Xu, L.; Wang, Y.; Karanja, B.K.; Zhu, X.; Cao, Y.; Liu, L. Identification and Transcript Analysis of MATE Genes Involved in Anthocyanin Transport in Radish (Raphanus sativus L.). Sci. Hortic. 2018, 238, 195–203. [Google Scholar] [CrossRef]
- Borevitz, J.O.; Xia, Y.; Blount, J.; Dixon, R.A.; Lamb, C. Activation Tagging Identifies a Conserved MYB Regulator of Phenylpropanoid Biosynthesis. Plant Cell 2000, 12, 2383–2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Liu, Y.; Liu, H.; Kang, L.; Geng, J.; Gai, Y.; Ding, Y.; Sun, H.; Li, Y. Identification and Expression Analysis of MATE Genes Involved in Flavonoid Transport in Blueberry Plants. PLoS ONE 2015, 10, e0118578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, M.-S.; Ku, Y.-S.; Yung, W.-S.; Cheng, S.-S.; Man, C.-K.; Yang, L.; Song, S.; Chung, G.; Lam, H.-M. MATE-Type Proteins Are Responsible for Isoflavone Transportation and Accumulation in Soybean Seeds. Int. J. Mol. Sci. 2021, 22, 12017. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-Y.; Tang, Y.-M.; Hu, Y.-Y.; Wang, Y.; Sun, B.; Wang, X.-R.; Tang, H.-R.; Chen, Q. FaTT12-1, a Multidrug and Toxin Extrusion (MATE) Member Involved in Proanthocyanidin Transport in Strawberry Fruits. Sci. Hortic. 2018, 231, 158–165. [Google Scholar] [CrossRef]
- Xu, L.; Shen, Z.-L.; Chen, W.; Si, G.-Y.; Meng, Y.; Guo, N.; Sun, X.; Cai, Y.-P.; Lin, Y.; Gao, J.-S. Phylogenetic Analysis of Upland Cotton MATE Gene Family Reveals a Conserved Subfamily Involved in Transport of Proanthocyanidins. Mol. Biol. Rep. 2019, 46, 161–175. [Google Scholar] [CrossRef]
- Yang, S.; Jiang, Y.; Xu, L.; Shiratake, K.; Luo, Z.; Zhang, Q. Molecular Cloning and Functional Characterization of DkMATE1 Involved in Proanthocyanidin Precursor Transport in Persimmon (Diospyros kaki Thunb.) Fruit. Plant Physiol. Biochem. 2016, 108, 241–250. [Google Scholar] [CrossRef]
- Goodman, C.D.; Casati, P.; Walbot, V. A Multidrug Resistance–Associated Protein Involved in Anthocyanin Transport in Zea Mays. Plant Cell 2004, 16, 1812–1826. [Google Scholar] [CrossRef] [Green Version]
- Behrens, C.E.; Smith, K.E.; Iancu, C.V.; Choe, J.; Dean, J.V. Transport of Anthocyanins and Other Flavonoids by the Arabidopsis ATP-Binding Cassette Transporter AtABCC2. Sci. Rep. 2019, 9, 437. [Google Scholar] [CrossRef] [Green Version]
- Francisco, R.M.; Regalado, A.; Ageorges, A.; Burla, B.J.; Bassin, B.; Eisenach, C.; Zarrouk, O.; Vialet, S.; Marlin, T.; Chaves, M.M.; et al. ABCC1, an ATP Binding Cassette Protein from Grape Berry, Transports Anthocyanidin 3-O-Glucosides. Plant Cell 2013, 25, 1840–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.-L.; Xie, X.-R.; Zhang, J.; Xiang, G.; Li, Y.; Wu, H.-B. In Silico Analysis of a MRP Transporter Gene Reveals Its Possible Role in Anthocyanins or Flavonoids Transport in Oryze sativa. Am. J. Plant Sci. 2013, 4, 555–560. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Qiu, Y.; Duan, M.; Wang, J.; Zhang, X.; Wang, H.; Song, J.; Li, X. Identification of Anthocyanin Biosynthesis Related MicroRNAs in a Distinctive Chinese Radish (Raphanus sativus L.) by High-Throughput Sequencing. Mol. Genet. Genom. 2017, 292, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Baxter, I.R.; Young, J.C.; Armstrong, G.; Foster, N.; Bogenschutz, N.; Cordova, T.; Peer, W.A.; Hazen, S.P.; Murphy, A.S.; Harper, J.F. A Plasma Membrane H+-ATPase Is Required for the Formation of Proanthocyanidins in the Seed Coat Endothelium of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2005, 102, 2649–2654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appelhagen, I.; Nordholt, N.; Seidel, T.; Spelt, K.; Koes, R.; Quattrochio, F.; Sagasser, M.; Weisshaar, B. TRANSPARENT TESTA 13 Is a Tonoplast P3A-ATPase Required for Vacuolar Deposition of Proanthocyanidins in Arabidopsis thaliana Seeds. Plant J. 2015, 82, 840–849. [Google Scholar] [CrossRef]
- Verweij, W.; Spelt, C.; Di Sansebastiano, G.-P.; Vermeer, J.; Reale, L.; Ferranti, F.; Koes, R.; Quattrocchio, F. An H+ P-ATPase on the Tonoplast Determines Vacuolar PH and Flower Colour. Nat. Cell Biol. 2008, 10, 1456–1462. [Google Scholar] [CrossRef]
- Sundaramoorthy, J.; Park, G.T.; Lee, J.-D.; Kim, J.H.; Seo, H.S.; Song, J.T. A P3A-Type ATPase and an R2R3-MYB Transcription Factor Are Involved in Vacuolar Acidification and Flower Coloration in Soybean. Front. Plant Sci. 2020, 11, 1880. [Google Scholar] [CrossRef]
- Sarafian, V.; Kim, Y.; Poole, R.J.; Rea, P.A. Molecular Cloning and Sequence of CDNA Encoding the Pyrophosphate-Energized Vacuolar Membrane Proton Pump of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1992, 89, 1775–1779. [Google Scholar] [CrossRef] [Green Version]
- Ichino, T.; Fuji, K.; Ueda, H.; Takahashi, H.; Koumoto, Y.; Takagi, J.; Tamura, K.; Sasaki, R.; Aoki, K.; Shimada, T.; et al. GFS9/TT9 Contributes to Intracellular Membrane Trafficking and Flavonoid Accumulation in Arabidopsis thaliana. Plant J. 2014, 80, 410–423. [Google Scholar] [CrossRef]
- Ichino, T.; Maeda, K.; Hara-Nishimura, I.; Shimada, T. Arabidopsis ECHIDNA Protein Is Involved in Seed Coloration, Protein Trafficking to Vacuoles, and Vacuolar Biogenesis. J. Exp. Bot. 2020, 71, 3999–4009. [Google Scholar] [CrossRef]
- Kulich, I.; Pečenková, T.; Sekereš, J.; Smetana, O.; Fendrych, M.; Foissner, I.; Höftberger, M.; Žárský, V. Arabidopsis Exocyst Subcomplex Containing Subunit EXO70B1 Is Involved in Autophagy-Related Transport to the Vacuole. Traffic 2013, 14, 1155–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathews, H.; Clendennen, S.K.; Caldwell, C.G.; Liu, X.L.; Connors, K.; Matheis, N.; Schuster, D.K.; Menasco, D.J.; Wagoner, W.; Lightner, J.; et al. Activation Tagging in Tomato Identifies a Transcriptional Regulator of Anthocyanin Biosynthesis, Modification, and Transport. Plant Cell 2003, 15, 1689–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butelli, E.; Titta, L.; Giorgio, M.; Mock, H.-P.; Matros, A.; Peterek, S.; Schijlen, E.G.W.M.; Hall, R.D.; Bovy, A.G.; Luo, J.; et al. Enrichment of Tomato Fruit with Health-Promoting Anthocyanins by Expression of Select Transcription Factors. Nat. Biotechnol. 2008, 26, 1301–1308. [Google Scholar] [CrossRef]
- Debeaujon, I.; Peeters, A.J.M.; Léon-Kloosterziel, K.M.; Koornneef, M. The TRANSPARENT TESTA12 Gene of Arabidopsis Encodes a Multidrug Secondary Transporter-like Protein Required for Flavonoid Sequestration in Vacuoles of the Seed Coat Endothelium. Plant Cell 2001, 13, 853–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimoda, K.; Otsuka, T.; Morimoto, Y.; Hamada, H.; Hamada, H. Glycosylation and Malonylation of Quercetin, Epicatechin, and Catechin by Cultured Plant Cells. Chem. Lett. 2007, 36, 1292–1293. [Google Scholar] [CrossRef]
- Pang, Y.; Peel, G.J.; Sharma, S.B.; Tang, Y.; Dixon, R.A. A Transcript Profiling Approach Reveals an Epicatechin-Specific Glucosyltransferase Expressed in the Seed Coat of Medicago truncatula. Proc. Natl. Acad. Sci. USA 2008, 105, 14210–14215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marles, M.A.S.; Ray, H.; Gruber, M.Y. New Perspectives on Proanthocyanidin Biochemistry and Molecular Regulation. Phytochemistry 2003, 64, 367–383. [Google Scholar] [CrossRef]
- Frangne, N.; Eggmann, T.; Koblischke, C.; Weissenböck, G.; Martinoia, E.; Klein, M. Flavone Glucoside Uptake into Barley Mesophyll and Arabidopsis Cell Culture Vacuoles. Energization Occurs by H+-Antiport and ATP-Binding Cassette-Type Mechanisms. Plant Physiol. 2002, 128, 726–733. [Google Scholar] [CrossRef]
- Klein, M.; Martinoia, E.; Hoffmann-Thoma, G.; Weissenböck, G. The ABC-like Vacuolar Transporter for Rye Mesophyll Flavone Glucuronides Is Not Species-Specific. Phytochemistry 2001, 56, 153–159. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Deroles, S.; Bennett, R.; Davies, K. New Insight into the Structures and Formation of Anthocyanic Vacuolar Inclusions in Flower Petals. BMC Plant Biol. 2006, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Grotewold, E.; Chamberlin, M.; Snook, M.; Siame, B.; Butler, L.; Swenson, J.; Maddock, S.; St. Clair, G.; Bowen, B. Engineering Secondary Metabolism in Maize Cells by Ectopic Expression of Transcription Factors. Plant Cell 1998, 10, 721–740. [Google Scholar] [CrossRef] [PubMed]
- Nozue, M.; Yasuda, H. Occurrence of Anthocyanoplasts in Cell Suspension Cultures of Sweet Potato. Plant Cell Rep. 1985, 4, 252–255. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Irani, N.G.; Grotewold, E. Sub-Cellular Trafficking of Phytochemicals Explored Using Auto-Fluorescent Compounds in Maize Cells. BMC Plant Biol. 2003, 3, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyder, B.A.; Nicholson, R.L. Synthesis of Phytoalexins in Sorghum as a Site-Specific Response to Fungal Ingress. Science 1990, 248, 1637–1639. [Google Scholar] [CrossRef] [PubMed]
- Malsam, J.; Kreye, S.; Söllner, T.H. Membrane Fusion: SNAREs and Regulation. Cell Mol. Life Sci. 2008, 65, 2814–2832. [Google Scholar] [CrossRef]
- Conn, S.; Franco, C.; Zhang, W. Characterization of Anthocyanic Vacuolar Inclusions in Vitis vinifera L. Cell Suspension Cultures. Planta 2010, 231, 1343–1360. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Pang, Y.; Dixon, R.A. The Mysteries of Proanthocyanidin Transport and Polymerization. Plant Physiol. 2010, 153, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, S.; Matsuda, F.; Tohge, T.; Yonekura-Sakakibara, K.; Yamazaki, M.; Saito, K.; Narumi, I. Metabolic Profiling and Cytological Analysis of Proanthocyanidins in Immature Seeds of Arabidopsis thaliana Flavonoid Accumulation Mutants. Plant J. 2010, 62, 549–559. [Google Scholar] [CrossRef]
- Nakamura, M. Development of Anthocyanoplasts in Relation to Coloration of ‘Kyoho’ Grapes. J. Jpn. Soc. Hortic. Sci. 1989, 58, 537–543. [Google Scholar] [CrossRef]
- Markham, K.R.; Gould, K.S.; Winefield, C.S.; Mitchell, K.A.; Bloor, S.J.; Boase, M.R. Anthocyanic Vacuolar Inclusions—Their Nature and Significance in Flower Colouration. Phytochemistry 2000, 55, 327–336. [Google Scholar] [CrossRef]
- Pecket, R.C.; Small, C.J. Occurrence, Location and Development of Anthocyanoplasts. Phytochemistry 1980, 19, 2571–2576. [Google Scholar] [CrossRef]
- Nozue, M.; Baba, S.; Kitamura, Y.; Xu, W.; Kubo, H.; Nogawa, M.; Shioiri, H.; Kojima, M. VP24 Found in Anthocyanic Vacuolar Inclusions (AVIs) of Sweet Potato Cells Is a Member of a Metalloprotease Family. Biochem. Eng. J. 2003, 3, 199–205. [Google Scholar] [CrossRef]
- Xu, W.; Moriya, K.; Yamada, K.; Nishimura, M.; Shioiri, H.; Kojima, M.; Nozue, M. Detection and Characterization of a 36-KDa Peptide in C-Terminal Region of a 24-KDa Vacuolar Protein (VP24) Precursor in Anthocyanin-Producing Sweet Potato Cells in Suspension Culture. Plant Sci. 2000, 160, 121–128. [Google Scholar] [CrossRef]
- Pourcel, L.; Irani, N.G.; Lu, Y.; Riedl, K.; Schwartz, S.; Grotewold, E. The Formation of Anthocyanic Vacuolar Inclusions in Arabidopsis thaliana and Implications for the Sequestration of Anthocyanin Pigments. Mol. Plant 2010, 3, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Small, C.J.; Pecket, R.C. The Ultrastructure of Anthocyanoplasts in Red-Cabbage. Planta 1982, 154, 97–99. [Google Scholar] [CrossRef] [PubMed]
- Nozue, M.; Kubo, H.; Nishimura, M.; Katou, A.; Hattori, C.; Usuda, N.; Nagata, T.; Yasuda, H. Characterization of Intravacuolar Pigmented Structures in Anthocyanin-Containing Cells of Sweet Potato Suspension Cultures. Plant Cell Physiol. 1993, 34, 803–808. [Google Scholar] [CrossRef]
- Conn, S.; Zhang, W.; Franco, C. Anthocyanic Vacuolar Inclusions (AVIs) Selectively Bind Acylated Anthocyanins in Vitis vinifera L. (Grapevine) Suspension Culture. Biotechnol. Lett. 2003, 25, 835–839. [Google Scholar] [CrossRef]
- Terrier, N.; Sauvage, F.X.; Ageorges, A.; Romieu, C. Changes in Acidity and in Proton Transport at the Tonoplast of Grape Berries during Development. Planta 2001, 213, 20–28. [Google Scholar] [CrossRef]
- Irani, N.G.; Grotewold, E. Light-Induced Morphological Alteration in Anthocyanin-Accumulating Vacuoles of Maize Cells. BMC Plant Biol. 2005, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Gendre, D.; Oh, J.; Boutté, Y.; Best, J.G.; Samuels, L.; Nilsson, R.; Uemura, T.; Marchant, A.; Bennett, M.J.; Grebe, M.; et al. Conserved Arabidopsis ECHIDNA Protein Mediates Trans–Golgi-Network Trafficking and Cell Elongation. Proc. Natl. Acad. Sci. USA 2011, 108, 8048–8053. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, K.; Huang, A.H.C. Tapetosomes in Brassica Tapetum Accumulate Endoplasmic Reticulum-Derived Flavonoids and Alkanes for Delivery to the Pollen Surface. Plant Cell 2007, 19, 582–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grunewald, S.; Marillonnet, S.; Hause, G.; Haferkamp, I.; Neuhaus, H.E.; Veß, A.; Hollemann, T.; Vogt, T. The Tapetal Major Facilitator NPF2.8 Is Required for Accumulation of Flavonol Glycosides on the Pollen Surface in Arabidopsis Thaliana. Plant Cell 2020, 32, 1727–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, T.; Ehmke, A.; Eilert, U.; von Borstel, K.; Theuring, C. Sites of Synthesis, Translocation and Accumulation of Pyrrolizidine Alkaloid N-Oxides in Senecio vulgaris L. Planta 1989, 177, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Selmar, D.; Lieberei, R.; Biehl, B. Mobilization and Utilization of Cyanogenic Glycosides: The Linustatin Pathway. Plant Physiol. 1988, 86, 711–716. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, M.E.; Nour-Eldin, H.H.; Halkier, B.A. Transport of Defense Compounds from Source to Sink: Lessons Learned from Glucosinolates. Trends Plant Sci. 2015, 20, 508–514. [Google Scholar] [CrossRef]
- Baldwin, I.T.; Karb, M.J.; Ohnmeiss, T.E. Allocation of 15N from Nitrate to Nicotine: Production and Turnover of a Damage-Induced Mobile Defense. Ecology 1994, 75, 1703–1713. [Google Scholar] [CrossRef]
- Weeks, W.W.; Bush, L.P. Alkaloid Changes in Tobacco Seeds during Germination. Plant Physiol. 1974, 53, 73–75. [Google Scholar] [CrossRef] [Green Version]
- Löffler, C.; Sahm, A.; Wray, V.; Czygan, F.-C.; Proksch, P. Soluble Phenolic Constituents from Cuscuta Reflexa and Cuscuta Platyloba. Biochem. Syst. Ecol. 1995, 23, 121–128. [Google Scholar] [CrossRef]
- Xu, Z.-S.; Yang, Q.-Q.; Feng, K.; Yu, X.; Xiong, A.-S. DcMYB113, a Root-Specific R2R3-MYB, Conditions Anthocyanin Biosynthesis and Modification in Carrot. Plant Biotechnol. J. 2020, 18, 1585–1597. [Google Scholar] [CrossRef] [Green Version]
- Pucker, B.; Iorizzo, M. Apiaceae FNS I Originated from F3H through Tandem Gene Duplication. bioRxiv 2022. [Google Scholar] [CrossRef]
- Shimizu, K.; Aoki, K. Development of Parasitic Organs of a Stem Holoparasitic Plant in Genus Cuscuta. Front. Plant Sci. 2019, 10, 1435. [Google Scholar] [CrossRef] [Green Version]
- Buer, C.S.; Muday, G.K.; Djordjevic, M.A. Flavonoids Are Differentially Taken up and Transported Long Distances in Arabidopsis. Plant Physiol. 2007, 145, 478–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenzel, C.L.; Hester, Q.; Mattsson, J. Identification of Genes Expressed in Vascular Tissues Using NPA-Induced Vascular Overgrowth in Arabidopsis. Plant Cell Physiol. 2008, 49, 457–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiyama, A.; Shitan, N.; Yazaki, K. Involvement of a Soybean ATP-Binding Cassette-Type Transporter in the Secretion of Genistein, a Signal Flavonoid in Legume-Rhizobium Symbiosis. Plant Physiol. 2007, 144, 2000–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pourcel, L.; Routaboul, J.-M.; Kerhoas, L.; Caboche, M.; Lepiniec, L.; Debeaujon, I. TRANSPARENT TESTA10 Encodes a Laccase-Like Enzyme Involved in Oxidative Polymerization of Flavonoids in Arabidopsis Seed Coat. Plant Cell 2005, 17, 2966–2980. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pucker, B.; Selmar, D. Biochemistry and Molecular Basis of Intracellular Flavonoid Transport in Plants. Plants 2022, 11, 963. https://doi.org/10.3390/plants11070963
Pucker B, Selmar D. Biochemistry and Molecular Basis of Intracellular Flavonoid Transport in Plants. Plants. 2022; 11(7):963. https://doi.org/10.3390/plants11070963
Chicago/Turabian StylePucker, Boas, and Dirk Selmar. 2022. "Biochemistry and Molecular Basis of Intracellular Flavonoid Transport in Plants" Plants 11, no. 7: 963. https://doi.org/10.3390/plants11070963
APA StylePucker, B., & Selmar, D. (2022). Biochemistry and Molecular Basis of Intracellular Flavonoid Transport in Plants. Plants, 11(7), 963. https://doi.org/10.3390/plants11070963