
Endophytic Fungus from Opuntia ficus-indica: A Source of Potential Bioactive Antimicrobial Compounds against Multidrug-Resistant Bacteria

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Endophytic Fungus Isolation and Identification
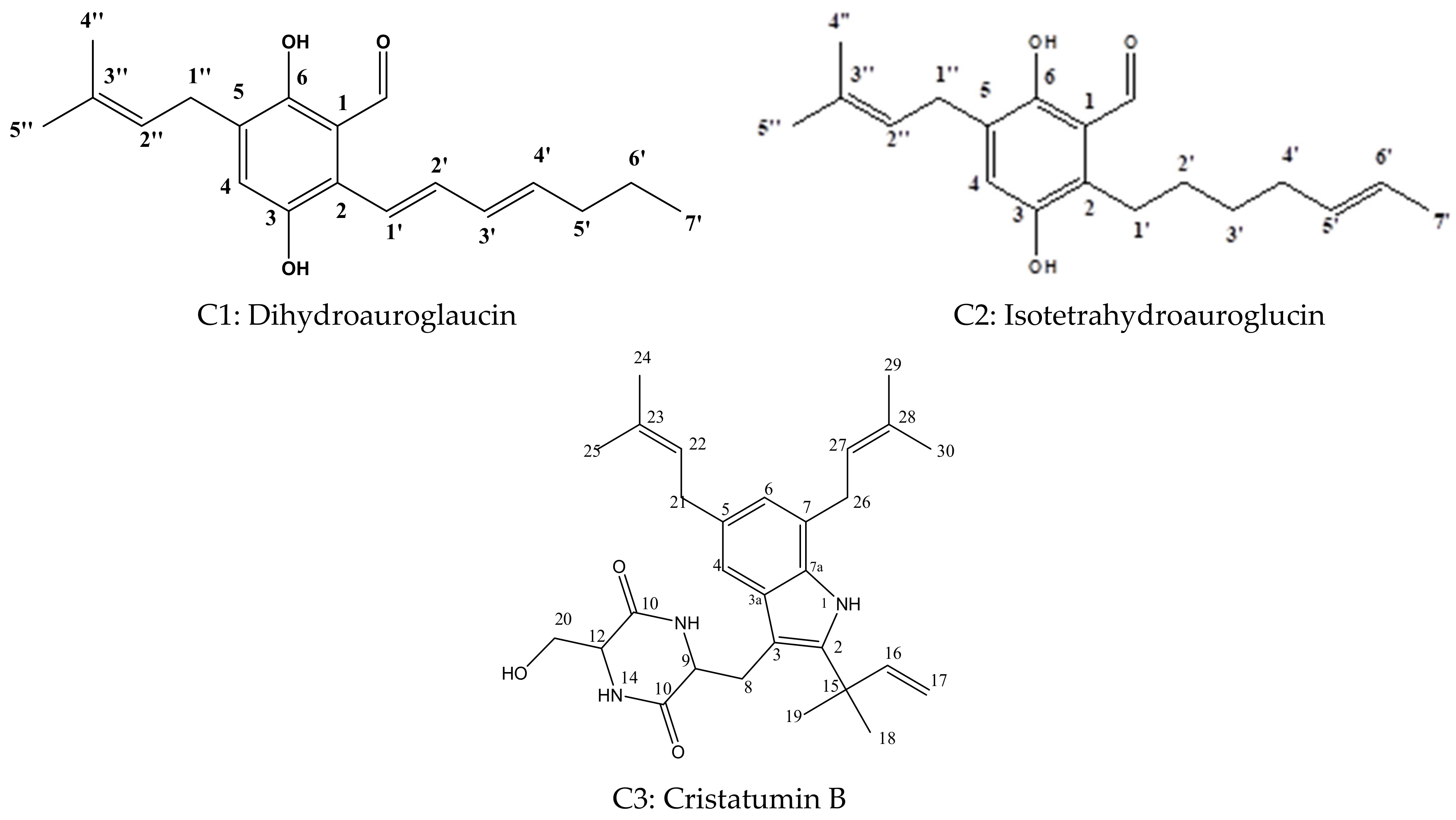
2.2. Identification of Fungal Ethyl Acetate Extract (FEA) Isolated Compounds
2.2.1. Dihydroauroglaucin (C1)
2.2.2. Isotetrahydroauroglaucin (C2)
2.2.3. Cristatumin B (C3)
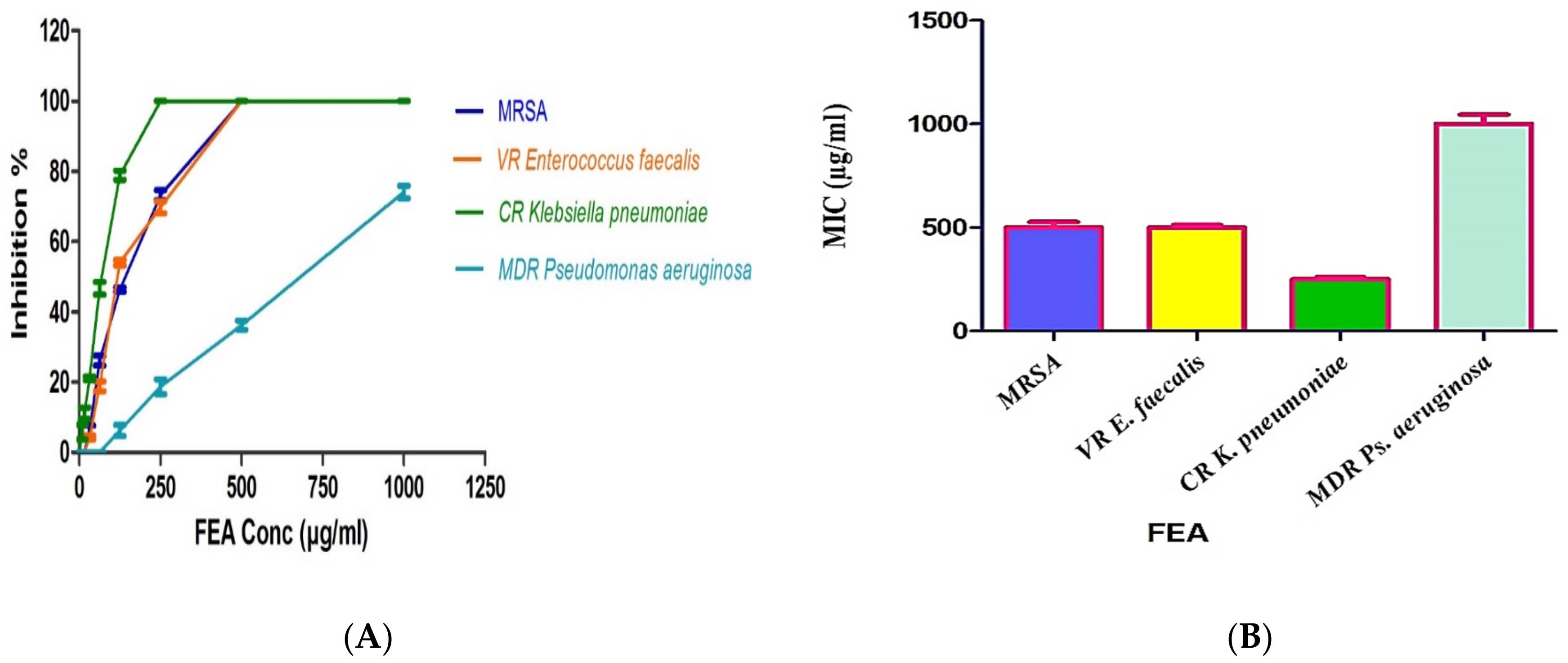
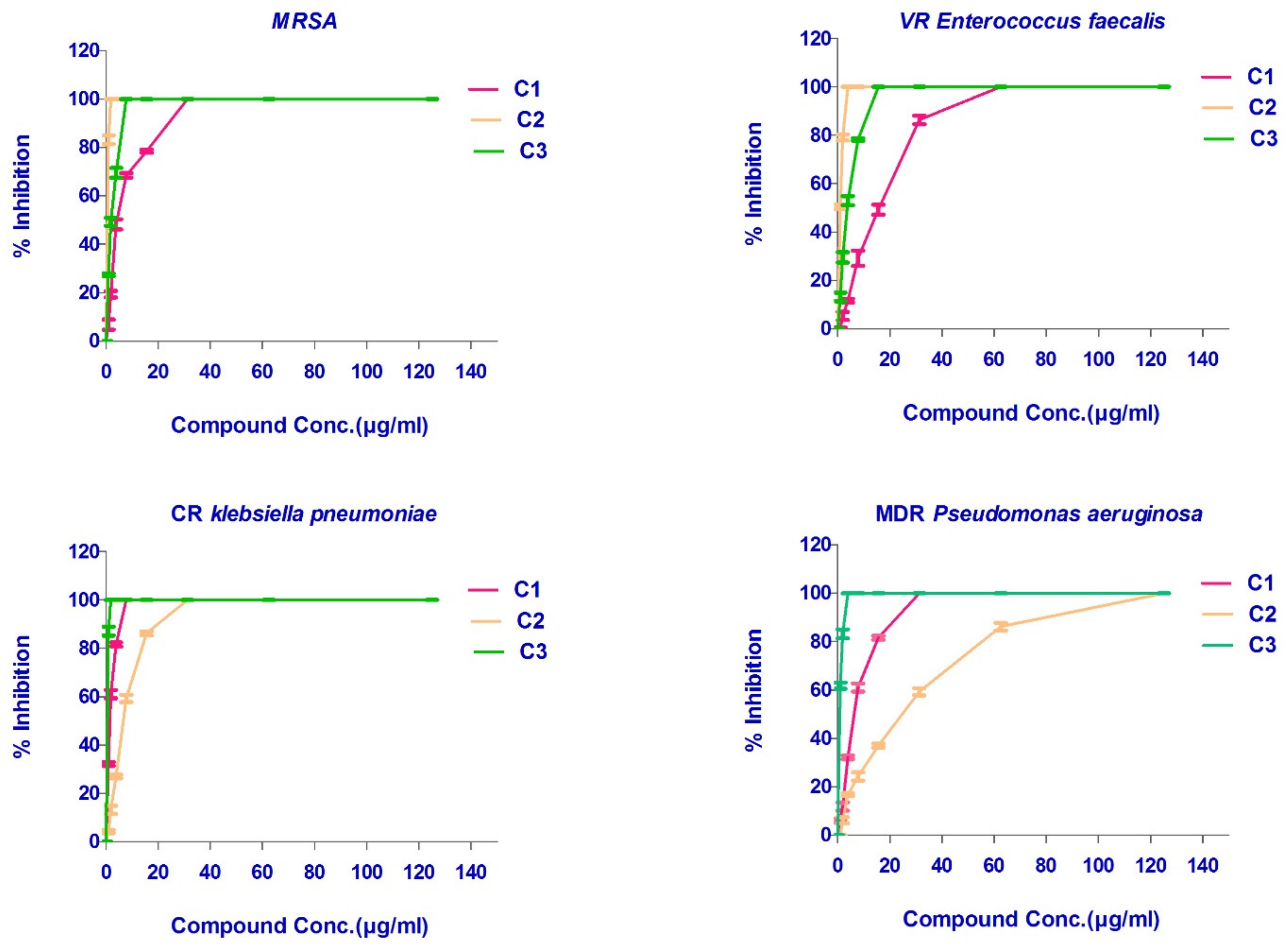
2.3. Antimicrobial Activity of the Isolated FEA and Its Compounds
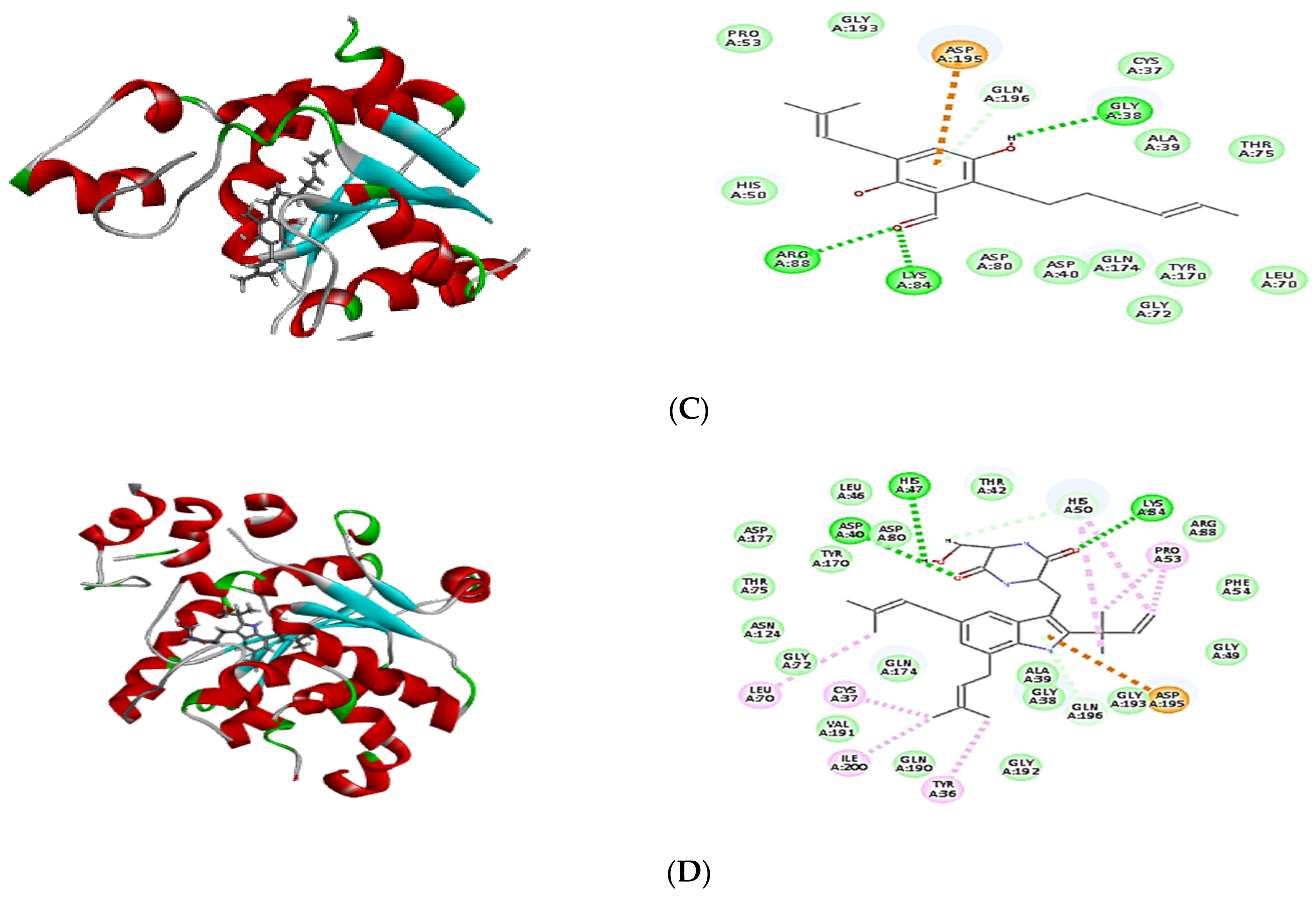
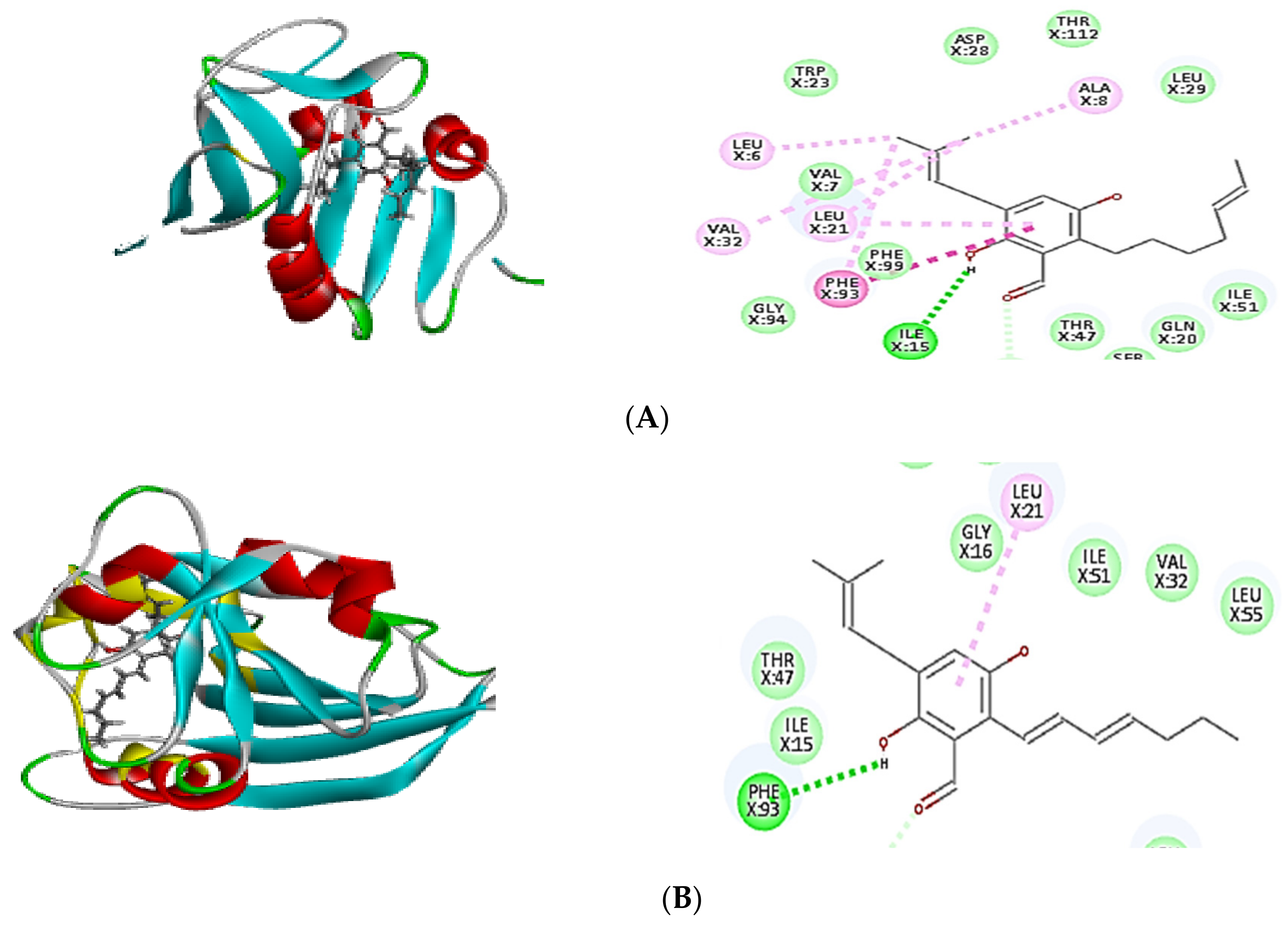
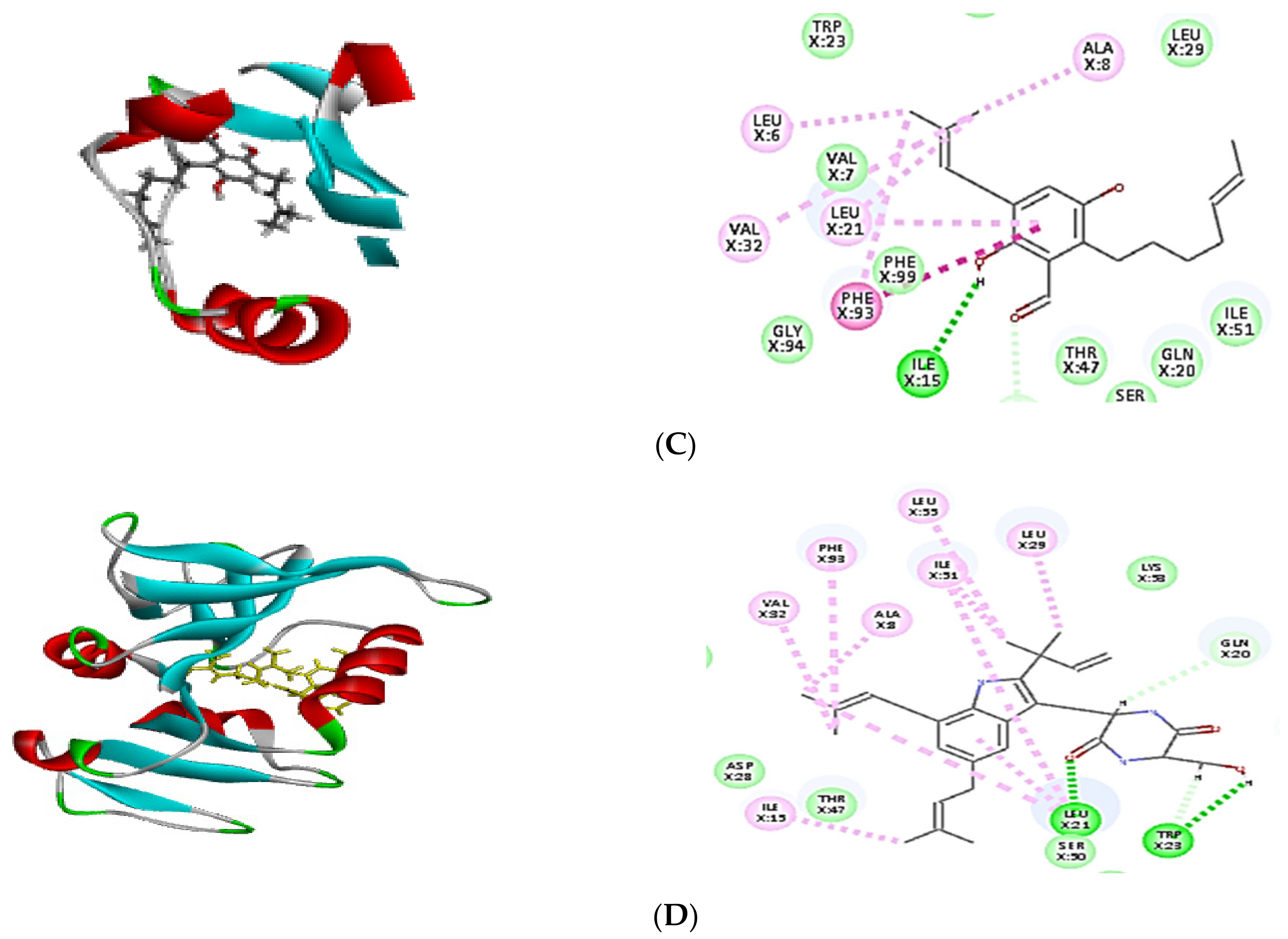
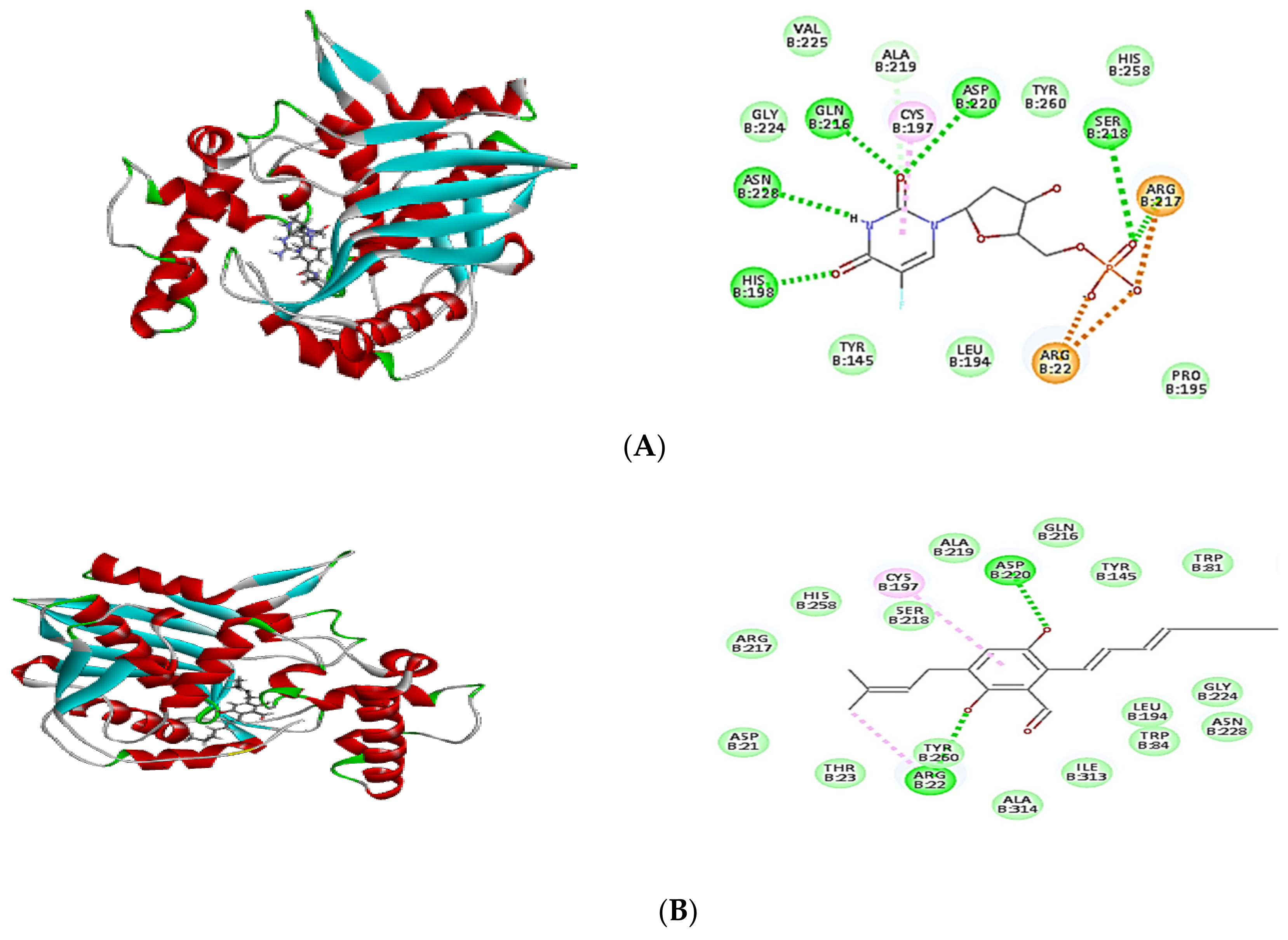
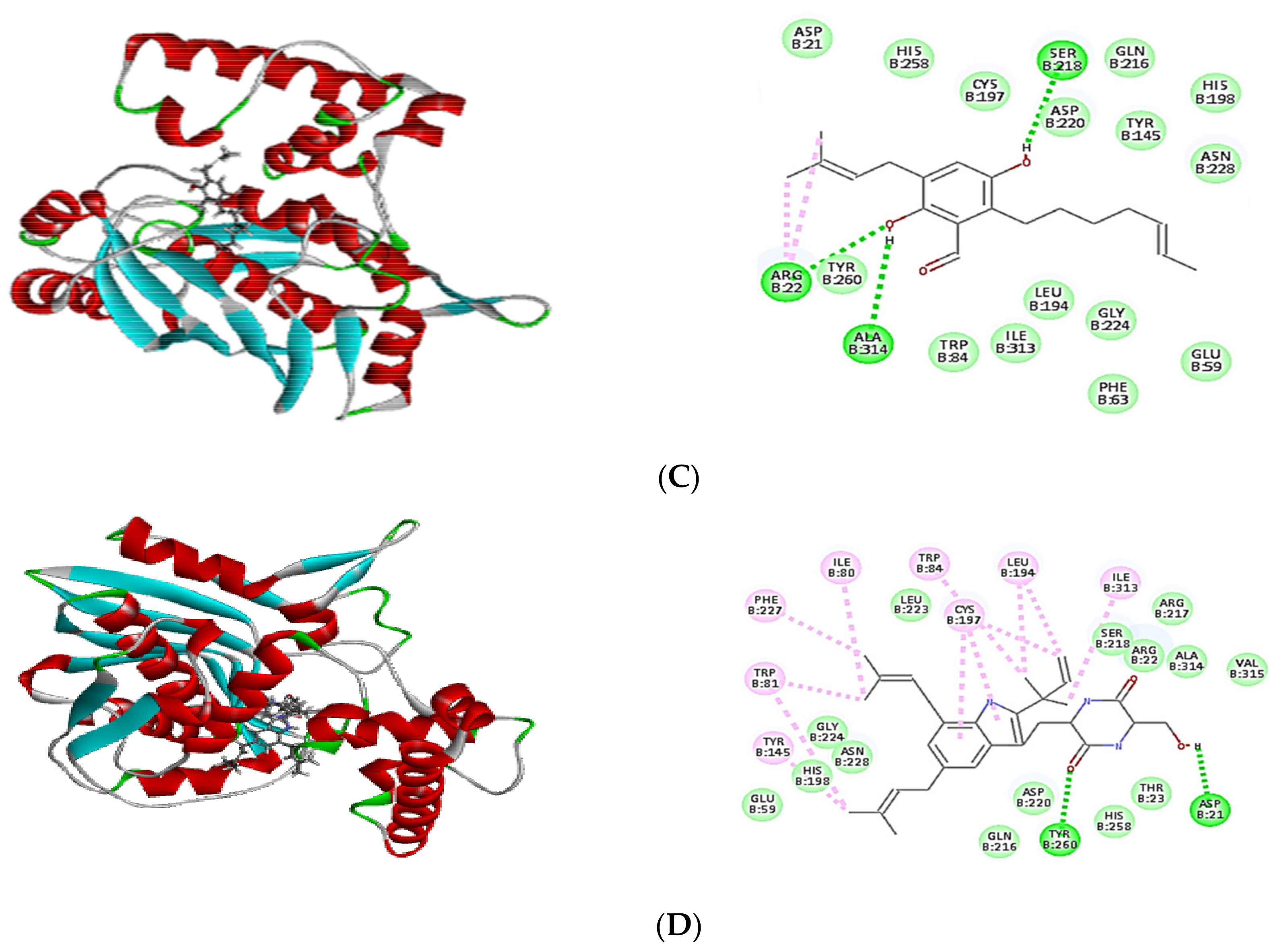
2.4. Molecular Docking of Isolated Compounds
3. Discussion
4. Materials and Methods
4.1. Plant Samples
4.2. Endophytic Fungus Isolation and Identification
4.3. Production and Extraction of the Fungus Metabolites
4.4. Compounds Fractionation and Purification
4.5. Identification of FEA Isolated Compounds
4.6. Screening of Antimicrobial Activity against Resistant Strains Using Total FEA and Its Purified Compounds
4.7. Molecular Docking
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Souza-Motta, C. Biologically Active Echinulin-Related Indolediketopiperazines from the Marine Sediment-Derived Fungus Aspergillus niveoglaucus. World J. Microbiol. Biotechnol. 2012, 28, 1989–1995. [Google Scholar] [CrossRef] [Green Version]
- Kaur, M.; Kaur, A.; Sharma, R. Pharmacological actions of Opuntia ficus indica: A Review. J. Appl. Pharm. Sci. 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Elkady, W.M.; Bishr, M.M.; Abdel-Aziz, M.M.; Salama, O.M. Identification and isolation of anti-pneumonia bioactive compounds from Opuntia ficus-indica fruit waste peels. Food Funct. 2020, 11, 5275–5283. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Chen, Y.; He, K.; Chen, Y.; Ye, Y.; Zhou, M. A New Cyclic Tetrapeptide from Endophytic Fungus Aspergillus versicolor E-2. Rec. Nat. Prod. 2021, 15, 363–367. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, J.; Li, G.; Zheng, Z.; Su, W. Antitumor and antifungal activities in endophytic fungi isolated from pharmaceutical plants Taxus mairei, Cephalataxus fortunei and Torreya grandis. FEMS Immunol. Med. Microbiol. 2001, 31, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted interactions between endophytes and plant: Developments and prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef]
- Corrêa, R.C.; Heleno, S.A.; Alves, M.J.; Ferreira, I.C. Bacterial Resistance: Antibiotics of last generation used in clinical practice and the arise of natural products as new therapeutic alternatives. Curr. Pharm. Des. 2020, 26, 815–837. [Google Scholar] [CrossRef]
- Bassegoda, A.; Ivanova, K.; Ramon, E.; Tzanov, T. Strategies to prevent the occurrence of resistance against antibiotics by using advanced materials. Appl. Microbiol. Biotechnol. 2018, 102, 2075–2089. [Google Scholar] [CrossRef] [Green Version]
- Fathallah, N.; Raafat, M.M.; Issa, M.Y.; Abdel-Aziz, M.M.; Bishr, M.; Abdelkawy, M.A.; Salama, O. Bio-guided fractionation of prenylated benzaldehyde derivatives as potent antimicrobial and antibiofilm from Ammi majus L. fruits-associated Aspergillus amstelodami. Molecules 2019, 24, 4118. [Google Scholar] [CrossRef] [Green Version]
- Miyake, Y.; Ito, C.; Kimura, T.; Suzuki, A.; Nishida, Y.; Itoigawa, M. Isolation of aromatic compounds produced by Eurotium herbariorum NU-2 from karebushi, a katsuobushi, and their DPPH-radical scavenging activities. Food Sci. Technol. Res. 2014, 20, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Elissawy, A.M.; Ebada, S.S.; Ashour, M.L.; El-Neketi, M.; Ebrahim, W.; Singab, A.B. New secondary metabolites from the mangrove-derived fungus Aspergillus sp. AV-2. Phytochem. Lett. 2019, 29, 1–5. [Google Scholar] [CrossRef]
- Palacio-Barrera, A.M.; Areiza, D.; Zapata, P.; Atehortúa, L.; Correa, C.; Peñuela-Vásquez, M. Induction of pigment production through media composition, abiotic and biotic factors in two filamentous fungi. Biotechnol. Rep. 2019, 21, e00308. [Google Scholar] [CrossRef]
- Quilico, A.; Panizzi, L. Chemische Untersuchungen über Aspergillus echinulatus, I. Mitteilung. Berichte der Deutschen Chemischen Gesellschaft (A and B Series) 1943, 76, 348–358. [Google Scholar] [CrossRef]
- Elsebai, M.F.; Schoeder, C.T.; Müller, C.E. Fintiamin: A diketopiperazine from the marine sponge derived fungus Eurotium sp. Arch. Der Pharm. 2021, 354, 2100206. [Google Scholar] [CrossRef] [PubMed]
- Du, F.-Y.; Li, X.-M.; Li, C.-S.; Shang, Z.; Wang, B.-G. Cristatumins A–D, new indole alkaloids from the marine-derived endophytic fungus Eurotium cristatum EN-220. Bioorg. Med. Chem. Lett. 2012, 22, 4650–4653. [Google Scholar] [CrossRef]
- Smetanina, O.; Kalinovskii, A.; Khudyakova, Y.V.; Slinkina, N.; Pivkin, M.; Kuznetsova, T. Metabolites from the marine fungus Eurotium repens. Chem. Nat. Compd. 2007, 43, 395–398. [Google Scholar] [CrossRef]
- Alanis, A.J. Resistance to antibiotics: Are we in the post-antibiotic era? Arch. Med. Res. 2005, 36, 697–705. [Google Scholar] [CrossRef]
- Wright, G.D. Mechanisms of resistance to antibiotics. Curr. Opin. Chem. Biol. 2003, 7, 563–569. [Google Scholar] [CrossRef]
- Al-Shaibani, A.B.A.; Al-Shakarchi, F.I.; Ameen, R.S. Extraction and characterization of antibacterial compound from Aspergillus niger. Al-Nahrain J. Sci. 2013, 16, 167–174. [Google Scholar] [CrossRef]
- Selim, K.; El-Beih, A.; AbdEl-Rahman, T.; El-Diwany, A. Biodiversity and antimicrobial activity of endophytes associated with Egyptian medicinal plants. Mycosphere 2011, 2, 669–678. [Google Scholar] [CrossRef]
- Adebayo, E.A.; Ibikunle, J.B.; Oke, A.M.; Lateef, A.; Azeez, M.A.; Oluwatoyin, A.O.; AyanfeOluwa, A.V.; Blessing, O.T.; Comfort, O.O.; Adekunle, O.O. Antimicrobial and antioxidant activity of silver, gold and silver-gold alloy nanoparticles phytosynthesized using extract of Opuntia ficus-indica. Rev. Adv. Mater. Sci. 2019, 58, 313–326. [Google Scholar] [CrossRef]
- Blando, F.; Russo, R.; Negro, C.; De Bellis, L.; Frassinetti, S. Antimicrobial and antibiofilm activity against Staphylococcus aureus of Opuntia ficus-indica (L.) Mill. cladode polyphenolic extracts. Antioxidants 2019, 8, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratnaweera, P.B.; de Silva, E.D.; Williams, D.E.; Andersen, R.J. Antimicrobial activities of endophytic fungi obtained from the arid zone invasive plant Opuntia dillenii and the isolation of equisetin, from endophytic Fusarium sp. BMC Complement. Altern. Med. 2015, 15, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akinyemi, A. Antimicrobial activities of secondary metabolites from fungal endophytes. IOSR J. Pharm. Biol. Sci. 2017, 12, 13–17. [Google Scholar]
- Al-Fakih, A.A.; Almaqtri, W.Q.A. Overview on antibacterial metabolites from terrestrial Aspergillus spp. Mycology 2019, 10, 191–209. [Google Scholar] [CrossRef] [Green Version]
- Enright, M.C.; Robinson, D.A.; Randle, G.; Feil, E.J.; Grundmann, H.; Spratt, B.G. The evolutionary history of methicillin-resistant Staphylococcus aureus (MRSA). Proc. Natl. Acad. Sci. USA 2002, 99, 7687–7692. [Google Scholar] [CrossRef] [Green Version]
- Murray, B.E. Vancomycin-resistant enterococcal infections. N. Engl. J. Med. 2000, 342, 710–721. [Google Scholar] [CrossRef]
- Netikul, T.; Kiratisin, P. Genetic characterization of carbapenem-resistant enterobacteriaceae and the spread of carbapenem-resistant Klebsiella pneumonia ST340 at a university hospital in Thailand. PLoS ONE 2015, 10, e0139116. [Google Scholar] [CrossRef]
- Poole, K. Pseudomonas aeruginosa: Resistance to the max. Front. Microbiol. 2011, 2, 65. [Google Scholar] [CrossRef] [Green Version]
- Slack, G.J.; Puniani, E.; Frisvad, J.C.; Samson, R.A.; Miller, J.D. Secondary metabolites from Eurotium species, Aspergillus calidoustus and A. insuetus common in Canadian homes with a review of their chemistry and biological activities. Mycol. Res. 2009, 113, 480–490. [Google Scholar] [CrossRef]
- Moni, J.N.R.; Adnan, M.; Tareq, A.M.; Kabir, M.; Reza, A.; Nasrin, M.; Chowdhury, K.H.; Sayem, S.A.J.; Rahman, M.A.; Alam, A. Therapeutic potentials of Syzygium fruticosum fruit (seed) reflect into an array of pharmacological assays and prospective receptors-mediated pathways. Life 2021, 11, 155. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.F.; Kader, F.B.; Arman, M.; Ahmed, S.; Lyzu, C.; Sakib, S.A.; Tanzil, S.M.; Zim, A.I.U.; Imran, M.A.S.; Venneri, T. Pharmacological insights and prediction of lead bioactive isolates of Dita bark through experimental and computer-aided mechanism. Biomed. Pharmacother. 2020, 131, 110774. [Google Scholar] [CrossRef] [PubMed]
- Wink, M.; Schimmer, O. Molecular modes of action of defensive secondary metabolites. Funct. Biotechnol. Plant Second. Metab. 2010, 39, 21–161. [Google Scholar] [CrossRef]
- Hazalin, N.A.; Ramasamy, K.; Lim, S.S.M.; Wahab, I.A.; Cole, A.L.; Abdul Majeed, A.B. Cytotoxic and antibacterial activities of endophytic fungi isolated from plants at the National Park, Pahang, Malaysia. BMC Complement. Altern. Med. 2009, 9, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubka, V.; Kolařík, M.; Kubátová, A.; Peterson, S.W. Taxonomic revision of Eurotium and transfer of species to Aspergillus. Mycologia 2013, 105, 912–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turan, C.; Nanni, I.M.; Brunelli, A.; Collina, M. New rapid DNA extraction method with Chelex from Venturia inaequalis spores. J. Microbiol. Methods 2015, 115, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Diba, K.; Kordbacheh, P.; Mirhendi, S.; Rezaie, S.; Mahmoudi, M. Identification of Aspergillus species using morphological characteristics. Pak. J. Med. Sci. 2007, 23, 867. [Google Scholar]
- Loures, F.v.V.; Levitz, S.M. XTT assay of antifungal activity. Bio-Protocol 2015, 5, e1543. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound 1 | Compound 2 | ||
|---|---|---|---|
| Atom No | δH, 1H-NMR | δC 13C-NMR/APT | δH, 1H-NMR |
| 1 | - | 130.8 C | - |
| 2 | - | 127.5 C | - |
| 3 | - | 145.5 C | - |
| 4 | 7.0 (s), 1H | 125.7 CH | - |
| 5 | - | 135.8 C | 6.36 (m), 1H |
| 6 | 11.75 (s), OH | 158.5 C | - |
| 7 | 10.1 (s) | 196.5 CH | 11.83 (s), 1H |
| 1′ | 6.59 (d, J = 15.8 Hz, 1H) | 140.7 CH | 10.12 (S), 1H |
| 2′ | 6.46 (dd, J = 15.8, 10.1 Hz, 1H) | 132.19 CH | 2.99 (m), 2H |
| 3′ | 6.3 (m), 1H | 125.49 CH | 1.6 (m), 2H |
| 4′ | 5.47 (m), 1H | 119.9 CH | 1.4 (m),2H |
| 5′ | 2.1 (m), 2H | 34.9 CH2 | 1.3 (m), 1H |
| 6′ | 1.3 (m), 2H | 29.9 CH2 | 5.3 (m),1H |
| 7′ | 0.9 (m), 3H | 13.40 CH3 | 5.4 (m), 1H |
| 1″ | 3.34 (d, J = 7.8 Hz, 1H) | 27.1 CH2 | 1.86 (d, J = 1.6 Hz, 1H) |
| 2″ | 5.3 (m), 1H | 121.2 CH | 3.44 (d, J = 7.6 Hz, 1H) |
| 3″ | - | 139.8 CH | 5.88 (m), 1H |
| 4″ | 1.7 (s), 3H | 17.7 CH3 | - |
| 5″ | 1.8 (s), 3H | 25.9 CH3 | 1.75 (s), 3H |
| Compound 3 | |||
|---|---|---|---|
| Atom No | δH, 1H-NMR | Atom No | δH, 1H-NMR |
| 1 | 7.7, br, s, 1NH | 16 | 5.9, m, 1H |
| 2 | - | 17 | 5.1, d, J = 17.4, 2H |
| 3 | - | 18 | 1.2, s, 3H |
| 4 | 7.3, s, 1H | 19 | 1.5, s, 3H |
| 5 | - | 20 | 3.9, m, 2H |
| 6 | 6.7, s, 1H | 21 | 3.39, m, 2H |
| 7 | - | 22 | 5.3, t, 1H |
| 8 | 3.67, s, 2H | 23 | - |
| 9 | 4.4, s, 1H | 24 | 2.1, 2, 3H |
| 10 | - | 25 | 2.0, s, 3H |
| 11 | 6.05, br, s, 1NH | 26 | 3.53, m, 2H |
| 12 | 4.03, s, 1H | 27 | 5.8, t, 1H |
| 13 | - | 28 | - |
| 14 | 5.3, br, s, 1NH | 29 | 1.8, s, 3H |
| 15 | - | 30 | 1.9, s, 3H |
| Compound | MIC (μg/mL) | |||
|---|---|---|---|---|
| Gram-Positive Strains | Gram-Negative Strains | |||
| MRSA ATCC-700788 | VR Enterococcus faecalis ATCC BAA-2365 | CR Klebsiella pneumonia ATCC BAA-2342 | MDR Pseudomonas aeruginosa ATCC-BAA-2111 | |
| C1 | 31.25 | 62.5 | 7.81 | 31.25 |
| C2 | 1.95 | 3.9 | 31.25 | 125 |
| C3 | 7.81 | 15.63 | 1.95 | 3.9 |
| Vancomycin | 1.95 | - | - | - |
| Linezolid | - | 1.95 | - | - |
| Colistin | - | - | 0.48 | 3.9 |
| Compounds | Binding Energy ∆G (kcal/mol) | ||
|---|---|---|---|
| Tyrosyl-tRNA Synthetase | Dihydrofolate Reductase | Thymidylate Synthase | |
| Co-crystalized ligand (629) | −64.82 | - | - |
| Co-crystalized ligand (Q27) | - | −48.63 | - |
| Co-crystalized ligand (FFO) | - | - | −64.12 |
| Dihydroauroglaucin (C1) | −41.59 | −41.77 | −42.16 |
| Isotetrahydroauroglucin (C2) | −44.32 | −42.75 | −43.98 |
| Cristatumin B (C3) | −63.77 | −55.50 | –54.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elkady, W.M.; Raafat, M.M.; Abdel-Aziz, M.M.; AL-Huqail, A.A.; Ashour, M.L.; Fathallah, N. Endophytic Fungus from Opuntia ficus-indica: A Source of Potential Bioactive Antimicrobial Compounds against Multidrug-Resistant Bacteria. Plants 2022, 11, 1070. https://doi.org/10.3390/plants11081070
Elkady WM, Raafat MM, Abdel-Aziz MM, AL-Huqail AA, Ashour ML, Fathallah N. Endophytic Fungus from Opuntia ficus-indica: A Source of Potential Bioactive Antimicrobial Compounds against Multidrug-Resistant Bacteria. Plants. 2022; 11(8):1070. https://doi.org/10.3390/plants11081070
Chicago/Turabian StyleElkady, Wafaa M., Marwa M. Raafat, Marwa M. Abdel-Aziz, Arwa A. AL-Huqail, Mohamed L. Ashour, and Noha Fathallah. 2022. "Endophytic Fungus from Opuntia ficus-indica: A Source of Potential Bioactive Antimicrobial Compounds against Multidrug-Resistant Bacteria" Plants 11, no. 8: 1070. https://doi.org/10.3390/plants11081070
APA StyleElkady, W. M., Raafat, M. M., Abdel-Aziz, M. M., AL-Huqail, A. A., Ashour, M. L., & Fathallah, N. (2022). Endophytic Fungus from Opuntia ficus-indica: A Source of Potential Bioactive Antimicrobial Compounds against Multidrug-Resistant Bacteria. Plants, 11(8), 1070. https://doi.org/10.3390/plants11081070