
Variations in Total Phenolic, Total Flavonoid Contents, and Free Radicals’ Scavenging Potential of Onion Varieties Planted under Diverse Environmental Conditions

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
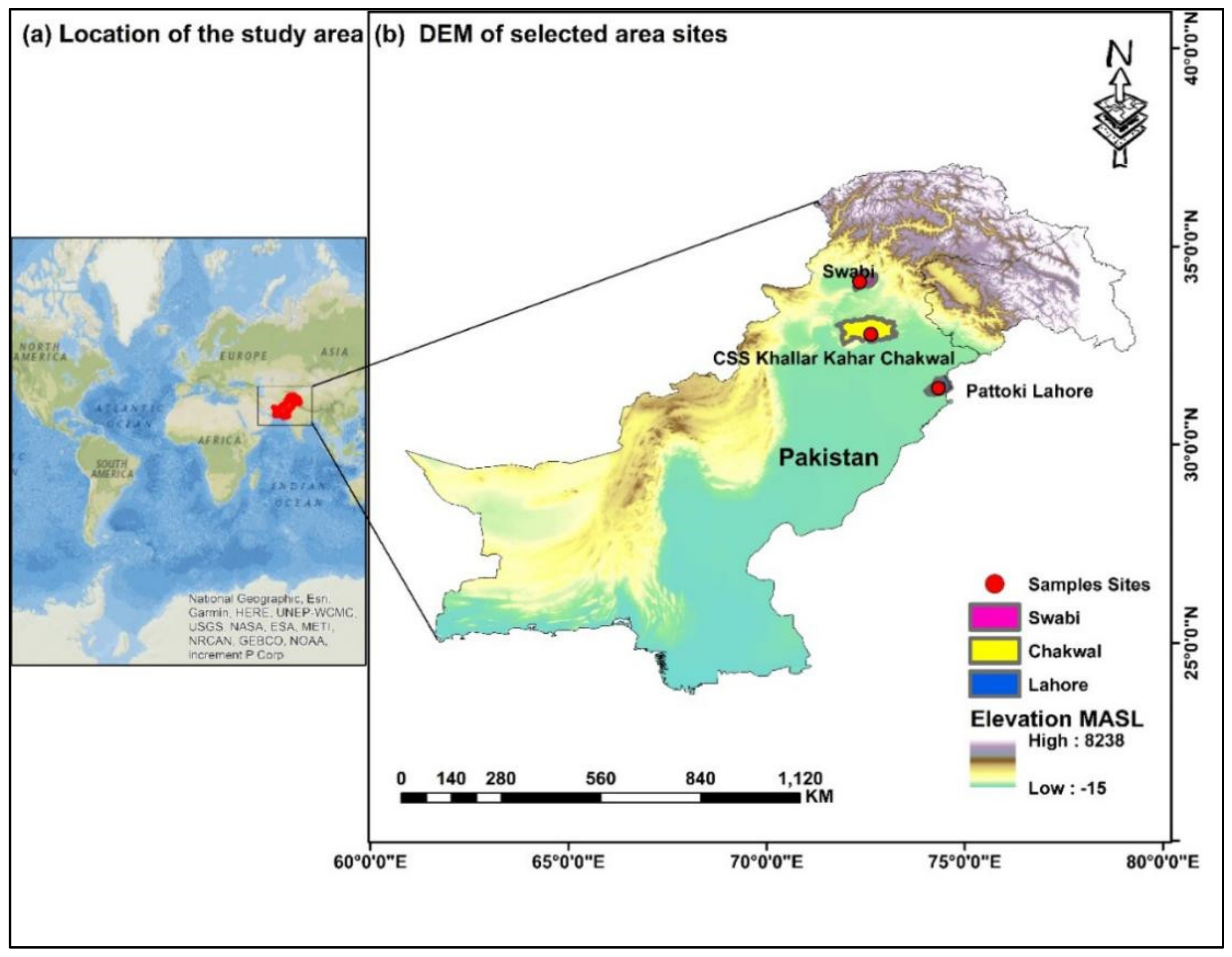
2.1. Sampling Sites, Samples’ Collection and Processing
2.2. Extraction
2.3. Quantification of Total Phenolic and Total Flavonoid Contents
2.4. Free Radicals’ Scavenging Assays
2.4.1. DPPH Assay
2.4.2. Ferric Ion Reducing Antioxidant Power (FRAP) Assay
2.4.3. Phosphomolybdenum Complex Assay (PMA)
2.4.4. ABTS Radical Scavenging Assay
2.4.5. Hydroxyl Radical (OH) Scavenging Activity
2.4.6. Hydrogen Peroxide (H2O2) Scavenging Activity
2.5. Climate Data
2.6. Analysis of Soil Properties
2.7. Statistical Analysis
3. Results and Discussion
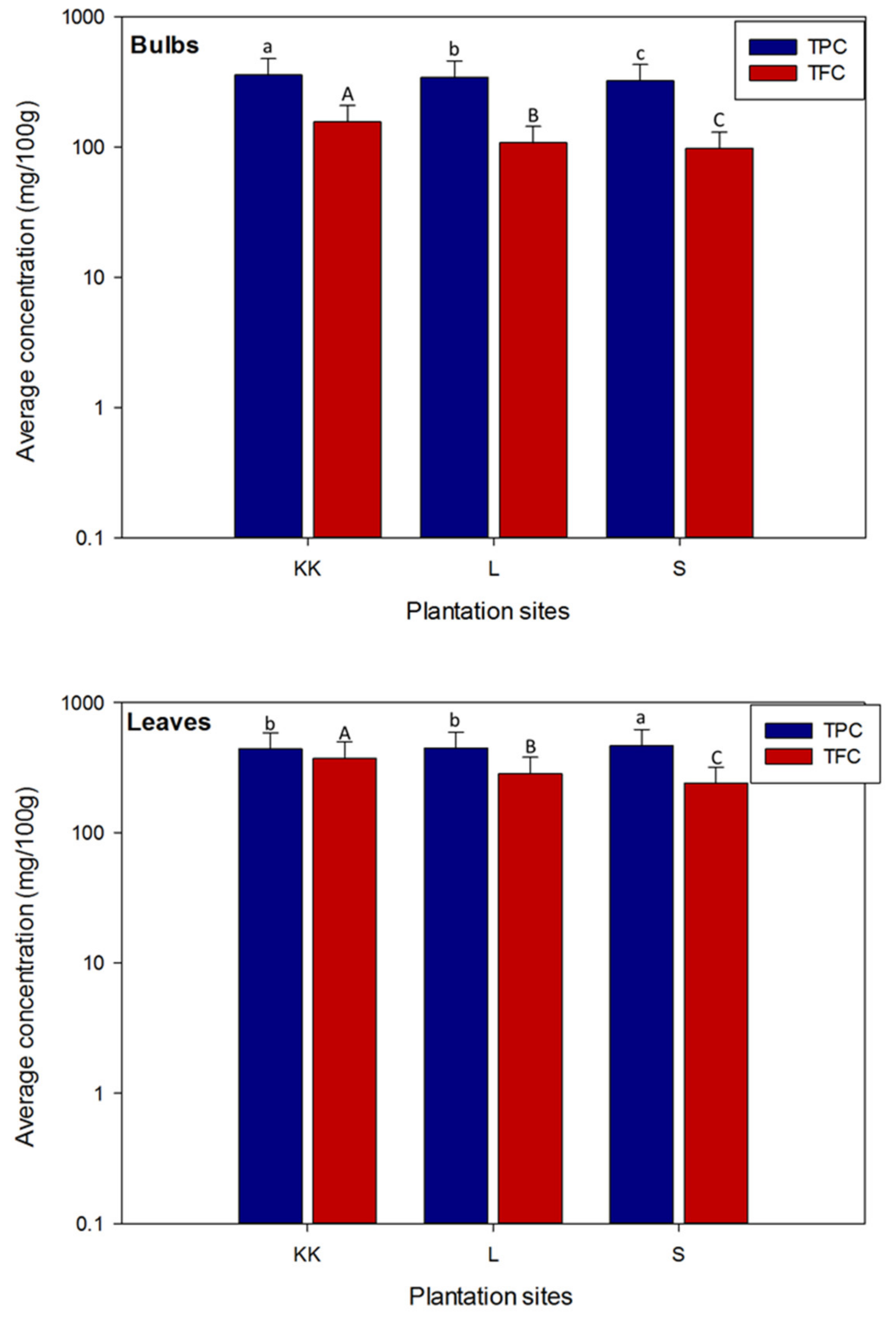
3.1. Variations in the Total Phenolic and Total Flavonoid Contents
3.2. Disparity in Free Radicals’ Scavenging/Antioxidant Potential
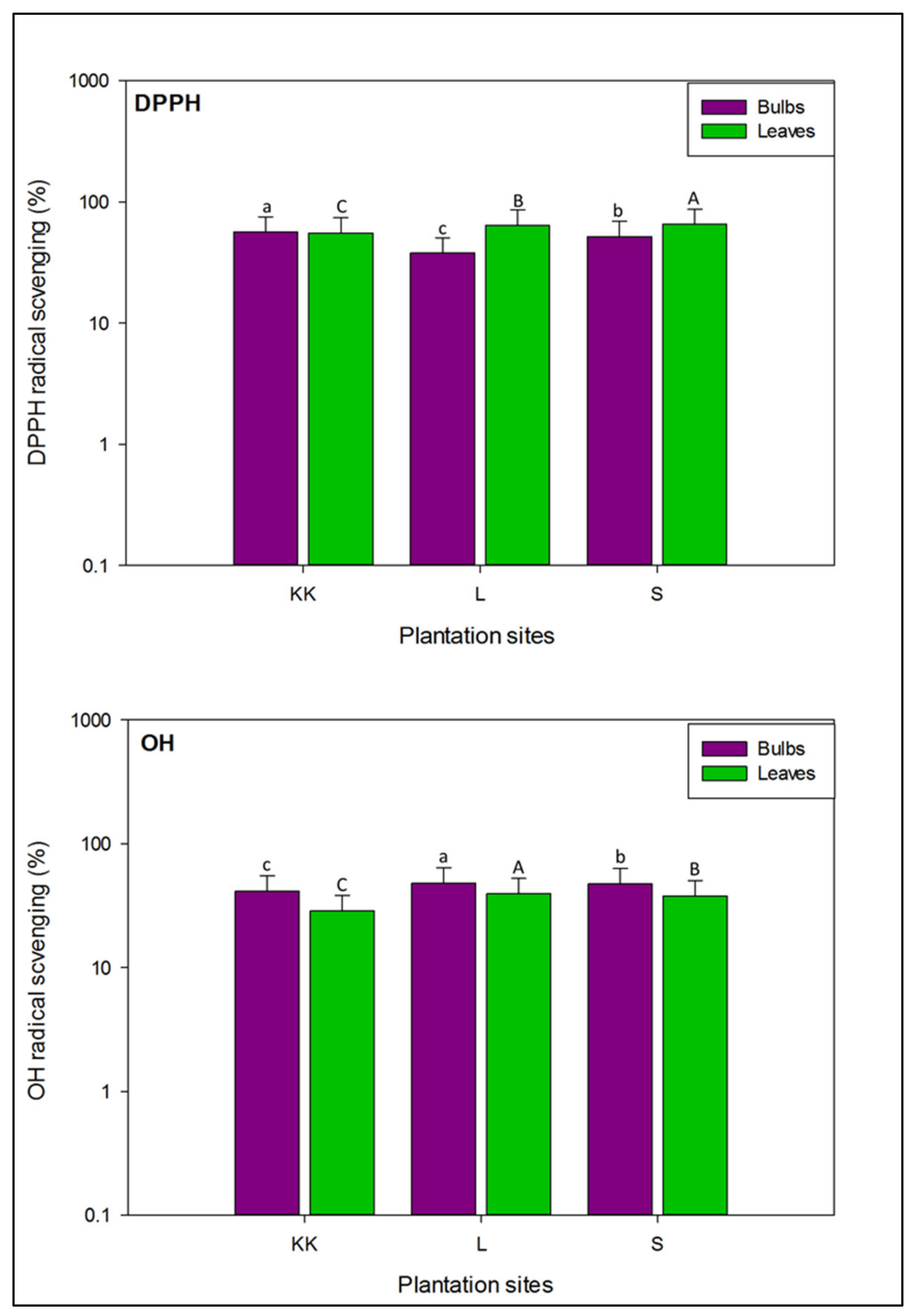
3.2.1. DPPH Radical Inhibition Potential in the Bulbs and Leaves of Onion Varieties
3.2.2. Hydroxyl Radical Scavenging Ability in Onion Varieties
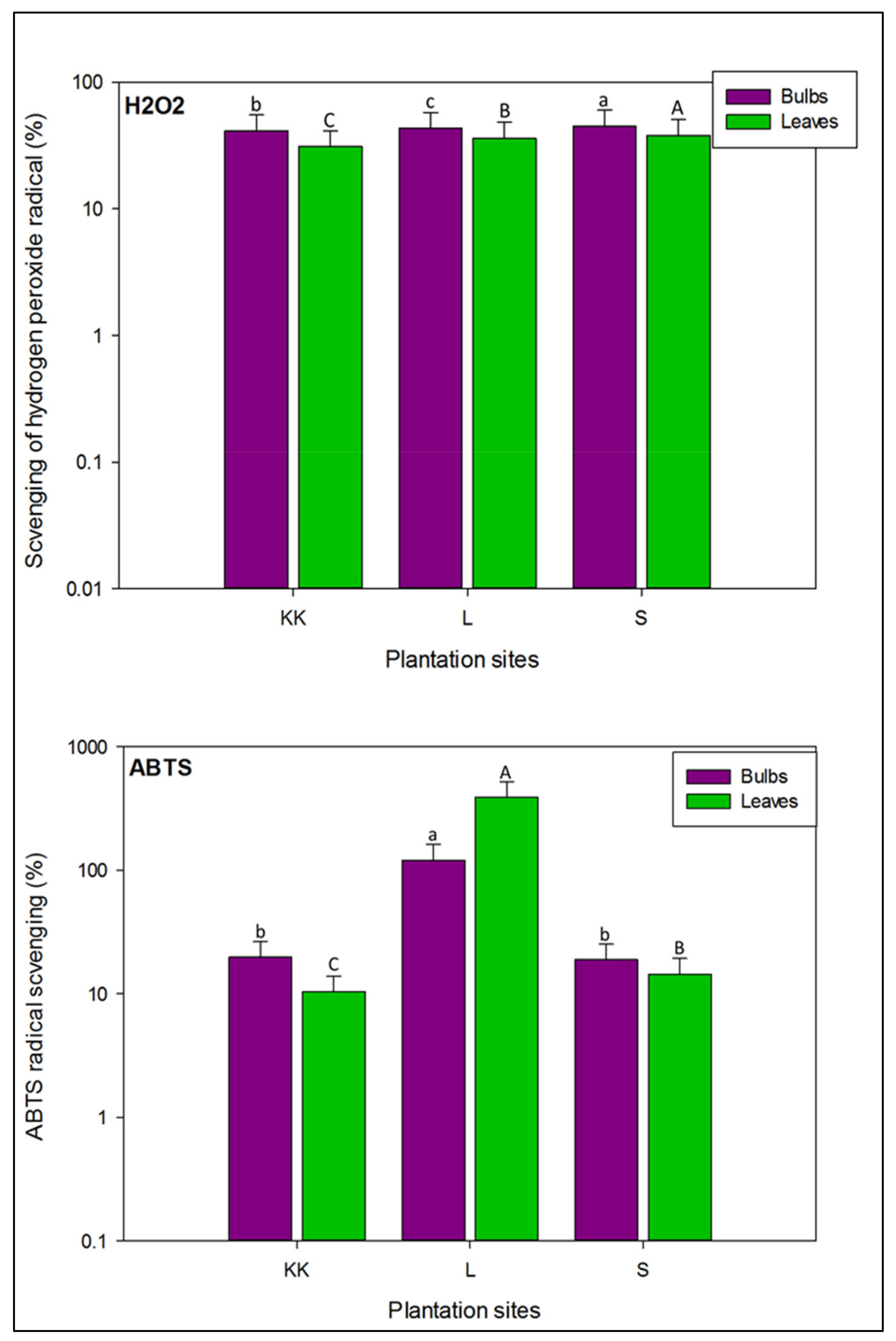
3.2.3. Hydrogen Peroxide Inhibition Capacity
3.2.4. ABTS Radical Scavenging Capacity
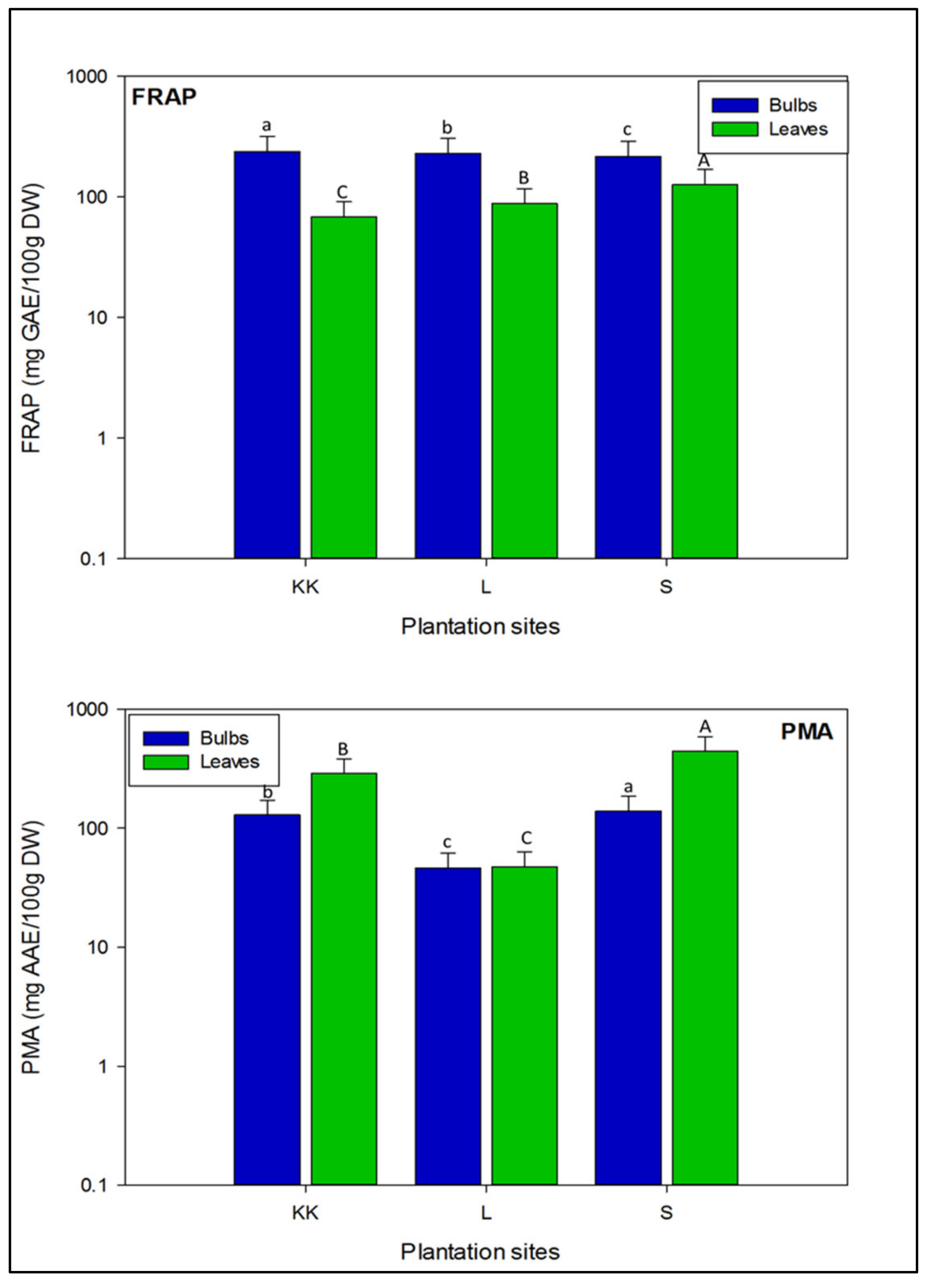
3.2.5. Ferric Ion Reducing Potential in Onion Varieties
3.2.6. Comparative Assessment of Total Antioxidant Capacity
3.3. Correlation Analysis
3.4. Impact of Growing Conditions Secondary Metabolites and Antioxidant Activities
3.4.1. Impact of Climatic Conditions on Total Phenolic, Total Flavonoid Contents and Antioxidant Activities
Impact of Temperature
Impact of Rain Fall and Humidity
Associations of Surface Pressure, Frost Point and Wind Speed
Impact of Altitudinal Variations
3.4.2. Impact of Edaphic Factors on Total Phenolic, Total Flavonoid Contents and Antioxidant Activities
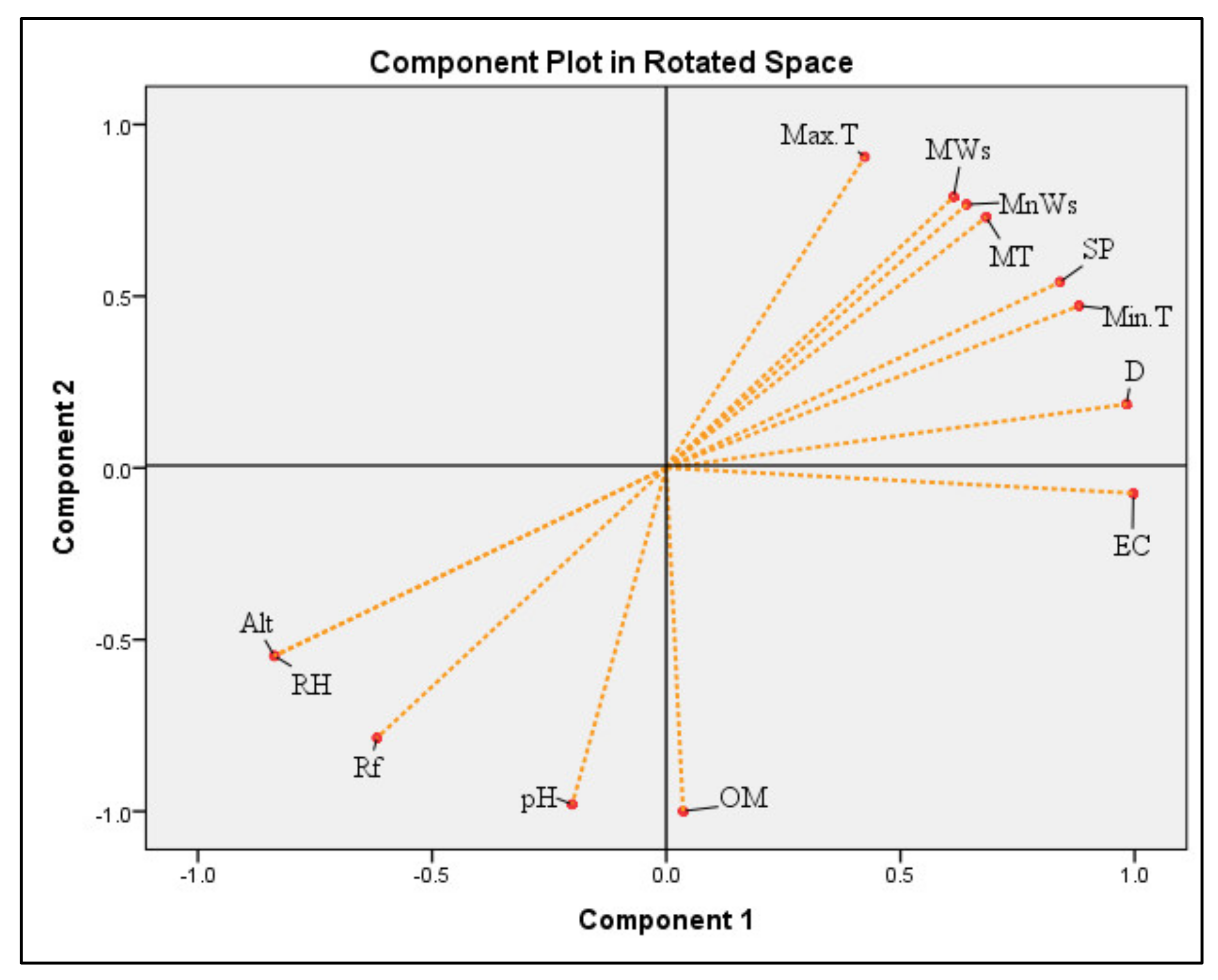
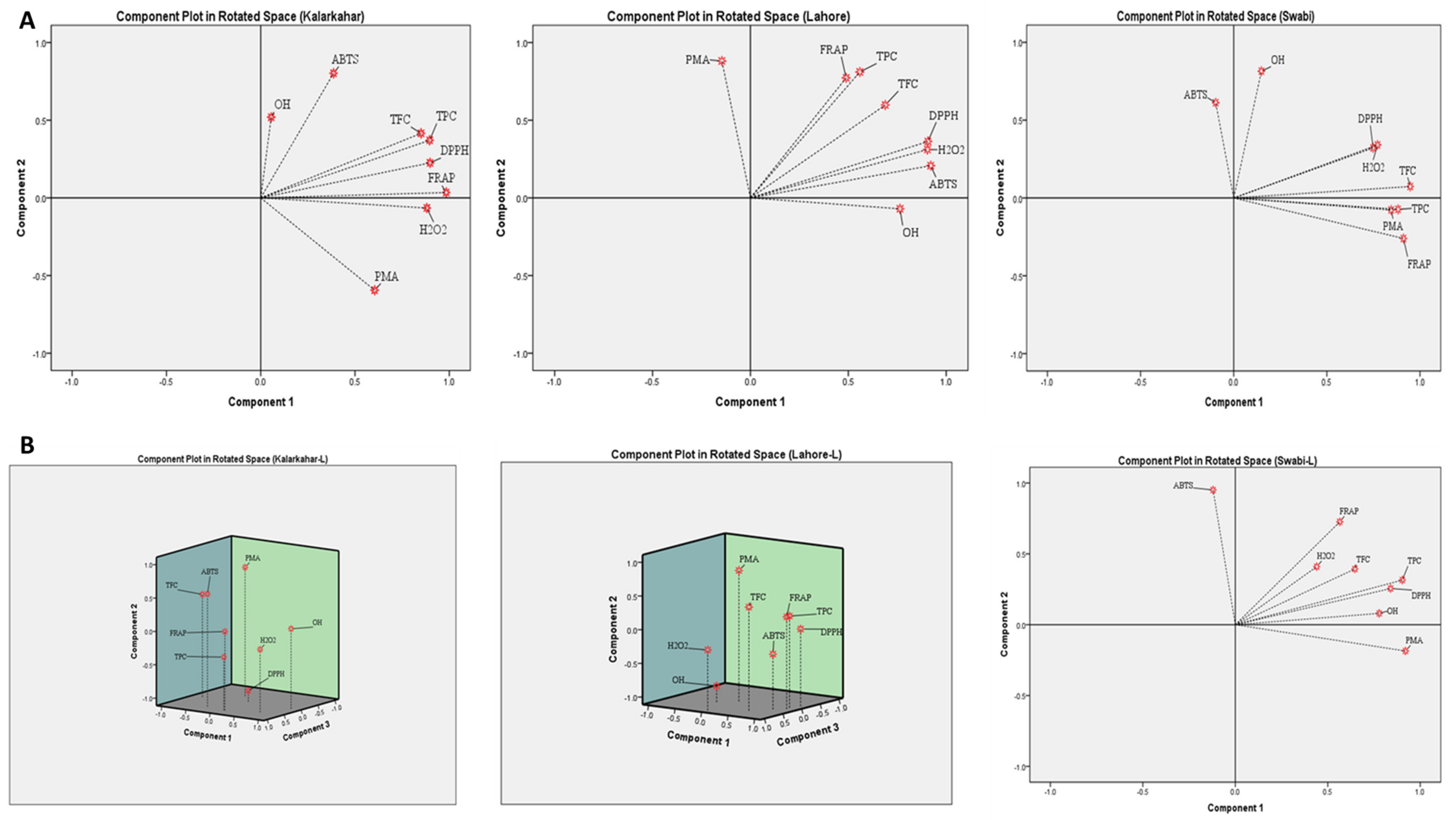
3.5. Principal Component Analysis (PCA)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caleja, C.; Ribeiro, A.; Filomena Barreiro, M.; Ferreira, I. Phenolic compounds as nutraceuticals or functional food ingredients. Curr. Pharm. Des. 2017, 23, 2787–2806. [Google Scholar] [CrossRef] [PubMed]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A concise overview on the chemistry, occurrence, and human health. Phytother. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trchounian, A.; Petrosyan, M.; Sahakyan, N. Plant Cell Redox Homeostasis and Reactive Oxygen Species. In Redox State as a Central Regulator of Plant-Cell Stress Responses; Springer: Cham, Switzerland, 2016; pp. 25–50. [Google Scholar]
- Sytar, O.; Zivcak, M.; Neugart, S.; Toutounchi, P.M.; Brestic, M. Precultivation of young seedlings under different color shades modifies the accumulation of phenolic compounds in Cichorium leaves in later growth phases. Environ. Exp. Bot. 2019, 165, 30–38. [Google Scholar] [CrossRef]
- Teshika, J.D.; Zakariyyah, A.M.; Zaynab, T.; Zengin, G.; Rengasamy, K.R.; Pandian, S.K.; Fawzi, M.M. Traditional and modern uses of onion bulb (Allium cepa L.): A systematic review. Crit. Rev. Food Sci. Nutr. 2019, 59 (Suppl. 1), S39–S70. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2016, 82, 513–523. [Google Scholar] [CrossRef]
- Mathesius, U. Flavonoid Functions in Plants and Their Interactions with Other Organisms. Plants 2018, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Tohidi, B.; Rahimmalek, M.; Arzani, A. Essential oil composition, total phenolic, flavonoid contents, and antioxidant activity of Thymus species collected from different regions of Iran. Food Chem. 2017, 220, 153–161. [Google Scholar] [CrossRef]
- Dudek, B.; Warskulat, A.-C.; Schneider, B. The occurrence of flavonoids and related compounds in flower sections of Papaver nudicaule. Plants 2016, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Zeinali, M.; Rezaee, S.A.; Hosseinzadeh, H. An overview on immunoregulatory and anti-inflammatory properties of chrysin and flavonoids substances. Biomed. Pharmacother. 2017, 92, 998–1009. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredotović, Ž.; Šprung, M.; Soldo, B.; Ljubenkov, I.; Budić-Leto, I.; Bilušić, T.; Puizina, J. Chemical composition and biological activity of Allium cepa L. and Allium × cornutum (Clementi ex Visiani 1842) methanolic extracts. Molecules 2017, 22, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razavi-Azarkhiavi, K.; Behravan, J.; Mosaffa, F.; Sehatbakhsh, S.; Shirani, K.; Karimi, G. Protective effects of aqueous and ethanol extracts of rosemary on H2O2-induced oxidative DNA damage in human lymphocytes by comet assay. J. Complement. Integr. Med. 2014, 11, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Hodaei, M.; Rahimmalek, M.; Arzani, A.; Talebi, M. The effect of water stress on phytochemical accumulation, bioactive compounds and expression of key genes involved in flavonoid biosynthesis in Chrysanthemum morifolium L. Ind. Crop. Prod. 2018, 120, 295–304. [Google Scholar] [CrossRef]
- Angelino, D.; Godos, J.; Ghelfi, F.; Tieri, M.; Titta, L.; Lafranconi, A.; Marventano, S.; Alonzo, E.; Gambera, A.; Sciacca, S.; et al. Fruit and vegetable consumption and health outcomes: An umbrella review of observational studies. Int. J. Food Sci. Nutr. 2019, 70, 652–667. [Google Scholar] [CrossRef]
- Wallace, T.C.; Bailey, R.L.; Blumberg, J.B.; Burton-Freeman, B.; Chen, C.O.; Crowe-White, K.M.; Wang, D.D. Fruits, vegetables, and health: A comprehensive narrative, umbrella review of the science and recommendations for enhanced public policy to improve intake. Crit. Rev. Food Sci. Nutr. 2020, 60, 2174–2211. [Google Scholar] [CrossRef] [Green Version]
- Aguiar, J.; Gonçalves, J.L.; Alves, V.L.; Câmara, J.S. Chemical fingerprint of free polyphenols and antioxidant activity in dietary fruits and vegetables using a non-targeted approach based on QuEChERS ultrasound-assisted extraction combined with UHPLC-PDA. Antioxidants 2020, 9, 305. [Google Scholar] [CrossRef] [Green Version]
- Mollavali, M.; Bolandnazar, S.A.; Schwarz, D.; Rohn, S.; Riehle, P.; ZaareNahandi, F. Flavonol glucoside and antioxidant enzyme biosynthesis affected by mycorrhizal fungi in various cultivars of onion (Allium cepa L.). J. Agric. Food Chem. 2016, 64, 71–77. [Google Scholar] [CrossRef]
- Barla, S.; Upasani, R. Study on Different Methods of Weed Management in Onion (Allium cepa L.). Curr. J. Appl. Sci. Technol. 2019, 33, 1–7. [Google Scholar] [CrossRef]
- Rady, M.O.; Semida, W.M.; Abd El-Mageed, T.A.; Hemida, K.A.; Rady, M.M. Up-regulation of antioxidative defense systems by glycine betaine foliar application in onion plants confer tolerance to salinity stress. Sci. Hortic. 2018, 240, 614–622. [Google Scholar] [CrossRef]
- Sagar, N.A.; Pareek, S.; Gonzalez-Aguilar, G.A. Quantification of flavonoids, total phenols and antioxidant properties of onion skin: A comparative study of fifteen Indian cultivars. J. Food Sci. Technol. 2020, 57, 2423–2432. [Google Scholar] [CrossRef] [PubMed]
- Munir, M.T.; Kheirkhah, H.; Baroutian, S.; Quek, S.Y.; Young, B.R. Subcritical water extraction of bioactive compounds from waste onion skin. J. Clean. Prod. 2018, 183, 487–494. [Google Scholar] [CrossRef]
- Manohar, C.M.; Xue, J.; Murayyan, A.; Neethirajan, S.; Shi, J. Antioxidant activity of polyphenols from Ontario grown onion varieties using pressurized low polarity water technology. J. Funct. Foods 2017, 31, 52–62. [Google Scholar] [CrossRef]
- Sami, R.; Bushnaq, T.; Radhi, K.; Benajiba, N.; Helal, M. Prevalence of thinness cases and dietary diversity among learners of various education stages in Taif Region, Saudi Arabia. Afr. J. Food Agric. Nutr. Dev. 2021, 20, 17081–17094. [Google Scholar] [CrossRef]
- Perez-Gregorio, R.M.; García-Falcón, M.S.; Símal-Gándara, J.; Rodrigues, A.S.; Almeida-Domingos, P.F. Identification and quantification of flavonoids in traditional cultivars of red and white onions at harvest. J. Food Compos. Anal. 2010, 23, 592–598. [Google Scholar] [CrossRef]
- Singh, B.N.; Singh, B.R.; Singh, R.L.; Prakash, D.; Singh, D.P.; Sarma, B.K.; Upadhyay, G.; Singh, H.B. Polyphenolics from various extracts/fraction of red onion (Allium cepa) peel with potential antioxidant and antimutagenic activities. Food Chem. Toxicol. 2009, 47, 1161–1167. [Google Scholar] [CrossRef]
- Marotti, M.; Piccaglia, R. Characterization of Flavonoids in Different Cultivars of Onion (Allium cepa L.). J. Food Sci. 2002, 67, 1229–1232. [Google Scholar] [CrossRef]
- Sellappan, S.; Akoh, C.C. Flavonoids and Antioxidant Capacity of Georgia-Grown Vidalia Onions. J. Agric. Food Chem. 2002, 50, 5338–5342. [Google Scholar] [CrossRef]
- Slimestad, R.; Fossen, T.; Vågen, I.M. Onions: A Source of Unique Dietary Flavonoids. J. Agric. Food Chem. 2007, 55, 10067–10080. [Google Scholar] [CrossRef]
- Bora, K.S.S.; Sharma, A. Phytoconstituents and therapeutic potential of Allium cepa Linn.—A review. Pharmacogn. Rev. 2009, 3, 170–180. [Google Scholar]
- Ignat, I.; Volf, I.; Popa, V.I. A critical review of methods for characterization of polyphenolic compounds in fruits and vegetables. Food Chem. 2011, 126, 1821–1835. [Google Scholar] [CrossRef] [PubMed]
- Bystrická, J.; Musilová, J.; Vollmannová, A.; Timoracká, M.; Kavalcová, P. Bioactive components of onion (Allium cepa L.)—A review. Acta Aliment. 2013, 42, 11–22. [Google Scholar] [CrossRef]
- El-Hadidy, E.M.; Mossa, M.E.A.; Habashy, H.N. Effect of freezing on the pungency and antioxidants activity in leaves and bulbs of greenonion in Giza 6 and Photon varieties. Ann. Agric. Sci. 2014, 59, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Pourzand, A.; Tajaddini, A.; Pirouzpanah, S.; Asghari-Jafarabadi, M.; Samadi, N.; Ostadrahimi, A.R.; Sanaat, Z. Associations between dietary allium vegetables and risk of breast cancer: A hospital-based matched case-control study. J. Breast Cancer 2016, 19, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Bahadoran, Z.; Mirmiran, P.; Momenan, A.A.; Azizi, F. Allium vegetable intakes and the incidence of cardiovascular disease, hypertension, chronic kidney disease, and type 2 diabetes in adults: A longitudinal follow-up study. J. Hypertens. 2017, 35, 1909–1916. [Google Scholar] [CrossRef] [PubMed]
- Santhosha, S.G.; Jamuna, P.; Prabhavathi, S.N. Bioactive components of garlicand their physiological role in health maintenance: A review. Food Biosci. 2013, 3, 59–74. [Google Scholar] [CrossRef]
- Suleria, H.A.R.; Butt, M.S.; Khalid, N.; Sultan, S.; Raza, A.; Aleem, M.; Abbas, M. Garlic (Allium sativum): Diet based therapy of 21st century—A review. Asian Pac. J. Trop. Dis. 2015, 5, 271–278. [Google Scholar] [CrossRef]
- Zargoosh, Z.; Ghavam, M.; Bacchetta, G.; Tavili, A. Effects of ecological factors on the antioxidant potential and total phenol content of Scrophularia striata Boiss. Sci. Rep. 2019, 9, 16021. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Liu, J.; Yin, D.; Zhao, X. Influence of ecological factors on the production of active substances in the anti-cancer plant Sinopodophyllum hexandrum (Royle) T.S. Ying. PLoS ONE 2015, 10, e0122981. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.E.; Ma, X.H.; Wei, Q.; Peng, S.B.; Zhang, S.C. Effects of growing location on the contents of secondary metabolites in the leaves of four selected superior clonesof Eucommia ulmoides. Ind. Crops Prod. 2011, 34, 1607–1614. [Google Scholar] [CrossRef]
- Bilger, W.; Rolland, M.; Nybakken, L. UV screening in higher plants induced by low temperature in the absence of UV-B radiation. Photochem. Photobiol. Sci. 2007, 6, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, K.; Ghasemi, Y.; Ehteshamnia, A.; Nabavi, S.M.; Nabavi, S.F.; Ebrahimzadeh, M.A.; Pourmorad, F. Influence of environmental factors on antioxidant activity, phenol and flavonoids contents of walnut (Juglans regia L.) green husks. J. Med. Plants Res. 2011, 5, 1128–1133. [Google Scholar]
- Mahmood, A.; Malik, R.N. Human health risk assessment of heavy metals via consumption of contaminated vegetables collected from different irrigation sources in Lahore, Pakistan. Arab. J. Chem. 2014, 7, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Jamal, A.; Sarim, M. Heavy metals distribution in different soil series of district Swabi, Khyber Pakhtunkhwa, Pakistan. World Sci. News 2018, 105, 1–13. [Google Scholar]
- Abbasi, A.M.; Guo, X.; Fu, X.; Zhou, L.; Chen, Y.; Zhu, Y.; Yan, H.; Liu, R.H. Comparative Assessment of Phenolic Content and in Vitro Antioxidant Capacity in the Pulp and Peel of Mango Cultivars. Int. J. Mol. Sci. 2015, 16, 13507–13527. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.S.; Tsai, P.H.; Kandaswami, C.C.; Cheng, C.H.; Ke, F.C.; Lee, P.P.; Hwang, J.J.; Lee, M.T. Effects of dietary flavonoids, luteolin, and quercetin on the reversal of epithelial–mesenchymal transition in A431 epidermal cancer cells. Cancer Sci. 2011, 102, 1829–1839. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, R.; Liu, C.; Zheng, X.; Liu, B. Enhancing antioxidant activity and antiproliferation of wheat bran through steam flash explosion. J. Food Sci. Technol. 2016, 53, 3028–3034. [Google Scholar] [CrossRef] [Green Version]
- Hazra, B.; Biswas, S.; Mandal, N. Antioxidant and free radical scavenging activity of Spondiaspinnata. BMC Complementary Altern. Med. 2008, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric Quantitation of Antioxidant Capacity through the Formation of a Phosphomolybdenum Complex: Specific Application to the Determination of Vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef]
- Karthiga, K.; Sivakumar, T. Antioxidant activity of three basidiomycete by ABTS+ radical cation decolourization assay method. Asian J. Chem. 2010, 22, 1647–1648. [Google Scholar]
- Wenli, Y.; Yaping, Z.; Bo, S. The radical scavenging activities of radix puerariaeisoflavonoids: A chemiluminescence study. Food Chem. 2004, 86, 525–529. [Google Scholar] [CrossRef]
- Aeyigoro, O.A.; Okoh, A.I. Preliminary phytochemical screening and in vitro antioxidant activities of the aqueous extract of Helichrysumlongifolium DC. BMC Complementary Altern. Med. 2010, 10, 21. [Google Scholar]
- Liu, W.; Yin, D.; Li, N.; Hou, X.; Wang, D.; Li, D.; Liu, J. Influence of environmental factors on the active substance production and antioxidant activity in Potentilla fruticosa L. and its quality assessment. Sci. Rep. 2016, 6, 28591. [Google Scholar] [CrossRef] [PubMed]
- Demirezen, D.; Aksoy, A. Heavy metal levels in vegetables in Turkey are within safe limits for Cu, Zn, Ni andexceeded for Cd and Pb. J. Food Qual. 2006, 29, 252–265. [Google Scholar] [CrossRef]
- Wang, Y.; Qiao, M.; Liu, Y.; Zhu, Y. Health risk assessment of heavy metals in soils and vegetables from wastewater irrigated area, Beijing-Tianjin city cluster, China. J. Environ. Sci. 2012, 24, 690–698. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L. Total carbon, organic carbon, and organic matter. Methods Soil Anal. Part 2 Chem. Microbiol. Prop. 1983, 9, 539–579. [Google Scholar]
- Downey, M.O.; Dokoozlian, N.K.; Krstic, M.P. Cultural practice and environmental impacts on the flavonoid composition of grapes and wine: A review of recent research. Am. J. Enol. Vitic. 2006, 57, 57–268. [Google Scholar]
- Orduña, R. Climate change associated effects on grape and wine quality and production. Food Res. Int. 2010, 43, 1844–1855. [Google Scholar] [CrossRef]
- Sharma, K.; Assefa, A.D.; Kim, S.; Ko, E.Y.; Lee, E.T.; Park, S.W. Evaluation of total phenolics, flavonoids and antioxidant activity of 18 Korean onion cultivars: A comparative study. J. Sci. Food Agric. 2014, 94, 1521–1529. [Google Scholar]
- Santas, J.; Carbo, R.; Gordon, M.H.; Almajano, M.P. Comparison of the antioxidant activity of two Spanish onion varieties. Food Chem. 2008, 107, 1210–1216. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, C.; Feng, B.; Chen, B.; Jin, L.; Shen, Y. UPLC-ESI-MS/MS based identification and antioxidant, antibacterial, cytotoxic activities of aqueous extracts from storey onion (Allium cepa L. var. proliferum Regel). Food Res. Int. 2020, 130, 108969. [Google Scholar] [CrossRef] [PubMed]
- Bernaert, N.; Paepe, D.D.; Bouten, C.; Clercq, H.D.; Stewart, D.; Bockstaele, E.V.; Loose, M.D.; Droogenbroeck, B.V. Antioxidant capacity, total phenolic and ascorbate content as a function of the genetic diversity of leek (Allium ampeloprasum var. porrum). Food Chem. 2012, 34, 669–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, M.N.; Bristi, N.J.; Rafiquzzaman, M. Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm. J. 2013, 21, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Sadeer, N.B.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The Versatility of Antioxidant Assays in Food Science and Safety—Chemistry, Applications, Strengths, and Limitations—Review. Antioxidants 2020, 9, 709. [Google Scholar] [CrossRef]
- Munteanu, I.G.; Apetrei, C. Analytical Methods Used in Determining Antioxidant Activity: A Review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]
- Apak, R.; Capanoglu, E.; Shahidi, F. Measurement of Antioxidant Activity & Capacity: Recent Trends and Applications, 1st ed.; Wiley: Chichester, UK, 2018. [Google Scholar] [CrossRef]
- Lopez-Alarcon, C.; Denicola, A. Evaluating the antioxidant capacity of natural products: A review on chemical and cellular-based assays. Anal. Chim. Acta 2013, 763, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Granato, D.; Shahidi, F.; Wrolstad, R.; Kilmartin, P.; Melton, L.D.; Hidalgo, F.J.; Miyashita, K.; van Camp, J.; Alasalvar, C.; Ismail, A.B.; et al. Antioxidant activity, total phenolics and flavonoids contents: Should we ban in vitro screening methods? Food Chem. 2018, 264, 471–475. [Google Scholar] [CrossRef]
- Yi, B.; Kim, M.J.; Lee, J. Antioxidant Properties of Astaxanthin in Oil-in-Water Emulsions with Differently-Charged Emulsifiers under Chlorophyll Photosensitization. J. Food Sci. 2018, 83, 589–596. [Google Scholar] [CrossRef]
- Lv, H.-P.; Zhang, Y.; Shi, J.; Lin, Z. Phytochemical profiles and antioxidant activities of Chinese dark teas obtained by different processing technologies. Food Res. Int. 2017, 100, 486–493. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized Methods for the Determination of Antioxidant Capacity and Phenolics in Foods and Dietary Supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Ola–Mudathir, F.K.; Wahab, A.A.; Moshood, A.I.; Obuotor, E.M. Comparative evaluation of antioxidant properties of methanolic extract of Allium cepa bulb, Allium cepa bulb peels and Allium fistulosum. Kragujev. J. Sci. 2018, 40, 131–141. [Google Scholar] [CrossRef]
- Cheng, A.; Chen, X.; Jin, Q.; Wang, W.; Shi, J.; Liu, Y. Comparison of phenolic content and antioxidant capacity of red and yellow onions. Czech J. Food Sci. 2013, 31, 501–508. [Google Scholar] [CrossRef]
- Issa, M.; Karabet, F.; Aljoubbeh, M. Total polyphenols, flavonoid content, kaempferol concentration and antioxidant activity of two onion Syrian (spring and white). Int. J. Chemtech. Res. 2013, 5, 2375–2380. [Google Scholar]
- Dontha, S. A review on antioxidant methods. Asian J. Pharm. Clin. Res. 2016, 9, 14–32. [Google Scholar]
- Moharram, H.; Youssef, M. Methods for determining the antioxidant activity: A review. Alex. J. Food Sci. Technol. 2014, 11, 31–42. [Google Scholar]
- Číž, M.; Čížová, H.; Denev, P.; Kratchanova, M.; Slavov, A.; Lojek, A. Different methods for control and comparison of the antioxidant properties of vegetables. Food Control 2010, 21, 518–523. [Google Scholar] [CrossRef]
- Benkeblia, N. Free-radical scavenging capacity and antioxidant properties of some selected onions (Allium cepa L.) and garlic (Allium sativum L.) extracts. Braz. Arch. Biol. Technol. 2005, 48, 753–759. [Google Scholar] [CrossRef]
- Zheng, L.; Zhao, M.M.; Xiao, C.Q.; Zhao, Q.Z.; Su, G.W. Practical problems when using ABTS assay to assess the radical-scavenging activity of peptides: Importance of controlling reaction pH and time. Food Chem. 2016, 192, 288–294. [Google Scholar] [CrossRef]
- Walker, R.B.; Everette, J.D. Comparative Reaction Rates of Various Antioxidants with ABTS Radical Cation. J. Agric. Food Chem. 2009, 57, 1156–1161. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free. Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Berker, K.I.; Guclu, K.; Tor, I.; Apak, R. Comparative evaluation of Fe(III) reducing power-based antioxidant capacity assays in the presence of phenanthroline, batho-phenanthroline, tripyridyltriazine (FRAP), and ferricyanide reagents. Talanta 2007, 72, 1157–1165. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Ann. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Saritha, V.; Anilakumar, K.R.; Khanum, F. Antioxidant and antibacterial activity of Aloe vera gel extracts. Int. J. Pharm. Biol. Arch. 2010, 1, 376–384. [Google Scholar]
- García-Macías, P.; Ordidge, M.; Vysini, E.; Waroonphan, S.; Battey, N.H.; Gordon, M.H.; Hadley, P.; John, P.; Lovegrove, J.A.; Wagstaffe, A. Changes in the Flavonoid and Phenolic Acid Contents and Antioxidant Activity of Red Leaf Lettuce (Lollo Rosso) Due to Cultivation under Plastic Films Varying in Ultraviolet Transparency. J. Agric. Food Chem. 2007, 55, 10168–10172. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Perret, J.; Harris, M.; Wilson, J.; Haley, S. Antioxidant Properties of Bran Extracts from “Akron” Wheat Grown at Different Locations. J. Agric. Food Chem. 2003, 51, 1566–1570. [Google Scholar] [CrossRef]
- Wang, S.Y.; Zheng, W. Effect of plant growth temperature on antioxidant capacity in strawberry. J. Agric. Food Chem. 2001, 49, 4977–4982. [Google Scholar] [CrossRef]
- Soumaya, K.; Chaouachi, F.; Ksouri, R.; El Gazzah, M. Polyphenolic composition in different organs of Tunisia populations of Cynara cardunculus L. and their antioxidant activity. J. Food Nutr. Res. 2013, 1, 1–6. [Google Scholar]
- Kabtni, S.; Sdouga, D.; Rebey, I.B.; Save, M.; Trifi-Farah, N.; Fauconnier, M.-L.; Marghali, S. Influence of climate variation on phenolic composition and antioxidant capacity of Medicago minima populations. Sci. Rep. 2020, 10, 8293. [Google Scholar] [CrossRef]
- Kaspar, S.; Matros, A.; Mock, H.-P. Proteome and Flavonoid Analysis Reveals Distinct Responses of Epidermal Tissue and Whole Leaves upon UV−B Radiation of Barley (Hordeum vulgare L.) Seedlings. J. Proteome Res. 2010, 9, 2402–2411. [Google Scholar] [CrossRef]
- Jordan, B.R.; James, P.E.; Mackerness, S.A.H. Factors affecting UV-B-induced changes in Arabidopsis thaliana L. gene expression: The role of development, protective pigments and the chloroplast signal. Plant Cell Physiol. 1998, 39, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Lake, J.A.; Field, K.J.; Davey, M.P.; Beerling, D.J.; Lomax, B.H. Metabolomic and physiological responses reveal multi-phasic acclimation of Arabidopsis thaliana to chronic UV radiation. Plant Cell Environ. 2009, 32, 1377–1389. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, H.; Zhang, B.; Deng, Z. The synergistic and antagonistic antioxidant interactions of dietary phytochemical combinations. Crit. Rev. Food Sci. Nutr. 2021, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Gargallo-Garriga, A.; Preece, C.; Sardans, J.; Oravec, M.; Urban, O.; Penuelas, J. Root exudate metabolomes change under drought and show limited capacity for recovery. Sci. Rep. 2018, 8, 12696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahuja, I.; De Vos, R.C.; Bones, A.M.; Hall, R.D. Plant molecular stress responses face climate change. Trends Plant Sci. 2010, 15, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Weidner, S.; Kordala, E.; Brosowska-Arendt, W.; Karamać, M.; Kosińska, A.; Amarowicz, R. Phenolic compounds and properties of antioxidants in grapevine roots (Vitis vinifera L.) under low-temperature stress followed by recovery. Acta Soc. Bot. Pol. 2009, 78, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Karakaya, S.; Bingol, Z.; Koca, M.; Dagoglu, S.; Pınar, N.M.; Demirci, B.; Gulcin, I.; Brestic, M.; Sytar, O. Identification of non-alkaloid natural compound of Angelica purpurascans (Apiaceae) Angelica purpurascens (Ava-Lall.) Gilli. (Apiaceae) with cholinesterase and carbonic anhydrase inhibition potential. Saudi Pharm. J. 2020, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yonekura-Sakakibara, K.; Higashi, Y.; Nakabayashi, R. The Origin and Evolution of Plant Flavonoid Metabolism. Front. Plant Sci. 2019, 10, 943. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Suleria, H.A.R.; Barrow, C.J.; Dunshea, F.R. Screening and Characterization of Phenolic Compounds and Their Antioxidant Capacity in Different Fruit Peels. Foods 2020, 9, 1206. [Google Scholar] [CrossRef]
- Peñuelas, J.; Llusià, J. Effects of carbon dioxide, water supply, and seasonally on tershakene content and emission by Rosmarinus officinalis. J. Chem. Ecol. 1997, 23, 979–993. [Google Scholar] [CrossRef] [Green Version]
- Dastoor, R.; Bakhshi, D.; Ali Akbar, A.R. Evaluation and comparison of total phenol, total flavonoid, resveratrol and antioxidant capacity in fruits of the species Vitis vinifera, (Pistacia vera), (Sambucus nigra) and (Ilex spinigera). Eco-Phytochem. J. Med. Plants 2017, 5, 37–48. [Google Scholar]
- Kavalcová, P.; Bystrická, J.; Tóth, T.; Trebichalský, P.; Hrstková, M.; Lenková, M.; Šiatkovský, O. Content of total polyphenols and antioxidant activity in selected varieties of onion (Allium cepa L.). Potravinarstvo 2015, 9, 494–500. [Google Scholar] [CrossRef] [Green Version]
- Holopainen, J.K.; Virjamo, V.; Ghimire, R.P.; Blande, J.D.; Julkunen-Tiitto, R.; Kivimäenpää, M. Climate change effects on secondary compounds of forest trees in the northern hemisphere. Front. Plant Sci. 2018, 9, 1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.S.; Deng, J.S.; Chen, H.J.; Lin, Y.H.; Huang, G.J. Antioxidant activities of two metallothionein-like proteins from sweet potato (Ipomoea batatas [L.] Lam. ‘Tainong 57′) storage roots and their synthesized peptides. Bot. Stud. 2014, 55, 64–73. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G.; Scheffer, J.J. Factors affecting secondary metabolite production in plants: Volatile components and essential oils. Flavour Fragr. J. 2008, 23, 213–226. [Google Scholar] [CrossRef]
- Dolkar, P.; Dolkar, D.; Angmo, S.; Kumar, B.; Stobdan, T. Variability in phenolics, flavonoids and antioxidants in seabuckthorn (Hippophaerhamnoides L.) seed from nine trans-Himalayan natural population. J. Berry Res. 2017, 7, 109–116. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, G.; Zhao, P.; Zhou, Q.; Zhao, X. The abundance of certain metabolites responds to drought stress in the highly drought tolerant plant Caragana korshinskii. Acta Physiol. Plant. 2017, 39, 116. [Google Scholar] [CrossRef]
- Kumar, S.; Yadav, M.; Yadav, A.; Yadav, J. Impact of spatial and climatic conditions on phytochemical diversity and in vitro antioxidant activity of Indian Aloe vera (L.) Burm.f. S. Afr. J. Bot. 2017, 111, 50–59. [Google Scholar] [CrossRef]
- Boussaa, F.; Zaouay, F.; Burlo-Carbonell, F.; Noguera-Artiaga, L.; Carbonell-Barrachina, A.; Melgarejo, P.; Hernandez, F.; Mars, M. Growing Location Affects Physical Properties, Bioactive Compounds, and Antioxidant Activity of Pomegranate Fruit (Punicagranatum L. var. Gabsi). Int. J. Fruit Sci. 2020, 20, S508–S523. [Google Scholar] [CrossRef]
- Kumar, S.; Budhwar, L.; Yadav, A.; Yadav, M.; Yadav, J.P. Phytochemical Screening and Antibacterial Activity of Aloe Vera Collected from Different Climatic Regions of India. Nat. Prod. J. 2016, 6, 73–82. [Google Scholar] [CrossRef]
- Moreira, S.A.; Pintado, M.E.; Saraiva, J.A. Optimization of antioxidant activity and bioactive compounds extraction of winter savory leaves by high hydrostatic pressure. High Press. Res. 2020, 40, 543–560. [Google Scholar] [CrossRef]
- Wang, S.Y. Effect of pre-harvest conditions on antioxidant capacity in fruits. Acta Hortic. 2006, 712, 299–305. [Google Scholar] [CrossRef]
- Mditshwa, A.; Fawole, O.A.; Al-Said, F.; Al-Yahyai, R.; Opara, U.L. Phytochemical content, antioxidant capacity and physicochemical properties of pomegranate grown in different microclimates in South Africa. S. Afr. J. Plants Soil 2013, 30, 81–90. [Google Scholar] [CrossRef]
- Attanayake, R.; Eeswaran, R.; Rajapaksha, R.; Weerakkody, P.; Bandaranayake, P.C.G. Biochemical composition and expression analysis of anthocyanin biosynthetic genes of a yellow peeled and pinkish ariled pomegranate (Punicagranatum L.) cultivar are differentially regulated in response to agro-climatic conditions. J. Agric. Food Chem. 2018, 66, 8761–8771. [Google Scholar] [CrossRef]
- Susanna, B.; Annamaria, L. Quality and Antioxidant Properties of Apricot Fruits at Ready-to-Eat: Influence of the Weather Conditions under Mediterranean Coastal Area. J. Food Process. Technol. 2015, 7, 1–6. [Google Scholar] [CrossRef]
- Maslennikov, P.; Golovina, E.; Artemenko, A. Ecological and Geochemical Conditions for the Accumulation of Antioxidants in the Leaves of Lathyrusmaritimus (L.) Bigel. Plants 2020, 9, 746. [Google Scholar] [CrossRef]
- Bernal, M.; Llorens, L.; Julkunen-Tiitto, R.; Badosac, J.; Verdaguer, D. Altitudinal and seasonal changes of phenolic compounds in Buxus sempervirens leaves and cuticles. Plant Physiol. Biochem. 2013, 70, 471–482. [Google Scholar] [CrossRef]
- Mpofu, A.; Sapirstein, H.D.; Beta, T. Genotype and environmental variation in phenolic content, phenolic acid composition, and antioxidant activity of hard spring wheat. J. Agric. Food Chem. 2006, 54, 1265–1270. [Google Scholar] [CrossRef]
- Zlati’c, N.M.; Stankovi’c, M.S. Variability of secondary metabolites of the species Cichoriumintybus L. from different habitats. Plants 2017, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- KhalsiAhvazi, L.; Heshmati, G.; Zofan, P.; Akbarlo, M. Phenolic, total flavonoid and antioxidant activity of medicinal plant in forage in different stages of growth in four habitats of the northeast of Khuzestan province. J. Ecoph. Med. Plants 2016, 4, 33–46. [Google Scholar]
- Saadatmad, L.; Gorbani, M.; Niakan, M. Investigation of Changes of the Most Important Substances Secondary Effects and Antioxidant Activity of Different Organs of Elaeagnus angustifolia L. in Different Sites of Khorasan Razavi Province. J. Med. Plants 2013, 1, 58–67. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Codes | Variety | TPC (mg GAE/100 g, DW) | |||||
|---|---|---|---|---|---|---|---|
| Kalar Kahar (KK) | Lahore (L) | Swabi (S) | |||||
| Bulbs | Leaves | Bulbs | Leaves | Bulbs | Leaves | ||
| V1 | Mustang | 584.3 ± 7.817 a | 472.2 ± 19.07 abc | 659.5 ± 6.593 a | 418.2 ± 10.49 d | 478.1 ± 14.25 b | 631.1 ± 8.580 a |
| V2 | Red Orb | 365.1 ± 11.12 c | 474.6 ± 10.15 abc | 394.9 ± 2.279 b | 469.9 ± 6.363 c | 557.2 ± 11.83 a | 574.3 ± 11.19 b |
| V3 | Super Sarhad | 367.2 ± 3.057 c | 486.9 ± 2.402 ab | 295.8 ± 2.821 e | 368.2 ± 0.697 e | 270.7 ± 7.218 d | 514.1 ± 11.92 c |
| V4 | Red Flame | 315.0 ± 3.750 d | 494.1 ± 8.797 a | 298.6 ± 8.377 de | 370.4 ± 0.701 e | 271.4 ± 1.590 d | 368.1 ± 2.970 d |
| V5 | Pulkara | 276.2 ± 0.985 e | 303.0 ± 21.29 e | 256.6 ± 16.27 f | 364.3 ± 6.126 e | 190.3 ± 0.000 e | 515.6 ± 16.64 c |
| V6 | Golden Orb | 506.7 ± 2.944 b | 414.5 ± 9.625 d | 355.0 ± 1.115 c | 443.6 ± 33.56 cd | 328.8 ± 2.492 c | 385.7 ± 18.00 d |
| V7 | White Pearl | 179.6 ± 1.093 g | 417.8 ± 5.671 d | 284.3 ± 10.95 e | 401.1 ± 11.17 de | 255.6 ± 9.869 d | 304.8 ± 31.81 e |
| V8 | Amazon | 243.8 ± 1.672 f | 435.1 ± 29.40 cd | 320.0 ± 6.041 d | 626.5 ± 2.870 a | 269.4 ± 16.55 d | 489.5 ± 7.562 c |
| V9 | Zeus | 374.6 ± 3.911 c | 445.4 ± 4.914 bcd | 217.9 ± 9.550 g | 530.0 ± 28.19 b | 274.6 ± 14.79 d | 404.5 ± 19.81 d |
| Codes | Variety | TFC (mg QE/100 g, DW) | |||||
| Kalar Kahar (KK) | Lahore (L) | Swabi (S) | |||||
| Bulbs | Leaves | Bulbs | Leaves | Bulbs | Leaves | ||
| V1 | Mustang | 303.0 ± 6.670 a | 340.2 ± 14.60 e | 229.7 ± 9.296 a | 318.2 ± 16.77 b | 157.7 ± 0.000 a | 253.2 ± 21.54 ab |
| V2 | Red Orb | 127.9 ± 2.865 c | 432.5 ± 10.36 ab | 230.3 ± 3.788 a | 414.3 ± 19.92 a | 145.7 ± 12.83 a | 284.5 ± 6.748 a |
| V3 | Super Sarhed | 183.6 ± 3.526 b | 368.3 ± 2.141 e | 93.87 ± 7.764 bc | 329.2 ± 10.68 b | 99.47 ± 7.289 c | 227.1 ± 10.36 b |
| V4 | Red Flame | 121.2 ± 4.036 c | 479.0 ± 34.05 a | 75.80 ± 6.428 de | 229.0 ± 12.99 c | 98.72 ± 1.361 c | 260.7 ± 22.76 ab |
| V5 | Pulkara | 101.6 ± 6.442 d | 238.6 ± 24.34 f | 64.59 ± 1.405 e | 202.1 ± 1.129 c | 45.30 ± 1.411 e | 241.5 ± 16.77 ab |
| V6 | Golden Orb | 303.0 ± 2.521 a | 352.3 ± 8.099 e | 110.8 ± 6.356 b | 226.5 ± 23.43 c | 122.7 ± 0.000 b | 223.8 ± 22.64 b |
| V7 | White Pearl | 53.29 ± 1.429 f | 393.1 ± 3.037 bc | 31.86 ± 1.430 f | 138.6 ± 11.48 d | 28.95 ± 1.408 e | 216.2 ± 5.986 b |
| V8 | Amazon | 88.94 ± 2.864 e | 375.3 ± 32.49 bc | 91.61 ± 8.726 cd | 384.6 ± 11.52 a | 77.95 ± 0.000 d | 232.9 ± 10.44 b |
| V9 | Zeus | 120.9 ± 2.457 c | 364.4 ± 23.71 e | 42.60 ± 1.377 f | 316.0 ± 18.27 b | 100.1 ± 8.399 c | 215.8 ± 14.16 b |
| Bulbs | DPPH (%) | OH (%) | |||||
|---|---|---|---|---|---|---|---|
| Codes | Variety | KK | L | S | KK | L | S |
| V1 | Mustang | 79.01 ± 1.49 a | 61.38 ± 2.09 a | 78.75 ± 0.19 a | 35.05 ± 0.35 f | 64.15 ± 0.84 b | 48.41 ± 1.03 c |
| V2 | Red Orb | 51.96 ± 1.90 c | 48.13 ± 0.38 b | 48.47 ± 0.63 cd | 45.40 ± 0.18 c | 40.87 ± 0.09 d | 53.80 ± 0.39 b |
| V3 | Super Sarhad | 72.20 ± 1.14 b | 41.60 ± 1.3 c | 74.38 ± 0.19 a | 38.54 ± 0.74 e | 42.52 ± 0.29 d | 47.18 ± 0.09 c |
| V4 | Red Flame | 53.52 ± 0.09 c | 38.32 ± 3.72 cd | 45.73 ± 0.00 d | 62.13 ± 0.11 a | 65.50 ± 0.19 ab | 56.86 ± 0.29 a |
| V5 | Pulkara | 75.30 ± 0.60 ab | 41.55 ± 3.50 cd | 29.16 ± 1.61 e | 41.42 ± 0.09 d | 58.76 ± 0.85 c | 52.82 ± 0.28 b |
| V6 | Golden Orb | 78.05 ± 0.61 a | 35.54 ± 2.24 d | 53.00 ± 1.99 c | 52.94 ± 0.11 b | 67.40 ± 0.09 a | 41.91 ± 1.12 d |
| V7 | White Pearl | 12.35 ± 3.02 e | 21.78 ± 0.49 e | 23.21 ± 1.53 e | 29.53 ± 1.03 g | 30.82 ± 0.09 f | 36.15 ± 0.37 e |
| V8 | Amazon | 41.17 ± 2.41 d | 35.90 ± 1.07 cd | 61.51 ± 1.04 b | 37.93 ± 0.54 e | 36.09 ± 1.03 e | 56.99 ± 0.18 a |
| V9 | Zeus | 45.31 ± 0.97 d | 17.08 ± 1.00 e | 50.79 ± 6.64 cd | 28.86 ± 0.00 h | 28.06 ± 0.42 g | 31.19 ± 0.85 f |
| Leaves | |||||||
| V1 | Mustang | 60.66 ± 1.50 a | 63.74 ± 1.13 b | 76.42 ± 0.47 a | 51.76 ± 2.23 a | 53.83 ± 1.58 b | 46.36 ± 3.17 ab |
| V2 | Red Orb | 63.85 ± 1.23 a | 57.89 ± 0.75 cd | 76.20 ± 1.15 a | 22.72 ± 3.30 c | 24.81 ± 1.16 de | 45.11 ± 1.00 bc |
| V3 | Super Sarhad | 63.28 ± 2.64 a | 50.92 ± 1.33 f | 72.02 ± 1.66 b | 24.36 ± 2.53 c | 30.17 ± 2.46 c | 41.00 ± 1.42 cd |
| V4 | Red Flame | 56.53 ± 1.33 b | 57.43 ± 1.37 cd | 63.94 ± 1.50 c | 19.91 ± 2.23 c | 54.12 ± 0.92 b | 28.64 ± 2.41 fg |
| V5 | Pulkara | 61.77 ± 0.87 a | 52.54 ± 1.33 ef | 69.29 ± 0.60 b | 24.82 ± 0.00 c | 54.89 ± 0.00 b | 50.00 ± 0.00 a |
| V6 | Golden Orb | 43.55 ± 1.27 d | 60.62 ± 2.02 bc | 60.41 ± 1.20 d | 48.87 ± 0.14 a | 61.21 ± 0.76 a | 31.61 ± 1.75 ef |
| V7 | White Pearl | 45.13 ± 1.37 cd | 55.47 ± 1.15 de | 51.71 ± 0.41 e | 11.63 ± 1.29 d | 23.75 ± 0.17 e | 33.91 ± 1.49 e |
| V8 | Amazon | 54.61 ± 1.16 b | 90.69 ± 0.26 a | 54.55 ± 1.58 e | 32.86 ± 0.14 b | 23.75 ± 1.16 e | 39.37 ± 0.50 d |
| V9 | Zeus | 49.13 ± 0.30 c | 88.65 ± 0.58 a | 65.50 ± 0.87 c | 21.31 ± 1.62 c | 28.07 ± 1.63 cd | 26.15 ± 0.29 g |
| Bulbs | H2O2 (%) | ABTS (%) | |||||
|---|---|---|---|---|---|---|---|
| Codes | Variety | KK | L | S | KK | L | S |
| V1 | Mustang | 53.72 ± 1.29 a | 55.35 ± 0.93 a | 59.76 ± 0.90 a | 30.17 ± 2.02 a | 65.38 ± 0.99 a | 29.88 ± 1.89 b |
| V2 | Red Orb | 33.33 ± 0.82 e | 51.09 ± 0.54 b | 46.43 ± 1.16 d | 24.74 ± 1.13 ab | 60.66 ± 1.46 b | 41.92 ± 0.35 a |
| V3 | Super Sarhad | 49.44 ± 1.84 b | 48.88 ± 1.11 bc | 50.02 ± 1.07 c | 19.35 ± 0.82 ab | 49.24 ± 0.89 d | 19.51 ± 0.86 c |
| V4 | Red Flame | 46.44 ± 1.01 c | 44.27 ± 0.66 d | 41.37 ± 1.28 e | 9.240 ± 1.04 b | 50.36 ± 0.43 cd | 19.88 ± 1.02 c |
| V5 | Pulkara | 37.44 ± 1.08 b | 41.89 ± 1.42 e | 41.89 ± 1.42 c | 31.37 ± 12.7 a | 49.27 ± 0.57 d | 11.03 ± 0.48 de |
| V6 | Golden Orb | 41.37 ± 1.28 d | 46.43 ± 1.16 cd | 53.72 ± 1.29 b | 32.80 ± 14.5 a | 52.98 ± 1.04 c | 16.91 ± 1.00 c |
| V7 | White Pearl | 39.14 ± 0.99 d | 31.31 ± 0.74 h | 26.97 ± 0.87 f | 9.221 ± 0.94 b | 30.28 ± 1.22 e | 11.44 ± 1.12 d |
| V8 | Amazon | 33.33 ± 0.82 e | 37.44 ± 1.08 f | 46.43 ± 1.16 d | 10.47 ± 1.89 b | 50.20 ± 0.54 d | 11.38 ± 1.14 d |
| V9 | Zeus | 46.43 ± 1.16 bc | 33.33 ± 0.82 g | 40.82 ± 0.92 e | 11.26 ± 1.59 b | 8.560 ± 0.60 f | 8.370 ± 0.74 e |
| Leaves | |||||||
| V1 | Mustang | 33.86 ± 1.32 b | 42.39 ± 0.47 ab | 47.87 ± 1.96 b | 10.04 ± 0.98 a | 41.36 ± 1.01 d | 12.33 ± 0.76 b |
| V2 | Red Orb | 39.80 ± 0.75 a | 44.18 ± 1.63 a | 42.34 ± 0.67 c | 11.25 ± 1.36 a | 42.89 ± 0.45 d | 12.89 ± 0.63 b |
| V3 | Super Sarhad | 31.46 ± 0.25 b | 33.69 ± 0.41 c | 37.13 ± 0.27 d | 6.890 ± 0.35 a | 37.39 ± 2.77 e | 13.46 ± 0.78 b |
| V4 | Red Flame | 33.69 ± 0.41 b | 40.87 ± 0.35 b | 40.87 ± 0.35 c | 11.84 ± 2.89 a | 56.18 ± 0.67 b | 17.58 ± 2.56 a |
| V5 | Pulkara | 22.11 ± 2.99 c | 32.73 ± 1.88 c | 29.23 ± 0.38 fg | 7.626 ± 1.77 a | 51.17 ± 1.14 c | 19.31 ± 0.87 a |
| V6 | Golden Orb | 40.87 ± 0.35 a | 44.18 ± 1.63 a | 31.46 ± 0.25 ef | 10.23 ± 1.62 a | 51.30 ± 1.24 c | 11.59 ± 0.43 b |
| V7 | White Pearl | 22.11 ± 2.99 c | 24.21 ± 0.70 d | 28.45 ± 1.22 g | 12.42 ± 8.99 a | 19.41 ± 0.77 f | 6.062 ± 0.66 c |
| V8 | Amazon | 24.21 ± 0.70 c | 39.80 ± 0.75 b | 51.86 ± 0.43 a | 11.53 ± 2.33 a | 63.55 ± 1.07 a | 19.68 ± 1.15 a |
| V9 | Zeus | 31.18 ± 0.85 b | 24.21 ± 0.70 d | 32.61 ± 0.55 e | 11.95 ± 1.21 a | 61.58 ± 0.28 a | 17.47 ± 0.55 a |
| Bulbs | FRAP (mg GAE/100 g DW) | PMA (mg AAE/100 g DW) | |||||
|---|---|---|---|---|---|---|---|
| Codes | Variety | KK | L | S | KK | L | S |
| V1 | Mustang | 415.1 ± 10.7 a | 373.5 ± 16.5 a | 271.2 ± 27.6 a | 141.3 ± 9.14 ab | 194.2 ± 9.54 a | 213.8 ± 0.00 a |
| V2 | Red Orb | 153.9 ± 2.56 e | 270.4 ± 9.21 b | 269.1 ± 13.8 a | 85.45 ± 8.35 e | 106.2 ± 9.89 de | 183.2 ± 17.8 b |
| V3 | Super Sarhad | 264.6 ± 4.13 c | 278.9 ± 17.2 b | 221.1 ± 14.2 bc | 112.6 ± 0.62 cde | 86.94 ± 1.30 ef | 122.2 ± 5.83 de |
| V4 | Red Flame | 255.9 ± 5.70 c | 182.8 ± 3.87 cd | 198.8 ± 9.92 bc | 137.0 ± 16.0 abc | 91.23 ± 4.58 ef | 141.4 ± 14.3 cd |
| V5 | Pulkara | 192.2 ± 11.4 d | 140.6 ± 2.41 e | 137.2 ± 15.5 d | 107.1 ± 2.08 de | 80.43 ± 2.27 f | 72.52 ± 4.55 f |
| V6 | Golden Orb | 315.6 ± 5.19 b | 203.3 ± 3.75 c | 235.5 ± 19.2 ab | 157.0 ± 9.43 a | 144.7 ± 1.18 bc | 133.2 ± 0.00 cd |
| V7 | White Pearl | 98.83 ± 3.24 f | 194.9 ± 4.95 c | 204.0 ± 0.70 bc | 136.1 ± 4.16 abc | 163.9 ± 15.8 b | 140.5 ± 9.55 cd |
| V8 | Amazon | 114.6 ± 0.71 f | 256.6 ± 13.3 b | 189.5 ± 12.8 c | 124.8 ± 18.1 bcd | 101.8 ± 11.0 def | 151.1 ± 8.46 c |
| V9 | Zeus | 328.3 ± 23.1 b | 152.6 ± 17.9 de | 219.8 ± 12.4 bc | 160.7 ± 4.03 a | 120.3 ± 10.3 cd | 99.48 ± 3.59 e |
| Leaves | |||||||
| V1 | Mustang | 74.58 ± 5.98 bc | 61.14 ± 10.4 e | 132.9 ± 3.90 bc | 156.7 ± 9.93 e | 179.1 ± 2.67 f | 606.8 ± 0.69 a |
| V2 | Red Orb | 92.41 ± 0.61 a | 91.90 ± 1.98 c | 157.7 ± 10.1 a | 160.4 ± 12.8 e | 460.0 ± 5.28 b | 570.7 ± 1.14 b |
| V3 | Super Sarhad | 55.12 ± 3.89 de | 75.30 ± 0.71 de | 128.2 ± 2.52 c | 225.3 ± 2.58 d | 523.0 ± 0.00 a | 623.6 ± 0.72 a |
| V4 | Red Flame | 55.89 ± 3.55 de | 85.43 ± 4.79 cd | 127.8 ± 3.18 c | 241.8 ± 5.06 d | 385.9 ± 16.3 d | 354.0 ± 16.4 d |
| V5 | Pulkara | 49.64 ± 4.63 e | 37.97 ± 3.81 f | 146.0 ± 4.16 ab | 332.8 ± 10.9 c | 329.6 ± 1.74 e | 366.7 ± 9.84 cd |
| V6 | Golden Orb | 65.84 ± 4.03 cd | 75.49 ± 5.07 cde | 120.5 ± 8.09 c | 349.3 ± 3.24 c | 408.5 ± 0.00 cd | 365.6 ± 14.4 cd |
| V7 | White Pearl | 63.62 ± 0.00 cd | 28.44 ± 3.44 f | 71.12 ± 1.82 d | 411.6 ± 13.0 a | 411.8 ± 0.00 cd | 364.7 ± 10.4 d |
| V8 | Amazon | 80.62 ± 5.21 ab | 184.2 ± 6.75 a | 119.9 ± 4.37 c | 380.7 ± 0.70 b | 424.0 ± 5.23 bc | 392.5 ± 0.00 c |
| V9 | Zeus | 74.36 ± 5.50 bc | 150.2 ± 8.17 b | 131.6 ± 4.45 bc | 336.9 ± 4.51 c | 397.9 ± 33.8 cd | 341.9 ± 11.8 d |
| Agro-Climatic Conditions | Codes | Localities | ||
|---|---|---|---|---|
| Kalar Kahar | Lahore | Swabi | ||
| Mean maximum temperature (°C) | Max.T | 31.33 ± 9.04 | 30.85 ± 8.91 | 28.60 ± 8.26 |
| Mean minimum temperature (°C) | Min.T | 17.94 ± 5.18 | 19.22 ± 5.55 | 16.43 ± 4.74 |
| Mean temperature (°C) | MT | 24.63 ± 7.11 | 25.03 ± 7.23 | 22.52 ± 6.50 |
| Average rain fall (mm) | Rf | 53.17 ± 15.3 | 52.35 ± 15.1 | 64.00 ± 18.5 |
| Relative humidity (%) | RH | 41.20 ± 11.9 | 39.18 ± 11.3 | 44.46 ± 12.8 |
| Surface pressure (kPa) | SP | 96.27 ± 27.8 | 98.33 ± 28.4 | 93.03 ± 26.9 |
| Dew/frost point (°C) | D | 8.637 ± 2.49 | 9.546 ± 2.76 | 8.302 ± 2.40 |
| Mean maximum wind speed (m/s) | MWs | 3.423 ± 0.99 | 3.457 ± 1.00 | 2.842 ± 0.82 |
| Mean minimum wind speed (m/s) | MnWs | 1.004 ± 0.29 | 1.045 ± 0.30 | 0.653 ± 0.19 |
| Altitude (m) | Alt. | 401.6 | 213.0 | 706.0 |
| pH | pH | 6.085 ± 0.03 | 6.195 ± 0.14 | 6.360 ± 0.05 |
| Electrical conductivity (dSm−1) | EC | 2.135 ± 0.36 | 12.77 ± 0.77 | 1.960 ± 0.73 |
| Soil organic matter (%) | Om | 0.875 ± 0.03 | 0.900 ± 0.07 | 0.925 ± 0.03 |
| Variables | Bulbs | Leaves | ||
|---|---|---|---|---|
| PC1 | PC2 | PC1 | PC2 | |
| Eigen value | 14.79 | 6.201 | 14.71 | 6.290 |
| Total variance (%) | 70.47 | 29.57 | 70.04 | 29.95 |
| Cumulative variance (%) | 70.47 | 100.0 | 70.04 | 100.0 |
| Total phenolic content (TPC) | 0.869 | −0.494 | 0.994 | 0.106 |
| Total flavonoid content (TFC) | 0.562 | −0.827 | −0.730 | −0.684 |
| FRAP assay | 0.851 | −0.525 | 0.967 | 0.256 |
| PMA assay | −0.940 | −0.341 | 0.809 | 0.588 |
| DPPH assay | −0.356 | −0.935 | 0.665 | 0.747 |
| OH assay | −0.292 | 0.957 | 0.468 | 0.884 |
| H2O2 assay | −0.815 | 0.580 | 0.749 | 0.662 |
| ABTS assay | 0.609 | 0.793 | −0.334 | 0.943 |
| Maximum temperature (Max.T) | 0.966 | −0.257 | −0.997 | −0.082 |
| Minimum temperature (Min.T) | 0.929 | 0.371 | −0.848 | 0.530 |
| Mean temperature (MT) | 0.999 | 0.052 | −0.974 | 0.228 |
| Rain fall (Rf) | −0.999 | 0.033 | 0.990 | −0.144 |
| Relative humidity (RH) | −0.958 | −0.288 | 0.892 | −0.453 |
| Surface pressure (SP) | 0.956 | 0.295 | −0.888 | 0.459 |
| Dew/frost point (D) | 0.773 | 0.635 | −0.648 | 0.761 |
| Maximum wind speed (MWs) | 0.999 | −0.042 | −0.991 | 0.135 |
| Minimum wind speed (MnWs) | 1.000 | −0.011 | −0.986 | 0.166 |
| pH | −0.878 | 0.480 | 0.948 | 0.317 |
| Electrical conductivity (EC) | 0.585 | 0.811 | −0.432 | 0.902 |
| Organic matter (OM) | −0.791 | 0.611 | 0.887 | 0.462 |
| Altitude (Alt.) | −0.957 | −0.289 | 0.891 | −0.454 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bibi, N.; Shah, M.H.; Khan, N.; Al-Hashimi, A.; Elshikh, M.S.; Iqbal, A.; Ahmad, S.; Abbasi, A.M. Variations in Total Phenolic, Total Flavonoid Contents, and Free Radicals’ Scavenging Potential of Onion Varieties Planted under Diverse Environmental Conditions. Plants 2022, 11, 950. https://doi.org/10.3390/plants11070950
Bibi N, Shah MH, Khan N, Al-Hashimi A, Elshikh MS, Iqbal A, Ahmad S, Abbasi AM. Variations in Total Phenolic, Total Flavonoid Contents, and Free Radicals’ Scavenging Potential of Onion Varieties Planted under Diverse Environmental Conditions. Plants. 2022; 11(7):950. https://doi.org/10.3390/plants11070950
Chicago/Turabian StyleBibi, Nusrat, Munir H. Shah, Nadeem Khan, Abdulrahman Al-Hashimi, Mohamed Soliman Elshikh, Akhtar Iqbal, Shakeel Ahmad, and Arshad Mehmood Abbasi. 2022. "Variations in Total Phenolic, Total Flavonoid Contents, and Free Radicals’ Scavenging Potential of Onion Varieties Planted under Diverse Environmental Conditions" Plants 11, no. 7: 950. https://doi.org/10.3390/plants11070950
APA StyleBibi, N., Shah, M. H., Khan, N., Al-Hashimi, A., Elshikh, M. S., Iqbal, A., Ahmad, S., & Abbasi, A. M. (2022). Variations in Total Phenolic, Total Flavonoid Contents, and Free Radicals’ Scavenging Potential of Onion Varieties Planted under Diverse Environmental Conditions. Plants, 11(7), 950. https://doi.org/10.3390/plants11070950