
Characteristics of Sunburn Browning Fruit and Rootstock-Dependent Damage-Free Yield of Ambrosia™ Apple after Sustained Summer Heat Events



Abstract
:1. Introduction
2. Results
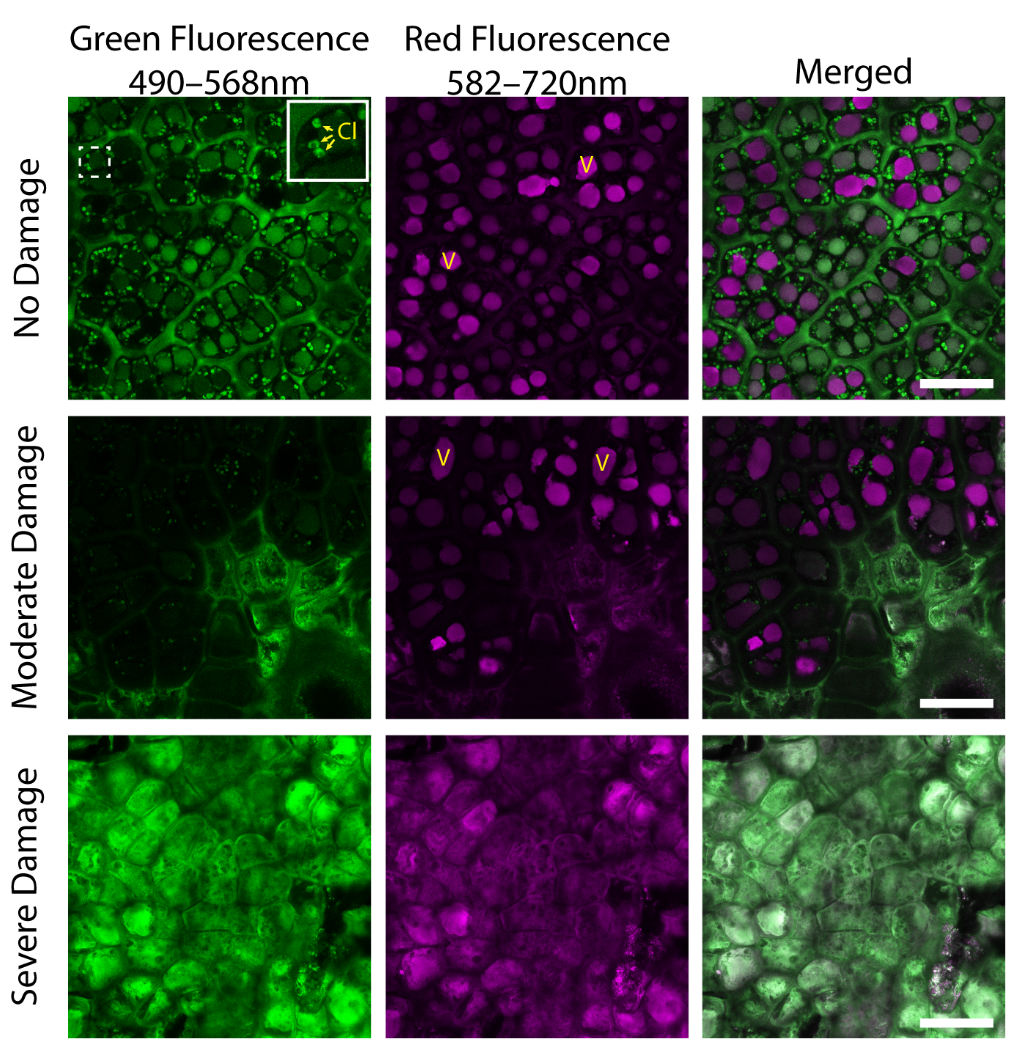
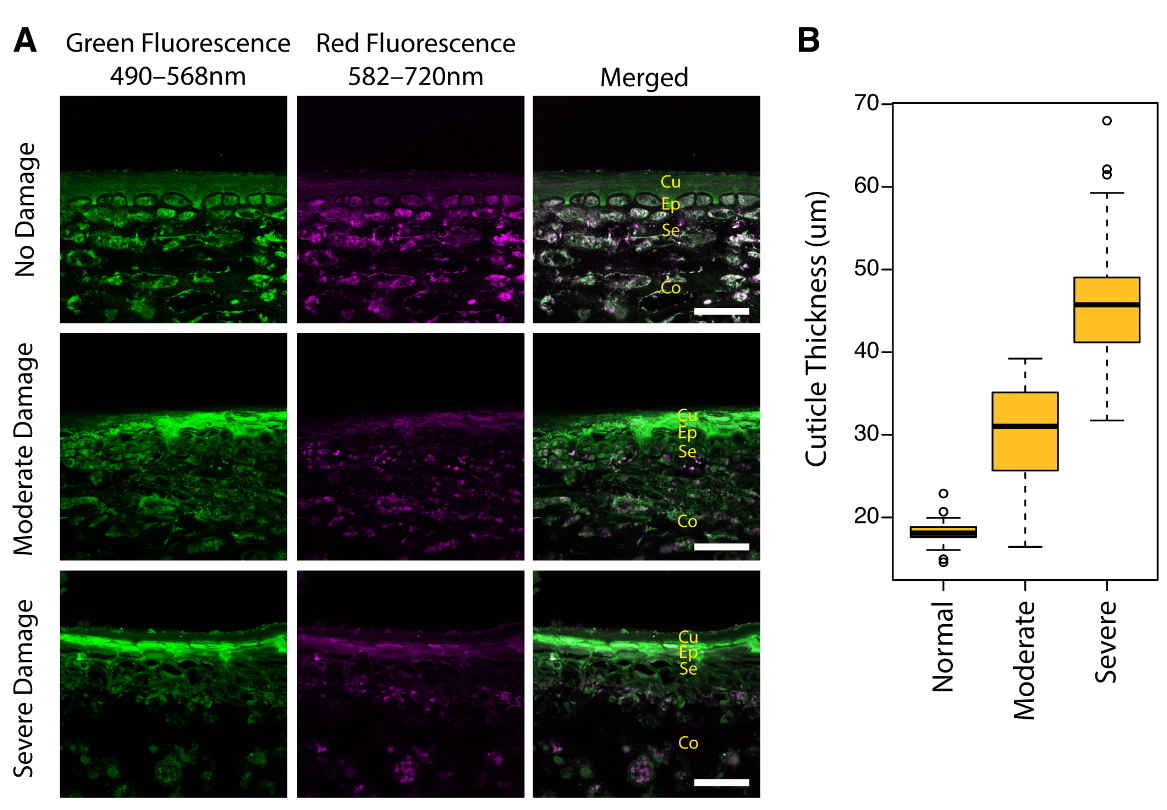
2.1. Cellular Structures of Fruit Skin in Sunburn Browning Apple
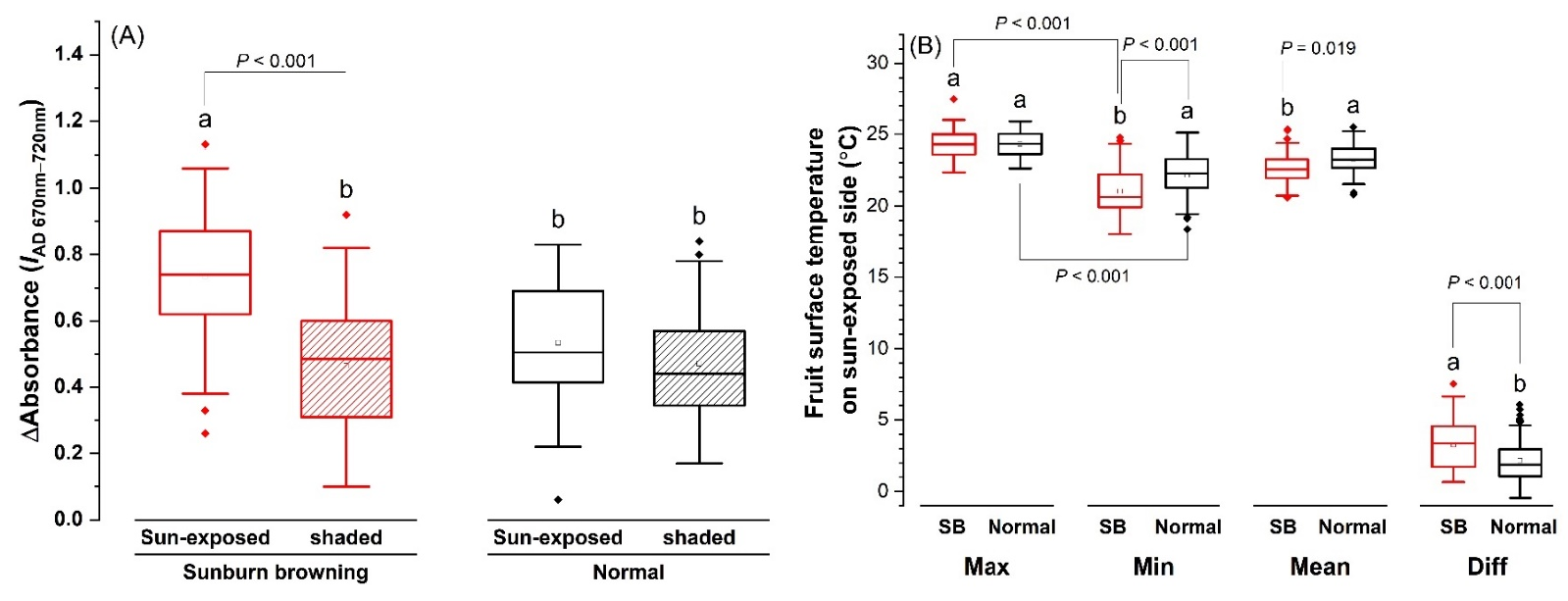
2.2. Spectral Characteristics of Fruit Skin in SB Apple
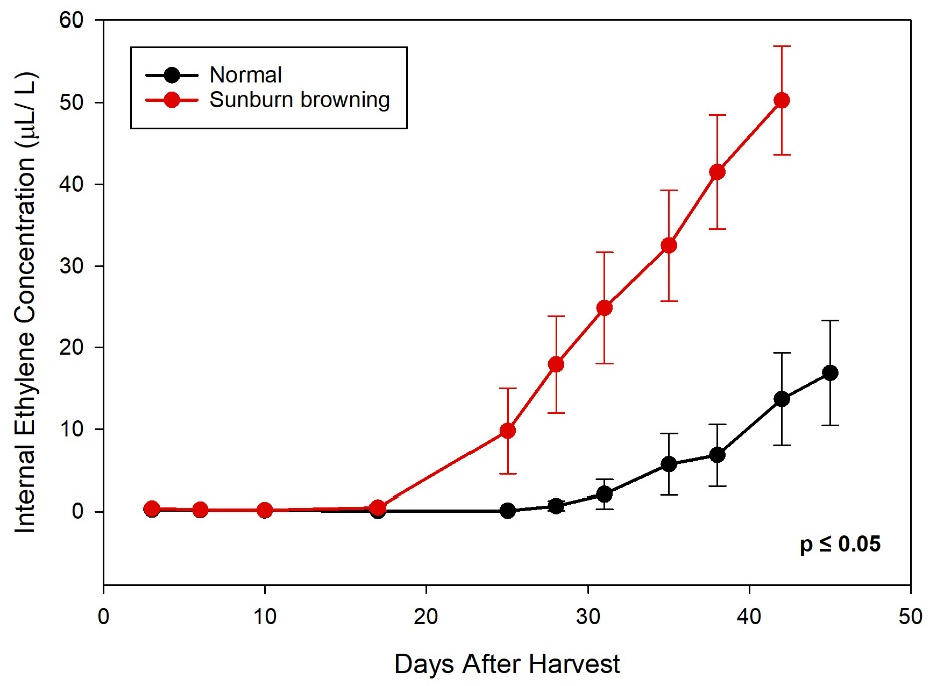
2.3. Dynamics of Internal Attributes of SB Apple
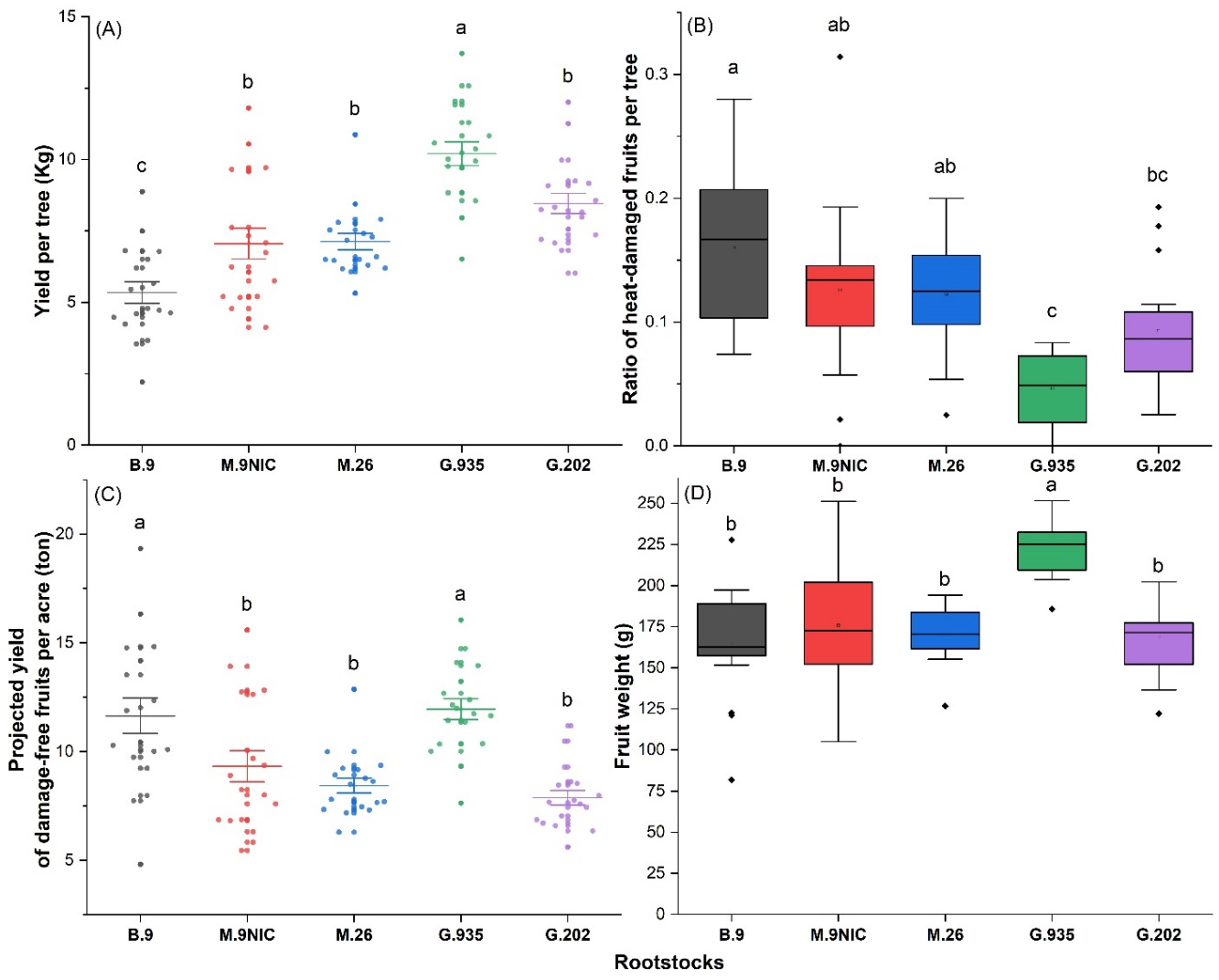
2.4. Vigor, Yield, and Ratio of Heat-Damaged Fruits on Five Rootstocks
3. Discussion
3.1. Fruit Responses to Heat and UV Stress
3.2. Heat, Water, and Rootstock Mitigation
3.3. Fruit Quality Deterioration in SB Apple
3.4. Detection Using Handheld Tools
3.5. Summary and Future Perspectives
4. Materials and Methods
4.1. Experimental Design and Study Materials
4.2. Spectral Characteristics of Fruit Skin
4.3. Confocal Microscopy
4.4. Fruit Compositional Attributes
4.5. Ethylene Emission
4.6. Rootstock Evaluation
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Greer, D.H. Temperature-dependent responses of the photosynthetic and chlorophyll fluorescence attributes of apple (Malus domestica) leaves during a sustained high temperature event. Plant Physiol. Biochem. 2015, 97, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Morales-Quintana, L.; Waite, J.M.; Kalcsits, L.; Torres, C.A.; Ramos, P. Sun injury on apple fruit: Physiological, biochemical and molecular advances, and future challenges. Sci. Hortic. 2020, 260, 108866. [Google Scholar] [CrossRef]
- Blankenship, S.M. Night-temperature effects on rate of apple fruit maturation and fruit quality. Sci. Hortic. 1987, 33, 205–212. [Google Scholar] [CrossRef]
- Caprio, J.M.; Quamme, H.A. Weather conditions associated with apple production in the Okanagan Valley of British Columbia. Can. J. Plant Sci. 1999, 79, 129–137. [Google Scholar] [CrossRef]
- Torres, C.A.; Sepulveda, A.; Gonzalez-Talice, J.; Yuri, J.A.; Razmilic, I. Fruit water relations and osmoregulation on apples (Malus domestica Borkh.) with different sun exposures and sun-injury levels on the tree. Sci. Hortic. 2013, 161, 143–152. [Google Scholar] [CrossRef]
- Racsko, J.; Schrader, L.E. Sunburn of Apple Fruit: Historical Background, Recent Advances and Future Perspectives. Crit. Rev. Plant Sci. 2012, 31, 455–504. [Google Scholar] [CrossRef]
- Felicetti, D.A.; Schrader, L.E. Changes in pigment concentrations associated with the degree of sunburn browning of ‘Fuji’ apple. J. Am. Soc. Hortic. Sci. 2008, 133, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Felicetti, D.A.; Schrader, L.E. Changes in pigment concentrations associated with sunburn browning of five apple cultivars. I. Chlorophylls and carotenoids. Plant Sci. 2009, 176, 78–83. [Google Scholar] [CrossRef]
- Felicetti, D.A.; Schrader, L.E. Changes in pigment concentrations associated with sunburn browning of five apple cultivars. II. Phenolics. Plant Sci. 2009, 176, 84–89. [Google Scholar] [CrossRef]
- Xu, H.; Ediger, D. Rootstocks with different vigor influenced scion–water relations and stress responses in Ambrosia™ apple trees (Malus domestica var. Ambrosia). Plants 2021, 10, 614. [Google Scholar] [CrossRef]
- Martin, L.B.; Rose, J.K. There’s more than one way to skin a fruit: Formation and functions of fruit cuticles. J. Exp. Bot. 2014, 65, 4639–4651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solovchenko, A.; Merzlyak, M. Optical properties and contribution of cuticle to UV protection in plants: Experiments with apple fruit. Photochem. Photobiol. Sci. 2003, 2, 861–866. [Google Scholar] [CrossRef] [PubMed]
- Lara, I.; Heredia, A.; Domínguez, E. Shelf life potential and the fruit cuticle: The unexpected player. Front. Plant Sci. 2019, 10, 770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, C.A.; Sepúlveda, A.; Leon, L.; Yuri, J.A. Early detection of sun injury on apples (Malus domestica Borkh.) through the use of crop water stress index and chlorophyll fluorescence. Sci. Hortic. 2016, 211, 336–342. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Chivkunova, O.B. Light-stress-induced pigment changes and evidence for anthocyanin photoprotection in apples. J. Photochem. Photobiol. Biol. 2020, 55, 155–163. [Google Scholar] [CrossRef]
- Donaldson, L. Autofluorescence in plants. Molecules 2020, 25, 2393. [Google Scholar] [CrossRef]
- Jiang, S.; Chen, M.; Wang, Z.; Ren, Y.; Wang, B.; Zhu, J.; Zhang, Y. Advances in understanding the causes, molecular mechanism, and perspectives of russeting on tree fruit. Front. Plant Sci. 2022, 13, 834109. [Google Scholar] [CrossRef]
- Kitin, P.; Nakaba, S.; Hunt, C.G.; Lim, S.; Funada, R. Direct fluorescence imaging of lignocellulosic and suberized cell walls in root and stems. Ann. Bot. 2020, 12, plaa032. [Google Scholar] [CrossRef]
- Reig, G.; Donahue, D.J.; Jentsch, P. The efficacy of four sunburn mitigation strategies and their effects on yield, fruit quality, and economic performance of Honeycrisp cv. Apples under Eastern New York (USA) Climatic Conditions. Int. J. Fruit Sci. 2020, 20, 541–561. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Webster, A.D.; Vaughan, S.; Lucas, A.S. Effects of root restriction on the physiology of apple tree growth. Acta Hortic. 1997, 451, 587–598. [Google Scholar] [CrossRef]
- Jones, H.G. How do rootstocks control shoot water relations? New Phytol. 2012, 194, 301–303. [Google Scholar] [CrossRef] [PubMed]
- Tworkoski, T.; Fazio, G. Effects of size-controlling apple rootstocks on growth, abscisic acid, and hydraulic conductivity of scion of different vigor. Int. J. Fruit Sci. 2015, 15, 369–381. [Google Scholar] [CrossRef]
- Xu, H.; Ediger, D.; Singh, A.; Pagliocchini, C. Rootstock–scion hydraulic balance influenced scion vigor and yield efficiency of Malus domestica cv. Honeycrisp on eight rootstocks. Horticulturae 2021, 7, 99. [Google Scholar] [CrossRef]
- Torres, C.A.; Sepúlveda, G.; Kahlaoui, B. Phytohormone Interaction Modulating Fruit Responses to Photooxidative and Heat Stress on Apple (Malus domestica Borkh.). Front. Plant Sci. 2017, 8, 2129. [Google Scholar] [CrossRef] [Green Version]
- Gapper, N.E.; McQuinn, R.P.; Giovannoni, J.J. Molecular and genetic regulation of fruit ripening. Plant Mol. Biol. 2013, 82, 575–591. [Google Scholar] [CrossRef]
- Knoche, M.; Khanal, B.P.; Brüggenwirth, M.; Thapa, S. Patterns of microcracking in apple fruit skin reflect those of the cuticular ridges and of the epidermal cell walls. Planta 2018, 248, 293–306. [Google Scholar] [CrossRef]
- Torres, C.A.; León, L.; Sánchez-Contreras, J. Spectral fingerprints during sun injury development on the tree in Granny Smith apples: A potential non-destructive prediction tool during the growing season. Sci. Hortic. 2016, 209, 165–172. [Google Scholar] [CrossRef]
- Lordan, J.; Francescatto, P.; Dominguez, L.I.; Robinson, T.L. Long-term effects of tree density and tree shape on apple orchard performance, a 20 year study—Part 1, agronomic analysis. Sci. Hortic. 2018, 238, 303–317. [Google Scholar] [CrossRef]
- Robinson, T. Apple Rootstock Performance—An Eastern Perspective. SCRI Apple Root to Fruit Webinar on Rootstock and Nutrition. 1 March 2022. Available online: https://youtube.com/playlist?list=PLYLbxsK4pTXXoyNLVkWZjKprOVOkkoQCL (accessed on 20 March 2022).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fruit Condition | Weight (g) | SSC (%) | Malic Acid (mg/100 mL) | Side of Fruit | Firmness (lbs) | DMC (%) | ΨFruit (MPa) |
|---|---|---|---|---|---|---|---|
| Normal | 188.49 ± 4.14 a | 14.23 ± 0.06 b | 772.82 ± 17.18 b | Sun-exposed | 19.00 ± 0.31 bc | 16.99 ± 0.18 b | −1.71 ± 0.07 a |
| Shaded | 18.68 ± 1.28 c | 15.58 ± 0.13 c | −1.68 ± 0.04 a | ||||
| Sunburn browning | 158.09 ± 2.48 b | 16.55 ± 0.08 a | 1092.89 ± 120.82 a | Sun-exposed | 21.22 ± 0.32 a | 22.14 ± 0.35 a | −2.47 ± 0.09 b |
| Shaded | 19.79 ± 1.22 b | 15.34 ± 0.20 c | −1.65 ± 0.04 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; Watanabe, Y.; Ediger, D.; Yang, X.; Iritani, D. Characteristics of Sunburn Browning Fruit and Rootstock-Dependent Damage-Free Yield of Ambrosia™ Apple after Sustained Summer Heat Events. Plants 2022, 11, 1201. https://doi.org/10.3390/plants11091201
Xu H, Watanabe Y, Ediger D, Yang X, Iritani D. Characteristics of Sunburn Browning Fruit and Rootstock-Dependent Damage-Free Yield of Ambrosia™ Apple after Sustained Summer Heat Events. Plants. 2022; 11(9):1201. https://doi.org/10.3390/plants11091201
Chicago/Turabian StyleXu, Hao, Yoichiro Watanabe, Danielle Ediger, Xiaotang Yang, and Davis Iritani. 2022. "Characteristics of Sunburn Browning Fruit and Rootstock-Dependent Damage-Free Yield of Ambrosia™ Apple after Sustained Summer Heat Events" Plants 11, no. 9: 1201. https://doi.org/10.3390/plants11091201
APA StyleXu, H., Watanabe, Y., Ediger, D., Yang, X., & Iritani, D. (2022). Characteristics of Sunburn Browning Fruit and Rootstock-Dependent Damage-Free Yield of Ambrosia™ Apple after Sustained Summer Heat Events. Plants, 11(9), 1201. https://doi.org/10.3390/plants11091201