
Potential Distribution of Cedrela odorata L. in Mexico according to Its Optimal Thermal Range for Seed Germination under Different Climate Change Scenarios
 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Current Distribution of C. odorata
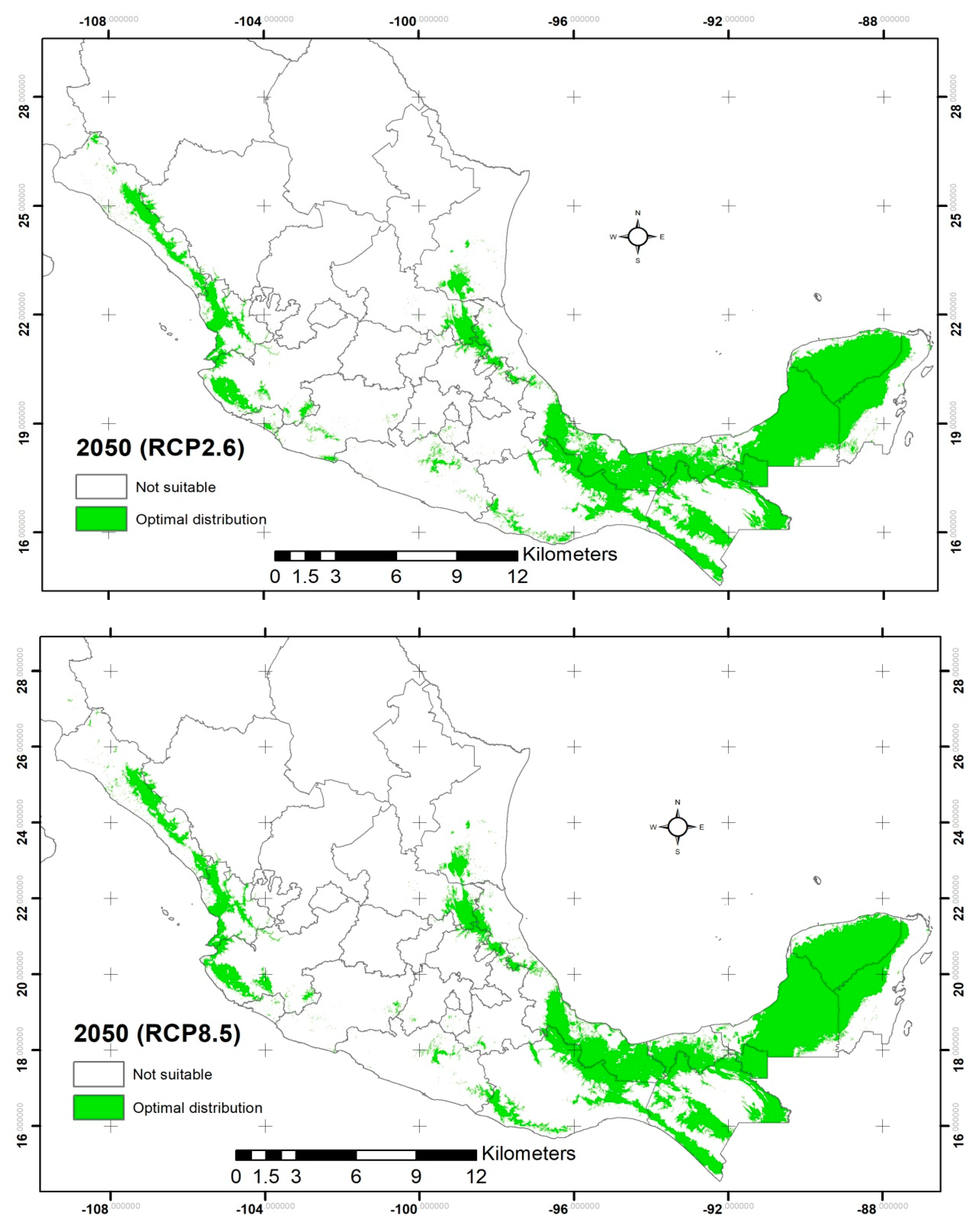
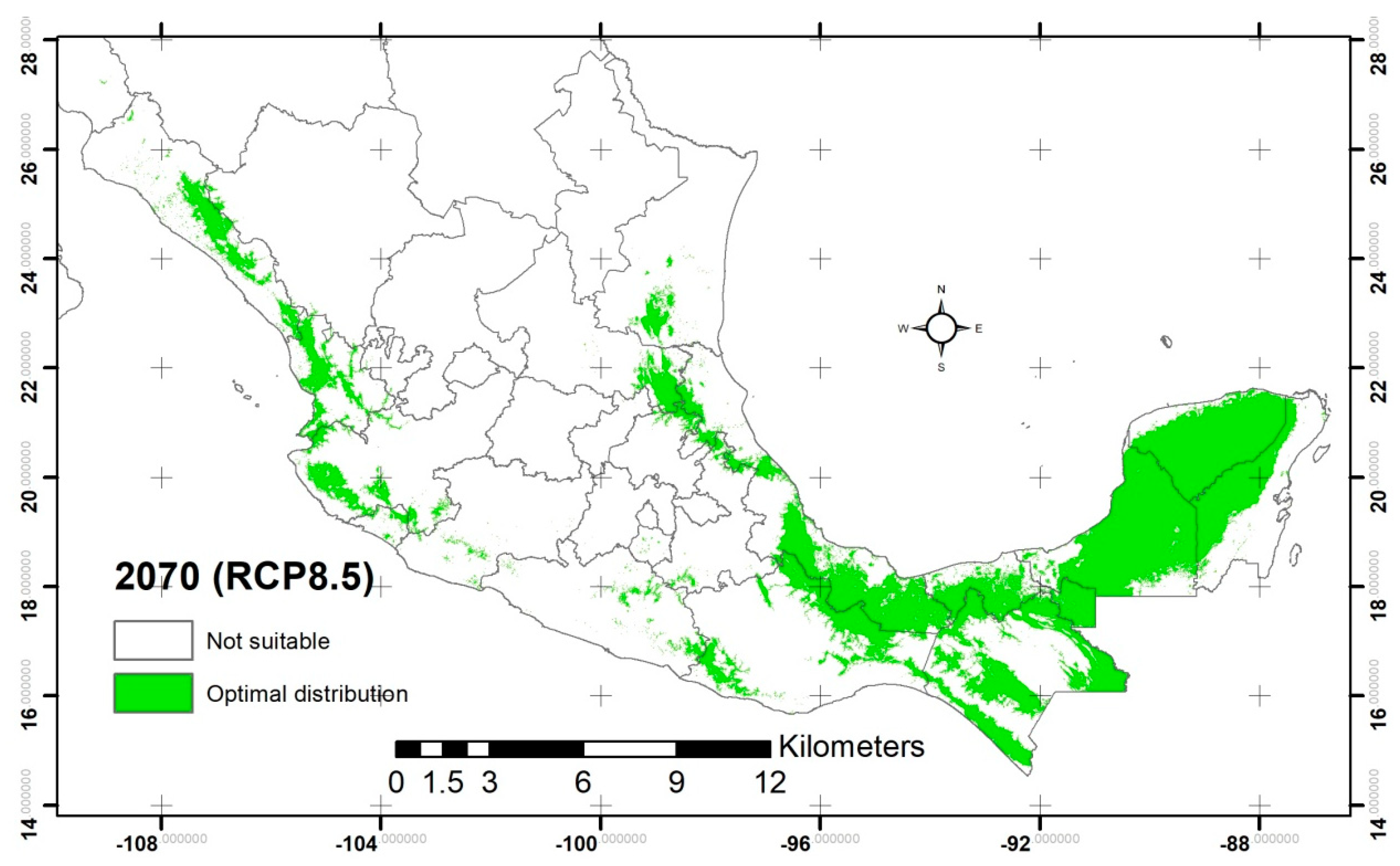
2.2. Potential Distribution of C. odorata
3. Discussion
3.1. Current Distribution of C. odorata
3.2. Potential Distribution of C. odorata
4. Materials and Methods
4.1. Study Species
4.2. Study Area
4.3. Data Acquisition
4.4. Climate Variables
4.5. Distribution Model and Optimum Germination Temperature Range
4.6. Modeling Current Distributions of C. odorata
4.7. Model of Potential Distribution of C. odorata
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franks, S.J.; Sim, S.; Weis, A.E. Rapid evolution of flowering time by an annual plant in response to a climate fluctuation. Proc. Natl. Acad. Sci. USA 2007, 104, 1278–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craufurd, P.Q.; Wheeler, T.R. Climate change and flowering time in annual crops. J. Exptl. Bot. 2009, 60, 2529–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, J.T.; Inouye, D.W.; McKinney, A.M.; Colauti, R.I.; Mitchell-Olds, T. Pheotypic plasticity and adaptive evolution contribute to advancing flowering phenology in response to climate change. Proc. R. Soc. B 2012, 279, 3843–3852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iler, A.M.; Høye, T.K.; Inouye, D.W.; Schmidt, N.M. Nonlinear flowering responses to climate: Are species approaching their limits of phonological change? Phil. Trans. R. Soc. B 2013, 368, 20120489. [Google Scholar] [CrossRef] [PubMed]
- Gaira, K.S.; Rawal, R.S.; Rawat, B.; Bhatt, I.D. Impact of climate change on the flowering of Rhododendron arboretum in central Himalaya, India. Curr. Sci. 2014, 106, 1735–1738. [Google Scholar]
- Wang, B.; Liu, D.L.; Asseng, S.; Macadam, I.; Yu, Q. Impact of climate change on wheat flowering time in eastern Australia. Agric. Forest. Meteorol. 2015, 209, 11–21. [Google Scholar] [CrossRef]
- Sampayo-Maldonado, S.; Ordoñez-Salanueva, C.A.; Mattana, E.; Ulian, T.; Way, M.; Castillo, E.; Davila-Aranda, P.D.; Lira-Sadde, R.; Téllez-Valdéz, O.; Rodríguez-Arevalo, N.I.; et al. Thermal time and cardinal temperatures for germination of Cedrela odorata L. Forests 2019, 10, 841. [Google Scholar] [CrossRef] [Green Version]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Root, T.L.; MacMynowski, D.P.; Mastrandrea, M.D.; Schneider, S.H. Human modified temperature induce species changes: Joint attribution. Proc. Natl. Acad. Sci. USA 2005, 102, 7465–7469. [Google Scholar] [CrossRef] [Green Version]
- Iverson, L.R.; Prasad, A.M.; Schwartz, M.W. Predicting potential changes in suitable habitat and distribution by 2100 for tree species in the eastern United States. J. Agric. Meteorol. 2005, 61, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Iverson, L.R.; Prasad, A.M.; Matthews, S.N.; Peters, M. Estimating potential habitat for 134 Eastern US tree species under six climate scenarios. For. Ecol. Manage 2008, 254, 390–406. [Google Scholar] [CrossRef]
- McKenney, D.W.; Pedlar, J.H.; Lawrence, K.; Campbell, K.; Hutchison, M.F. Potential impacts of climate change on the distribution of North American trees. Bioscience 2007, 57, 939–948. [Google Scholar] [CrossRef]
- Kelly, A.E.; Goulden, M.L. Rapid shifts in plant distribution with recent climate change. Proc. Natl. Acad. Sci. USA 2007, 105, 11823–11826. [Google Scholar] [CrossRef] [Green Version]
- Thuiller, W.; Albert, C.; Araújo, M.B. Predicting global change impacts on plant species distributions: Future challenges. Perspect. Plant. Ecol. Evol. Syst. 2008, 9, 137–152. [Google Scholar] [CrossRef]
- Pautasso, M. Forest ecosystems and global change: The case study of insurbia. Ann. Bot. 2013, 3, 1–29. [Google Scholar]
- Iverson, L.R.; McKenzie, D. Tree-species range shifts in the changing climate: Detecting, modeling, assisting. Landsc. Ecol. 2013, 28, 879–889. [Google Scholar] [CrossRef]
- Kurpis, J.; Serrato-Cruz, M.A.; Arroyo, T.P.F. Modeling the effects of climate change on the distribution of Tagetes lucida Cav. (Asteraceae). Glob. Ecol. Conserv. 2019, 20, e00747. [Google Scholar] [CrossRef]
- Walk, J.L.; Hidayati, S.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate change and plant regeneration from seed. Glob. Chang. Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Fernández-Pascual, E.; Mattana, E.; Pritchard, H.W. Seeds for future past: Climate change and the thermal memory of plant reproductive traits. Biol. Rev. 2019, 94, 439–456. [Google Scholar] [CrossRef]
- Romo-Lozano, J.L.; Vargas-Hernández, J.; López-Upton, J.; Ávila-Angulo, M.L. Estimate of the financial value of timber stocks of red cedar (Cedrela odorata L) in Mexico. Madera y Bosques 2017, 23, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Sampayo-Maldonado, S.; Jiménez-Casas, M.; López-Upton, J.; Sánchez-Monsalvo, V.; Jasso-Mata, J.; Equihua-Martínez, A.; Castillo-Martínez, C.R. Cedrela odorata L. mini-cutting rooting. Agrociencias 2016, 50, 919–929. [Google Scholar]
- Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES). In Proceedings of the CoP14 Prop. 33, Fourteenth Meeting of the Conference of the Parties, The Hague, The Netherlands, 3–15 June 2007.
- Secretaria del Medio Ambiente y Recursos Naturales (SEMARNAT). NORMA Oficial Mexicana NOM-059-SEMARNAT-2010, Protección Ambiental -Especies Nativas de México de Flora & Fauna Silvestres- Categorías de Riesgo & Especificaciones Para su Inclusión, Exclusión o cambio-Lista de Especies en Riesgo. Available online: http://www.cec.org/wp-content/uploads/wpallimport/files/18-2-det1412_es.pdf (accessed on 12 September 2022).
- Chun-Jing, W.; Zhi-Xiang, Z.; Ji-Zhong, W. Vulnerability of global Forest ecoregions to future climate. Glob. Ecol. Conserv. 2019, 20, 1–10. [Google Scholar] [CrossRef]
- Gómez-Díaz, J.D.; Monterroso-Rivas, A.I.; Tinoco-Rueda, J.A. Distribution of red cedar (Cedrela odorata L.) in the Hidalgo state, under current conditions and scenarios of climate change. Madera y Bosques 2007, 13, 29–49. [Google Scholar] [CrossRef]
- Hernández-Ramos, J.; Reynoso-Santos, R.; Hernández-Ramos, A.; García-Cuevas, X.; Hernández-Máximo, E.; Cob-Uicab, J.V.; Sumano-López, D. Historical, current and future distribution of Cedrela odorata in Mexico. Acta Bot. Mex. 2018, 124, 117–134. [Google Scholar] [CrossRef]
- CICS. Pridicting Species Distribution Use of Climatic Parametrs. Canadian Institute for Climate Studies. 2006. Available online: http://www.cics.uvic.ca/ (accessed on 13 December 2022).
- Duan, X.; Li, J.; Wu, S. MaxEnt Modeling to Estimate the Impact of Climate Factors on Distribution of Pinus densiflora. Forests 2022, 13, 402. [Google Scholar] [CrossRef]
- Duval, V.S.; Benedetti, G.M.; Campo, A.M. Relación clima-vegetación: Adaptaciones de la comunidad del jarillal al clima semiárido, Parque Nacional Lihué Calel, provincia de la Pampa, Argentina. Investig. Geog. 2015, 88, 33–44. [Google Scholar] [CrossRef] [Green Version]
- Funes, G.; Díaz, S.; Venier, P. La temperatura como principal determinante de la germinación en especies del Chaco seco de Argentina. Ecol. Austral 2009, 19, 129–138. [Google Scholar]
- Durán-Puga, N.; Ruíz-Corral, J.A.; González-Eguiarte, D.R.; Núñez-Hernández, G.; Padilla-Ramírez, F.J.; Contreras-Rodríguez, S.H. Development cardinal temperatures of the planting-emergence stage for 11 forage grasses. Rev. Mex. Cienc. Pecu. 2011, 2, 347–357. [Google Scholar]
- Nakao, E.A.; Cardoso, V.J.M. Analysis of thermal dependence on the germination of braquiarão seeds using the thermal time model. Braz. J. Biol. 2016, 76, 162–168. [Google Scholar] [CrossRef] [Green Version]
- González, J.E. Seed collection and germination of 26 tree species from the tropical rain forest. Rev. Biol. Trop. 1991, 39, 47–51. [Google Scholar] [CrossRef]
- Garcia-Huidobro, J.; Monteith, J.L.; Squire, G.R. Time, temperature and germination of pearl millet (Pennisetum typhoides S. & H.). I. Constant temperature. J. Exp. Bot. 1982, 33, 288–296. [Google Scholar]
- Gilbertson, P.K.; Berti, M.T.; Johnson, B.L. Borage cardinal germination temperatures and seed development. Ind. Crops Prod. 2014, 59, 202–209. [Google Scholar] [CrossRef]
- Calzada-López, S.G.; Kohashi-Shibata, J.; Uscanga-Mortera, E.; García-Esteva, A.; Yáñez-Jiménez, P. Cardinal temperatures and germination rate in husk tomato cultivars. Rev. Mex. Cienc. Agrí. 2014, 8, 1451–1458. [Google Scholar]
- Ruíz-Corral, J.A.; Flores-López, H.E.; Ramírez-Díaz, J.L.; González-Eguiarte, D.R. Cardinal temperatures and length of maturation cycle of maize hybrid H-311 under rainfed conditions. Agrociencia 2002, 36, 569–577. [Google Scholar]
- Pernús-Alvarez, M.; Sánchez-Rendón, J.A. Efecto de la procedencia sobre rasgos seminales y requerimientos germinativos de Cedrela odorata (Meliaceae). Revista Jard. Bot. Nac. 2016, 37, 203–211. [Google Scholar]
- Adam, N.R.; Dierig, D.A.; Coffelt, T.A.; Wintermeyer, M.J.; Mackey, B.E.; Wall, G.W. Cardinal temperatures for germination and early growth of two Lesquerella species. Ind. Crops Prod. 2007, 25, 24–33. [Google Scholar] [CrossRef]
- Hardegree, S.P. Predicting germination response to temperature. I. Cardinal temperature models and subpopulation-specific regression. Ann. Bot. 2006, 97, 1115–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Sáenz-Romero, C.; Rehfeldt, G.E.; Crookston, N.L.; Duval, P.; St-Amant, R.; Beaulieu, J.; Richardson, B.A. Spline models of contemporary, 2030, 2060 and 2090 climates for Mexico and their use in understanding climate-plant impacts on vegetation. Clim. Chang. 2010, 102, 595–623. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Rehfeldt, G.E.; Duval, P.; Lindig-Cisneros, R.A. Abies religiosa habitat prediction in climatic change scenarios and implications for monarch butterfly conservation in Mexico. For. Ecol. Manag. 2012, 275, 98–106. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Rehfeldt, G.E.; Ortega-Rodríguez, J.M.; Marín-Togo, M.C.; Madrigal-Sánchez, X. Pinus leiophylla suitable habitat for 1961-1990 and future climate. Bot. Sci. 2015, 93, 709–718. [Google Scholar] [CrossRef] [Green Version]
- Garza-López, M.; Ortega-Rodríguez, L.M.; Zamudio-Sánchez, F.J.; López-Toledo, J.F.; Domínguez-Álvarez, F.A.; Sáenz-Romero, C. Calakmul como refugio de Swietenia macrophylla King ente el cambio climático. Bot. Sci. 2016, 94, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Sampayo-Maldonado, S.; Ordoñez-Salanueva, C.A.; Mattana, E.; Way, M.; Castillo-Lorenzo, E.; Dávila-Aranda, P.D.; Lira-Sadde, R.; Téllez-Valdéz, O.; Rodríguez-Arevalo, N.I.; Ulian, T.; et al. Thermal Niche for Seed Germination and Species Distribution Modelling of Swietenia macrophylla King (Mahogany) under Climate Change Scenarios. Plants 2021, 10, 2377. [Google Scholar] [CrossRef] [PubMed]
- Villaseñor, J.L.; Ortiz, E. Biodiversity of flowering plants (Division Magnoliophyta) in Mexico. Rev. Mex. Biodivers. 2014, 85, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, H.T.; Kester, D.E.; Davies, F.T.; Geneve, R.L. Plant Propagation: Principles and Practices, 8th ed.; Prentice-Hall: Hoboken, NJ, USA, 2013; 913p. [Google Scholar]
- Beigaité, R.; Tang, H.; Bryn, A.; Skarpaas, O.; Stordal, F.; Bjerke, J.W.; Zliobaite, I. Identifying climate thresholds for dominant natural vegetation types at the global scale using machine learning: Average climate versus extremes. Glob. Chang. Biol. 2022, 28, 3557–3579. [Google Scholar] [CrossRef]
- Leal-Narez, O.; Mendoza, M.E.; Pérez-Salicrup, D.; Geneletti, D.; López-Granados, E.; Carranza, E. Distribución potencial de Pinus Martinezii: Un modelo espacial basado en conocimiento ecológico y análisis multicriterio. Rev. Mex. Biodivers. 2012, 83, 1152–1170. [Google Scholar] [CrossRef]
- Gutiérrez, E.; Trejo, I. Effect of climatic change on the potential distribution of five species of temperate forest trees in Mexico. Rev. Mex. Biodivers. 2014, 85, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Parmoon, G.; Moosavi, S.A.; Akbari, H.; Ebadi, A. Quantifying cardinal temperatures and thermal time required for germination of Silybum marianum seed. Crop J. 2015, 3, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Daibes, L.F.; Amoêdo, S.C.; Nascimento-Moraes, J.; Fenelon, N.; da Silva, D.R.; de Melo-Lopes, M.J.; Vargas, L.A.; Monteiro, E.F.; Frigeri, R.B.C. Thermal requirements of seed germination of ten tree species occurring in the western Brazilian Amazon. Seed Sci. Res. 2019, 29, 115–123. [Google Scholar] [CrossRef]
- Ortiz-Solorio, C.A. Elementos de Agrometeorología Cuantitativa Con Aplicaciones en la República Mexicana; Universidad Autónoma Chapingo: Texcoco, Mexico, 1987; 327p. [Google Scholar]
- Fremout, T.; Thomas, E.; Gaisberger, H.; Van-Meerbeek, K.; Muenchow, J.; Briers, S.; Gutierrez-Miranda, C.E.; Marcelo-Peña, J.L.; Kindt, R.; Atkinson, R.; et al. Mapping tree species vulnerability to multiple threats as a guide to restoration and conservation of tropical dry forests. Glob. Chang. Biol. 2020, 26, 3552–3568. [Google Scholar] [CrossRef]
- Pennington, T.D.; Sarukhán, J. Tropical trees of Mexico: Manual for the Identification of the Main Species, 3rd ed.; University Scientific Text; UNAM: Mexico City, Mexico, 2005; pp. 294–296. [Google Scholar]
- Mendizábal-Hernández, L.C.; Alba-Landa, J.; Márquez-Ramírez, J.; Cruz-Jiménez, H.; Ramírez-García, E.O. Carbon sequestration by Cedrela odorata L. in a genetic trial. Rev. Mex. Cienc. Forest. 2011, 2, 107–113. [Google Scholar]
- Instituto Nacional de Estadística y Geografía (INEGI). Conjunto de Datos Vectoriales Edafológicos, Escala 1: 250,000 Serie IV; Continuo Nacional, Escala: 1: 250,000 Ed. 2; Instituto Nacional de Estadística y Geografía: Aguascalientes, Mexico, 2014. [Google Scholar]
- Global Biodiversity Information Facility. GBIF Ocurrence Cedrela odorata L. Available online: https://api.gbif.org/v1/occurrence/download/request/0216476-220831081235567.zip (accessed on 18 November 2021).
- Instituto Nacional de Estadística y Geografía (INEGI). Conjunto De Datos Vectoriales de Uso Del Suelo Y Vegetación. Escala 1: 250,000 Serie VI; Continuo Nacional, Escala: 1: 250,000 Ed. 2; Instituto Nacional de Estadística y Geografía: Aguascalientes, Mexico, 2018. [Google Scholar]
- Hijmans, R.j.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very higt resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- WorldClim. Free Climate Data for Ecological Modeling and GIS. WorldClim-Global Climate Data. 2006. Available online: http://www.worldclim.org/ (accessed on 18 November 2021).
- Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). Red mundial de información sobre biodiversidad (REMIB). Comisión Nacional para el Conocimiento & Uso de la Biodiversidad. Mexico City, México. 2015. Available online: https://www.academia.edu/23597234/Comisi%C3%B3n_Nacional_para_el_Conocimiento_y_Uso_de_la_Biodiversidad_Comisi%C3%B3n_Nacional_de_%C3%81reas_Naturales_Protegidas (accessed on 25 November 2020).
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of MaxEnt. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Baldwin, R.A. Use of maximum entropy modeling in wildlife research. Entropy 2009, 11, 854–866. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistic explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modelling. Ecol. Modell. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Fernández-Eguiarte, A.; Zavala-Hidalgo, J.; Romero-Centeno, R.; Conde-Álvarez, A.C.; Trejo-Vázquez, R.I. Actualización de los escenarios de Cambio Climático Para Estudios de Impactos, Vulnerabilidad y Adaptación en México y Centroamérica; Centro de Ciencias de la Atmósfera, Universidad Nacional Autónoma de México. Instituto Nacional de Ecología y Cambio Climático: Mexico City, Mexico, 2015; 22p. [Google Scholar]
- Weyant, J.; Azar, C.; Kainuma, M.; Kejun, J.; Nakicenovic, N.; Shukla, P.R.; La Rovere, E.; Yohe, G. Future Intergovernmental Panel on Climate Change activities- New scenarios. Report of 2.6 Versus 2.9 Watts/m2 Representative Concentration Pathways (RCP´s) Evaluation Panel. In Proceedings of the Intergovernmental Panel on Climate Change (IPCC)-XXX/INF.6, Antalya, Turkey, 21–23 April 2009. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| State | Current (km2) | 2050 (km2) | 2070 (km2) | ||
|---|---|---|---|---|---|
| RCP2.6 | RCP8.5 | RCP2.6 | RCP8.5 | ||
| Campeche | 49,206.37 | 46,696.03 | 46,596.33 | 45,717.05 | 46,299.80 |
| Chiapas | 26,474.27 | 31,882.02 | 32,529.20 | 32,424.94 | 33,982.65 |
| Chihuahua | 433.45 | 28.61 | 2.23 | 16.59 | - |
| Colima | 335.29 | 572.07 | 208.92 | 475.87 | 929.28 |
| Durango | 1508.06 | 1128.22 | 773.41 | 1101.02 | 759.43 |
| Guanajuato | - | 15.38 | 6.64 | 65.10 | 2.70 |
| Guerrero | 6033.25 | 2138.62 | 2268.99 | 2150.80 | 2684.70 |
| Hidalgo | 668.78 | 1867.85 | 1566.70 | 1831.65 | 1755.27 |
| Jalisco | 5044.10 | 8603.37 | 8863.37 | 9131.17 | 8990.25 |
| Mexico | 433.54 | 67.76 | 80.22 | 46.47 | 4.41 |
| Michoacan | 435.00 | 1391.38 | 409.87 | 1,301.65 | 798.17 |
| Morelos | 802.23 | 122.15 | 70.71 | 62.82 | 72.15 |
| Nayarit | 8899.30 | 8895.37 | 9016.18 | 9022.26 | 8872.49 |
| Oaxaca | 16,410.20 | 20,896.92 | 20,852.32 | 18,317.95 | 19,579.65 |
| Puebla | 1341.35 | 848.88 | 1368.41 | 1309.98 | 1862.81 |
| Queretaro | 71.61 | 137.77 | 96.53 | 158.12 | 65.26 |
| Quintana Roo | 20,136.19 | 17,307.75 | 16,330.78 | 15,596.01 | 18,351.02 |
| San Luis Potosi | 6848.31 | 4888.47 | 5112.18 | 4445.60 | 5200.63 |
| Sinaloa | 6586.40 | 9883.49 | 9728.92 | 9754.78 | 9823.81 |
| Sonora | 16.45 | 11.42 | 32.58 | 3.78 | 31.98 |
| Tabasco | 14,217.66 | 17,776.39 | 16,326.25 | 18,410.85 | 16,609.55 |
| Tamaulipas | 4623.80 | 3757.83 | 3467.14 | 3779.01 | 3526.20 |
| Veracruz | 24,716.46 | 31,145.75 | 31,821.38 | 31,705.65 | 32,537.99 |
| Yucatan | 33,538.96 | 33,827.35 | 34,003.95 | 33,109.55 | 33,941.25 |
| Suitability area (km2) | 228,780.66 | 243,890.85 | 241,533.21 | 239,938.67 | 246,681.45 |
| Increment area (km2) | N/A | 15,110.19 | 12,752.55 | 11,158.01 | 17,900.79 |
| Percentage of increment (%) | N/A | 6.60 | 5.57 | 4.87 | 7.82 |
| Abbreviation | Environmental Variable | Units | Used for Modeling |
|---|---|---|---|
| Bio1 | Annual average temperature | (°C) | No |
| Bio2 | Diurnal temperature oscillation | (°C) | Yes |
| Bio3 | Isothermality (Bio2/Bio7) × 100 | (°C) | No |
| Bio4 | Temperature seasonality (standard deviation × 100) | (°C) | Yes |
| Bio5 | Maximum average temperature of the warmest period | (°C) | No |
| Bio6 | Minimum temperature of the coldest month | (°C) | Yes |
| Bio7 | Annual temperature oscillation (Bio5–Bio6) | (°C) | Yes |
| Bio8 | Average temperature of the wettest month | (°C) | No |
| Bio9 | Average temperature of the driest month | (°C) | Yes |
| Bio10 | Average temperature of the warmest quarter | (°C) | No |
| Bio11 | Average temperature of the coldest quarter | (°C) | No |
| Bio12 | Annual precipitation | (mm) | Yes |
| Bio13 | Precipitation during the wettest period | (mm) | No |
| Bio14 | Precipitation during the driest period | (mm) | Yes |
| Bio15 | Precipitation seasonality (coefficient of variation) | CV | No |
| Bio16 | Precipitation during the wettest trimester | (mm) | No |
| Bio17 | Precipitation during the driest trimester | (mm) | No |
| Bio18 | Precipitation during the warmest trimester | (mm) | No |
| Bio19 | Precipitation during the coldest trimester | (mm) | No |
| Altitude | Elevation | (m) | No |
| Soils | Soil type | - | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sampayo-Maldonado, S.; Ordoñez-Salanueva, C.A.; Mattana, E.; Way, M.; Castillo-Lorenzo, E.; Dávila-Aranda, P.D.; Lira-Saade, R.; Téllez-Valdés, O.; Rodríguez-Arévalo, N.I.; Flores-Ortiz, C.M.; et al. Potential Distribution of Cedrela odorata L. in Mexico according to Its Optimal Thermal Range for Seed Germination under Different Climate Change Scenarios. Plants 2023, 12, 150. https://doi.org/10.3390/plants12010150
Sampayo-Maldonado S, Ordoñez-Salanueva CA, Mattana E, Way M, Castillo-Lorenzo E, Dávila-Aranda PD, Lira-Saade R, Téllez-Valdés O, Rodríguez-Arévalo NI, Flores-Ortiz CM, et al. Potential Distribution of Cedrela odorata L. in Mexico according to Its Optimal Thermal Range for Seed Germination under Different Climate Change Scenarios. Plants. 2023; 12(1):150. https://doi.org/10.3390/plants12010150
Chicago/Turabian StyleSampayo-Maldonado, Salvador, Cesar A. Ordoñez-Salanueva, Efisio Mattana, Michael Way, Elena Castillo-Lorenzo, Patricia D. Dávila-Aranda, Rafael Lira-Saade, Oswaldo Téllez-Valdés, Norma I. Rodríguez-Arévalo, Cesar M. Flores-Ortiz, and et al. 2023. "Potential Distribution of Cedrela odorata L. in Mexico according to Its Optimal Thermal Range for Seed Germination under Different Climate Change Scenarios" Plants 12, no. 1: 150. https://doi.org/10.3390/plants12010150
APA StyleSampayo-Maldonado, S., Ordoñez-Salanueva, C. A., Mattana, E., Way, M., Castillo-Lorenzo, E., Dávila-Aranda, P. D., Lira-Saade, R., Téllez-Valdés, O., Rodríguez-Arévalo, N. I., Flores-Ortiz, C. M., & Ulian, T. (2023). Potential Distribution of Cedrela odorata L. in Mexico according to Its Optimal Thermal Range for Seed Germination under Different Climate Change Scenarios. Plants, 12(1), 150. https://doi.org/10.3390/plants12010150