
Antiviral Activity of Selected Essential Oils against Cucumber Mosaic Virus

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. The Composition of EOs
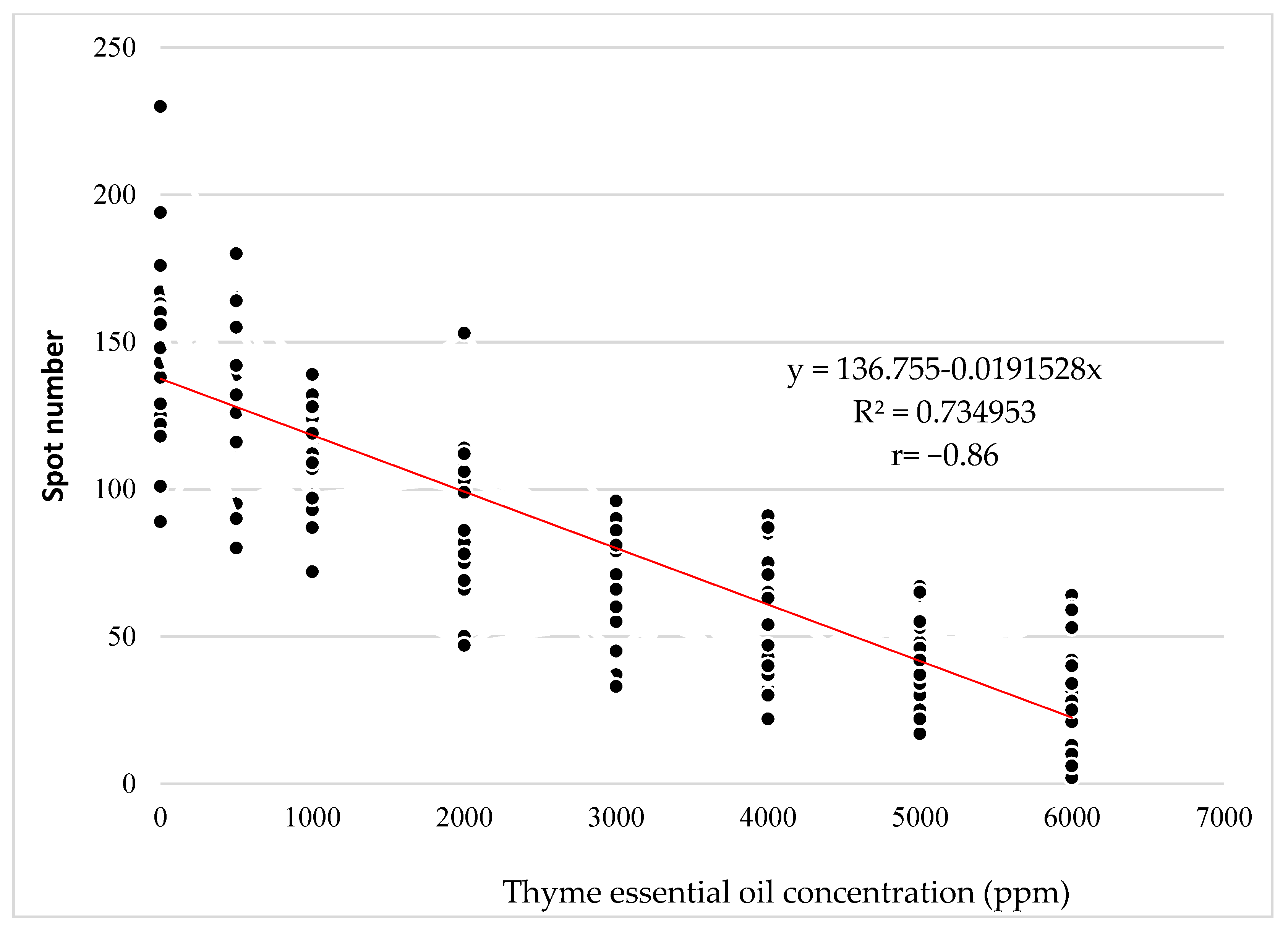
2.2. Evaluation of the Antiviral Activity of the EOs against the CMV-S21 Isolate (In Vitro)
2.3. Evaluation of the Antiviral Activity of the EOs against the CMV-S21 Isolate (In Vivo)
3. Discussion
4. Materials and Methods
4.1. Plant Raw Materials Used for Distillation of EOs
4.2. EOs Extraction and GC-MS/GC-FID Analysis
4.3. Virus and Plant Host
4.4. Antiviral Effects of the EOs on the Ch. Quinoa Plants
4.5. DAS-ELISA Test
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Islam, W.; Zaynab, M.; Qasim, M.; Wu, Z. Plant-virus interaction disease resistance in Focus. Hosts Viruses 2017, 4, 5–20. [Google Scholar] [CrossRef]
- Taglienti, A.; Donati, L.; Ferretti, L.; Tomassoli, L.; Sapienza, F.; Sabatino, M.; Di Massimo, G.; Fiorentino, S.; Vecchiarelli, V.; Nota, P.; et al. In vivo antiphytoviral activity of essential oils and hydrosols from Origanum vulgare, Thymus vulgaris, and Rosmarinus officinalis to control Zucchini Yellow Mosaic Virus and Tomato Leaf Curl New Delhi Virus in Cucurbita pepo L. Front. Microbiol. 2022, 13, 840893. [Google Scholar] [CrossRef] [PubMed]
- Reichling, J.; Schnitzler, P.; Suschke, U.; Saller, R. Essential oils of aromatic plants with antibacterial, antifungal, antiviral, and cytotoxic properties—An overview. Forsch. Komplementärmedizin 2009, 16, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezić, N.; Dunkić, V.; Vuko, E. Antiphytoviral activity of essential oils of some Lamiaceae species and there most important compounds on CMV and TMV. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Méndez-Vilas, A., Ed.; FORMATEX: Badajoz, Spain, 2013; pp. 982–987. [Google Scholar]
- Akthar, M.S.; Degaga, B.; Azam, T. Antimicrobial activity of essential oils extracted from medicinal plants against the pathogenic microorganisms: A review. Issues Biol. Sci. Pharm. Res. 2014, 2, 1588. [Google Scholar]
- Arshad, Z.; Hanif, M.A.; Qadri, R.W.K.; Khan, M. Role of essential oils in plant diseases protection: A review. IJCBS 2014, 6, 11–17. [Google Scholar]
- Șener, B. Antiviral activityof naural products and herbal extracts. GMJ 2020, 31, 474–477. [Google Scholar] [CrossRef]
- Islam, W.; Adnan, A.; Tayyab, M.; Hussain, M.; Islama, S.U. Phyto-metabolites; An Impregnable Shield against Plant Viruses. Nat. Prod. Commun. 2018, 13, 41–51. [Google Scholar] [CrossRef]
- Shukla, H.S.; Dubey, P.; Chaturvedi, R.V. Antiviral properties of essential oils of Foeniculum vulgare and Pimpinella anisum L. Agronomie 1989, 9, 277–279. [Google Scholar] [CrossRef] [Green Version]
- Othman, B.A.; Shoman, S.A. Antiphytoviral activity of the Plectranthus tenuiflorus on some important viruses. Int. J. Agric. Biol. 2004, 6, 844–849. [Google Scholar]
- Chen, J.; Yan, X.-H.; Dong, J.-H.; Sang, P.; Fang, X.; Di, Y.-T.; Zhang, Z.-K.; Hao, X.-J. Tobacco mosaic virus (TMV) inhobitors from Picrasma quassioides Benn. J. Agric. Food Chem. 2009, 57, 6590–6595. [Google Scholar] [CrossRef]
- Dunkić, V.; Bezić, N.; Vuko, E.; Cukrov, D. Antiphytoviral Activity of Satureja montana L. ssp. Variegata (Host) P. W. Ball Essential Oil and Phenol Compounds on CMV and TMV. Molecules 2010, 15, 6713–6721. [Google Scholar] [CrossRef] [Green Version]
- Jerković-Mujkić, A.; Mahmutović, I.; Bešta-Gajević, R. Antiphytoviral Effects of Three Different Essential Oils on Tobacco mosaic virus; Works of the Faculty of Forestry; University of Sarajevo: Sarajevo, Yugoslavia, 2013; Volume 2, pp. 41–51. [Google Scholar] [CrossRef]
- Min, L.; Han, Z.; Xu, Y.; Yao, L. In vitro and in vivo anti-Tobacco mosaic virus activities of essential oils and individual compounds. J. Microbiol. Biotechnol. 2013, 23, 771–778. [Google Scholar] [CrossRef]
- Elsharkawy, M.M.; El-Sawy, M.M. Control of Bean common mosaic virus by plant extracts in bean plants. Int. J. Pest Manag. 2015, 61, 54–59. [Google Scholar] [CrossRef]
- Petrov, N.; Stoyanova, M.; Valkova, M. Antiviral activity of plant extract from Tanacetum vulgare against Cucumber mosaic virus and Potato virus Y. J. BioSci. Biotechnol. 2016, 5, 189–194. [Google Scholar]
- Dikova, B.; Dobreva, A.; Djurmanski, A. Essential oils of Lavender and fennel for inhibiting Tomato Spotted Wilt Virus in pepper plants. Acta Microbiol. Bulg. 2017, 33/1, 36–43. [Google Scholar]
- Elbeshehy, E.K.F. Inhibitor of different medicinal plants extracts from Thuja orientalis, Nigell sativa L., Azadirachta indica and Bougainvillea spectabilis against Zuchcchini yellow mosaic virus (ZYMV) infecting Citrullus lanatus. Biotechnol. Biotechnol. Equip. 2017, 31, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Helal, I.M. Use of biocides for controlling viral diseases that attack common bean and cucumber plants. Folia Hortic. 2019, 31, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Kowalczyk, A.; Przychodna, M.; Sopata, S.; Bodalska, A.; Fecka, I. Thymol and Thyme essential oil—New insights into selected therapeutic applications. Molecules 2020, 25, 4125. [Google Scholar] [CrossRef]
- Król, S.K.; Skalicka-Woźniak, K.; Kandefer-Szerszeń, M.; Stepulak, A. Aktywność biologiczna i farmakologiczna olejków eterycznych w leczeniu i profilaktyce chorób infekcyjnych. Postępy Hig. Med. Doświadczalnej 2013, 67, 1000–1007. [Google Scholar] [CrossRef]
- Gibbs, A.J.; Harrison, B.D. Cucumber mosaic virus. In CMI/AAB Descriptions of Plants Viruses; Commonwealth Mycological Institute: Surrey, UK, 1970; No. 1. [Google Scholar]
- Dunkić, V.; Vuko, E.; Bezić, N.; Kremer, D.; Ruščić, M. Composition and Antiviral Activity of the Essential Oils of Eryngium alpinum and E. amethystinum. Chem. Biodivers. 2013, 10, 1894–1902. [Google Scholar] [CrossRef]
- Bezić, N.; Vuko, E.; Dunkić, V.; Ruščić, M.; Blažević, I.; Burčul, F. Antiphytoviral activity of sesquiterpene-rich essential oils from four Croatian Teucrium species. Molecules 2011, 16, 8119–8129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negai, A.; Duarte Lígia, M.L.; Santos Déborah, Y.A.C. Influence of viral infection on essential oil composition of Ocimum basilicum (Lamiaceae). Nat. Prod. Commun. 2011, 6, 1189–1192. [Google Scholar] [CrossRef] [Green Version]
- Vuko, E.; Dunkić, V.; Bezić, N.; Ruščić, M.; Kremer, D. Chemical composition and antiphytoviral activity of essential oil of Micromeria graeca. Nat. Prod. Commun. 2012, 7, 1227–1230. [Google Scholar] [CrossRef] [Green Version]
- Hamidson, H.; Damiri, N.; Angraini, E. Effect of medicinal plants extracts on the incidence of mosaic disease caused by Cucumber mosaic virus and growth of chili. IOP Conf. Ser. Earth Environ. Sci. 2018, 102, 012062. [Google Scholar] [CrossRef]
- Ruščić, M.; Vuko, E.; Bezić, N.; Kremer, D.; Dunkić, V. Croatian Micromeria fruticulose—Essential oil composition and antiphytoviral activity. Nat. Prod. Commun. 2017, 12, 1133–1135. [Google Scholar] [CrossRef] [Green Version]
- Vuko, E.; Rusak, G.; Dunkić, V.; Kremer, D.; Kosalec, I.; Rađa, B.; Bezić, N. Inhibition of satellite RNA associated Cucumber mosaic virus infection by essential oil of Micromeria croatica (Pers.) Schott. Molecules 2019, 24, 1342. [Google Scholar] [CrossRef] [Green Version]
- Babushok, V.I.; Linstrom, P.J.; Zenkevich, I.G. Retention indices for frequently reported compounds of plant essential oils. J. Phys. Chem. Ref. Data 2011, 40, 043101. [Google Scholar] [CrossRef] [Green Version]
- Barra, A. Factors affecting chemical variability of essential oils: A review of recent developments. Nat. Prod. Commun. 2009, 4, 1147–1154. [Google Scholar] [CrossRef] [Green Version]
- Bylaite, E.; Venscutonis, R.; Roozen, J.P.; Posthumus, M.A. Composition of essential oil of costmary [Balsamita major (L.) desf.] at different growth phases. J. Agric. Food Chem. 2000, 48, 2409–2414. [Google Scholar] [CrossRef]
- De Martino, L.; De Feo, V.; Formisano, C.; Mignola, E.; Senatore, F. Chemical composition and antimicrobial activity of the essential oils from three chemotypes of Origanum vulgareL. ssp. hirtum (Link) Ietswaart growing wild in Campania (Southern Italy). Molecules 2009, 14, 2735–2746. [Google Scholar] [CrossRef] [Green Version]
- Hassanpouraghdam, M.B.; Tabatabaie, S.J.; Nazemiyeh, H.; Vojodi, L.; Aazami, M.A. Volatile oil constituents of alecost [Tanacetum balsamita L. ssp. balsamitoides (Schultz-Bip.)] growing wild in North-West of Iran. Herba Pol. 2009, 55, 53–59. [Google Scholar]
- Bączek, K.; Kosakowska, O.; Przybył, J.; Pióro-Jabrucka, E.; Costa, R.; Luigi Mondello, L.; Gniewosz, M.; Synowiec, A.; Węglarz, Z. Antibacterial and antioxidant activity of essential oils and extracts from costmary (Tanacetum balsamita L.) and tansy (Tanacetum vulgare L.). Ind. Crops Prod. 2017, 102, 154–163. [Google Scholar] [CrossRef]
- Węglarz, Z.; Kosakowska, O.; Przybył, J.L.; Pióro-Jabrucka, E.; Bączek, K. The Quality of Greek Oregano (O. vulgare L. subsp. hirtum (Link) Ietswaart) and Common Oregano (O. vulgare L. subsp. vulgare) Cultivated in the Temperate Climate of Central Europe. Foods 2020, 9, 1671. [Google Scholar] [CrossRef]
- European Pharmacopoeia. European Directorate for the Quality of Medicines and Health Care (EDQM), 9th ed.; Council of Europe: Strasbourg, France, 2017. [Google Scholar]
- Bączek, K.; Kosakowska, O.; Przybył, J.L.; Kuźma, P.; Ejdys, M.; Obiedziński, M.; Węglarz, Z. Intraspecific variability of yarrow (Achillea millefolium L. s.l.) in respect of developmental and chemical traits. Herba Pol. 2015, 61, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Clark, M.F.; Adams, A.N. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Relative Area (%) | ||||||
|---|---|---|---|---|---|---|
| No. | Compound | RI 1 | RI 2 | Greek Oregano | Thyme | Costmary |
| 1 | (Z)-salvene | 935 | - | - | - | 0.28 ± 0.02 |
| 2 | (E)-salvene | 949 | - | - | - | 0.05 ± 0.01 |
| 3 | α-thujene | 1023 | 1012–1039 | 0.21 ± 0.02 | 0.33 ± 0.03 | 0.01 ± 0.00 |
| 4 | α-pinene | 1029 | 1008–1039 | 2.26 ± 0.19 | 2.03 ± 0.16 | 0.08 ± 0.01 |
| 5 | camphene | 1075 | 1043–1086 | 0.40 ± 0.04 | 0.37 ± 0.05 | 0.19 ± 0.06 |
| 6 | β-pinene | 1111 | 1085–1138 | 0.27 ± 0.00 | 0.18 ± 0.01 | 0.10 ± 0.01 |
| 7 | sabinene | 1125 | 1098–1140 | 0.08 ± 0.01 | 0.08 ± 0.02 | 0.02 ± 0.00 |
| 8 | β-myrcene | 1167 | 1155–1169 | 2.37 ± 0.24 | 1.83 ± 0.22 | - |
| 9 | α-terpinene | 1183 | 1154–1195 | 2.17 ± 0.22 | 1.57 ± 0.06 | 0.39 ± 0.09 |
| 10 | D-limonene | 1203 | 1178–1219 | 0.20 ± 0.05 | 0.21 ± 0.04 | 0.36 ± 0.04 |
| 11 | 1.8-cineole | 1216 | 1186–1231 | - | 0.08 ± 0.01 | 1.82 ± 0.16 |
| 12 | β-ocimene | 1236 | 1211–1251 | 0.06 ± 0.00 | 0.07 ± 0.01 | - |
| 13 | γ-terpinene | 1253 | 1222–1266 | 19.73 ± 1.88 | 12.78 ± 1.28 | 0.37 ± 0.10 |
| 14 | p-cymene | 1276 | 1246–1291 | 4.29 ± 0.30 | 9.98 ± 0.60 | 0.88 ± 0.10 |
| 15 | terpinolene | 1285 | 1261–1300 | 0.09 ± 0.01 | 0.09 ± 0.01 | 0.22 ± 0.02 |
| 16 | hexanol | 1349 | 1344–1360 | - | 0.12 ± 0.01 | - |
| 17 | octan-3-ol | 1392 | 1372–1408 | - | 0.14 ± 0.03 | - |
| 18 | α-thujone | 1418 | 1385–1441 | - | - | 2.55 ± 0.33 |
| 19 | β-thujone | 1437 | 1400–1452 | - | - | 90.60 ± 5.03 |
| 20 | 1-octen-3-ol | 1446 | 1411–1465 | 0.51 ± 0.06 | - | - |
| 21 | menthone | 1459 | 1450–1475 | 0.29 ± 0.03 | - | 0.03 ± 0.00 |
| 22 | α-copaene | 1494 | 1462–1522 | - | 0.49 ± 0.12 | - |
| 23 | camphor | 1509 | 1481–1537 | 0.22 ± 0.03 | 0.14 ± 0.02 | 0.13 ± 0.01 |
| 24 | β-cubebene | 1536 | 1518–1560 | - | 1.10 ± 0.12 | - |
| 25 | linalool | 1542 | 1507–1564 | 0.25 ± 0.03 | 1.39 ± 0.12 | 0.06 ± 0.01 |
| 26 | bornyl acetate | 1577 | 1549–1597 | 1.20 ± 0.18 | 1.06 ± 0.17 | - |
| 27 | β-caryophyllene | 1593 | 1570–1685 | 0.40 ± 0.14 | 0.64 ± 0.09 | 0.33 ± 0.03 |
| 28 | terpinen-4-ol | 1597 | 1564–1630 | 0.84 ± 0.08 | 0.67 ± 0.06 | - |
| 29 | pulegone | 1623 | 1626–1663 | - | - | 0.26 ± 0.04 |
| 30 | menthol | 1631 | 1599–1651 | 0.21 ± 0.01 | - | 0.15 ± 0.02 |
| 31 | β-terpineol | 1636 | 1616–1644 | 0.88 ± 0.06 | ||
| 32 | borneol | 1687 | 1653–1728 | - | 0.29 ± 0.03 | - |
| 33 | carvone | 1711 | 1699–1751 | 0.62 ± 0.09 | - | - |
| 34 | germacrene D | 1716 | 1676–1726 | 0.44 ± 0.07 | 0.12 ± 0.02 | - |
| 35 | caryophyllene oxide | 1976 | 1936–2023 | 0.17± 0.02 | - | - |
| 36 | (−)-spathulenol | 2124 | 2074–2150 | 0.18 ± 0.01 | - | - |
| 37 | thymol | 2164 | 2100–2205 | 1.19 ± 0.12 | 59.34 ± 5.23 | 0.21 ± 0.03 |
| 38 | α-bisabolol | 2197 | 2178–2234 | 0.21 ± 0.02 | - | - |
| 39 | carvacrol | 2211 | 2140–2246 | 59.41 ± 5.13 | 2.41 ± 0.36 | 0.15 ± 0.02 |
| 40 | β-eudesmol | 2235 | 2196–2272 | - | - | 0.11 ± 0.01 |
| Total identified | 98.98 | 97.68 | 99.35 | |||
| Monoterpene hydrocarbons | 32.13 | 29.52 | 2.62 | |||
| Oxygenated monoterpenes | 4.51 | 3.63 | 95.60 | |||
| Phenolic monoterpenes | 60.60 | 61.75 | 0.36 | |||
| Sesquiterpene hydrocarbons | 0.84 | 2.35 | 0.33 | |||
| Oxygenated sesquiterpenes | 0.39 | 0.17 | 0.11 | |||
| Other compounds | 0.51 | 0.26 | 0.33 | |||
| ** Experimental Group | Average Number of Local Viral Lesions | SD | Inhibition of Local Lesions (%) | |
|---|---|---|---|---|
| CMV-S21 + Thyme 6000 ppm | 30.4 | a * | 18.60 | 79.5 |
| CMV-S21 + Thyme 5000 ppm | 42.0 | b | 15.72 | 71.6 |
| CMV-S21 + Thyme 4000 ppm | 56.0 | c | 21.66 | 62.2 |
| CMV-S21 + Thyme 3000 ppm | 73.0 | d | 18.00 | 50.7 |
| CMV-S21 + Thyme 2000 ppm | 90.1 | e | 27.05 | 39.1 |
| CMV-S21 + Thyme 1000 ppm | 112.6 | f | 16.61 | 23.9 |
| CMV-S21 + Greek oregano 6000 ppm | 121.0 | fg | 11.94 | 18.2 |
| CMV-S21 + Greek oregano 5000 ppm | 123.6 | fgh | 10.84 | 16.5 |
| CMV-S21 + Greek oregano 4000 ppm | 125.6 | fghi | 12.01 | 15.1 |
| CMV-S21 + Costmary 6000 ppm | 127.6 | fghi | 10.99 | 13.8 |
| CMV-S21 + Greek oregano 3000 ppm | 128.9 | fghij | 11.00 | 12.9 |
| CMV-S21 + Thyme 500 ppm | 130.5 | ghij | 28.47 | 11.8 |
| CMV-S21 + Costmary 5000 ppm | 130.9 | ghij | 11.71 | 11.5 |
| CMV-S21 + Costmary 4000 ppm | 132.7 | ghij | 12.08 | 10.3 |
| CMV-S21 + Greek oregano 2000 ppm | 132.8 | ghij | 13.80 | 10.3 |
| CMV-S21 + Costmary 3000 ppm | 135.7 | ghij | 13.61 | 8.3 |
| CMV-S21 + Greek oregano 1000 ppm | 137.0 | ghij | 15.17 | 7.4 |
| CMV-S21 + Costmary 2000 ppm | 138.9 | ghij | 15.70 | 6.2 |
| CMV-S21 + Greek oregano 500 ppm | 141.1 | hij | 16.56 | 4.7 |
| CMV-S21 + Costmary 1000 ppm | 142.9 | hij | 26.69 | 3.4 |
| CMV-S21 + Costmary 500 ppm | 144.2 | ij | 24.78 | 2.6 |
| *** Control group 3 | 148.0 | j | 32.78 | |
| Essential Oil | Concentration (ppm) | ||||||
|---|---|---|---|---|---|---|---|
| 500 | 1000 | 2000 | 3000 | 4000 | 5000 | 6000 | |
| Absorbance at λ = 405 nm | |||||||
| Greek oregano | 1.000 | 0.954 | 0.919 | 0.893 | 0.823 | 0.765 | 0.715 |
| Thyme | 0.989 | 0.802 | 0.765 | 0.690 | 0.591 | 0.515 | 0.491 |
| Costmary | 1.012 | 0.989 | 0.976 | 0.912 | 0.892 | 0.812 | 0.801 |
| Plant Inoculation/Thyme Oil Application (6000 ppm) | Time of Application (h) | ||
|---|---|---|---|
| 24 | 48 | 72 | |
| Absorbance at λ = 405 nm | |||
| * Test A CMV-S21 isolate + T6000 | 0.452 | 0.431 | 0.390 |
| ** Test B T6000 + CMV-S21 isolate | 0.341 | 0.322 | 0.301 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paduch-Cichal, E.; Mirzwa-Mróz, E.; Wojciechowska, P.; Bączek, K.; Kosakowska, O.; Węglarz, Z.; Szyndel, M.S. Antiviral Activity of Selected Essential Oils against Cucumber Mosaic Virus. Plants 2023, 12, 18. https://doi.org/10.3390/plants12010018
Paduch-Cichal E, Mirzwa-Mróz E, Wojciechowska P, Bączek K, Kosakowska O, Węglarz Z, Szyndel MS. Antiviral Activity of Selected Essential Oils against Cucumber Mosaic Virus. Plants. 2023; 12(1):18. https://doi.org/10.3390/plants12010018
Chicago/Turabian StylePaduch-Cichal, Elżbieta, Ewa Mirzwa-Mróz, Patrycja Wojciechowska, Katarzyna Bączek, Olga Kosakowska, Zenon Węglarz, and Marek Stefan Szyndel. 2023. "Antiviral Activity of Selected Essential Oils against Cucumber Mosaic Virus" Plants 12, no. 1: 18. https://doi.org/10.3390/plants12010018
APA StylePaduch-Cichal, E., Mirzwa-Mróz, E., Wojciechowska, P., Bączek, K., Kosakowska, O., Węglarz, Z., & Szyndel, M. S. (2023). Antiviral Activity of Selected Essential Oils against Cucumber Mosaic Virus. Plants, 12(1), 18. https://doi.org/10.3390/plants12010018