
Morphological and Genetic Diversity of Cucumber (Cucumis sativus L.) Fruit Development


{kind=link}
{kind=link}
{kind=link}
Abstract
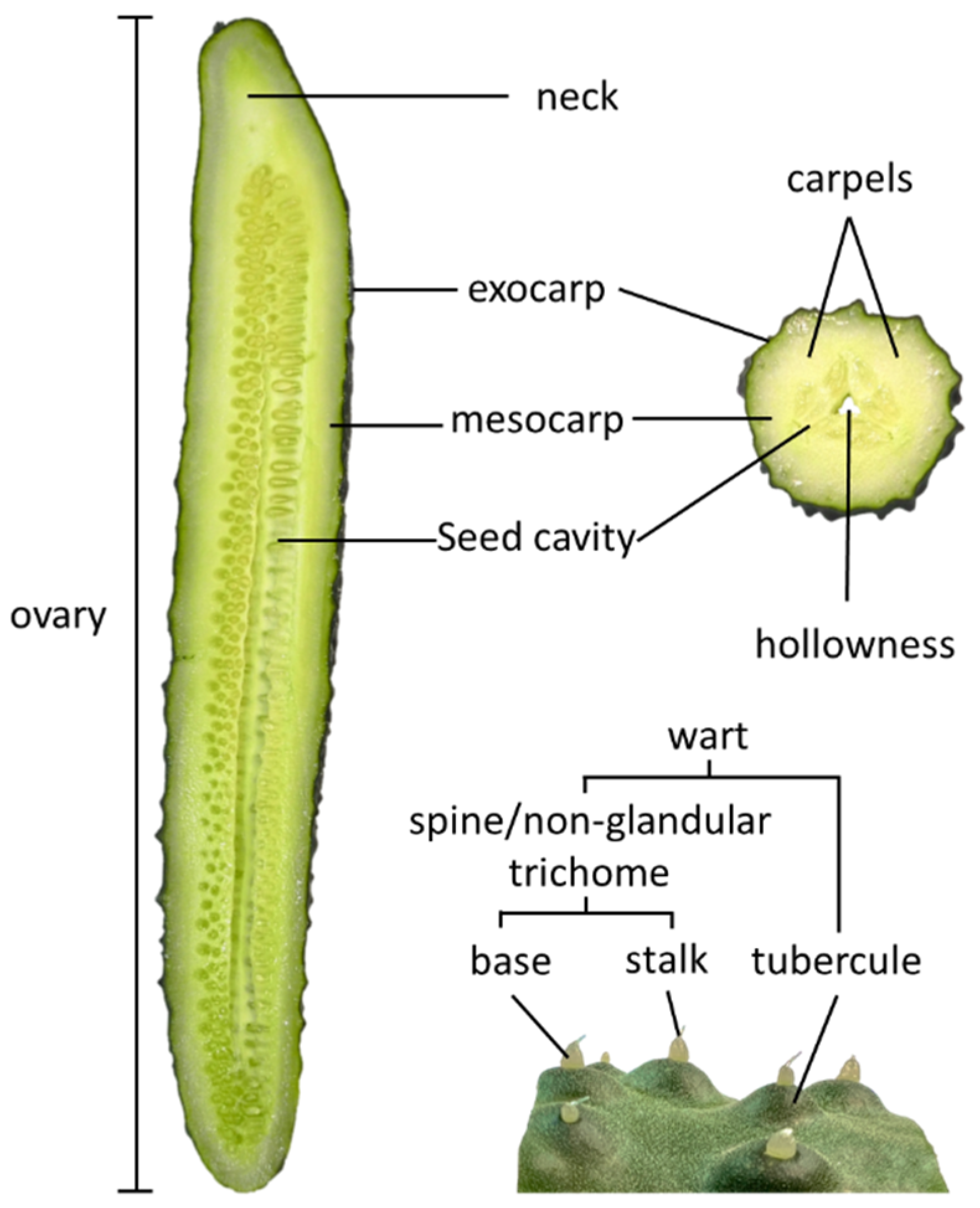
:1. Introduction
2. Fruit Development, Size, and Shape
2.1. Fruit Size
2.2. Parthenocarpy
2.3. Fruit Shape
3. Fruit Surface Features
3.1. Trichomes
3.2. Tubercules
3.3. Cuticle and Wax
3.4. Netting
4. Fruit Color
4.1. Skin Color
4.2. Flesh Color
5. Internal Features
5.1. Flesh Thickness
5.2. Seed Cavity and Hollowness
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, X.; Bao, K.; Reddy, U.K.; Bai, Y.; Hammar, S.A.; Jio, C.; Wehner, T.C.; Madera, A.R.; Weng, X.; Grumet, R.; et al. The USDA cucumber (Cucumis sativus L.) collection: Genetic diversity, population structure, genome-wide association studies and core collection development. Hortic. Res. 2018, 5, 64. [Google Scholar] [CrossRef] [PubMed]
- Robinson, R.W.; Decker-Walters, D.S. Cucurbits; CAB International: Wallingford, UK, 1997. [Google Scholar]
- Jiang, L.; Yan, S.; Yang, W.; Li, Y.; Xiz, Y.; Xia, M.; Cheng, Z.; Wang, Q.; Yan, L.; Song, X.; et al. Transcriptomic analysis reveals the roles of microtubule-related genes and transcription factors in fruit length regulation in cucumber (Cucumis sativus L.). Sci. Rep. 2015, 5, 8031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colle, M.; Weng, Y.; Kang, Y.; Ophir, R.; Sherman, A.; Grumet, R. Variation in cucumber (Cucumis sativus L.) fruit size and shape results from multiple components acting pre-anthesis and post-pollination. Planta 2017, 246, 641–658. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.X.; Pan, Y.P.; Liu, C.; Ding, Y.Y.; Wang, X.; Cheng, Z.H.; Meng, H.W. Cucumber fruit size and shape variations explored from the aspects of morphology, histology, and endogenous hormones. Plants 2020, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.; Colle, M.; Wang, Y.; Yang, L.; Rubenstein, M.; Sherman, A.; Ophir, R.; Grumet, R. QTL mapping in multiple populations and development stages reveals dynamic quantitative trait loci for fruit size in cucumbers of different market classes. Theor. Appl. Genet. 2015, 128, 1747–1763. [Google Scholar] [CrossRef]
- Zhang, T.T.; Li, X.Z.; Yang, Y.T.; Guo, X.; Feng, Q.; Dong, X.Y.; Chen, S.X. Genetic analysis and QTL mapping of fruit length and diameter in a cucumber (Cucumber sativus L.) recombinant inbred line (RIL) population. Sci. Hortic. 2019, 250, 214–222. [Google Scholar] [CrossRef]
- Gao, Z.H.; Zhang, H.Y.; Cao, C.X.; Han, J.; Li, H.; Ren, Z.H. QTL Mapping for cucumber fruit size and shape with populations from long and round fruited inbred lines. Hortic. Plant J. 2020, 6, 132–144. [Google Scholar] [CrossRef]
- Cui, L.; Li, J.; Zhang, T.; Guo, Q.; Xu, J.; Lou, Q.; Chen, J. Identification and expression analysis of D-type cycling genes in early developing fruit of cucumber (Cucumis sativus L.). Plant Mol. Biol. Rep. 2014, 32, 209–218. [Google Scholar] [CrossRef]
- Fu, F.Q.; Mao, W.H.; Shi, K.; Zhou, Y.H.; Asami, T.; Yu, J.Q. Spatio-temporal changes in cell division, endoreduplication and expression of cell cycle-related genes in pollinated and plant growth substances-treated ovaries of cucumber. Plant Biol. 2010, 12, 98–107. [Google Scholar] [CrossRef]
- Ando, K.; Carr, K.M.; Grumet, R. Transcriptome analyses of early cucumber fruit growth identifies distinct gene modules associated with phases of development. BMC Genom. 2012, 13, 518–534. [Google Scholar] [CrossRef]
- Yang, X.Y.; Wang, Y.; Jiang, W.; Liu, X.L.; Zhang, X.M.; Yu, J.H.; Huang, S.W.; Liu, G.Q. Characterization and expression profiling of cucumber kinesin genes during early fruit development: Revealing the roles of kinesins in exponential cell production and enlargement in cucumber fruit. J. Exp. Bot. 2013, 64, 4541–4557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Sun, J.; Yang, F.; Weng, Y.; Chen, P.; Du, S.; Wei, A.; Li, Y. CsKTN1 for a katanin p60 subunit is associated with the regulation of fruit elongation in cucumber (Cucumis sativus L.). Theor Appl Genet. 2021, 134, 2429–2441. [Google Scholar] [CrossRef]
- Luptovčiak, I.; Komis, G.; Takáč, T.; Ovečka, M.; Šamaj, J. Katanin: A sword cutting microtubules for cellular, developmental, and physiological purposes. Front Plant Sci. 2017, 8, 1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Wang, B.W.; Wang, S.H.; Lin, T.; Yang, L.; Zhao, Z.L.; Zhang, Z.H.; Huang, S.W.; Yang, X.Y. Genome-wide target mapping shows histone deacetylase complex1 regulates cell proliferation in cucumber fruit. Plant Physiol. 2020, 182, 167–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Jiang, L.; Che, G.; Pan, Y.; Li, Y.; Hou, Y.; Zhao, W.; Zhong, Y.; Ding, L.; Yan, S.; et al. A functional allele of CsFUL1 regulates fruit length through repressing CsSUP and inhibiting auxin transport in cucumber. Plant Cell 2019, 31, 1289–1307. [Google Scholar] [CrossRef] [PubMed]
- Boonkorkaew, P.; Hikosaka, S.; Sugiyama, N. Effect of pollination on cell division, cell enlargement, and endogenous hormones in fruit development in a gynoecious cucumber. Sci. Hortic. 2008, 116, 1–7. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.; Yao, X.; Wang, J.; Feng, S.; Sun, L.; Ma, S.; Xu, K.; Chen, L.Q.; Sui, X. Hexose transporter CsSWEET7a in cucumber mediates phloem unloading in companion cells for fruit development. Plant Physiol. 2021, 186, 640–654. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, X.; Zhao, Y.L.; Nie, J.; Yao, X.H.; Lv, L.J.; Yang, J.W.; Ma, N.; Guo, Y.C.; Li, Y.X.; et al. Alkaline alpha-galactosidase 2 (CsAGA2) plays a pivotal role in mediating source-sink communication in cucumber. Plant Physiol. 2022, 189, 1501–1518. [Google Scholar] [CrossRef]
- Zhang, Z.; Deng, Y.; Song, X.; Miao, M. Trehalose-6-phosphate and SNF1-related protein kinase 1 are involved in the first-fruit inhibition of cucumber. J. Plant Physiol. 2015, 177, 110–120. [Google Scholar] [CrossRef]
- Shnaider, Y.; Mitra, D.; Miller, G.; Baniel, A.; Doniger, T.; Kuhalskaya, A.; Scossa, F.; Fernie, A.R.; Brotman, Y.; Perl-Treves, R. Cucumber ovaries inhibited by dominant fruit express a dynamic developmental program, distinct from either senescence-determined or fruit-setting ovaries. Plant J. 2018, 96, 651–669. [Google Scholar] [CrossRef]
- Fu, F.Q.; Mao, W.H.; Shi, K.; Zhou, Y.H.; Asami, T.; Yu, J.Q. A role of brassinosteroids in early fruit development in cucumber. J. Exp. Bot. 2008, 59, 2299–2308. [Google Scholar] [CrossRef] [PubMed]
- Xin, T.X.; Zhang, Z.; Li, S.; Zhang, S.; Li, Q.; Zhang, Z.H.; Huang, S.W.; Yang, X.Y. Genetic regulation of ethylene dosage for cucumber fruit elongation. Plant Cell 2019, 31, 1063–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Li, J.; Cui, L.; Zhang, T.; Wu, Z.; Zhu, P.Y.; Meng, Y.J.; Zhang, K.J.; Yu, X.Q.; Lou, Q.F.; et al. New insights into the roles of cucumber TIR1 homologs and miR393 in regulating fruit/seed set development and leaf morphogenesis. BMC Plant Biol. 2017, 17, 130–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.D.; Luo, W.R.; Chang, H.C.; Li, Z.X.; Zhou, J.G.; Li, X.Z.; Zheng, J.L.; Hao, M.X. Identification of miRNAs and their target genes involved in cucumber fruit expansion using small RNA and degradome sequencing. Biomolecules 2019, 9, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Q.Z.; Fu, W.Y.; Wang, Y.Z.; Qin, X.D.; Wang, J.; Li, J.; Lou, Q.F.; Chen, J.F. Rapid identification of fruit length loci in cucumber (Cucumis sativus L.) using next-generation sequencing (NGS)-based QTL analysis. Sci Rep. 2016, 6, 27496. [Google Scholar] [CrossRef]
- Zhu, W.Y.; Huang, L..; Chen, L.; Yang, J.T.; Wu, J.N.; Qu, M.L.; Yao, D.Q.; Guo, C.L.; Lian, H.L.; He, H.L.; et al. A high-density genetic linkage map for cucumber (Cucumis sativus L.): Based on specific length amplified fragment (SLAF) sequencing and QTL analysis of fruit traits in cucumber. Front Plant Sci. 2016, 7, 437. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.P.; Wang, Y.H.; McGregor, C.; Liu, S.; Luan, F.S.; Gao, M.L.; Weng, Y. Genetic architecture of fruit size and shape variation in cucurbits: A comparative perspective. Theor. Appl. Genet. 2020, 133, 1–21. [Google Scholar] [CrossRef]
- Sheng, Y.Y.; Pan, Y.P.; Li, Y.H.; Yang, L.M.; Weng, Y.Q. Quantitative trait loci for fruit size and flowering time-related traits under domestication and diversifying selection in cucumber (Cucumis sativus). Plant Breed. 2020, 139, 176–191. [Google Scholar] [CrossRef]
- Wang, Y.; Bo, K.; Gu, X.; Pan, J.; Li, Y.; Chen, J.; Wen, C.; Ren, Z.; Ren, H.; Chen, X.; et al. Molecularly tagged genes and quantitative trait loci in cucumber with recommendations for QTL Nomenclature. Hortic. Res. 2020, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.J.; Van Houten, J.; Gonzalez, G.; Xiao, H.; van der Knapp, E. Genome-wide identification, phylogeny and expression analysis of SUN, OFP and YABBY gene family in tomato. Mol Gen Genom. 2013, 288, 111–129. [Google Scholar] [CrossRef]
- Monforte, A.J.; Diaz, A.; Cano-Delgado, A.; van der Knapp, E. The genetic basis of fruit morphology in horticultural crops: Lessons from tomato and melon. J. Exp. Bot. 2014, 65, 4625–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, S.E.; Running, M.P.; Meyerowitz, E.M. CLAVATA1, a regulator of meristem and flower development in Arabidopsis. Development 1993, 119, 397–418. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Song, M.; Zhang, M.; Cheng, C.; Chen, J.; Lou, Q. A SNP mutation in the CsCLAVATA1 leads to pleiotropic variation in plant architecture and fruit morphogenesis in cucumber (Cucumis sativus L.). Plant Sci. 2022, 323, 111397. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, H.; Zhao, J.; Pan, Y.; Cheng, S.; Lietzow, C.D.; Wen, C.; Zhang, X.; Weng, Y. LITTLELEAF (LL) encodes a WD40 repeat domain-containing protein associated with organ size variation in cucumber. Plant J. 2018, 95, 834–847. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Ning, K.; Wang, Z.; Liu, X.; Zhong, Y.; Ding, H.; Zi, H.; Cheng, Z.; Li, X.; Shan, H.; et al. CsIVP functions in vasculature development and downy mildew resistance in cucumber. PLoS Biol. 2020, 18, e3000671. [Google Scholar] [CrossRef] [Green Version]
- Lietzow, C.D.; Zhu, H.; Pandey, S.; Havey, M.J.; Weng, Y. QTL mapping of parthenocarpic fruit set in North American processing cucumber. Theor. Appl. Genet. 2016, 129, 2387–2401. [Google Scholar] [CrossRef]
- Dhatt, A.S.; Kaur, G. Parthenocarpy: A potential trait to exploit in vegetable crops: A review. Agric. Rev. 2016, 37, 300–308. [Google Scholar] [CrossRef] [Green Version]
- Su, L.; Rahat, S.; Ren, N.; Kojima, M.; Takebayashi, Y.; Sakakibara, H.; Wang, M.; Chen, X.; Qi, X. Cytokinin and auxin modulate cucumber parthenocarpy fruit development. Sci. Hortic. 2021, 282, 110026. [Google Scholar] [CrossRef]
- Mandal, N.K.; Kumari, K.; Kundu, A.; Arora, A.; Bhowmick, P.K.; Iquebal, M.A.; Jaiswal, S.; Behera, T.K.; Munshi, A.D.; Dey, S.S. Cross-talk between the cytokinin, auxin, and gibberellin regulatory networks in determining parthenocarpy in cucumber. Front Genet. 2022, 13, 957360. [Google Scholar] [CrossRef]
- Beyer, E.M.; Quebedeaux, B. Parthenocarpy in cucumber: Mechanism of action of auxin transport inhibitors. J. Am. Soc. Hortic. Sci. 1974, 99, 385–390. [Google Scholar] [CrossRef]
- Cui, L.; Zhang, T.; Li, J.; Lou, Q.; Chen, J. Cloning and expression analysis of Cs-TIR1/AFB2: The fruit development-related genes of cucumber (Cucumis sativus L.). Acta Physiol. Plant. 2014, 36, 139–149. [Google Scholar] [CrossRef]
- Li, J.; Xu, J.; Guo, Q.W.; Wu, Z.; Zhang, T.; Zhang, K.J.; Chen, C.Y.; Zhu, P.Y.; Lou, Q.F.; Chen, J.F. Proteomic insight into fruit set of cucumber (Cucumis sativus L.) suggests the cues of hormone-independent parthenocarpy. BMC Genom. 2017, 18, 896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Su, L.; Cong, Y.; Chen, J.; Geng, Y.; Qian, Z.; Xu, Q. Sugars enhance parthenocarpic fruit formation in cucumber by promoting auxin and cytokinin signaling. Sci. Hortic. 2021, 283, 110061. [Google Scholar] [CrossRef]
- Gou, C.; Zhu, P.; Meng, Y.; Yang, F.; Xu, Y.; Xia, P.; Chen, J.; Li, J. Evaluation and genetic analysis of parthenocarpic germplasms in cucumber. Genes 2022, 13, 225. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.Y.; Lou, L.N.; Li, X.L.; Feng, Z.H.; Lou, Q.F.; Chen, J.F. Evaluation of parthenocarpy in cucumber germplasm. Acta Hortic. Sin. 2009, 36, 975–982. [Google Scholar] [CrossRef]
- Sun, Z.; Staub, J.; Chung, S.; Lower, R. Identification and comparative analysis of quantitative trait loci associated with parthenocarpy in processing cucumber. Plant Breed. 2006, 125, 281–287. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, T.; Li, L.; Xu, J.; Qin, X.; Zhang, T.; Cui, L.; Lou, Q.; Li, J.; Chen, J. Identification of a stable major-effect QTL (Parth 2.1) controlling parthenocarpy in cucumber and associated candidate gene analysis via whole genome re-sequencing. BMC Plant Biol. 2016, 16, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, Z.H.; Song, X.F.; Li, X.L.; Guo, X.Y.; He, S.Q.; He, L.J.Z.; Feng, Z.H.; Sun, C.Z.; Yan, L.Y. Inheritance and QTL mapping for parthenocarpy in cucumber. Sci. Agric. Sin. 2020, 53, 160–171. [Google Scholar] [CrossRef] [Green Version]
- Grumet, R.; Colle, M. Genomic analysis of cucurbit fruit growth. In Genetics and Genomics of the Cucurbitaceae; Grumet, R., Katzir, N., Garcia-Mas, J., Eds.; Springer Publishing: Chan, Switzerland, 2017; pp. 307–320. [Google Scholar] [CrossRef]
- Saito, S.; Fujii, N.; Miyazawa, Y.; Yamasaki, S.; Matsuura, S.; Mizusawa, H.; Fujita, Y.; Takashashi, H. Correlation between development of female flowers and expression of the CsACS2 gene in cucumber plants. J. Exp. Bot. 2007, 58, 2897–2907. [Google Scholar] [CrossRef]
- Li, Z.; Pan, J.S.; Guan, Y.; Tao, Q.Y.; He, H.L.; Si, L.T.; Cair, R. Development and fine mapping of three co-dominant SCAR markers linked ot the M/m gene in the cucumber plant (Cucumis sativus L). Theor. Appl. Genet. 2008, 117, 1253–1260. [Google Scholar] [CrossRef]
- Boualem, A.; Troadec, C.; Kovalski, I.; Sari, M.; Perl-Treves, R.; Bendahmane, A. A conserved ethylene biosynthesis enzyme leads to andromonoecy. in two Cucumis species. PLoS ONE 2008, 4, e6144. [Google Scholar] [CrossRef]
- Manzano, S.; Martinez, C.; Megias, Z.; Gomez, P.; Garrido, D.; Jamilena, M. The role of ethylene and brassinosteroids in the control of sex expression and flower development in Cucurbita pepo. Plant Growth Reg. 2011, 65, 213–221. [Google Scholar] [CrossRef]
- Pitrat, M. Phenotypic diversity in wild and cultivated melons (Cucumis melo). Plant Biotechnol. 2013, 30, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Switzenberg, J.A.; Beaudry, R.M.; Grumet, R. Effect of CRC::etr1-1 transgene expression on ethylene production, sex expression, fruit set and fruit ripening in transgenic melon (Cucumis melo L.). Transgenic. Res. 2015, 24, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Tao, Q.; Niu, H.; Zhang, Z.; Li, D.; Gong, Z.; Weng, Y.; Li, Z. A novel allele of monecious (m) locus is responsible for elongated fruit shape and perfect flowers in cucumber (Cucumis sativus L.). Theor. Appl. Genet. 2015, 128, 2483–2493. [Google Scholar] [CrossRef]
- Li, S.; Pan, Y.; Wen, C.; Li, Y.; Liu, X.; Zhang, X.; Behera, T.K.; Xing, G.; Weng, Y. Integrated analysis in bi-parental and natural populations reveals CsCLAVATA3 (CsCLV3) underlying carpel number variations in cucumber. Theor. Appl. Genet. 2016, 129, 1007–1022. [Google Scholar] [CrossRef]
- Yadav, R.K.; Perales, M.; Gruel, J.; Girke, T.; Jönsson, H.; Reddy, G.V. WUSCHEL protein movement mediates stem cell homeostasis in the Arabidopsis shoot apex. Genes Dev. 2011, 25, 2025–2030. [Google Scholar] [CrossRef] [Green Version]
- Che, G.; Gu, R.; Zhao, J.; Liu, X.; Song, X.; Zi, H.; Cheng, Z.; Shen, J.; Wang, Z.; Liu, R.; et al. Gene regulatory network controlling carpel number variation in cucumber. Development 2020, 147, dev184788. [Google Scholar] [CrossRef]
- Fernandez-Silva, I.; Moreno, E.; Essafi, A.; Fergany, M.; Garcia-Mas, J.; Martín-Hernandez, A.M.; Alvarez, J.M.; Monforte, A.J. Shaping melons: Agronomi.c and genetic characterization of QTLs that modify melon fruit morphology. Theor. Appl. Genet. 2010, 121, 931–940. [Google Scholar] [CrossRef]
- Liu, X.; Hao, N.; Li, H.; Ge, D.; Du, Y.; Liu, R.; Wen, C.; Li, Y.; Zhang, X.; Wu, T. PINOID is required for lateral organ morphogenesis and ovule development in cucumber. J. Exp. Bot. 2019, 70, 5715–5730. [Google Scholar] [CrossRef]
- Liu, X.F.; Ning, K.; Che, G.; Yan, S.S.; Han, L.J.; Gu, R.; Li, Z.; Weng, Y.Q.; Zhang, X.L. CsSPL functions as an adaptor between HD-ZIPIII and CsWUS transcription factors regulating anther and ovule development in Cucumis sativus (cucumber). Plant J. 2018, 94, 535–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, H.; Liu, X.; Tong, C.; Wang, H.; Li, S.; Lu, L.; Pan, Y.; Zhang, X.; Weng, Y.; Li, Z. The WUSCHEL-related homeobox1 gene of cucumber regulates reproductive organ development. J. Exp. Bot. 2018, 69, 5373–5387. [Google Scholar] [CrossRef] [Green Version]
- Owens, K.W.; Bliss, F.A.; Peterson, C.E. Genetic analysis of fruit length and weight in two cucumber populations using the inbred backcross lines. J. Am. Soc. Hortic. Sci. 1985, 110, 431–436. [Google Scholar] [CrossRef]
- Strefeler, M.S.; Wehner, T.C. Estimates of heritabilities and genetic variances of three yield and five quality traits in three fresh market cucumber populations. J. Am. Soc. Hortic. Sci. 1986, 111, 599–605. [Google Scholar] [CrossRef]
- Shimomura, K.; Fukino, N.; Sugiyama, M.; Kawazu, Y.; Sakata, Y.; Yoshioka, Y. Quantitative trait locus analysis of cucumber fruit morphological traits based on image analysis. Euphytica 2017, 213, 138. [Google Scholar] [CrossRef]
- Yuan, X.J.; Pan, J.S.; Cai, R.; Guan, Y.; Liu, L.Z.; Zhang, W.W.; Li, Z.; He, H.L.; Zhang, C.; Si, L.T. Genetic mapping and QTL analysis of fruit and flower related traits in cucumber (Cucumis sativus L.) using recombinant inbred lines. Euphytica 2008, 164, 473–491. [Google Scholar] [CrossRef]
- Miao, H.; Gu, X.F.; Zhang, S.P. Mapping QTLs for fruit-associated traits in Cucumis sativus. L. Sci. Agric. Sin. 2011, 44, 531–540. [Google Scholar] [CrossRef]
- Pan, Y.; Liang, X.; Gao, M.; Liu, H.; Meng, H.; Cheng, Z. Round fruit shape in WI7239 cucumber is controlled by two interacting quantitative trait loci with one putatively encoding a tomato SUN homolog. Theor. Appl. Genet. 2017, 130, 573–586. [Google Scholar] [CrossRef]
- Wang, X.; Li, H.; Gao, Z.; Wang, L.; Ren, Z. Localization of quantitative trait loci for cucumber fruit shape by a population of chromosome segment substitution lines. Sci. Rep. 2020, 10, 11030. [Google Scholar] [CrossRef]
- Wu, S.; Clevenger, J.P.; Sun, L.; Visa, S.; Kamiya, Y.; Jikumaru, Y.; Blakeslee, J.; van der Knapp, E. The control of tomato fruit elongation orchestrated by sun, ovate, and fs8.1 in a wild relative of tomato. Plant Sci. 2015, 238, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Zhang, B.Y.; Keyhaninejad, N.; Rodríguez, G.R.; Kim, H.J.; Chakrabarti, M.; Illa-Berenguer, E.; Taitano, N.K.; Gonzalo, M.J.; Díaz, A.; et al. Tomato reveals a common mechanism underlying morphological diversification in domesticated plants. Nat. Commun. 2018, 9, 4734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, M.; Yang, Y.; Cheng, Z.; Meng, H. Dynamic changes of fruit shape traits and its correlation to the morphology of other organs in cucumber. Agronomy 2020, 10, 1111. [Google Scholar] [CrossRef]
- Shimomura, K.; Horiea, H.; Sugiyama, M.; Kawazua, Y.; Yoshioka, Y. Quantitative evaluation of cucumber fruit texture and shape traits reveals extensive diversity and differentiation. Sci. Hortic. 2016, 199, 133–141. [Google Scholar] [CrossRef]
- Zhao, J.; Li, Y.; Ding, L.; Yan, S.; Liu, M.; Jiang, L.; Zhao, W.; Wang, Q.; Yan, L.; Liu, R.; et al. Phloem transcriptome signatures underpin the physiological differentiation of the pedicel, stalk and fruit of cucumber (Cucumis sativus L.). Plant Cell Physiol. 2016, 57, 19–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.W.; Wei, C.X.; Liu, Q.Y.; Qu, W.Q.; Qi, X.H.; Xu, Q.; Chen, X.H. The major-effect quantitative trait locus Fnl7.1 encodes a late embryogenesis abundant protein associated with fruit neck length in cucumber. Plant Biotechnol. J. 2020, 18, 1598–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Zhou, Z.; Wang, L.; Yan, S.; Cheng, Z.; Liu, X.; Han, L.; Chen, G.; Wang, S.; Song, W.; et al. The CsHEC1-CsOVATE module contributes to fruit neck length variation via modulating auxin biosynthesis in cucumber. Proc. Natl. Acad. Sci. USA 2022, 119, e2209717119. [Google Scholar] [CrossRef]
- Li, S.; Wang, C.; Zhou, X.; Liu, D.; Liu, C.; Luan, J.; Qin, Z.; Xin, M. The curvature of cucumber fruits is associated with spatial variation in auxin accumulation and expression of a YUCCA biosynthesis gene. Hortic. Res. 2020, 7, 135. [Google Scholar] [CrossRef]
- Zhu, P.Y.; Meng, Y.J.; Zhang, K.J.; Wang, X.Y.; Liang, K.; Wang, T.T.; Xu, J.; Qin, X.D.; Wu, Z.; Cheng, C.Y.; et al. Mapping of fruit apex shape related QTL across multi-genetic backgrounds in cucumber (Cucumis sativus L.). Hortic. Plant J. 2022, 8, 328–340. [Google Scholar] [CrossRef]
- Klosinska, U.; Kozik, E.U.; Wehner, T.C. Inheritance of a new trait-twin fused fruit-in cucumber. HortScience 2006, 41, 313–314. [Google Scholar] [CrossRef]
- Liu, X.; Yang, X.; Xie, Q.; Miao, H.; Bo, K.; Dong, S.; Xin, T.; Gu, X.; Sun, J.; Zhang, Z. NS encodes an auxin transporter that regulates the ‘numerous spines’ trait in cucumber (Cucumis sativus) fruit. Plant J. 2022, 110, 325–336. [Google Scholar] [CrossRef]
- Chen, C.; Liu, M.; Jiang, L.; Liu, X.; Zhao, J.; Yan, S.; Yang, S.; Ren, H.; Liu, R.; Zhang, X. Transcriptome profiling reveals roles of meristem regulators and polarity genes during fruit trichome development in cucumber (Cucumis sativus L.). J. Exp. Bot. 2014, 65, 4943–4958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Yin, S.; Liu, X.; Liu, B.; Yang, S.; Xue, S.; Cai, Y.; Black, K.; Liu, H.; Dong, M.; et al. The WD-repeat protein CsTTG1 regulates fruit wart formation through interaction with the homeodomain-leucine zipper I protein Mict. Plant Physiol. 2016, 71, 1156–1168. [Google Scholar] [CrossRef] [Green Version]
- Xue, S.; Dong, M.; Liu, X.; Xu, S.; Pang, J.; Zhang, W.; Weng, Y.; Ren, H. Classification of fruit trichomes in cucumber and effects of plant hormones on type II fruit trichome development. Planta 2019, 249, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, S.; Miao, H.; Wang, M.; Liu, P.; Wehner, T.C.; Gu, X. Molecular mapping and candidate gene analysis for numerous spines on the fruit of cucumber. J. Hered. 2016, 107, 471–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Q.; Liu, P.; Shi, L.; Miao, H.; Bo, K.; Wang, Y.; Gu, X.; Zhang, S. Combined fine mapping, genetic diversity, and transcriptome profiling reveals that the auxin transporter gene ns plays an important role in cucumber fruit spine development. Theor. Appl. Genet. 2018, 131, 1239–1252. [Google Scholar] [CrossRef]
- Bo, K.; Miao, H.; Wang, M.; Xie, X.; Song, Z.; Xie, Q.; Shi, L.; Wang, W.; Wei, S.; Zhang, S.; et al. Novel loci fsd6.1 and Csgl3 regulate ultra-high fruit spine density in cucumber. Theor. Appl. Genet. 2019, 132, 27–40. [Google Scholar] [CrossRef]
- Yang, C.; Li, H.; Zhang, J.; Wang, T.; Ye, Z. Fine-mapping of the woolly gene controlling multicellular trichome formation and embryonic development in tomato. Theor. Appl. Genet. 2011, 123, 625–633. [Google Scholar] [CrossRef]
- Liu, X.; Bartholomew, E.; Cai, Y.; Ren, H. Trichome-related mutants provide a new perspective on multicellular trichome initiation and development in cucumber (Cucumis sativus L). Front Plant Sci. 2016, 10, 1187. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Bo, K.; Cheng, Z.; Weng, Y. The loss-of-function GLABROUS 3 mutation in cucumber is due to LTR-retrotransposon insertion in a class IV HD-ZIP transcription factor gene CsGL3 that is epistatic over CsGL1. BMC Plant Biol. 2015, 15, 302. [Google Scholar] [CrossRef]
- Cui, J.Y.; Miao, H.; Ding, L.H.; Wehner, T.C.; Liu, P.N.; Wang, Y.; Zhang, S.P.; Gu, X.F. A new glabrous gene (csgl3) identified in trichome development in cucumber (Cucumis sativus L.). PLoS ONE 2016, 11, e0148422. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.L.; Nie, J.T.; Chen, H.M.; Guo, C.L.; Pan, J.; He, H.L.; Pan, J.S.; Cai, R. Identification and mapping of Tril, a homeodomain-leucine zipper gene involved in multicellular trichome initiation in Cucumis sativus. Theor. Appl. Genet. 2016, 9, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, L.; Zheng, S.; Liu, Z.; Wu, X.; Gao, Z.; Cao, C.; Li, Q.; Ren, Z. A fragment substitution in the promoter of CsHDZIV11/CsGL3 is responsible for fruit spine density in cucumber (Cucumis sativus L.). Theor. Appl. Genet. 2016, 129, 1289–1301. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Wang, G.; Pan, J.; Chen, Y.; Xiao, T.; Zhang, L.; Zhang, K.; Wen, H.; Xiong, L.; Yu, Y.; et al. The HD-ZIP IV transcription factor Tril regulates fruit spine density through gene dosage effects in cucumber. J. Exp. Bot. 2020, 22, 6297–6310. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Cai, Y.; Liu, X.; Dong, M.; Zhang, Y.; Chen, S.; Zhang, W.; Li, Y.; Tang, M.; Zhang, X.; et al. A CsMYB6-CsTRY module regulates fruit trichome initiation in cucumber. J. Exp. Bot. 2018, 69, 1887–1902. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Cao, C.; Zhang, C.; Zhen, S.; Wang, Z.; Wang, L.; Ren, Z. The identification of Cucumis sativus Glabrous 1 (CsGL1) required for the formation of trichomes uncovers a novel function for the homeodomain-leucine zipper I gene. J. Exp. Bot. 2015, 66, 2515–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.L.; Pan, J.S.; Guan, Y.; Zhang, W.W.; Bie, B.B.; Wang, Y.L.; He, H.L.; Lian, H.L.; Cai, R. Micro-trichome as a class I homeodomain-leucine zipper gene regulates multicellular trichome development in Cucumis sativus. J. Integr. Plant Biol. 2015, 57, 925–935. [Google Scholar] [CrossRef]
- Zhang, Y.; Shen, J.; Bartholomew, E.S.; Dong, M.; Chen, S.; Yin, S.; Zhai, X.; Feng, Z.; Ren, H.; Liu, X. TINY BRANCHED HAIR functions in multicellular trichome development through an ethylene pathway in Cucumis sativus L. Plant J. 2021, 106, 753–765. [Google Scholar] [CrossRef]
- Elhiti, M.; Stasolla, C. Structure and function of homodomain-leucine zipper (HD-Zip) proteins. Plant Signal Behav. 2009, 4, 86–88. [Google Scholar] [CrossRef] [Green Version]
- Sessa, G.; Carabelli, M.; Possenti, M.; Morelli, G.; Ruberti, I. Multiple Links between HD-Zip Proteins and Hormone Networks. Int. J. Mol. Sci. 2018, 19, 047. [Google Scholar] [CrossRef]
- Liu, X.W.; Wang, T.; Bartholomew, E.; Black, K.; Dong, M.; Zhang, Y.; Yang, S.; Cai, Y.; Xue, S.; Weng, Y.; et al. Comprehensive analysis of NAC transcription factors and their expression during fruit spine development in cucumber (Cucumis sativus L.). Hortic. Res. 2018, 1, 31. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, Y.; Zhou, P.; Bao, W.; Yang, Y.; Xu, T.; She, W.; Xu, L.; Yu, P.; Pan, J. Identification and fine mapping of molecular markers closely linked to fruit spines size ss gene in cucumber (Cucumis sativus L.). Euphytica 2018, 214, 213. [Google Scholar] [CrossRef]
- Yang, S.; Wang, Y.; Zhu, H.; Zhang, M.; Wang, D.; Xie, K.; Fan, P.; Dou, J.; Liu, D.; Liu, B.; et al. A novel HD-Zip I/C2H2-ZFP/WD-repeat complex regulates the size of spine base in cucumber. New Phytol. 2022, 233, 2643–2658. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Chang, H.; Li, Q.; Wang, L.; Ren, Z.; Ren, H.; Chen, C. Transcriptome profiling reveals genes involved in spine development during CsTTG1-regulated pathway in cucumber (Cucumis sativus L.). Plant Sci. 2020, 291, 110354. [Google Scholar] [CrossRef]
- Zhang, L.; Lv, D.; Pan, J.; Zhang, K.; Wen, H.; Chen, Y.; Du, H.; He, H.; Cai, R.; Pan, J.; et al. A SNP of HD-ZIP I transcription factor leads to distortion of trichome morphology in cucumber (Cucumis sativus L.). BMC Plant Biol. 2021, 21, 182. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wen, C.; Weng, Y. Fine mapping of the pleiotropic locus B for black spine and orange mature fruit color in cucumber identifies a 50 kb region containing a R2R3-MYB transcription factor. Theor. Appl. Genet. 2013, 126, 2187–2196. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, C.; Duan, L.; Luan, Z.; Li, J.; Yang, Y.; Qi, X.; Ren, Z. CsMYB60 is a key regulator of flavonols and proanthocyanidans that determine the colour of fruit spines in cucumber. J. Exp. Bot. 2019, 70, 69–84. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Win, K.T.; Kim, Y.C.; Lee, S. Two types of mutations in the HEUKCHEEM gene functioning in cucumber spine color development can be used as signatures for cucumber domestication. Planta 2019, 250, 1491–1504. [Google Scholar] [CrossRef]
- Samuels, A.L.; Glass, A.D.M.; Ehret, D.L.; Menzies, J.G. The effects of silicon supplementation on cucumber fruit: Changes in surface characteristics. Annals Bot. 1993, 72, 433–440. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, W.; He, H.; Nie, J.; Bie, B.; Zhao, J.; Ren, G.; Li, Y.; Zhang, D.; Pan, J.; et al. Tuberculate fruit gene Tu encodes a C2 H2 zinc finger protein that is required for the warty fruit phenotype in cucumber (Cucumis sativus L.). Plant J. 2014, 7, 1034–1046. [Google Scholar] [CrossRef]
- Zhao, L.; Zhu, H.; Zhang, K.; Wang, Y.; Wu, L.; Chen, C.; Liu, X.; Yang, S.; Ren, H.; Yang, L. The MIXTA-LIKE transcription factor CsMYB6 regulates fruit spine and tubercule formation in cucumber. Plant Sci. 2020, 300, 110636. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, L.; Han, L.; Cheng, Z.; Li, X.; Wang, S.; Liu, L.; Chen, J.; Song, W.; Zhao, J.; et al. HECATE2 acts with GLABROUS3 and Tu to boost cytokinin biosynthesis and regulate cucumber fruit wart formation. Plant Physiol. 2021, 187, 1619–1635. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wen, C.; Liu, B.; Cai, Y.; Xue, S.; Bartholomew, E.S.; Dong, M.; Jian, C.; Xu, S.; Wang, T.; et al. A CsTu-TS1 regulatory module promotes fruit tubercule formation in cucumber. Plant Biotechnol. J. 2019, 17, 289–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeats, T.H.; Rose, J.K.C. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hen-Avivi, S.; Lashbrooke, J.; Costa, F.; Aharoni, A. Scratching the surface: Genetic regulation of cuticle assembly in fleshy fruit. J. Exp. Bot. 2014, 65, 4653–4664. [Google Scholar] [CrossRef] [Green Version]
- Ando, K.; Carr, K.M.; Colle, M.; Mansfeld, B.N.; Grumet, R. Exocarp properties and transcriptomic analysis of cucumber (Cucumis sativus) fruit expressing age-related resistance to Phytophthora capsici. PLoS ONE 2015, 10, e0142133. [Google Scholar] [CrossRef] [PubMed]
- Rett-Cadman, S.; Colle, M.; Mansfeld, B.; Barry, C.S.; Wang, Y.; Weng, Y.; Gao, L.; Fei, Z.; Grumet, R. QTL and transcriptomic analyses implicate cuticle transcription factor SHINE as a source of natural variation for epidermal traits in cucumber fruit. Front. Plant Sci. 2019, 10, 1536. [Google Scholar] [CrossRef]
- Lara, I.; Heredia, A.; Domínguez, E. Shelf life potential and the fruit cuticle: The unexpected player. Front. Plant Sci. 2019, 10, 770. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Du, Y.; Long, Z.; Li, Y.; Kong, W.; Wang, H.; Wei, A.; Du, S.; Yang, R.; Li, J.; et al. Fine mapping of a novel QTL CsFSG1 for fruit skin gloss in cucumber (Cucumis sativus L.). Molec. Breed. 2022, 42, 1–8. [Google Scholar] [CrossRef]
- Compagnon, V.; Diehl, P.; Benveniste, I.; Meyer, D.; Schaller, H.; Schreiber, L.; Franke, R.; Pinot, F. CYP86B1 is required for very long chain omega-hydroxyacid and alpha, omega -dicarboxylic acid synthesis in root and seed suberin polyester. Plant Physiol. 2009, 150, 1831–1843. [Google Scholar] [CrossRef]
- Isaacson, T.; Kosma, D.K.; Matas, A.J.; Buda, G.J.; He, Y.; Yu, B.; Pravitasari, A.; Batteas, J.D.; Stark, R.E.; Jenks, M.A.; et al. Cutin deficiency in the tomato fruit cuticle consistently affects resistance to microbial infection and biomechanical properties, but not transpirational water loss. Plant J. 2009, 60, 363–377. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, Y.; Xu, C.; Ren, J.; Liu, X.; Black, K.; Gai, X.; Wang, Q.; Ren, H. Cucumber ECERIFERUM1 (CsCER1), which influences the cuticle properties and drought tolerance of cucumber, plays a key role in VLC alkanes biosynthesis. Plant Molec. Biol. 2014, 87, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, S.; Li, M.; Hou, L. Cloning and expression analysis of Cucumis sativus L. CER4 involved in cuticular wax biosynthesis in cucumber. Biotechnol. Biotechnol. Equip. 2018, 32, 1113–1118. [Google Scholar] [CrossRef] [Green Version]
- Aharoni, A.; Dixit, S.; Jetter, R.; Thoenes, E.; Van Arkel, G.; Pereira, A. The SHINE clade of AP2 domain transcription factors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in Arabidopsis. Plant Cell 2004, 16, 2463–2480. [Google Scholar] [CrossRef] [Green Version]
- Broun, P.; Poindexter, P.; Osborne, E.; Jiang, C.Z.; Riechmann, J.L. WIN1, a transcriptional activator of epidermal wax accumulation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 4706–4711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Wang, Y.; Tan, J.; Weng, Y. Functional copy number variation of CsSHINE1 is associated with fruit skin netting intensity in cucumber, Cucumis sativus. Theor. Appl. Genet. 2022, 135, 2101–2119. [Google Scholar] [CrossRef]
- Knoche, M.; Lang, A. Ongoing growth challenges fruit skin integrity. Crit. Rev. Plant Sci. 2017, 36, 190–215. [Google Scholar] [CrossRef]
- Cohen, H.; Dong, Y.; Szymanski, J.; Lashbrooke, J.; Meir, S.; Almekias-Siegl, E.; Zeisler-Diehl, V.V.; Schreiber, L.; Aharoni, A. A multilevel study of melon fruit reticulation provides insight into skin ligno-suberization hallmarks. Plant Physiol. 2019, 179, 1486–1501. [Google Scholar] [CrossRef] [Green Version]
- Nomberg, G.; Marinov, O.; Karavani, E.; Manasherova, E.; Zelinger, E.; Yarden, O.; Cohen, H. Cucumber fruit skin reticulation affects post-harvest traits. Postharvest. Biol. Technol. 2022, 194, 112071. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, B.; Dymerski, R.; Xu, X.; Weng, Y. Quantitative trait loci for horticulturally important traits defining the Sikkim cucumber, Cucumis sativus var. sikkimensis. Theor. Appl. Genet. 2021, 134, 229–247. [Google Scholar] [CrossRef]
- Wang, M.; Chen, L.; Liang, Z.; He, X.; Liu, W.; Jiang, B.; Yan, J.; Sun, P.; Cao, Z.; Peng, Q.; et al. Metabolome and transcriptome analyses reveal chlorophyll and anthocyanin metabolism pathway associated with cucumber fruit skin color. BMC Plant Biol. 2020, 20, 386. [Google Scholar] [CrossRef]
- Jo, H.E.; Son, S.Y.; Lee, C.H. Comparison of metabolome and functional properties of three Korean cucumber cultivars. Front. Plant Sci. 2022, 13, 882120. [Google Scholar] [CrossRef] [PubMed]
- Kishor, D.S.; Alavilli, H.; Lee, S.C.; Kim, J.G.; Song, K. Development of SNP markers for white immature fruit skin color in cucumber (Cucumis sativus L.) using QTL-seq and marker analyses. Plants 2021, 10, 2341. [Google Scholar] [CrossRef] [PubMed]
- Lun, Y.; Wang, X.; Zhang, C.; Yang, L.; Gao, D.; Chen, H.; Huang, S. A CsYcf54 variant conferring light green coloration in cucumber. Euphytica 2016, 208, 509–517. [Google Scholar] [CrossRef]
- Zhou, Q.; Wang, S.; Hu, B.; Chen, H.; Zhang, Z.; Huang, S. An ACCUMULATION AND REPLICATION OF CHLOROPLASTS 5 gene mutation confers light green peel in cucumber. J. Integr. Plant Biol. 2015, 57, 936–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, N.; Du, Y.; Li, H.; Wang, C.; Wang, C.; Gong, S.; Zhou, S.; Wu, T. CsMYB36 is involved in the formation of yellow green peel in cucumber (Cucumis sativus L.). Theor. Appl. Genet. 2018, 131, 1659–1669. [Google Scholar] [CrossRef]
- Huang, H.; Yang, Q.; Zhang, L.; Kong, W.; Wang, H.; Wei, A.; Du, S.; Yang, R.; Li, J.; Lin, T.; et al. Genome-wide association analysis reveals a novel QTL CsPC1 for pericarp color in cucumber. BMC Genom. 2022, 23, 383. [Google Scholar] [CrossRef]
- Liu, H.; Jiao, J.; Liang, X.; Liu, J.; Meng, H.; Chen, S.; Li, Y.; Cheng, Z. Map-based cloning, identification and characterization of the w gene controlling white immature fruit color in cucumber (Cucumis sativus L.). Theor. Appl. Genet. 2016, 129, 1247–1256. [Google Scholar] [CrossRef]
- Jiao, J.; Liu, H.; Liu, J.; Cui, M.; Xu, J.; Meng, H.; Li, Y.; Chen, S.; Cheng, Z. Identification and functional characterization of APRR2 controlling green immature fruit color in cucumber (Cucumis sativus L.). Plant Growth Regul. 2017, 83, 233–243. [Google Scholar] [CrossRef]
- Tang, H.Y.; Dong, X.; Wang, J.K.; Xia, J.H.; Xie, F.; Zhang, Y.; Yao, X.; Xu, Y.J.; Wang, Z.J. Fine mapping and candidate gene prediction for white immature fruit skin in cucumber (Cucumis sativus L.). Int. J. Mol. Sci. 2018, 19, 1493. [Google Scholar] [CrossRef]
- Qi, J.; Liu, X.; Shen, D.I.; Miao, H.; Xie, B.; Li, X.; Zeng, P.; Wang, S.; Shang, Y.; Gu, X.; et al. A genomic variation map provides insights into the genetic basis of cucumber domestication and diversity. Nat. Genet. 2013, 45, 1510–1515. [Google Scholar] [CrossRef]
- Kishor, D.S.; Lee, H.Y.; Alavilli, H.; You, C.R.; Kim, J.G.; Lee, S.Y.; Kang, B.C.; Song, K. Identification of an allelic variant of the CsOr gene controlling fruit endocarp color in cucumber (Cucumis sativus L.) using genotyping-by-sequencing (GBS) and whole-genome sequencing. Front. Plant Sci. 2021, 22, 802864. [Google Scholar] [CrossRef] [PubMed]
- Tzuri, G.; Zhou, X.; Chayut, N.; Yuan, H.; Portnoy, V.; Meir, A.; Sa’ar, U.; Baumkoler, F.; Mazourek, M.; Lewinsohn, E.; et al. A ‘golden’ SNP in CmOr governs the fruit flesh color of melon (Cucumis melo). Plant J. 2015, 82, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Bo, K.; Wei, S.; Wang, W.; Miao, H.; Dong, S.; Zhang, S.; Gu, X. QTL mapping and genome-wide association study reveal two novel loci associated with green flesh color in cucumber. BMC Plant Biol. 2019, 19, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.; Chen, C.; Liu, X.; Yang, K.; Wang, C.; Lu, X.; Tian, Y.; Chen, H. The formation of hollow trait in cucumber (Cucumis sativus L.) fruit is controlled by CsALMT2. Int. J. Mol. Sci. 2022, 23, 6173. [Google Scholar] [CrossRef]
- Ando, K.; Grumet, R. Transcriptional profiling of rapidly growing cucumber fruit by 454-pyrosequencing analysis. J. Am. Soc. Hortic. Sci. 2010, 135, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Lu, L.; Zhu, B.; Xu, Q.; Qi, X.; Chen, X. QTL mapping of cucumber fruit flesh thickness by SLAF-seq. Sci. Rep. 2015, 5, 15829. [Google Scholar] [CrossRef] [Green Version]
- Horvath, D.P.; Schaffer, R.; West, M.; Wisman, E. Arabidopsis microarrays identify conserved and differentially expressed genes involved in shoot growth and development from distantly related plant species. Plant J. 2003, 34, 125–134. [Google Scholar] [CrossRef]
- Qi, X.; Zhu, Y.; Li, S.; Zhou, H.; Xu, X.; Xu, Q.; Chen, X. Identification of genes related to mesocarp development in cucumber. Hortic. Plant J. 2020, 6, 293–300. [Google Scholar] [CrossRef]
- Lin, X.; Zhang, N.; Song, H.; Lin, K.; Pang, E. Population-specific, recent positive selection signatures in cultivated Cucumis sativus L. (cucumber). G3 2022, 12, jkac119. [Google Scholar] [CrossRef]
- Wenzel, G.; Kennard, W.C.; Havey, M.J. Quantitative trait analysis of fruit quality in cucumber: QTL detection, confirmation, and comparison with mating-design variation. Theor Appl Genet. 1995, 91, 53–61. [Google Scholar] [CrossRef]
- Yuan, X.J.; Li, X.Z.; Pan, J.S.; Wang, G.; Jiang, S.; Li, X.H.; Deng, S.L.; He, H.L.; Si, M.X.; Lai, L.; et al. Genetic linkage map construction and location of QTLs for fruit-related traits in cucumber. Plant Breed. 2008, 127, 180–188. [Google Scholar] [CrossRef]
- Wilson, J.E.; Baker, L.R. Inheritance of carpel separation in mature fruits of pickling cucumbers. J. Am. Soc. Hortic. Sci. 1976, 101, 66–69. [Google Scholar] [CrossRef]
- Cheng, Z.; Song, X.; Liu, X.; Yan, S.; Song, W.; Wang, Z.; Han, L.; Zhao, J.; Yan, L.; Zhou, Z.; et al. SPATULA and ALCATRAZ confer female sterility and fruit cavity via mediating pistil development in cucumber. Plant Physiol. 2022, 189, 1553–1569. [Google Scholar] [CrossRef] [PubMed]
- Jawad, M.U.; Gao, L.; Gebremeskel, H.; Safdar, L.B.; Yuan, P.; Zhao, S.; Xuqiang, L.; Nan, H.; Zhu, H.; Liu, W. Expression pattern of sugars and organic acids regulatory genes during watermelon fruit development. Sci. Hortic. 2020, 265, 109102. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grumet, R.; Lin, Y.-C.; Rett-Cadman, S.; Malik, A. Morphological and Genetic Diversity of Cucumber (Cucumis sativus L.) Fruit Development. Plants 2023, 12, 23. https://doi.org/10.3390/plants12010023
Grumet R, Lin Y-C, Rett-Cadman S, Malik A. Morphological and Genetic Diversity of Cucumber (Cucumis sativus L.) Fruit Development. Plants. 2023; 12(1):23. https://doi.org/10.3390/plants12010023
Chicago/Turabian StyleGrumet, Rebecca, Ying-Chen Lin, Stephanie Rett-Cadman, and Ajaz Malik. 2023. "Morphological and Genetic Diversity of Cucumber (Cucumis sativus L.) Fruit Development" Plants 12, no. 1: 23. https://doi.org/10.3390/plants12010023
APA StyleGrumet, R., Lin, Y. -C., Rett-Cadman, S., & Malik, A. (2023). Morphological and Genetic Diversity of Cucumber (Cucumis sativus L.) Fruit Development. Plants, 12(1), 23. https://doi.org/10.3390/plants12010023