
The Influence of Chestnut Extract and Its Components on Antibacterial Activity against Staphylococcus aureus
 , , ,
, , , 
Abstract
:1. Introduction
2. Results and Discussion
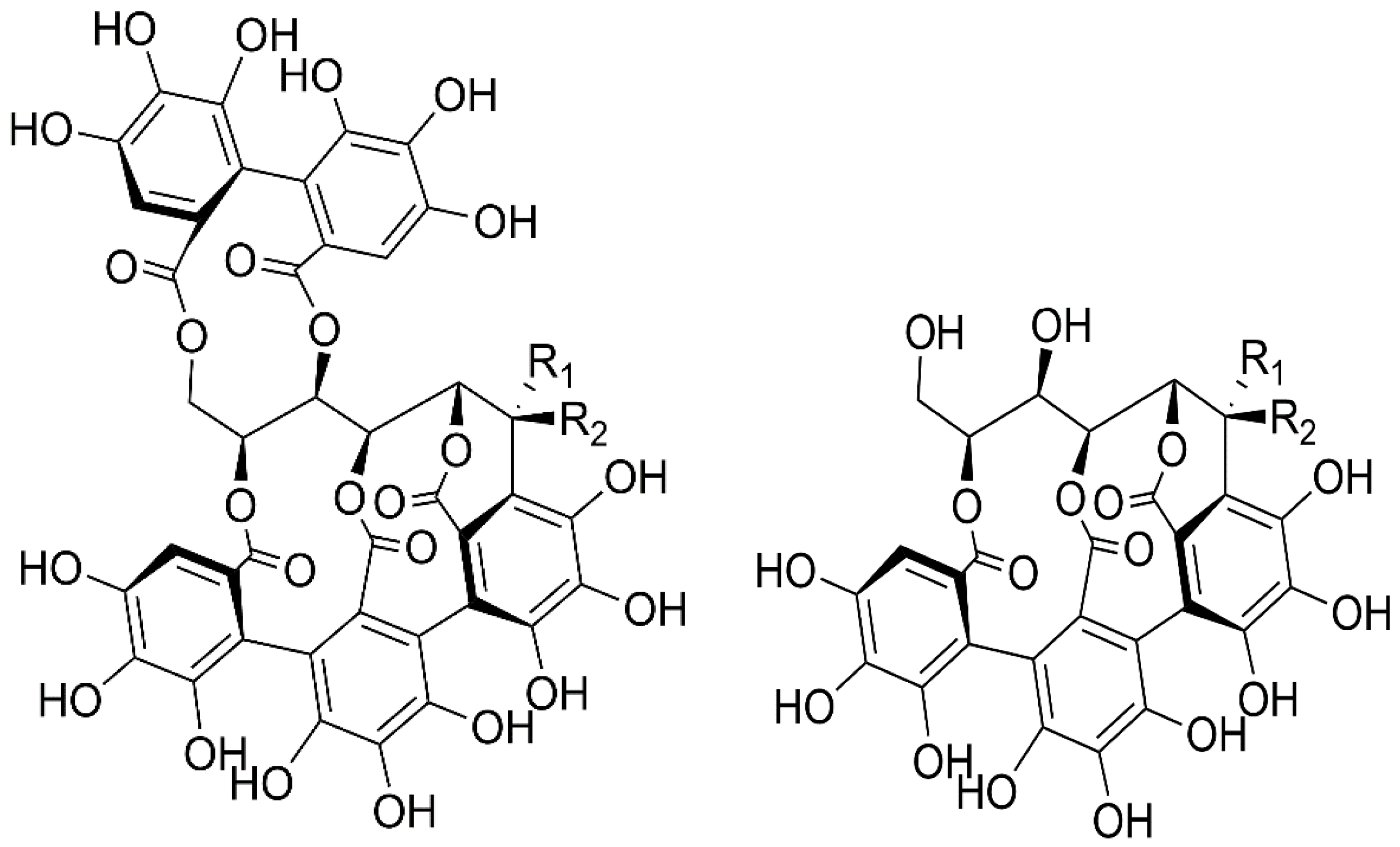
2.1. Identification of Isolated Vescalin, Castalin, Vescalagin and Castalagin
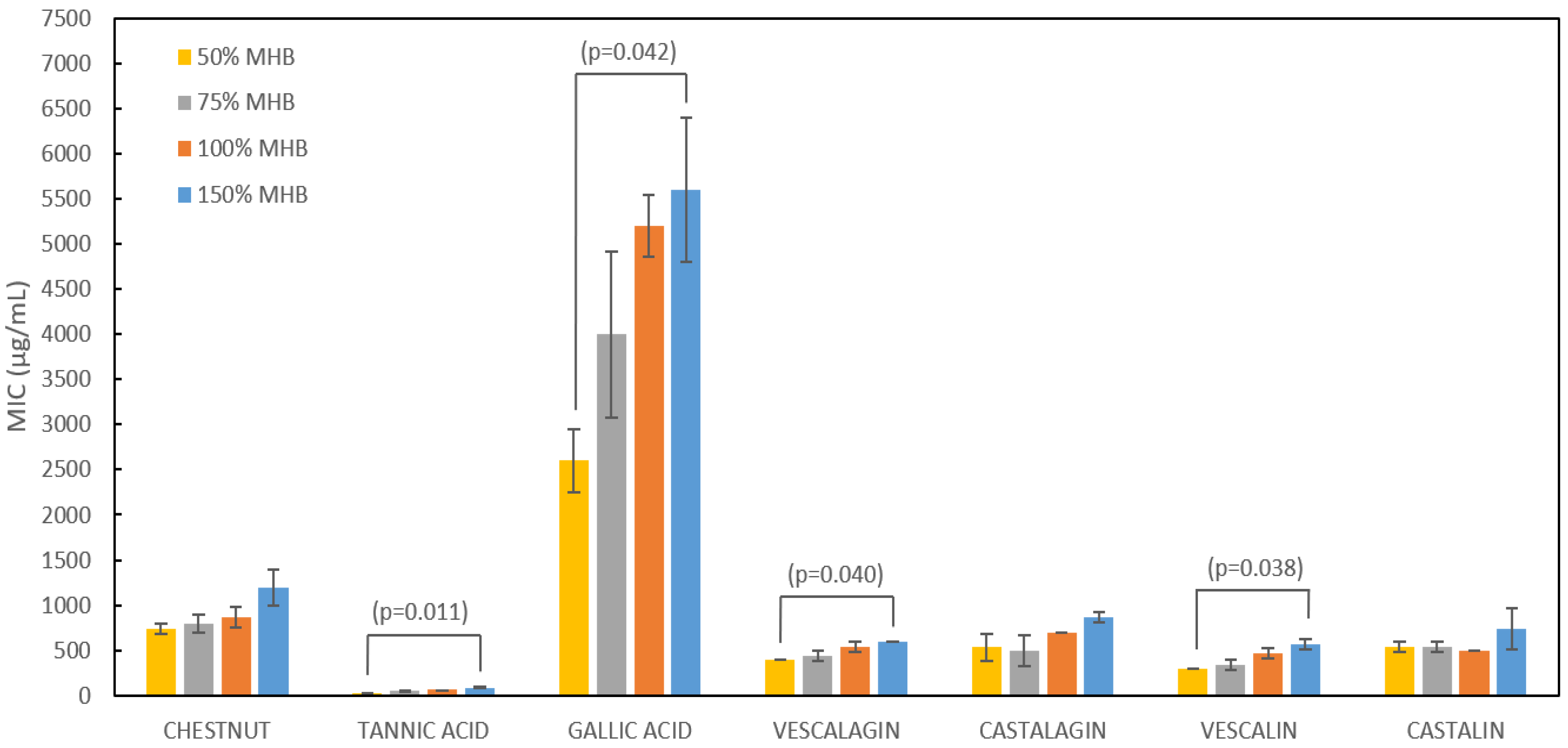
2.2. Variation of the Growth Medium Strength and Its Influence on MIC
2.3. Minimum Inhibitory Concentrations (MICs) of Studied Samples against Staphylococcus aureus
2.4. Minimum Bactericidal Concentrations (MBCs) of Studied Samples against S. aureus
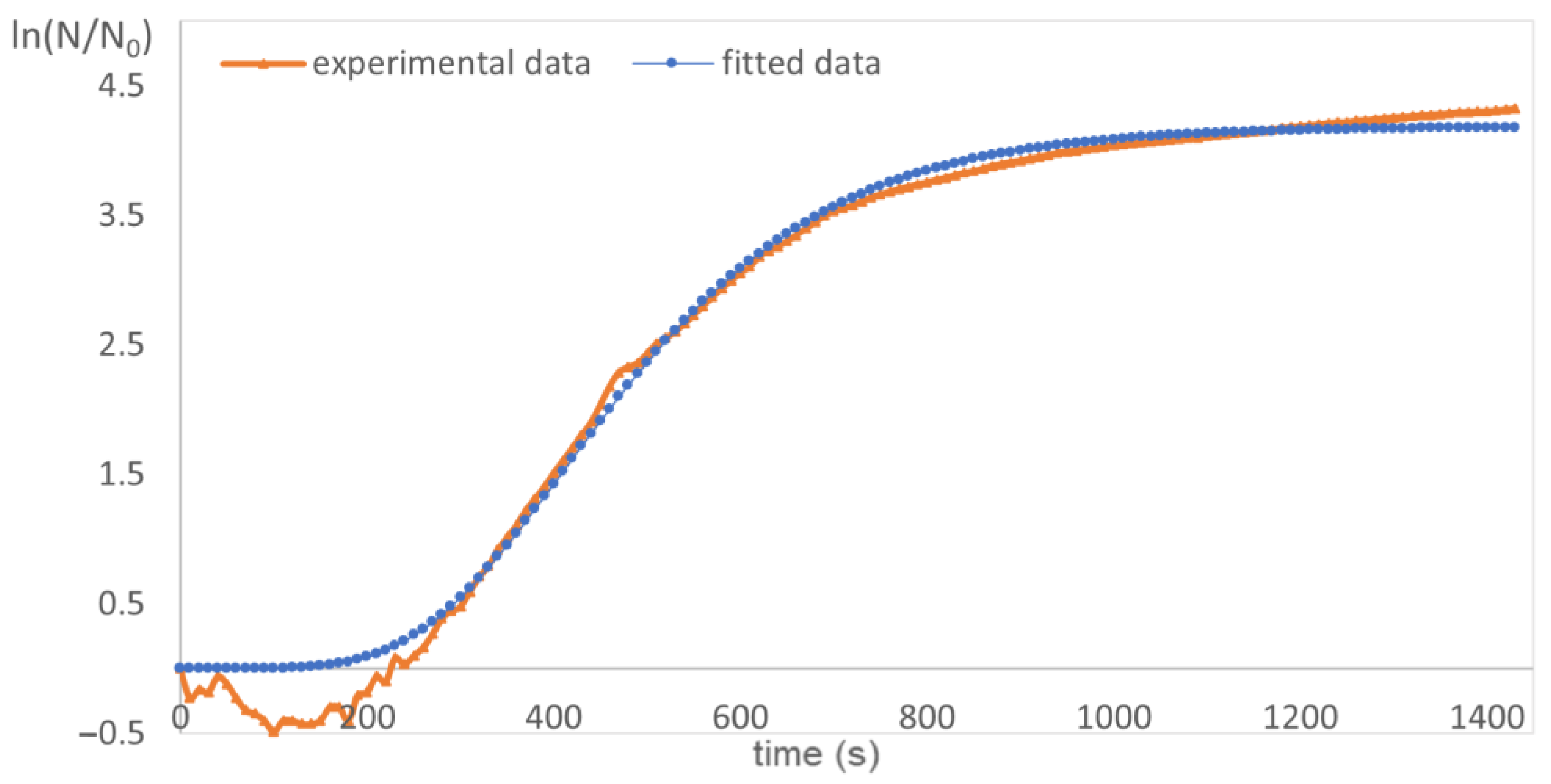
2.5. Lag Phases, Growth Rates and Generation Times
3. Materials and Methods
3.1. Antibacterial Agents
3.2. Isolation of Vescalin and Castalin
3.3. Identification of Isolated Compounds Using HPLC
3.4. Antibacterial Assay
3.4.1. Microorganisms and Culture Conditions
3.4.2. Preparation of Antimicrobial Agents
3.4.3. Broth Microdilution Assay
3.4.4. Determination of Minimum Inhibitory Concentration (MIC)
3.4.5. Determination of Minimum Bactericidal Concentration (MBC)
3.5. Statistical Analysis
3.6. Lag Phases, Growth Rates and Generation Times
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Poole, K. Overcoming antimicrobial resistance by targeting resistance mechanisms. J. Pharm. Pharmacol. 2001, 53, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Hostnik, G.; Gladović, M.; Bren, U. Tannin Basic Building Blocks as Potential Scavengers of Chemical Carcinogens: A Computational Study. J. Nat. Prod. 2019, 82, 3279–3287. [Google Scholar] [CrossRef] [PubMed]
- Tosovic, J.; Bren, U. Antioxidative Action of Ellagic Acid-A Kinetic DFT Study. Antioxidants 2020, 9, 587. [Google Scholar] [CrossRef] [PubMed]
- Braga, N.; Rodrigues, F.; Oliveira, M.B. Castanea sativa by-products: A review on added value and sustainable application. Nat. Prod. Res. 2015, 29, 1–18. [Google Scholar] [CrossRef]
- Stumpf, S.; Hostnik, G.; Primožič, M.; Leitgeb, M.; Salminen, J.-p.; Bren, U. The effect of growth medium strength on minimum inhibitory concentrations of tannins and tannin extracts against E. coli. Molecules 2020, 25, 2947. [Google Scholar] [CrossRef]
- Marston, H.D.; Dixon, D.M.; Knisely, J.M.; Palmore, T.N.; Fauci, A.S. Antimicrobial Resistance. JAMA 2016, 316, 1193–1204. [Google Scholar] [CrossRef]
- Nicoloff, H.; Hjort, K.; Levin, B.R.; Andersson, D.I. The high prevalence of antibiotic heteroresistance in pathogenic bacteria is mainly caused by gene amplification. Nat. Microbiol. 2019, 4, 504–514. [Google Scholar] [CrossRef]
- Abreu, A.C.; McBain, A.J.; Simoes, M. Plants as sources of new antimicrobials and resistance-modifying agents. Nat. Prod. Rep. 2012, 29, 1007–1021. [Google Scholar] [CrossRef]
- Conedera, M.; Krebs, P.; Tinner, W.; Pradella, M.; Torriani, D. The cultivation of Castanea sativa (Mill.) in Europe, from its origin to its diffusion on a continental scale. Veg. Hist. Archaeobotany 2004, 13, 161–179. [Google Scholar] [CrossRef]
- De Vasconcelos, M.C.; Bennett, R.N.; Rosa, E.A.; Ferreira-Cardoso, J.V. Composition of European chestnut (Castanea sativa Mill.) and association with health effects: Fresh and processed products. J. Sci. Food Agric. 2010, 90, 1578–1589. [Google Scholar] [CrossRef]
- Gonçalves, B.; Borges, O.; Costa, H.S.; Bennett, R.; Santos, M.; Silva, A.P. Metabolite composition of chestnut (Castanea sativa Mill.) upon cooking: Proximate analysis, fibre, organic acids and phenolics. Food Chem. 2010, 122, 154–160. [Google Scholar] [CrossRef]
- Khanbabaee, K.; Ree, T.V. Tannins: Classification and definition. R. Soc. Chem. 2001, 18, 641–649. [Google Scholar] [CrossRef]
- Serrano, J.; Puupponen-Pimiä, R.; Dauer, A.; Aura, A.-M.; Saura-Calixto, F. Tannins: Current knowledge of food sources, intake, bioavailability and biological effects. Mol. Nutr. Food Resour. 2009, 53, 310–329. [Google Scholar] [CrossRef] [PubMed]
- Mämmelä, P.; Savolainen, H.; Lindroos, L.; Kangas, J.; Vartiainen, T. Analysis of oak tannins by liquid chromatography-electrospray ionisation mass spectrometry. J. Chromatogr. A 2000, 891, 75–83. [Google Scholar] [CrossRef]
- Salminen, J.-P.; Ossipov, V.; Loponen, J.; Haukioja, E.; Pihlaja, K. Characterisation of hydrolysable tannins from leaves of Betula pubescens by high-performance liquid chromatography—Mass spectrometry. J. Chromatogr. A 1999, 864, 283–291. [Google Scholar] [CrossRef]
- Mueller-Harvey, I. Analysis of hydrolysable tannins. Anim. Feed. Sci. Technol. 2001, 91, 3–20. [Google Scholar] [CrossRef]
- Živković, J.; Mujić, I.; Zeković, Z.; Nikolić, G.; Vidović, S.; Mujić, A. Extraction and analysis of condensed tannins in Castanea sativa Mill. J. Cent. Eur. Agric. 2009, 10, 283–288. [Google Scholar]
- Das, A.K.; Islam, M.N.; Faruk, M.O.; Ashaduzzaman, M.; Dungani, R. Review on tannins: Extraction processes, applications and possibilities. S. Afr. J. Bot. 2020, 135, 58–70. [Google Scholar] [CrossRef]
- Sanz, M.; Cadahia, E.; Esteruelas, E.; Munoz, A.M.; Fernandez de Simon, B.; Hernandez, T.; Estrella, I. Phenolic compounds in chestnut (Castanea sativa Mill.) heartwood. Effect of toasting at cooperage. J. Agric. Food Chem. 2010, 58, 9631–9640. [Google Scholar] [CrossRef]
- Scalbert, A.; Monties, B.; Janin, G. Tannins in Wood: Comparison of Different Estimation Methods. J. Agric. Food Chem. 1989, 37, 1324–1329. [Google Scholar] [CrossRef]
- Scalbert, A. Antimicrobial properties of tannins. Phytochemistry 1991, 30, 3875–3883. [Google Scholar] [CrossRef]
- Chu, E.H.Y.; Arbor, A.; Hollaender, M.A.; Nicoletti, B.; Kopits, S.E.; Ascani, E.; McKusick, V.A.; Sutherland, B.M.; Woodhead, A.D.; Harling, O.K.; et al. Plant Polyphenols: Synthesis, Properties, Significance; Plenum Press: New York, NY, USA, 1991; Volume 59. [Google Scholar]
- Tian, F.; Li, B.; Ji, B.; Zhang, G.; Luo, Y. Identification and structure–activity relationship of gallotannins separated from Galla chinensis. LWT—Food Sci. Technol. 2009, 42, 1289–1295. [Google Scholar] [CrossRef]
- Ikigai, H.; Nakae, T.; Hara, Y.; Shimamura, T. Bactericidal catechins damage the lipid bilayer. Biochim. Biophys. Acta 1993, 1147, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Taguri, T.; Tanaka, T.; Kouno, I. Antibacterial Spectrum of Plant Polyphenols and Extracts Depending upon Hydroxyphenyl Structure. Biol. Pharm. Bull. 2006, 29, 2226–2235. [Google Scholar] [CrossRef]
- Anderson, R.C.; Vodovnik, M.; Min, B.R.; Pinchak, W.E.; Krueger, N.A.; Harvey, R.B.; Nisbet, D.J. Bactericidal effect of hydrolysable and condensed tannin extracts on Campylobacter jejuni in vitro. Folia Microbiol. 2012, 57, 253–258. [Google Scholar] [CrossRef]
- Akiyama, H.; Fujii, K.; Yamasaki, O.; Oono, T.; Iwatsuki, K. Antibacterial action of several tannins against Staphylococcus aureus. J. Antimicrob. Chemother. 2001, 48, 487–491. [Google Scholar] [CrossRef]
- Basile, A.; Sorbo, S.; Giordano, S.; Ricciardi, L.; Ferrara, S.; Montesano, D.; Castaldo Cobianchi, R.; Vuotto, M.L.; Ferrara, L. Antibacterial and allelopathic activity of extract from Castanea sativa leaves. Fitoterapia 2000, 71, S110–S116. [Google Scholar] [CrossRef]
- Puljula, E.; Walton, G.; Woodward, M.J.; Karonen, M. Antimicrobial Activities of Ellagitannins against Clostridiales perfringens, Escherichia coli, Lactobacillus plantarum and Staphylococcus aureus. Molecules 2020, 25, 3714. [Google Scholar] [CrossRef]
- Funatogawa, K.; Hayashi, S.; Shimomura, H.; Yoshida, T.; Hatano, T.; Ito, H.; Hirai, Y. Antibacterial Activity of Hydrolyzable Tannins Derived from Medicinal Plants against Helicobacter pylori. Microbiol. Immunol. 2004, 48, 251–261. [Google Scholar] [CrossRef]
- Freser, F.; Hostnik, G.; Tosovic, J.; Bren, U. Dependence of the Fe(II)-Gallic Acid Coordination Compound Formation Constant on the pH. Foods 2021, 10, 2689. [Google Scholar] [CrossRef] [PubMed]
- Araujo, A.R.; Araujo, A.C.; Reis, R.L.; Pires, R.A. Vescalagin and Castalagin Present Bactericidal Activity toward Methicillin-Resistant Bacteria. ACS Biomater. Sci. Eng. 2021, 7, 1022–1030. [Google Scholar] [CrossRef] [PubMed]
- Engström, M.T.; Arvola, J.; Nenonen, S.; Virtanen, V.T.J.; Leppä, M.M.; Tähtinen, P.; Salminen, J.P. Structural Features of Hydrolyzable Tannins Determine Their Ability to Form Insoluble Complexes with Bovine Serum Albumin. J. Agric. Food Chem. 2019, 67, 6798–6808. [Google Scholar] [CrossRef] [PubMed]
- Engstrom, M.T.; Virtanen, V.; Salminen, J.P. Influence of the Hydrolyzable Tannin Structure on the Characteristics of Insoluble Hydrolyzable Tannin-Protein Complexes. J. Agric. Food. Chem. 2022, 70, 13036–13048. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, V.; Green, R.J.; Karonen, M. Interactions between Hydrolysable Tannins and Lipid Vesicles from Escherichia coli with Isothermal Titration Calorimetry. Molecules 2022, 27, 3204. [Google Scholar] [CrossRef]
- Taguri, T.; Tanaka, T.; Kouno, I. Antimicrobial Activity of 10 Different Plant Polyphenols against Bacteria Causing Food-Borne Disease. Biol. Pharm. Bull. 2004, 27, 1965–1969. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.T.; Stevens, S.E., Jr.; Lin, W.F.; Wei, C.I. Growth inhibition of selected food-borne bacteria by tannic acid, propyl gallate and related compounds. Lett. Appl. Microbiol. 1993, 17, 29–32. [Google Scholar] [CrossRef]
- Li, B.; Qiu, Y.; Shi, H.; Yin, H. The importance of lag time extension in determining bacterial resistance to antibiotics. Analyst 2016, 141, 3059–3067. [Google Scholar] [CrossRef]
- Vogel, S.J.; Tank, M.; Goodyear, N. Variation in detection limits between bacterial growth phases and precision of an ATP bioluminescence system. Lett. Appl. Microbiol. 2014, 58, 370–375. [Google Scholar] [CrossRef]
- Hamill, P.G.; Stevenson, A.; McMullan, P.E.; Williams, J.P.; Lewis, A.D.R.; Sudharsan, S.; Stevenson, K.E.; Farnsworth, K.D.; Khroustalyova, G.; Takemoto, J.Y.; et al. Microbial lag phase can be indicative of, or independent from, cellular stress. Sci. Rep. 2020, 10, 5948. [Google Scholar] [CrossRef]
- Stumpf, S.; Hostnik, G.; Primozic, M.; Leitgeb, M.; Bren, U. Generation Times of E. coli Prolong with Increasing Tannin Concentration while the Lag Phase Extends Exponentially. Plants 2020, 9, 1680. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Minimum Inhibitory Concentration (MIC) | ||
|---|---|---|
| Sample | (µg/mL) | µmol/mL |
| chestnut extract | 867 ± 115 | / |
| tannic acid | 60 ± 0 | 0.04 ± 0.00 |
| gallic acid | 5200 ± 346 | 27.64 ± 0.61 |
| vescalagin | 533 ± 58 | 0.57 ± 0.06 |
| castalagin | 700 ± 0 | 0.75 ± 0.00 |
| vescalin | 350 ± 58 | 0.55 ± 0.16 |
| castalin | 450 ± 0 | 0.71 ± 0.00 |
| streptomycin | 3.25 ± 0.87 | 0.004 ± 0.001 |
| Minimum Bactericidal Concentration (MBC) | |
|---|---|
| Samples | (µg/mL) |
| chestnut extract | 2000 |
| tannic acid | aNI |
| gallic acid | 10,000 |
| vescalagin | 1500 |
| castalagin | 1500 |
| vescalin | 1000 |
| castalin | 1000 |
| streptomycin | 8 |
| Samples | Concentration (µg/mL) | µ (min−1) | lag (min) | td (min) |
|---|---|---|---|---|
| negative control | 0 | 0.0208 | 100 | 33 |
| chestnut extract | 400 | 0.0141 | 269 | 49 |
| 500 | 0.0144 | 267 | 48 | |
| 600 | 0.0119 | 284 | 58 | |
| 700 | 0.0119 | 587 | 58 | |
| tannic acid | 300 | 0.0109 | 189 | 64 |
| 400 | 0.0074 | 299 | 94 | |
| 500 | 0.0101 | 289 | 69 | |
| gallic acid | 2400 | 0.0141 | 179 | 49 |
| 3000 | 0.0168 | 273 | 41 | |
| 3600 | 0.0120 | 312 | 58 | |
| 4200 | 0.0082 | 355 | 84 | |
| vescalagin | 500 | 0.0126 | 513 | 55 |
| castalagin | 300 | 0.0113 | 247 | 61 |
| 400 | 0.0098 | 253 | 71 | |
| 500 | 0.0085 | 260 | 81 | |
| 600 | 0.0056 | 407 | 123 | |
| vescalin | 100 | 0.0168 | 281 | 41 |
| 200 | 0.0144 | 382 | 48 | |
| 300 | 0.0111 | 537 | 62 | |
| castalin | 400 | 0.0151 | 438 | 46 |
| streptomycin | 0.25 | 0.018 | 164 | 39 |
| 0.5 | 0.012 | 220 | 57 | |
| 1 | 0.013 | 320 | 53 | |
| 2 | 0.0045 | 922 | 152 |
| t (min) | Solvent A (%) | Solvent B (%) | Solvent C (%) |
|---|---|---|---|
| 0.0–60.0 | 100 | 0 | 0 |
| 60.4–135.0 | 70 | 30 | 0 |
| 135.4–185.0 | 50 | 50 | 0 |
| 185.4–197.5 | 90 | 0 | 10 |
| 197.9–210.0 | 70 | 0 | 30 |
| 210.4–222.5 | 50 | 0 | 50 |
| 222.9–240.9 | 25 | 0 | 75 |
| 248.3–260.0 | 50 | 50 | 0 |
| 260.4–309.1 | 100 | 0 | 0 |
| t (min) | Solvent A (%) | Solvent B (%) |
|---|---|---|
| 0.0–6.4 | 98 | 2 |
| 8.6–20.1 | 2 | 98 |
| 22.2–29.1 | 98 | 2 |
| t (min) | Solvent A (%) | Solvent B (%) |
|---|---|---|
| 0.0—2.0 | 98 | 2 |
| 14.0 | 50 | 50 |
| 15.0–20.0 | 10 | 90 |
| 22.0–26.0 | 98 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Štumpf, S.; Hostnik, G.; Langerholc, T.; Pintarič, M.; Kolenc, Z.; Bren, U. The Influence of Chestnut Extract and Its Components on Antibacterial Activity against Staphylococcus aureus. Plants 2023, 12, 2043. https://doi.org/10.3390/plants12102043
Štumpf S, Hostnik G, Langerholc T, Pintarič M, Kolenc Z, Bren U. The Influence of Chestnut Extract and Its Components on Antibacterial Activity against Staphylococcus aureus. Plants. 2023; 12(10):2043. https://doi.org/10.3390/plants12102043
Chicago/Turabian StyleŠtumpf, Sara, Gregor Hostnik, Tomaž Langerholc, Maša Pintarič, Zala Kolenc, and Urban Bren. 2023. "The Influence of Chestnut Extract and Its Components on Antibacterial Activity against Staphylococcus aureus" Plants 12, no. 10: 2043. https://doi.org/10.3390/plants12102043
APA StyleŠtumpf, S., Hostnik, G., Langerholc, T., Pintarič, M., Kolenc, Z., & Bren, U. (2023). The Influence of Chestnut Extract and Its Components on Antibacterial Activity against Staphylococcus aureus. Plants, 12(10), 2043. https://doi.org/10.3390/plants12102043