
Human Settlement and Landscape Anthropization of Remote Oceanic Islands: A Comparison between Rapa Nui (Pacific Ocean) and the Azores (Atlantic Ocean)

Abstract
:1. Introduction
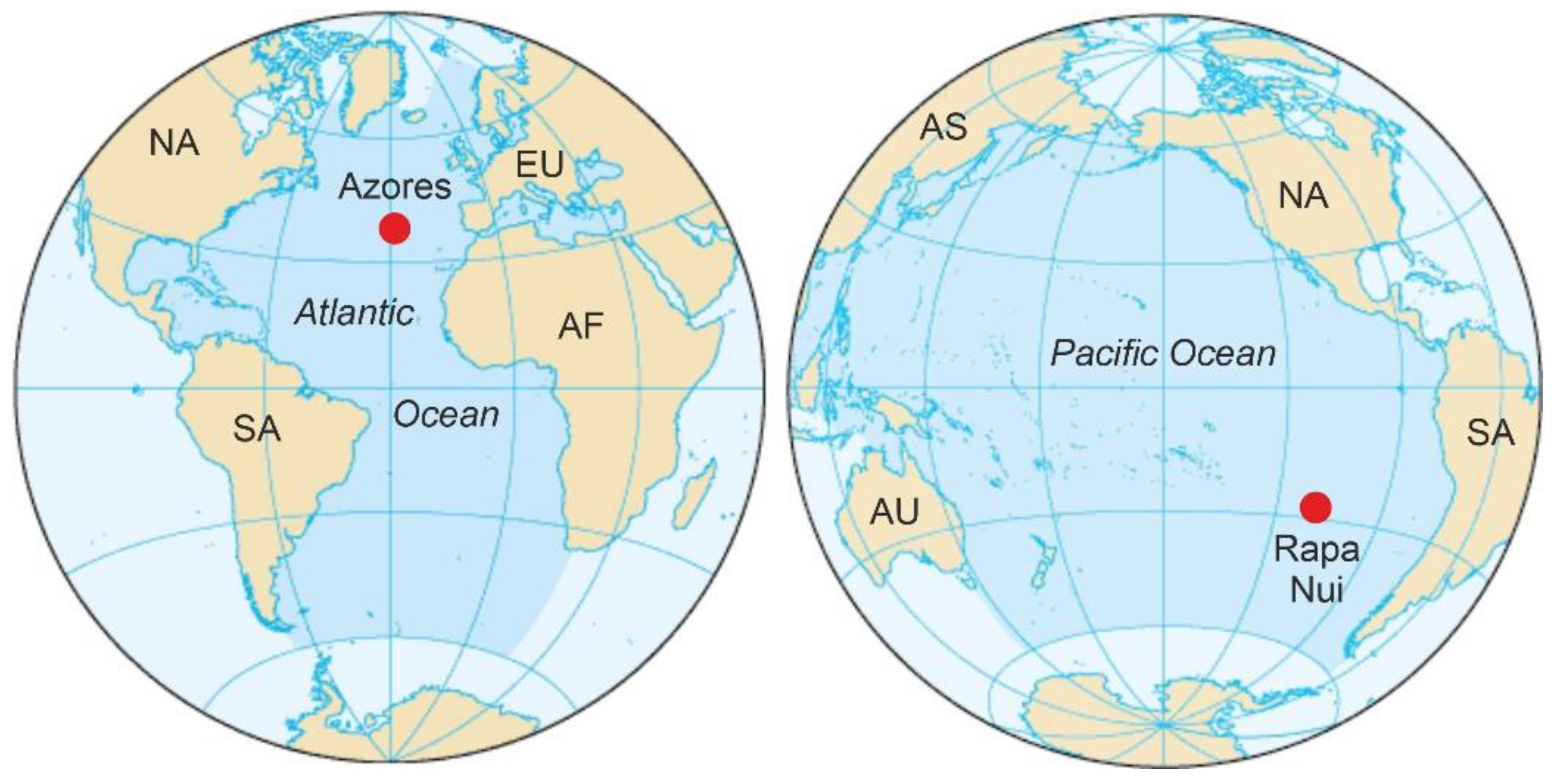
2. Rapa Nui and the Azores
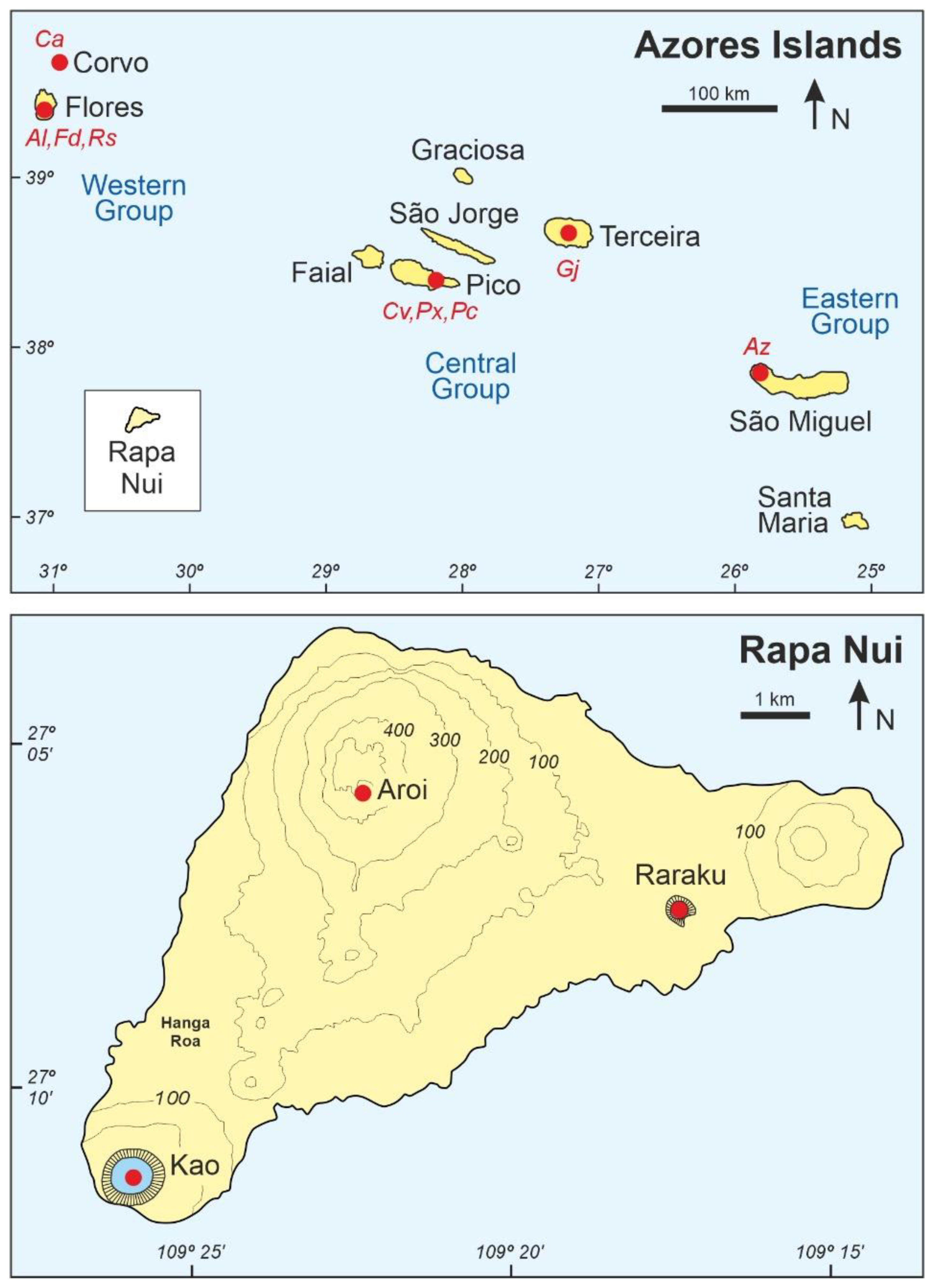
2.1. Rapa Nui
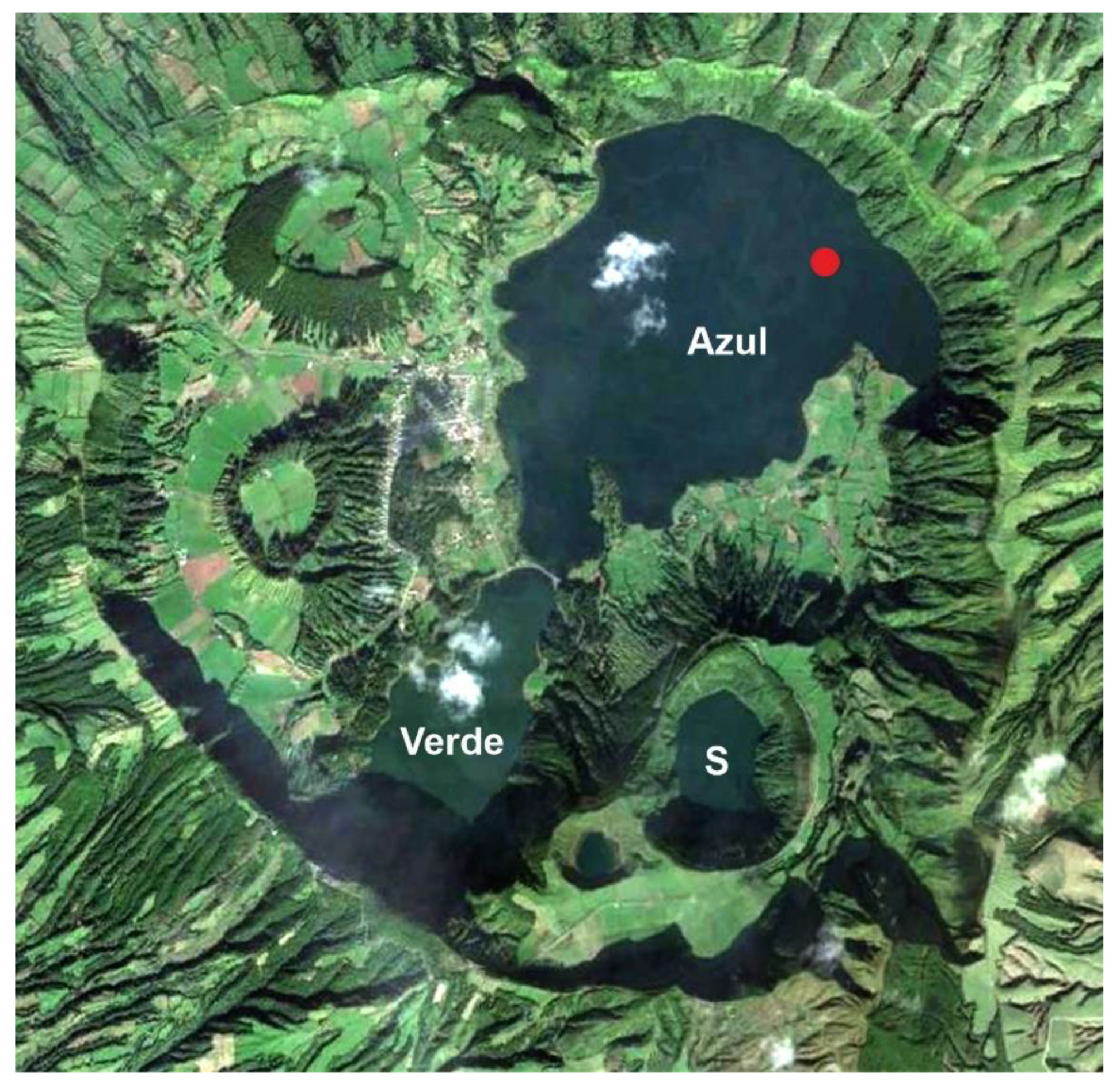
2.2. The Azores
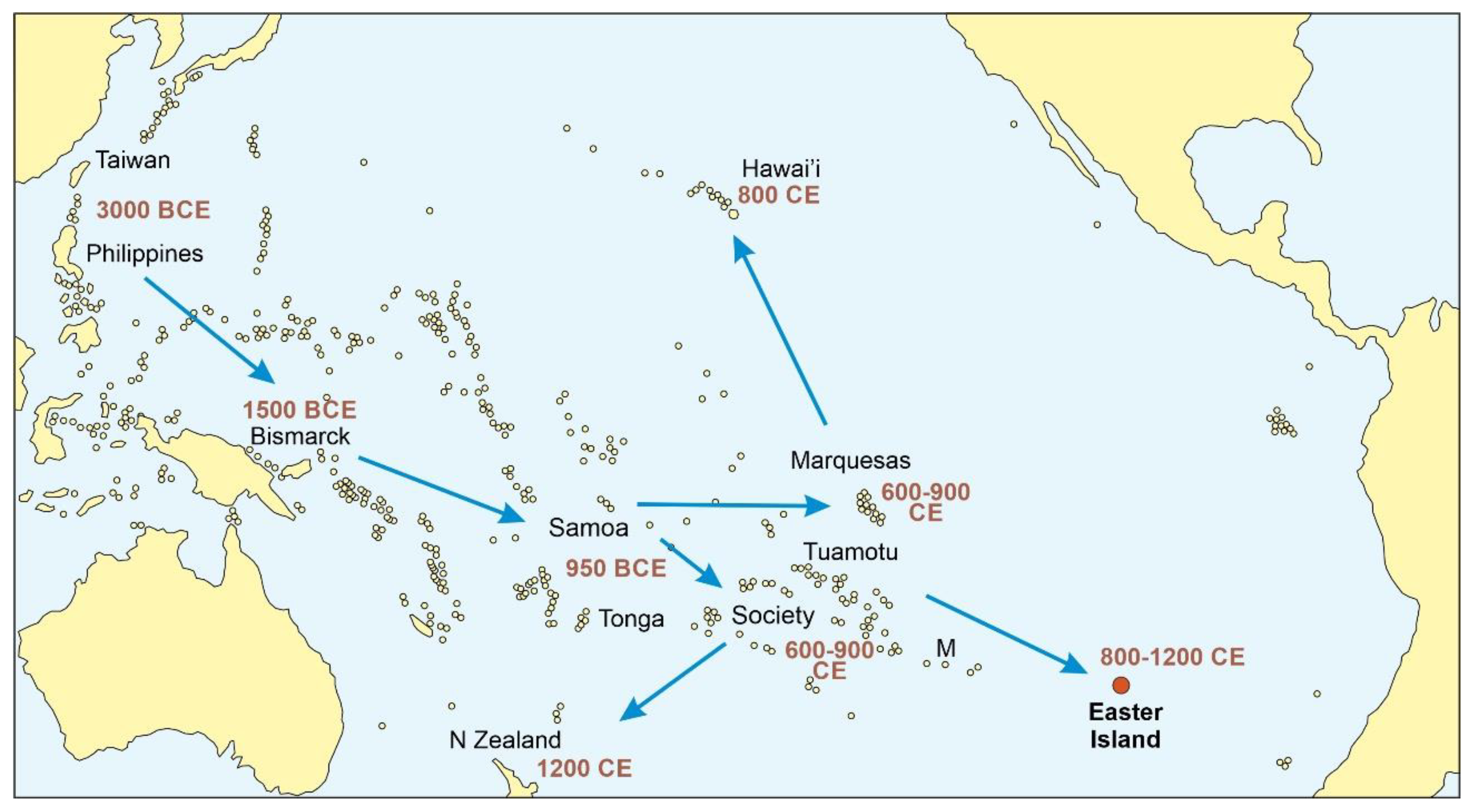
3. Discovery and Settlement
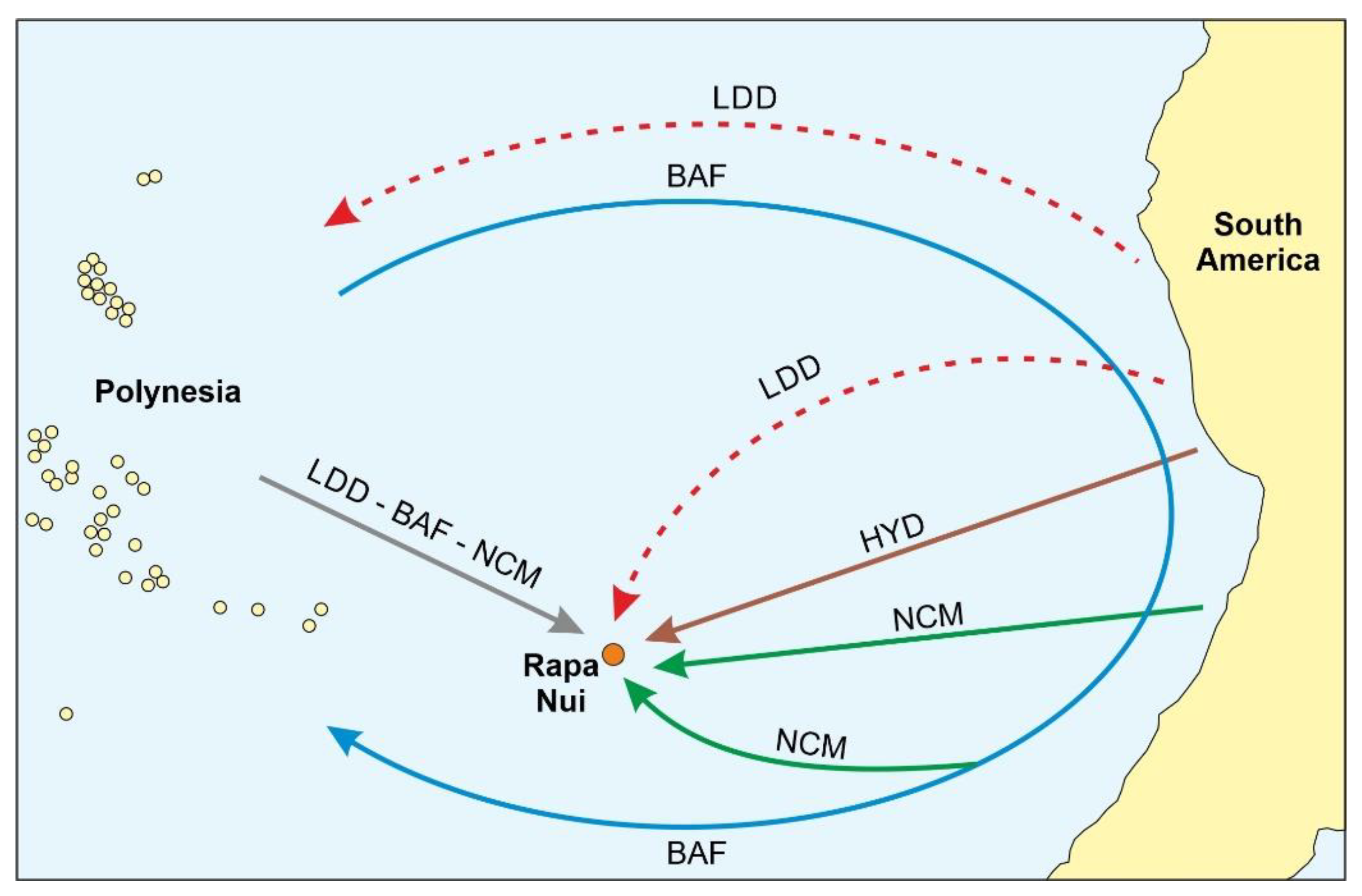
3.1. Rapa Nui
3.2. Azores
4. Deforestation and Landscape Transformation
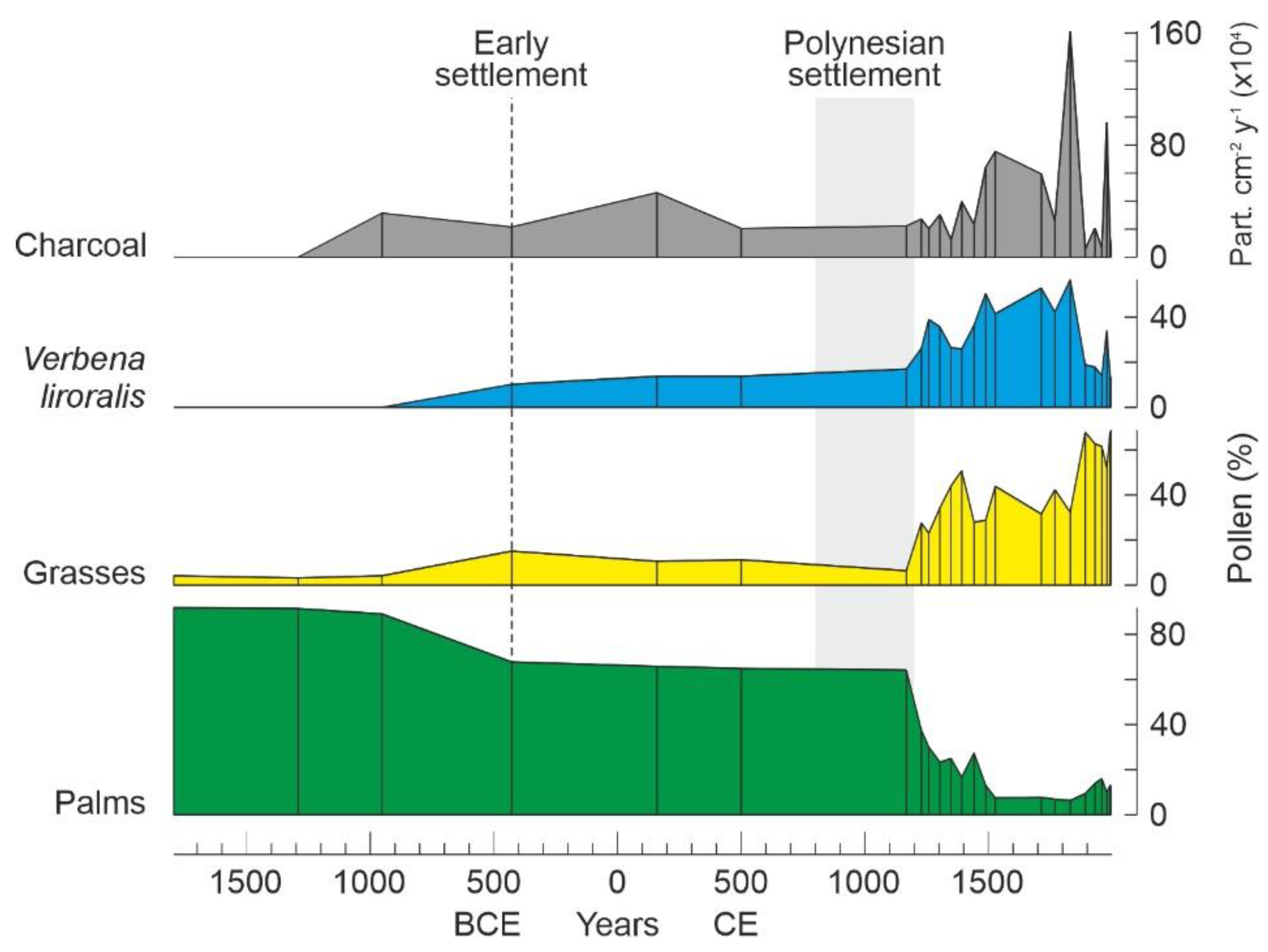
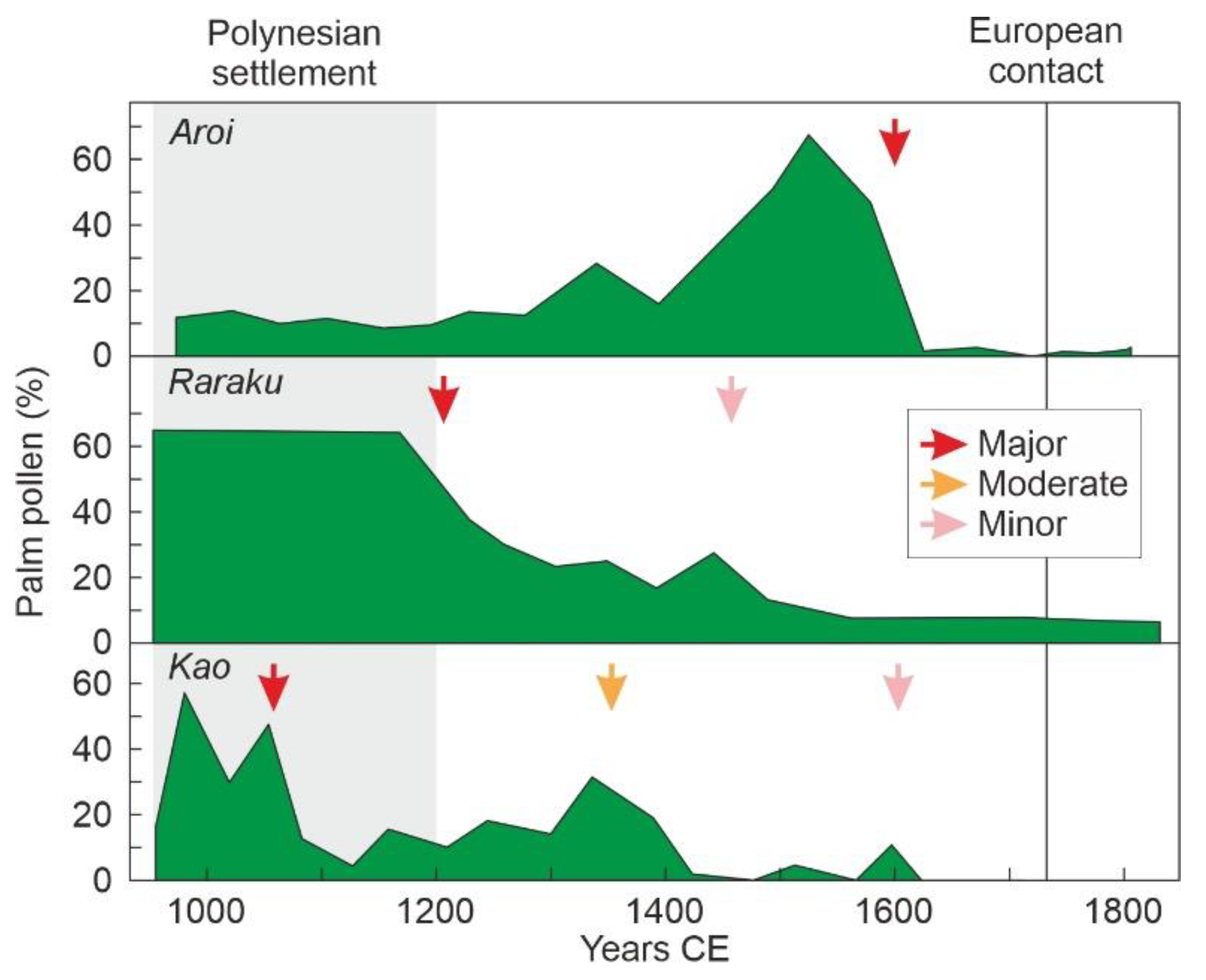
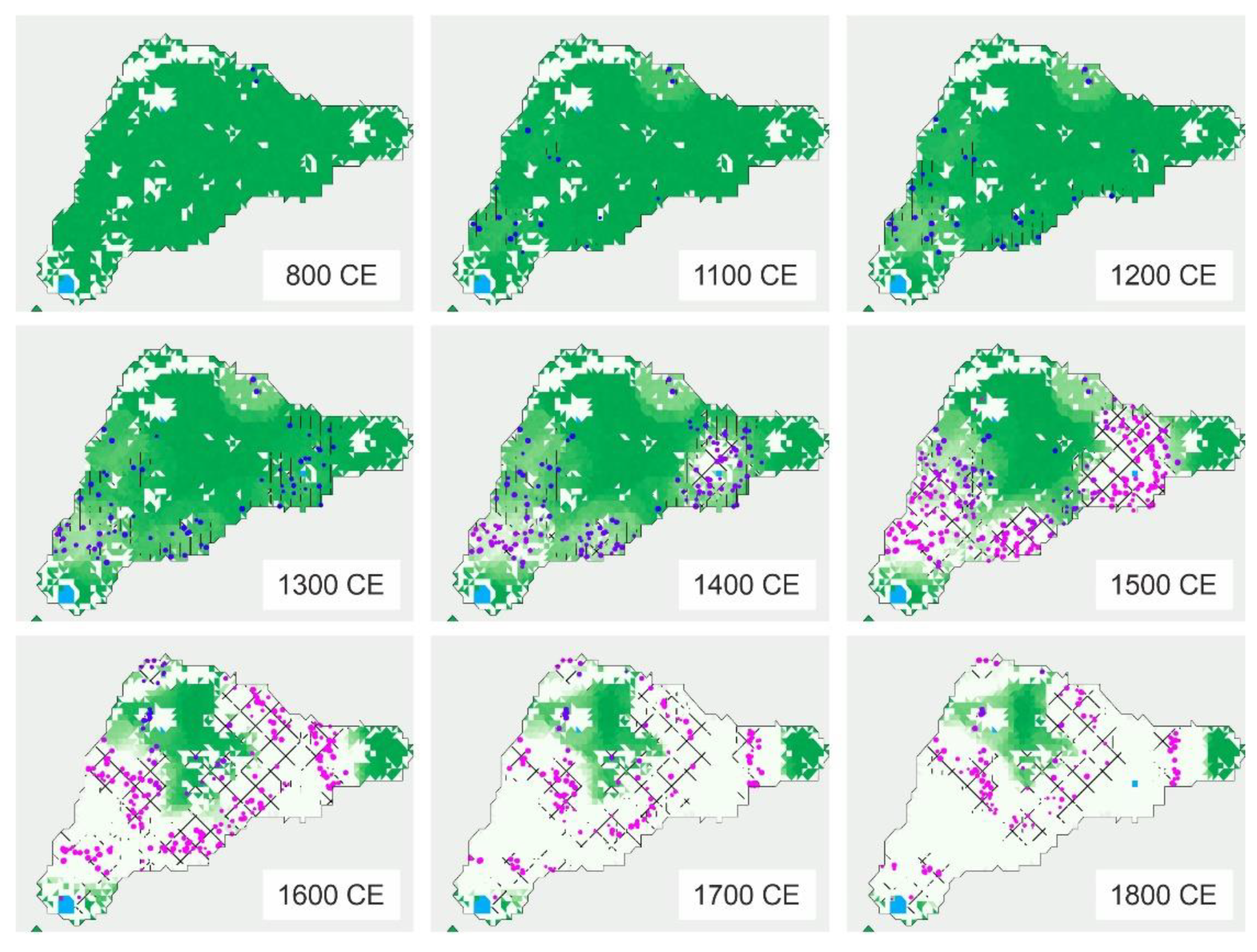
4.1. Rapa Nui
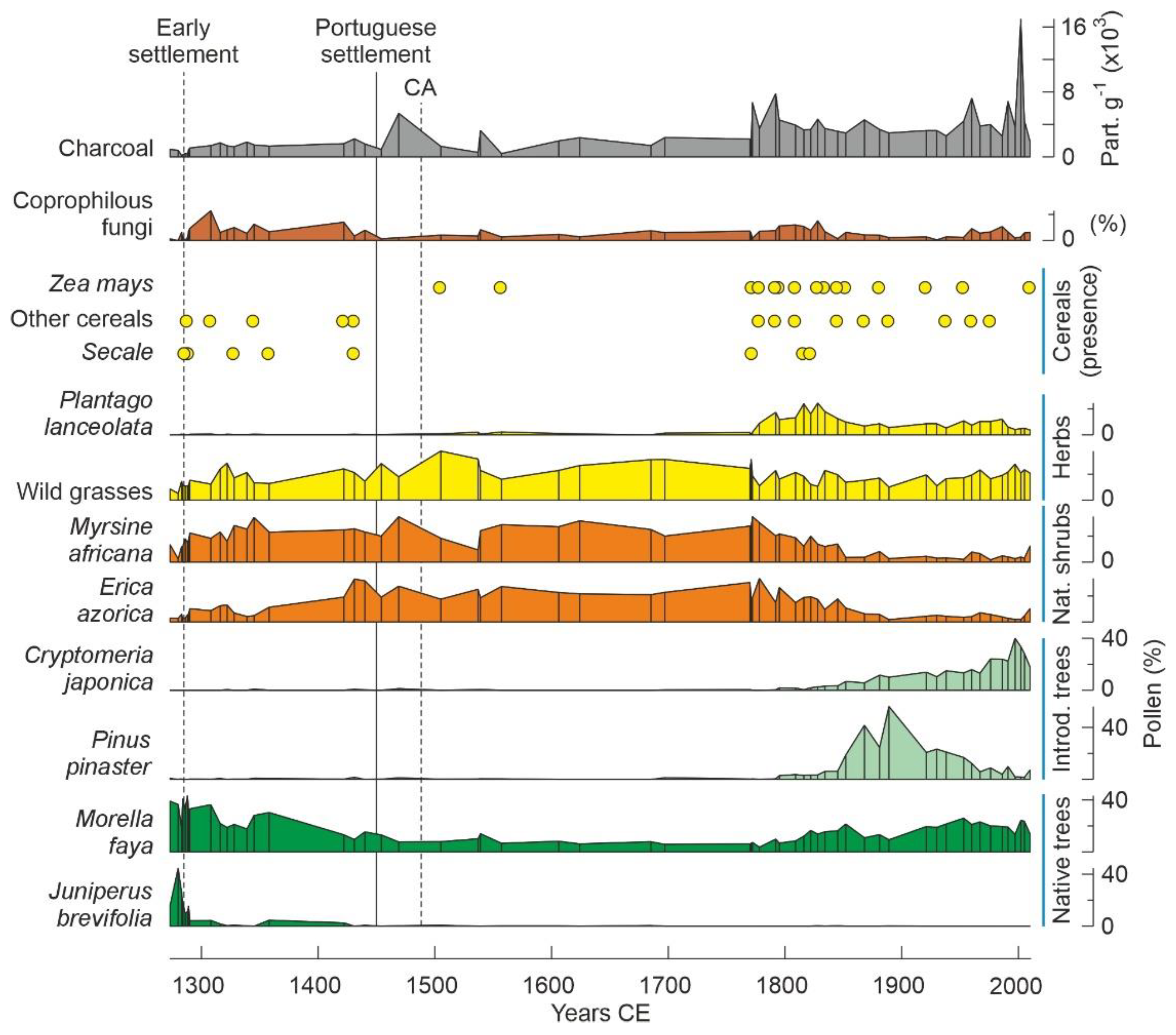
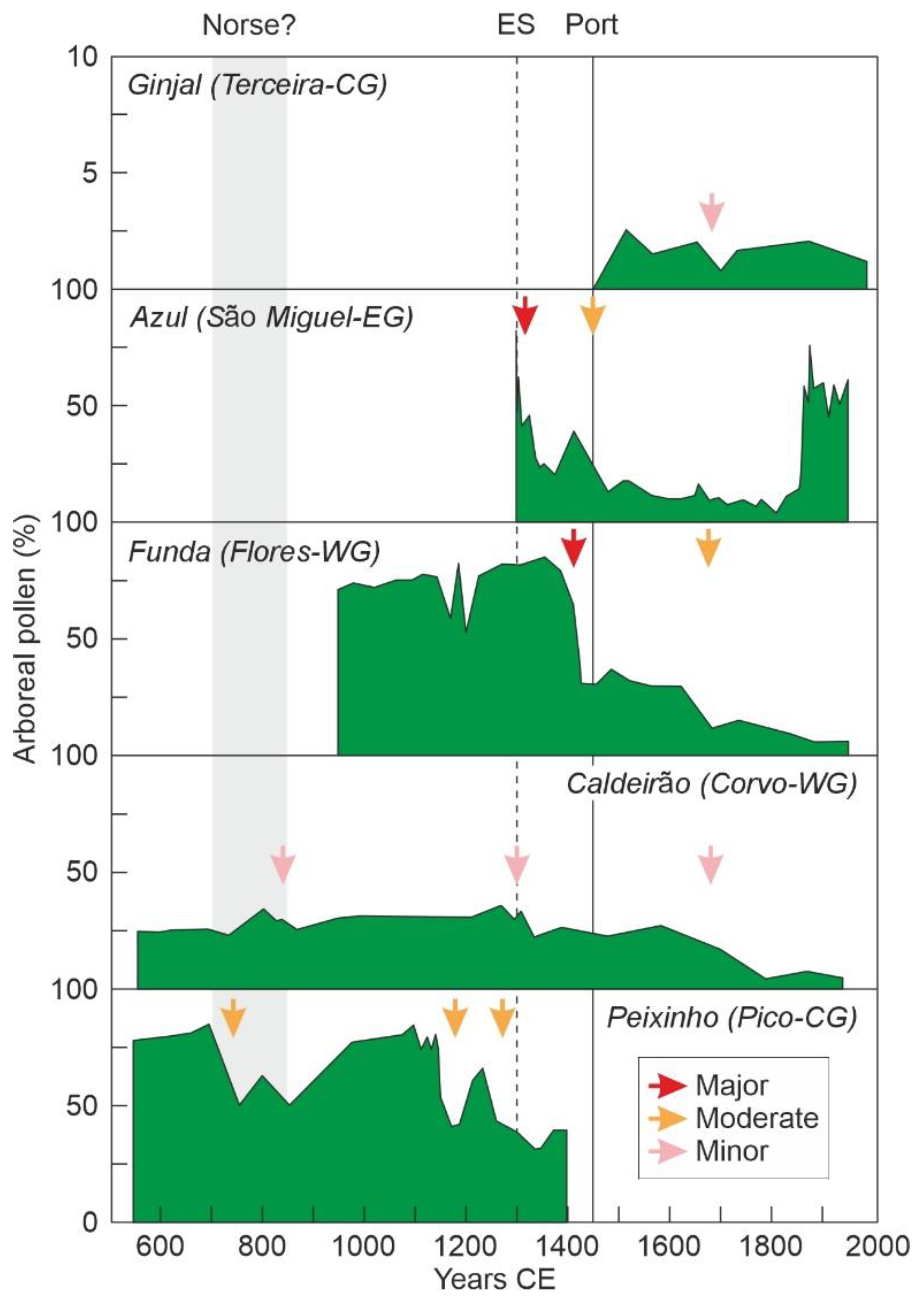
4.2. Azores
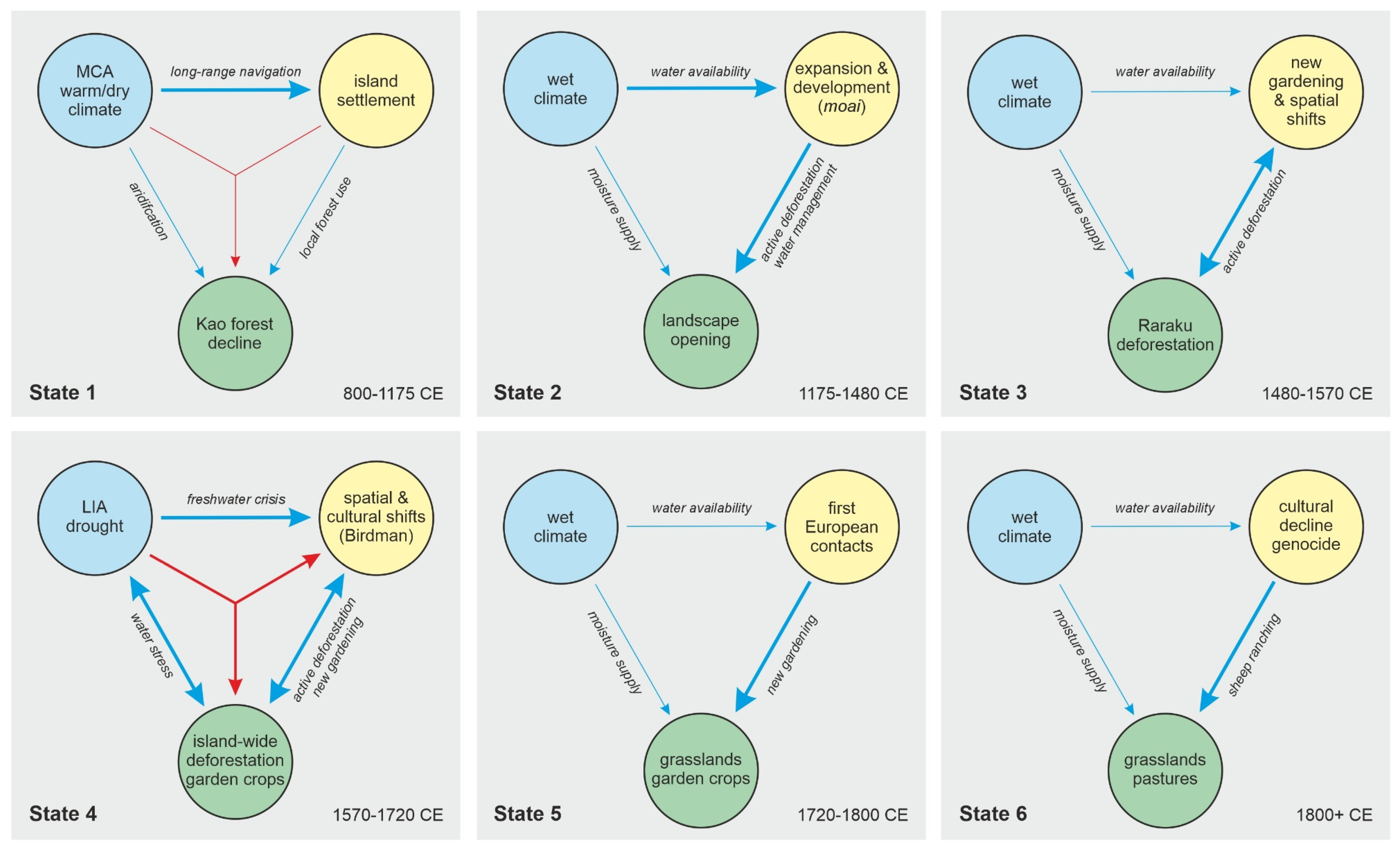
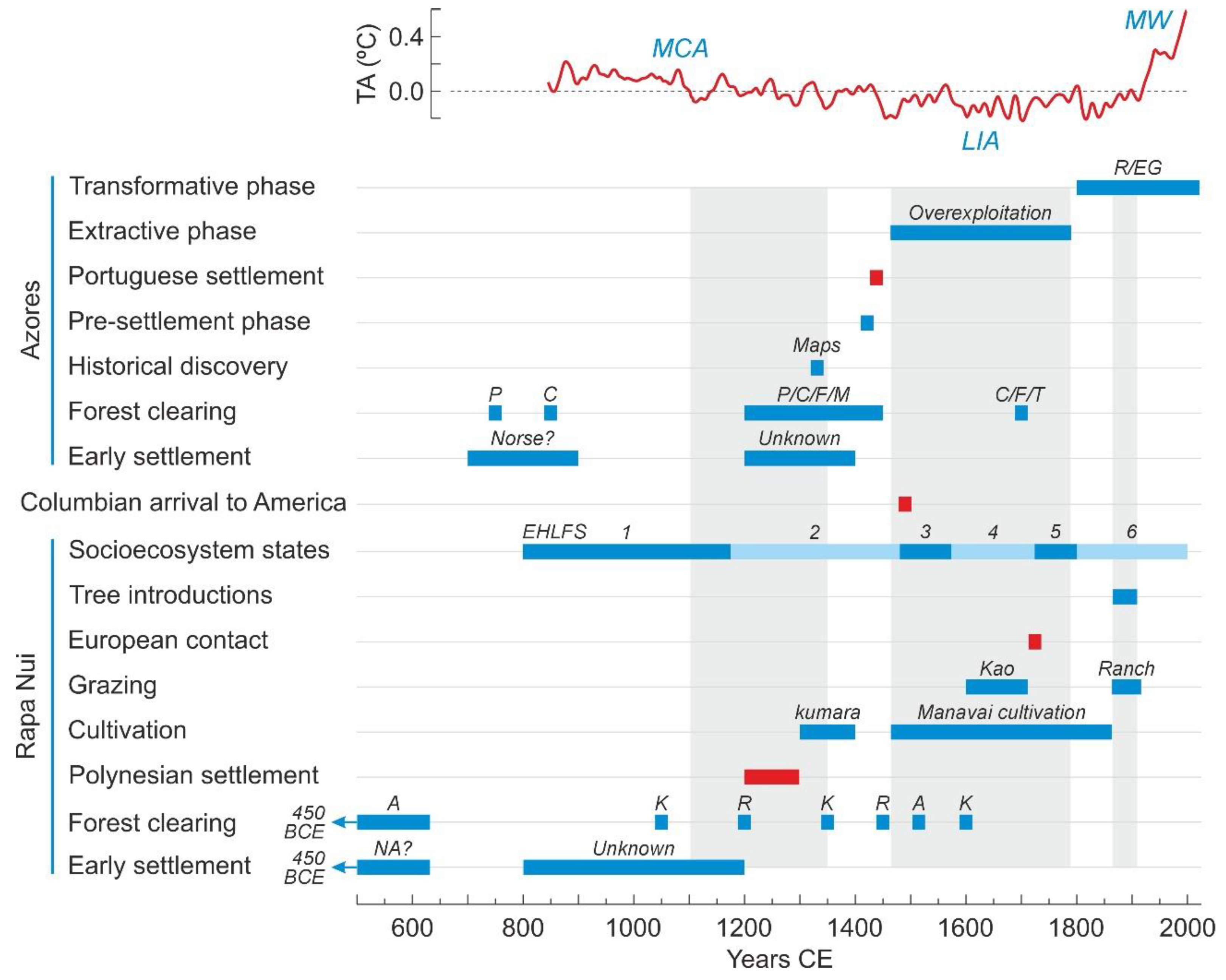
5. Potential Influence of Climatic Changes
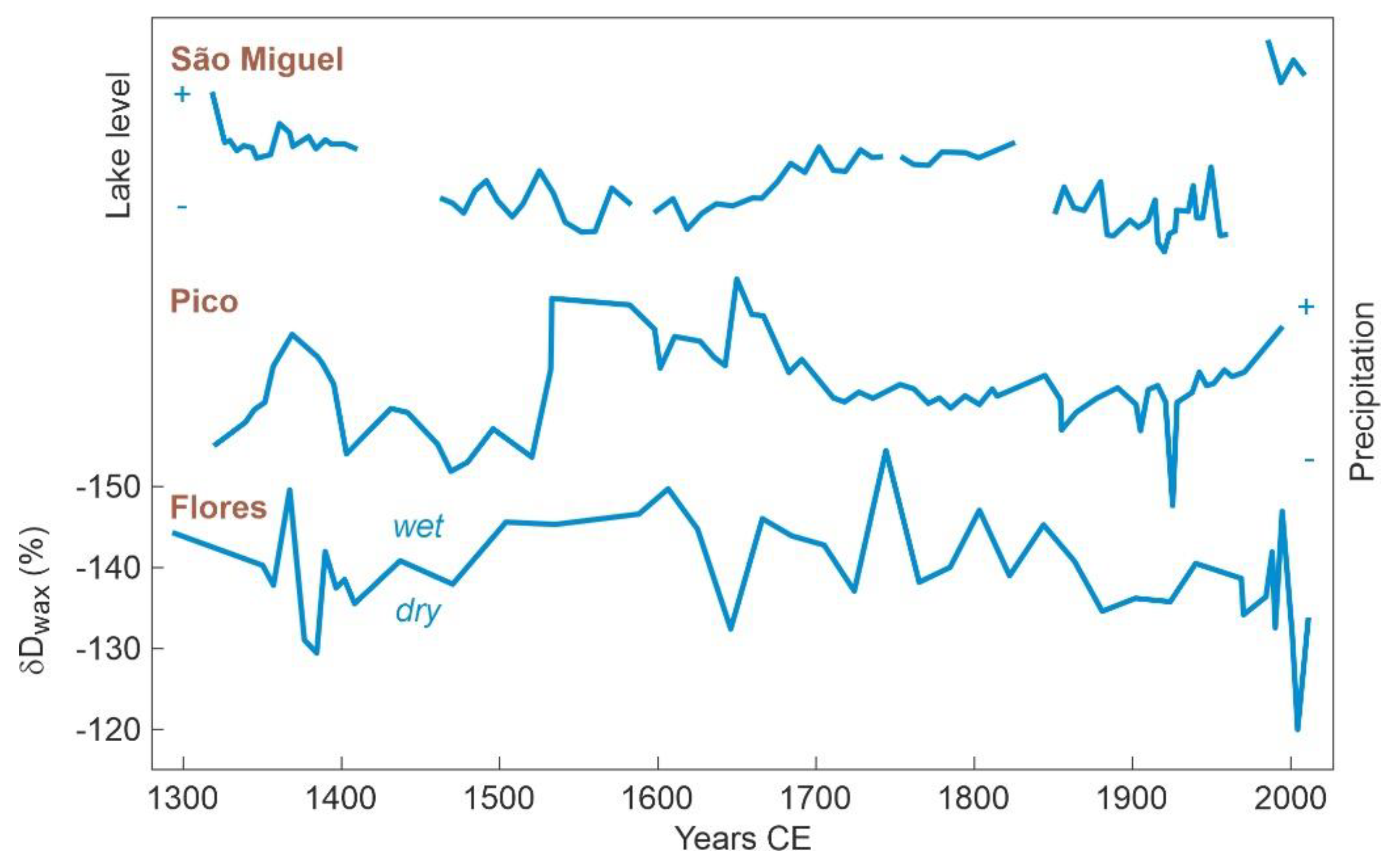
5.1. Rapa Nui
5.2. Azores
6. Comparisons between Rapa Nui and the Azores
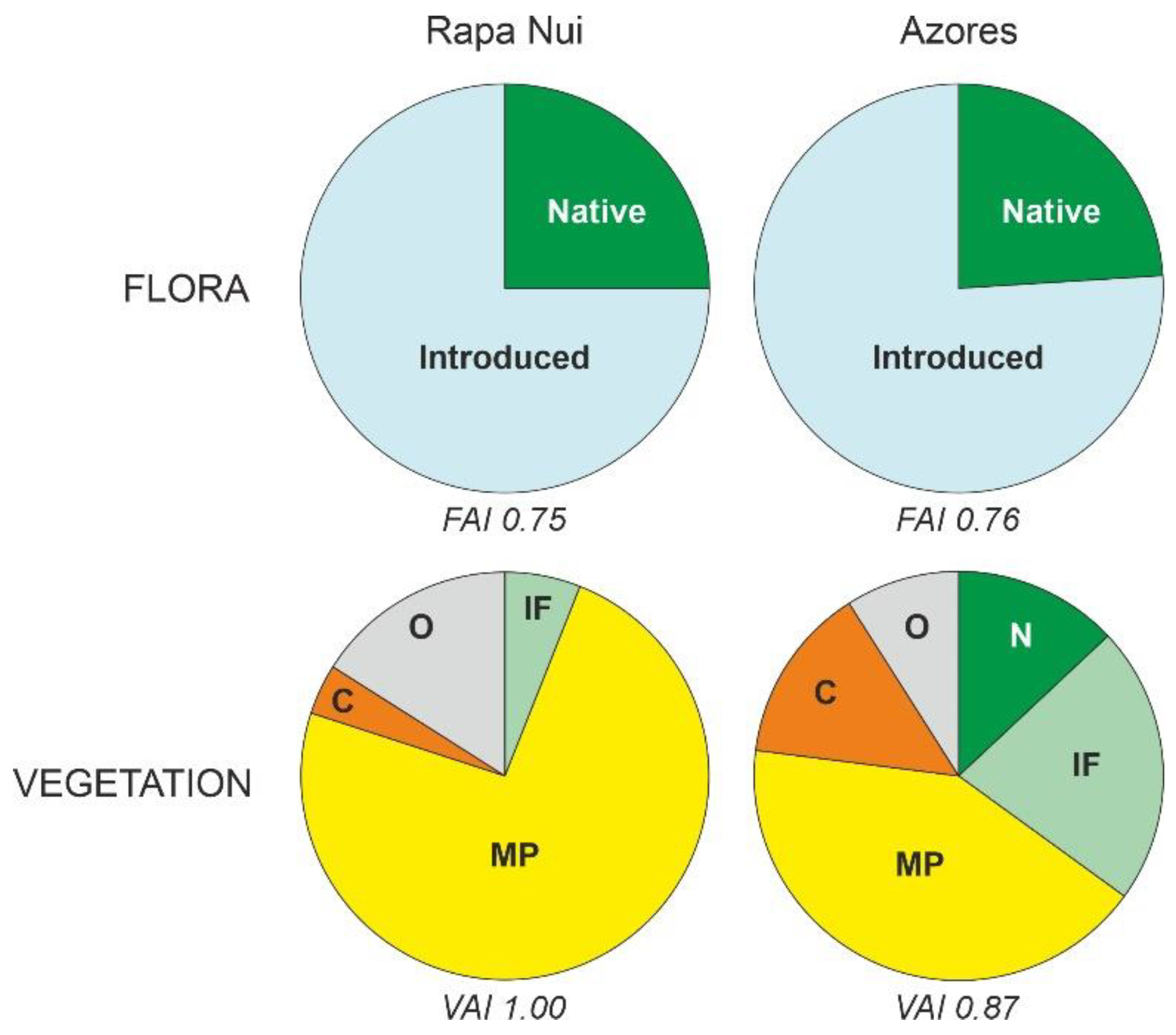
6.1. Present Conditions
6.2. Early Settlement
6.3. Permanent Settlement
6.4. Deforestation
6.5. Landscape Transformation
7. Conclusions
8. Prospects for Future Research
Funding
Acknowledgments
Conflicts of Interest
References
- Nogué, S.; de Nascimento, L.; Froyd, C.A.; Wilmshurst, J.M.; de Boer, E.J.; Coffey, E.E.D.; Whittaker, R.J.; Fernández-Palacios, J.M.; Willis, K.J. Island biodiversity conservation needs paleoecology. Nat. Ecol. Evol. 2017, 1, 0181. [Google Scholar] [CrossRef] [PubMed]
- Nogué, S.; Santos, A.M.C.; Birks, H.J.B.; Björck, S.; Castilla-Beltrán, A.; Connor, S.; de Boer, E.J.; de Nascimento, L.; Felde, V.A.; Fernández-Palacios, J.M.; et al. The human dimension of biodiversity changes on islands. Science 2021, 372, 488–491. [Google Scholar] [CrossRef] [PubMed]
- Rull, V. Pristinity, degradation and replacement: The three dimensions of human impact on island vegetation. Progr. Phys. Geogr. 2021, 45, 407–426. [Google Scholar] [CrossRef]
- Rull, V. Biodiversity crisis or sixth mass extinction? EMBO Rep. 2021, 23, e54193. [Google Scholar] [CrossRef]
- Steadman, D.W. Prehistoric extinctions of Pacific island birds: Biodiversity meets zooarchaeology. Science 1995, 267, 1123–1131. [Google Scholar] [CrossRef]
- Flenley, J.R. A palynologist looks at the colonization of the Pacific. In The Gotland Papers. Selected Papers from the VII International Conference on Easter Island and the Pacific. Migration, Identity, and Cultural Heritage; Wallin, P., Martinson-Wallin, H., Eds.; Gotland University Press: Uppsala, Sweden, 2007; pp. 15–34. [Google Scholar]
- Whittaker, R.J.; Fernández-Palacios, J.M. Island Biogeography. Ecology, Evolution and Conservation; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Prebble, M.; Dowe, J.L. The late Quaternary decline and extinction of palms on oceanic Pacific Islands. Quat. Sci. Rev. 2008, 27, 2546–2567. [Google Scholar] [CrossRef]
- Prebble, M.; Wilmshurst, J.M. Detecting the initial impact of humans and introduced species on island environments in Remote Oceania using palynology. Biol. Invasions 2009, 11, 1529–1556. [Google Scholar] [CrossRef]
- Gillespie, R.G.; Clague, D.A. Encyclopedia of Islands; University of California Press: Berkeley, CA, USA, 2009. [Google Scholar]
- Hay, J.E.; Mimura, N. Vulnerability, risk, and adaptation assessment methods in the Pacific Islands region: Past approaches, and considerations for the future. Sustain. Sci. 2013, 8, 391–405. [Google Scholar] [CrossRef]
- Newman, R.J.S.; Capitani, C.; Courtney-Mustaphi, C.; Thorn, J.P.R.; Kariuki, R.; Enns, C.; Marchant, R. Integrating insights from social-ecological interactions into sustainable land use change scenarios for small islands in the western Indian Ocean. Sustainability 2020, 12, 1340. [Google Scholar] [CrossRef]
- Rick, T.; Reeder-Meters, L.; Braje, T.J.; Wake, T. Human ecology, paleogeography, and biodiversity on California’s small islands. J. Isl. Coast. Archaeol. 2020, 113, 14536–14543. [Google Scholar] [CrossRef]
- Rull, V.; Lara, A.; Rubio-Inglés, M.J.; Giralt, S.; Gonçalves, V.; Raposeiro, P.; Hernández, A.; Sánches-López, G.; Vázquez-Loureiro, D.; Roberto, B.; et al. Vegetation and landscape dynamics under natural and anthropogenic forcing on the Azores islands: A 700-year pollen record from the So Miguel Island. Quat. Sci. Rev. 2017, 159, 155–168. [Google Scholar] [CrossRef]
- Vezzoli, L.; Acocella, V. Easter Island, SE Pacific: And end-member type of hotspot volcanism. Geol. Soc. Am. Bull. 2009, 121, 869–886. [Google Scholar] [CrossRef]
- Hildenbrand, A.; Weis, D.; Madureira, P.; Marques, F.O. Recent plate reorganization at the Azores triple junction: Evidence from combined geochemical and geochronological data on Faial S Jorge and Terceira volcanic islands. Lithos 2014, 210–211, 27–39. [Google Scholar] [CrossRef]
- Ramalho, R.S.; Helffrich, G.; Madeira, J.; Costa, M.; Thomas, C.; Quartau, R.; Hipólito, A.; Rovere, A.; Hearthy, P.J.; Ávila, S.P. Emergence and evolution of Santa Maria Island (Azores)–The conundrum of uplifted islands revisited. Geol. Soc. Am. Bull. 2017, 129, 372–390. [Google Scholar] [CrossRef]
- Sibrant, A.L.R.; Hildenbrant, A.; Marques, F.O.; Weiss, B.; Boulesteix, T.; Hübscher, C.; Lüdmann, T.; Costa, A.C.G.; Catalão, J.C. Morpho-structural evolution of a volcanic island developed inside an active oceanic rift: S Miguel Island (Terceira rift Azores). J. Volcanol. Geotherm. Res. 2015, 301, 90–106. [Google Scholar] [CrossRef]
- McLaughlin, S. The Complete Guide to Easter Island; Easter Island Foundation: Los Osos, CA, USA, 2007. [Google Scholar]
- Rull, V. Paleoecological Research on Easter Island; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Connor, S.E.; van Leeuwen, J.F.N.; Rittenour, T.M.; van der Knapp, O.; Ammann, B.; Björk, S. The ecological impact of oceanic island colonization—A palaeoecological perspective from the Azores. J. Biogeogr. 2012, 39, 1007–1023. [Google Scholar] [CrossRef]
- Connor, S.E.; van der Knapp, O.; van Leeuwen, J.F.N.; Kuněs, P. Holocene palaeoclimate and palaeovegetational on the islands of Flores and Pico. In Climate Change Perspectives from the Atlantic: Past, Present and Future; Fernández-Palacios, J.M., de Nascimento, L., Hernández, J.C., Clemente, S., González, A., Díaz-González, J.P., Eds.; Universidad de La Laguna: La Laguna, Spain, 2013; pp. 149–162. [Google Scholar]
- Raposeiro, P.M.; Hernández, A.; Pla-Rabes, S.; Gonçalves, V.; Bao, R.; Sáez, A.; Shanahan, T.; Benavente, M.; de Boer, E.J.; Richter, N.; et al. Climate change facilitated the early colonization of the Azores Archipelago during medieval times. Proc. Natl. Acad. Sci. USA 2021, 118, e2108236118. [Google Scholar] [CrossRef]
- Herrera, C.; Custodio, E. Conceptual hydrogeological model of volcanic Easter Island (Chile) after chemical and isotopic surveys. Hydrogeol. J. 2008, 16, 1329–1348. [Google Scholar] [CrossRef]
- Puleston, C.O.; Ladefoged, T.N.; Haoa, S.; Chadwick, O.A.; Vitousek, P.M.; Stevenson, C.M. Rain, sun, soil and sweat: A consideration of population limits on Rapa Nui (Easter Island) before European contact. Front. Ecol. Evol. 2017, 5, 69. [Google Scholar] [CrossRef]
- Garreaud, R.; Aceituno, P. Interannual rainfall variability over the South American Altiplano. J. Clim. 2001, 14, 2779–2789. [Google Scholar] [CrossRef]
- Bradley, R.S.; D’Andrea, W.J.; Diaz, H.F.; Ning, L. Climatology of Rapa Nui (Easter Island, Isla de Pascua). In The Prehistory of Rapa Nui (Easter Island); Rull, V., Stevenson, C.M., Eds.; Springer Nature: Cham, Switzerland, 2023; pp. 259–274. [Google Scholar]
- Delcroix, T.; Michel, S.L.L.; Swingedouw, D.; Malaizé, B.; Daniau, A.-L.; Abarca-del-Rio, R.; Caley, T.; Sémah, A.-M. Clarifying the role of ENSO on Easter Island precipitation changes: Potential environmental implications for the last millennium. Paleoceanogr. Paleoclimatol. 2022, 37, e2022PA004514. [Google Scholar] [CrossRef]
- Steiger, N.J.; D’Andrea, W.J.; Smerdon, J.E.; Bradley, R.S. Large infrequent rain events dominate the hydroclimate of Rapa Nui (Easter Island). Clim. Dyn. 2022, 59, 595–608. [Google Scholar] [CrossRef]
- Zizka, G.; Zizka, A. Te flora and vegetation of Easter Island: Past and present. In The Prehistory of Rapa Nui (Easter Island); Rull, V., Stevenson, C.M., Eds.; Springer Nature: Cham, Switzerland, 2023; pp. 347–376. [Google Scholar]
- CIREN. Mapa de Vegetación de la Isla de Pascua; Ministerio de Agricultura: Santiago, Chile, 2013.
- Borges, P.A.W.; Santos, A.M.C.; Elias, R.B.; Gabriel, R. The Azores Archipelago: Biodiversity erosion and conservation. In Encyclopedia of the World’s Biomes; Elsevier: Amsterdam, The Netherlands, 2020; pp. 101–113. [Google Scholar]
- Volkov, D.L.; Fu, L.-L. On the reasons for the formation and variability of the Azores current. J. Phys. Oceanogr. 2010, 40, 2197–2220. [Google Scholar] [CrossRef]
- Yamamoto, A.; Palter, J.B. The absence of an Atlantic imprint on the multidecadal variability of wintertime European temperature. Nat. Commun. 2016, 7, 10930. [Google Scholar] [CrossRef]
- Cropper, T.; Hanna, E.; Valente, M.A.; Jónsson, T. A daily Azores–Iceland North Atlantic Oscillation index back to 1850. Geosci. Data J. 2015, 2, 12–24. [Google Scholar] [CrossRef]
- Schaeffer, H. Chorology and diversity of the Azorean flora. Diss. Bot. 2003, 374, 1–130. [Google Scholar]
- Carta de Ocupação do Solo da Região Autónoma dos Açores; Direcção Regional do Ordenamento do Território e dos Recursos Hídricos: Ponta Delgada, Portugal, 2007.
- Frutuoso, G. Saudades da Terra; Instituto de Cultura: Ponta Delgada, Portugal, 1589; reprinted 1998. [Google Scholar]
- Moreira, J.M. Alguns aspetos de Intervenção humana na Evolução da Paisagem da Ilha de São Miguel (Açores); Serviço Nacional de Parques, Reservas e Conservação da Naturaleza: Lisboa, Portugal, 1968. [Google Scholar]
- Hunt, T.L.; Lipo, C.P. Late colonization of Easter Island. Science 2006, 311, 1603–1606. [Google Scholar] [CrossRef]
- Wilmshurst, J.M.; Hunt, T.L.; Lipo, C.P.; Anderson, A.J. High-precision radiocarbon dating shows recent and rapid initial human colonization of East Polynesia. Proc. Natl. Acad. Sci. USA 2011, 108, 1815–1820. [Google Scholar] [CrossRef]
- Boersema, J.J. The Survival of Easter Island. Dwilling Resources and Cultural Resilience; Cambridge University Press: Cambridge, UK, 2015. [Google Scholar]
- Fischer, S.R. Island at the End of the World. The Turbulent History of Easter Island; Reaktion Books: London, UK, 2005. [Google Scholar]
- Flenley, J.R.; King, S. Late Quaternary pollen records from Easter Island. Nature 1984, 307, 47–50. [Google Scholar] [CrossRef]
- Flenley, J.R.; King, A.S.M.; Jackson, J.; Chew, C.; Teller, J.T.; Prentice, M.E. The Late Quaternary vegetation and climatic history of Easter Island. J. Quat. Sci. 1991, 6, 85–115. [Google Scholar] [CrossRef]
- Butler, K.; Prior, C.A.; Flenley, J.R. Anomalous radiocarbon dates from Easter Island. Radiocarbon 2004, 46, 395–420. [Google Scholar] [CrossRef]
- Mann, D.; Chase, J.; Edwards, J.; Beck, W.; Reanier, R.; Mass, M.; Finney, B.; Loret, J. Prehistoric destruction of the primeval soils and vegetation of Rapa Nui (Isla de Pascua, Easter Island). In Easter Island. Scientific Exploration into the World’s Environmental Problems in Microcosm; Loret, J., Tanacredi, J.T., Eds.; Kluwer Academic/Plenum: New York, NY, USA, 2003; pp. 133–153. [Google Scholar]
- Mieth, A.; Bork, H.R. Humans, climate or introduced rats–Which is to blame for the woodland destruction on prehistoric Rapanui (Easter Island)? J. Archaeol. Sci. 2010, 37, 417–426. [Google Scholar] [CrossRef]
- Rull, V. Contributions of paleoecology to Easter Island’s prehistory: A thorough review. Quat. Sci. Rev. 2021, 252, 106751. [Google Scholar] [CrossRef]
- Rull, V. Prehistoric paleoecology of Easter Island. In The Prehistory of Rapa Nui (Easter Island); Rull, V., Stevenson, C.M., Eds.; Springer Nature: Cham, Switzerland, 2023; pp. 275–309. [Google Scholar]
- Cañellas-Boltà, N.; Rull, V.; Sáez, A.; Margalef, O.; Bao, R.; Pla-Rabes, S.; Blaauw, M.; Valero-garcés, B.; Giralt, S. Vegetation changes and human settlement of Easter Island during the last millennia: A multiproxy study of the Lake Raraku sediments. Quat. Sci. Rev. 2013, 72, 36–48. [Google Scholar] [CrossRef]
- Butler, K.R.; Flenley, J.R. The Rano Kau 2 pollen diagram: Palaeoecology revealed. Rapa Nui J. 2010, 24, 5–10. [Google Scholar]
- Horrocks, M.; Baisden, W.T.; Flenley, J.; Feek, D.; González-Nualart, L.; Haoa-Cardinali, S.; Edmunds Gorman, T. Fossil plant remains at Rano Raraku, Easter Island’s statue quarry: Evidence for past elevated lake level and ancient Polynesian agriculture. J. Paleolimnol. 2012, 46, 767–783. [Google Scholar] [CrossRef]
- McCall, G. Easter Island. In Encyclopedia of Islands; Gillespie, R.G., Clague, D.A., Eds.; University of California Press: Berkeley, CA, USA, 2009; pp. 244–251. [Google Scholar]
- Piperno, D.R.; Holst, I. The presence of starch grains on prehistoric stone tools from the humid Neotropics: Indications of early tuber use and agriculture in Panama. J. Archaeol. Sci. 1998, 25, 765–776. [Google Scholar] [CrossRef]
- Hather, J.; Kirch, P.V. Prehistoric sweet potato (Ipomoea batatas) from Mangaia Island, central Polynesia. Antiquity 1991, 65, 887–893. [Google Scholar] [CrossRef]
- Orliac, C.; Orliac, M. The disappearance of Easter Island’s forest: Over-exploitation or climatic catastrophe? In Easter Island in Pacific Context; Stevenson, C.M., Lee, G., Morin, F.J., Eds.; Easter Island Foundation: Los Osos, CA, USA, 1998; pp. 129–134. [Google Scholar]
- Rull, V. Human settlement and discovery of the remote Easter Island. Quaternary 2019, 2, 15. [Google Scholar] [CrossRef]
- Anderson, A. Ex Oriente Lux: Amerindian seafaring and Easter Island contact revisited. In The Prehistory of Rapa Nui (Easter Island); Rull, V., Stevenson, C.M., Eds.; Springer Nature: Cham, Switzerland, 2023; pp. 19–40. [Google Scholar]
- Muñoz-Rodríguez, P.; Wood, J.R.I.; Scotland, R.W. Sweet potato on Rapa Nui: Insights from a monographic study of the genus Ipomoea. In The Prehistory of Rapa Nui (Easter Island); Rull, V., Stevenson, C.M., Eds.; Springer Nature: Cham, Switzerland, 2023; pp. 63–83. [Google Scholar]
- Temmen, J.; Montenegro, A.; Juras, S.; Field, J.S.; DeGrand, J. Floating the sweet potato to polynesia: Considering the feasibility of oceanic drift for the prehistoric introduction of the sweet potato (Ipomoea batatas) to Pacific Islands. Quat. Sci. Rev. 2022, 295, 107709. [Google Scholar] [CrossRef]
- Thompson, V.A.; Herrera, M.; Austin, J.I. Commensals/domesticates on Rapa Nui: What can their phylogeographic patterns tell us about the discovery and settlement of the island? In The Prehistory of Rapa Nui (Easter Island); Rull, V., Stevenson, C.M., Eds.; Springer Nature: Cham, Switzerland, 2023; pp. 41–62. [Google Scholar]
- Van Tilburg, J.A.; Huebert, J.M.; Sherwood, S.C.; Barrier, C.R. Pre-European contact sweet potato (Ipomoea batatas) at Rapa Nui: Macrobotanical evidence from recent excavations in Rano Raraku quarry, Rapa Nui. In The Prehistory of Rapa Nui (Easter Island); Rull, V., Stevenson, C.M., Eds.; Springer Nature: Cham, Switzerland, 2023; pp. 85–108. [Google Scholar]
- Jones, T.L.; Storey, A.A.; Matisoo-Smith, E.; Ramírez-Aliaga, J.M. Polynesians in America. Pre-Columbian Contacts with the New World; Altamira Press: Lanham, MD, USA, 2011. [Google Scholar]
- Heyerdahl, T. American Indians in the Pacific: The Theory behind the Kon-Tiki Expedition; Allen & Unwin: London, UK, 1952. [Google Scholar]
- Flenley, J.R.; Bahn, P.G. The Enigmas of Easter Island; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Ioannidis, A.G.; Blanco-Portillo, J.; Sandoval, K.; Hagelberg, E.; Miquel-Poblete, J.F.; Moreno-Mayar, J.V.; Rodríguez-Rodríguez, J.E.; Quinto-Cortés, C.D.; Auckland, K.; Parks, T.; et al. Native American gene flow into Polynesia predating Easter Island settlement. Nature 2020, 583, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Thorsby, E. Genetic evidence of a contribution of Native Americans to the early settlement of Rapa Nui (Easter Island). Front. Ecol. Evol. 2016, 4, 118. [Google Scholar] [CrossRef]
- Stevenson, C.M.; Ramírez, J.M.; Haoa, S.; Allen, T. Archaeological investigations at ‘Anakena beach and other near-coastal locations. In Easter island Arcaheology: Research on Early Rapanui Culture; Stevenson, C.M., Ayres, W.S., Eds.; Easter Island Foundation: Los Osos, CA, USA, 2000; pp. 147–172. [Google Scholar]
- Flenley, J.R.; Bahn, P.G. Hunt, terry and carl Lipo. The statues that walked. Unravelling the mystery of Easter Island. Rapa Nui J. 2011, 25, 60–62. [Google Scholar]
- Elias, R.B.; Connor, S.E.; Góis-Marques, C.A.; Schaefer, H.; Silva, L.; Sequeira, M.M.; Moura, M.; Borges, P.A.V.; Gabriel, R. Is there solid evidence of widespread landscape disturbance in the Azores before the arrival of the Portuguese? Proc. Natl. Acad. Sci. USA 2022, 119, e2119218119. [Google Scholar] [CrossRef] [PubMed]
- Raposeiro, P.M.; Hernández, A.; Pla-Rabes, S.; Gonçalves, V.; Bao, R.; Sáez, A.; Shanahan, T.; Benavente, M.; de Boer, E.J.; Richter, N.; et al. Multiproxy evidence of widespread landscape disturbance in multiple Azorean lakes before the Portuguese arrival. Proc. Natl. Acad. Sci. USA 2022, 119, e2120107119. [Google Scholar] [CrossRef] [PubMed]
- Rull, V. Human settlement and anthropization of the Azores Islands. EarthArXiv 2023. [Google Scholar] [CrossRef]
- Dransfield, J.; Flenley, J.R.; King, S.M.; Harkness, D.D.; Rapu, S. A recently extinct palm from Easter Island. Nature 1984, 312, 750–752. [Google Scholar] [CrossRef]
- Zizka, G. Flowering Plants of Easter Island; Hortus Fracofurtensis: Frankfurt Main, Germany, 1991. [Google Scholar]
- Mieth, A.; Bork, H.R. Degradation of resources and successful land-use management of prehistoric Rapanui: Two sides of the same coin. In Easter Island: Collapse or Transformation? A State of the Art; Clauwe, N., De Drapper, M., Eds.; Royal Academy of Overseas Sciences: Brussels, Belgium, 2015; pp. 91–112. [Google Scholar]
- Mieth, A.; Bork, H.R. A vanished landscape–Phenomena and eco-cultural consequences of extensive deforestation in the prehistory of Rapa Nui. In Cultural and Environmental Change on Rapanui; Haoa Cardinali, S., Ingersoll, K.B., Ingersoll, D.W., Stevenson, C.M., Eds.; Routledge: London, UK, 2017; pp. 32–58. [Google Scholar]
- Rull, V. The deforestation of Easter Island. Biol. Rev. 2020, 95, 124–141. [Google Scholar] [CrossRef]
- Horrocks, M.; Baisden, W.T.; Harper, M.A.; Marra, M.; Flenley, J.; Feek, D.; Haoa-Cardinali, S.; Keller, E.D.; González Nualart, L.; Edmunds Gorman, T. A plant microfossil record of Late Quaternary environments and human activity from Rano Aroi and surroundings, Easter Island. J. Paleolimnol. 2015, 54, 279–303. [Google Scholar] [CrossRef]
- Seco, I.; Rull, V.; Montoya, E.; Cañellas-Boltà, N.; Giralt, S.; Margalef, O.; Pla-Rabes, S.; D’Andrea, W.J.; Bradley, R.S.; Sáez, A. A continuous palynological record of forest clearing at Rano Kao (Easter Island, SE Pacific) during the last millennium: Preliminary report. Quaternary 2019, 2, 22. [Google Scholar] [CrossRef]
- Rull, V.; Cañellas-Boltà, N.; Margalef, O.; Sáez, A.; Pla-Rabes, S.; Giralt, S. Late Holocene vegetation dynamics and deforestation in Rano Aroi: Implications for Easter Island’s ecological and cultural history. Quat. Sci. Rev. 2015, 126, 219–226. [Google Scholar] [CrossRef]
- Mieth, A.; Bork, H.R. History, origin and extent of soil erosion on Easter Island (Rapa Nui). Catena 2005, 63, 244–260. [Google Scholar] [CrossRef]
- Stevenson, C.M.; Wozniak, J.A.; Haoa, S. Prehistoric agricultural production on Easter Island (Rapa Nui). Antiquity 1999, 73, 801–812. [Google Scholar] [CrossRef]
- Wozniak, J.A. Prehistoric horticultural practices on Easter Island: Lithic mulched gardens and field streams. Rapa Nui J. 1999, 13, 95–99. [Google Scholar]
- Bork, H.R.; Mieth, A.; Tschochner, B. Nothing but stones? Review on the extent and technical efforts of prehistoric stone mulching on Rapa Nui. Rapa Nui J. 2004, 18, 10–14. [Google Scholar]
- Steiglechner, P.; Merico, A. Spatio-temporal patterns of deforestation, settlement, and land use on Easter Island prior to European arrivals. In The Prehistory of Rapa Nui (Easter Island); Rull, V., Stevenson, C.M., Eds.; Springer Nature: Cham, Switzerland, 2023; pp. 401–426. [Google Scholar]
- Horrocks, M.; Wozniak, J.A. Plant microfossil analysis reveals disturbed forest and a mixed-crop, dryland production system at Te Niu, Easter Island. J. Archaeol. Sci. 2008, 35, 126–142. [Google Scholar] [CrossRef]
- Horrocks, M.; Baisden, W.T.; Nieuwoudt, M.K.; Flenley, J.; Feek, D.; González-Nualart, L.; Haoa-Cardinali, S.; Edmunds Gorman, T. Microfossils of Polynesian cultigens in lake sediment cores from Rano Kau, Easter Island. J. Paleolimnol. 2012, 47, 185–204. [Google Scholar] [CrossRef]
- Horrocks, M.; Marra, M.; Baisden, W.T.; Flenley, J.; Feek, D.; González-Nualart, L.; Haoa-Cardinali, S.; Edmunds Gorman, T. Pollen, phytoliths, arthropods and high-resolution 14C sampling from Rano Kau, Easter Island: Evidence for Late Quaternary environments, ant (formicidae) distributions and human activity. J. Paleolimnol. 2013, 50, 417–432. [Google Scholar] [CrossRef]
- Horrocks, M.; Baisden, W.T.; Flenley, J.R.; Feek, D.; Love, C.; Haoa-Cardinali, S.; González-Nualart, L.; Edmunds Gorman, T. Pollen, phytolith and starch analysis of dryland soils from Easter Island (Rapa Nui) show widespread vegetation clearance and Polynesian-induced crops. Palynology 2016, 41, 339–350. [Google Scholar] [CrossRef]
- Dias, E. A chegada dos portugueses às ilhas–O antes e o depois. In Açores e Madeira: A Floresta das Ilhas; Silva, L., Ed.; Fundação Luso Americana: Lisboa, Portugal, 2007; pp. 137–295. [Google Scholar]
- Schaeffer, H.; Moura, M.; Graciete, M.; Silva, L.; Rumsey, F.J.; Carine, M.A. The Linnean shortfall in oceanic biogeography: A case study in the Azores. J. Biogeogr. 2011, 38, 1345–1355. [Google Scholar] [CrossRef]
- Silva, L.; Smith, C.W. A characterization of the non-indigenous flora of the Azores Archipelago. Biol. Invasions 2004, 6, 193–204. [Google Scholar] [CrossRef]
- Rull, V. Quaternary Ecology, Evolution and Biogeography; Academic Press: London, UK, 2020. [Google Scholar]
- Nunn, P.D. Climate, Environment and Society in the Pacific during the Last Millennium; Elsevier: Amstredam, The Netherlands, 2007. [Google Scholar]
- Godwin, I.D.; Browning, S.A.; Anderson, A. Climate windows for Polynesian voyaging to New Zealand and Easter Island. Proc. Natl. Acad. Sci. USA 2014, 111, 14716–14721. [Google Scholar] [CrossRef] [PubMed]
- Sear, D.A.; Allen, M.S.; Hassall, J.D.; Maloney, A.E.; Langdon, P.G.; Morrison, A.E.; Henderson, A.C.G.; Mackay, H.; Croudace, I.W.; Clarke, C.; et al. Human settlement of east Polynesia earlier, incremental, and coincident with prolonged South Pacific drought. Proc. Natl. Acad. Sci. USA 2020, 117, 8813–8819. [Google Scholar] [CrossRef] [PubMed]
- Montenegro, Á.; Callaghan, R.T.; Fitzpatrick, S.M. Using seafaring simulations and shortest-hop trajectories to model prehistoric colonization of Remote Oceania. Proc. Natl. Acad. Sci. USA 2016, 113, 12685–12690. [Google Scholar] [CrossRef]
- Margalef, O.; Álvarez-Gómes, J.A.; Pla-Rabes, S.; Cañellas-Boltà, N.; Rull, V.; Sáez, A.; Geyer, A.; Peñuelas, J.; Sardans, J.; Gitalt, S. Revisiting the role of high-energy Pacific events in the environmental and cultural history of Easter Island (Rapa Nui). Geogr. J. 2018, 184, 310–322. [Google Scholar] [CrossRef]
- Bahn, P.; Flenley, J.R. Easter Island, Earth Island; Tames & Hudson: London, UK, 1992. [Google Scholar]
- Roman, M.; McWethy, D.B.; Kehrwald, N.M.; Erheni, E.O.; Myrbo, A.E.; Ramirez-Aliaga, J.M.; Pauchard, A.; Turetta, C.; Barbante, C.; Prebble, M.; et al. A multi-decadal geochemical record from Rano Aroi (Easter Island/Rapa Nui): Implications for the environment, climate and humans during the last two millennia. Quat. Sci. Rev. 2021, 268, 107115. [Google Scholar] [CrossRef]
- Rull, V. Drought, freshwater availability and cultural resilience on Easter Island (SE Pacific) during the Little Ice Age. Holocene 2020, 30, 774–780. [Google Scholar] [CrossRef]
- Rull, V. Strong Fuzzy EHLFS: A general conceptual framework to address past records of environmental, ecological and cultural change. Quaternary 2018, 1, 10. [Google Scholar] [CrossRef]
- Rull, V.; Stevenson, C.M. (Eds.) Towards a holistic approach to Easter Island’s prehistory. In The Prehistory of Rapa Nui (Easter Island); Springer Nature: Cham, Switzerland, 2023; pp. 611–628. [Google Scholar]
- Björk, S.; Rittenour, T.; Rosén, P.; França, Z.; Möller, P.; Snowball, I.; Wastegård, S.; Bennike, O.; Kromer, B. A Holocene lacustrine record in the central North Atlantic: Proxies for volcanic activity, and long-term precipitation changes. Quat. Sci. Rev. 2006, 25, 9–32. [Google Scholar] [CrossRef]
- Hernández, A.; Sáez, A.; Bao, R.; Raposeiro, P.M.; Trigo, R.M.; Doolittle, S.; Masqué, P.; Rull, V.; Gonçalves, V.; Vázquez-Loureiro, D.; et al. The influences of the AMO and NAO on the sedimentary infill in an Azores Archipelago lake since ca. 1350 CE. Glob. Planet. Change 2017, 154, 61–74. [Google Scholar] [CrossRef]
- Richter, N.; Russell, J.M.; Amaral-Zettler, L.; DeGroff, W.; Raposeiro, P.M.; Gonçalves, V.; de Boer, E.J.; Pla-Rabes, S.; Hernández, A.; Benavente, M.; et al. Long-term hydroclimate variability in the sub-tropical North Atlantic and anthropogenic impacts on lake ecosystems: A case study from Flores Island, the Azores. Quat. Sci. Rev. 2022, 285, 107525. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. An equilibrium theory of insular biogeography. Evolution 1963, 17, 373–387. [Google Scholar] [CrossRef]
- Carine, M.A.; Schaeffer, H. The Azorean diversity enigma: Why are there so few Azorean endemic flowering plants and why they are so widespread? J. Biogeogr. 2010, 37, 77–89. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2021. The Physical Basis; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Kirch, P.V. Peopling of the Pacific: A holistic anthropological perspective. Annu. Rev. Anthropol. 2010, 39, 131–148. [Google Scholar]
- Ioannidis, A.G.; Blanco-Portillo, J.; Sandoval, K.; Hagelberg, E.; Barberena-Jonas, C.; Hill, A.V.S.; Rodríguez-Rodríguez, J.E.; Fox, K.; Robson, K.; Haoa-Cardinali, S.; et al. Paths and 478.timings of the peopling of Polynesia inferred from genomic networks. Nature 2021, 597, 522–526. [Google Scholar] [CrossRef]
- Kirch, P.V. When early Polynesians migrated estawards. Nature 2021, 597, 477–478. [Google Scholar] [CrossRef]
- Finney, B. Colonizing an island world. Trans. Am. Philos. Soc. 1996, 86, 71–116. [Google Scholar] [CrossRef]
- Kirch, P.V. On the Road of the Winds: An Archaeological History of the Pacific Islands before European Contact; University of California Press: Berkeley, CA, USA, 2017. [Google Scholar]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term | Meaning | Types | Meaning | Evidence (Examples) |
|---|---|---|---|---|
| Discovery | Finding the island for the first time | Sighting | See the island in the distance | Historical (maps, logbooks) |
| Landing | Physically arrive on the island | |||
| Settlement | Physical occupancy of the island | Ephemeral | Temporary occupancy | Historical (written documents); archaeological (housing, monuments, art); paleoecological and archaeobotanical (biomarkers, cultivated plants, fire); anthropological (DNA) |
| Intermittent | Recurrent occupancy and abandonment | |||
| Permanent | Long-lasting occupancy | |||
| Local | Restricted to one or a few sites | |||
| Full | Extensive to the whole island | |||
| Contact | Arrival of a new culture to an already settled island | Visit | Stopover with or without cultural incidence | Historical (written documents); archaeological (housing, monuments, art); anthropological (fossil skeletons, DNA, ethnography) |
| Admixture | Cultural/biological merging | |||
| Invasion | Violent occupation or slavery practices |
| Rapa Nui | Azores | ||
|---|---|---|---|
| Extent | Total surface (km2) | 164 | 2351 |
| Isolation | Nearest continent (km) | 3700 | 1400 |
| Nearest island (km) | 2100 | 900 | |
| Vascular flora | Total species (n) | 201 | 811 |
| Species/area | 1.22 | 0.34 | |
| Native (%) | 25 | 24 | |
| Introduced (%) | 75 | 76 | |
| Endemics (%) | 12 | 30 | |
| Vegetation cover | Native types (%) | 0 | 13 |
| Introduced forests (%) | 6 | 22 | |
| Meadows/pastures (%) | 74 | 42 | |
| Crops (%) | 4 | 14 | |
| Others (%) | 16 | 9 | |
| Anthropization | FAI | 0.75 | 0.76 |
| VAI | 1 | 0.87 |
| Process | Similarities | Differences |
|---|---|---|
| Early settlement | Consistent evidence (forest clearing, fire, cultivated plants/weeds) | Timing and origin: RN—Amerindian component (450 BCE) AZ—Unknown origin (13th–14th centuries) |
| Permanent settlement | Timing (1200–1450 CE) | Origin and expansion routes: RN—Ancient Polynesians (late eastward expansion) AZ—Modern Portuguese (early westward imperial expansion) |
| Forest clearing | Overexploitation and time of initiation (15th century) | Intensity: RN—Full deforestation (1600 CE) by random local pulses AZ—Partial deforestation by generalized events |
| Landscape transformation | Timing of major transformations (15th–18th centuries) and maximum anthropization (19th century) | Type of transformation: RN—expansion of gardening cultivation and extensive grazing AZ—extractive practices and further reforestation (18th century) |
| Present conditions | Floristic anthropization (75%) | Species/area relationships: RN—1.22; AZ—0.34 Endemic species (%): RN—12%; AZ—30% Vegetation anthropization: RN—1.00; AZ—0.87 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rull, V. Human Settlement and Landscape Anthropization of Remote Oceanic Islands: A Comparison between Rapa Nui (Pacific Ocean) and the Azores (Atlantic Ocean). Plants 2023, 12, 2089. https://doi.org/10.3390/plants12112089
Rull V. Human Settlement and Landscape Anthropization of Remote Oceanic Islands: A Comparison between Rapa Nui (Pacific Ocean) and the Azores (Atlantic Ocean). Plants. 2023; 12(11):2089. https://doi.org/10.3390/plants12112089
Chicago/Turabian StyleRull, Valentí. 2023. "Human Settlement and Landscape Anthropization of Remote Oceanic Islands: A Comparison between Rapa Nui (Pacific Ocean) and the Azores (Atlantic Ocean)" Plants 12, no. 11: 2089. https://doi.org/10.3390/plants12112089
APA StyleRull, V. (2023). Human Settlement and Landscape Anthropization of Remote Oceanic Islands: A Comparison between Rapa Nui (Pacific Ocean) and the Azores (Atlantic Ocean). Plants, 12(11), 2089. https://doi.org/10.3390/plants12112089