

Do Native and Alien Species Differ in Their Ecological Strategies? A Test with Woody Plants in Tropical Rainforests on Réunion Island (Mascarene Archipelago, Indian Ocean)
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
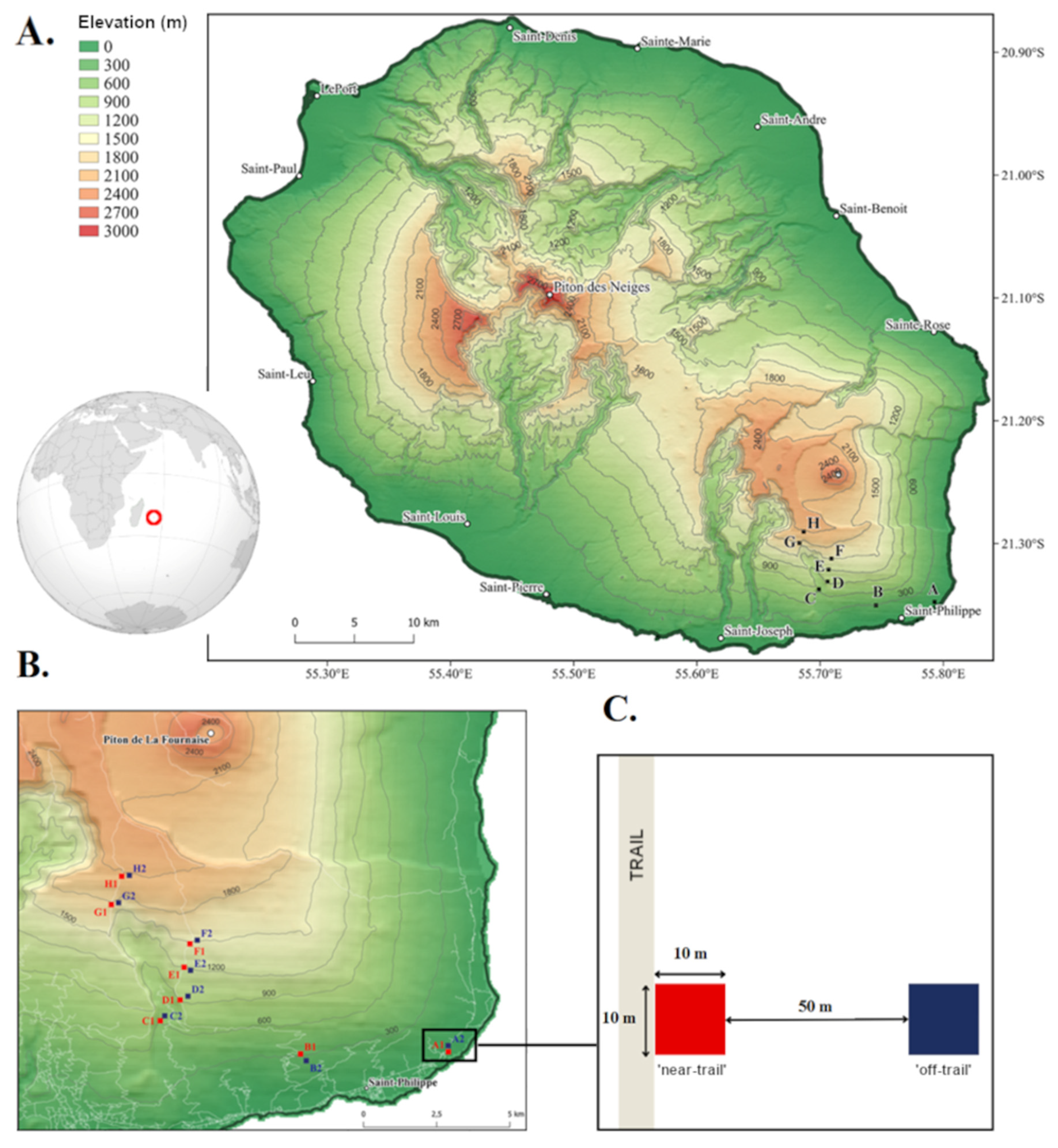
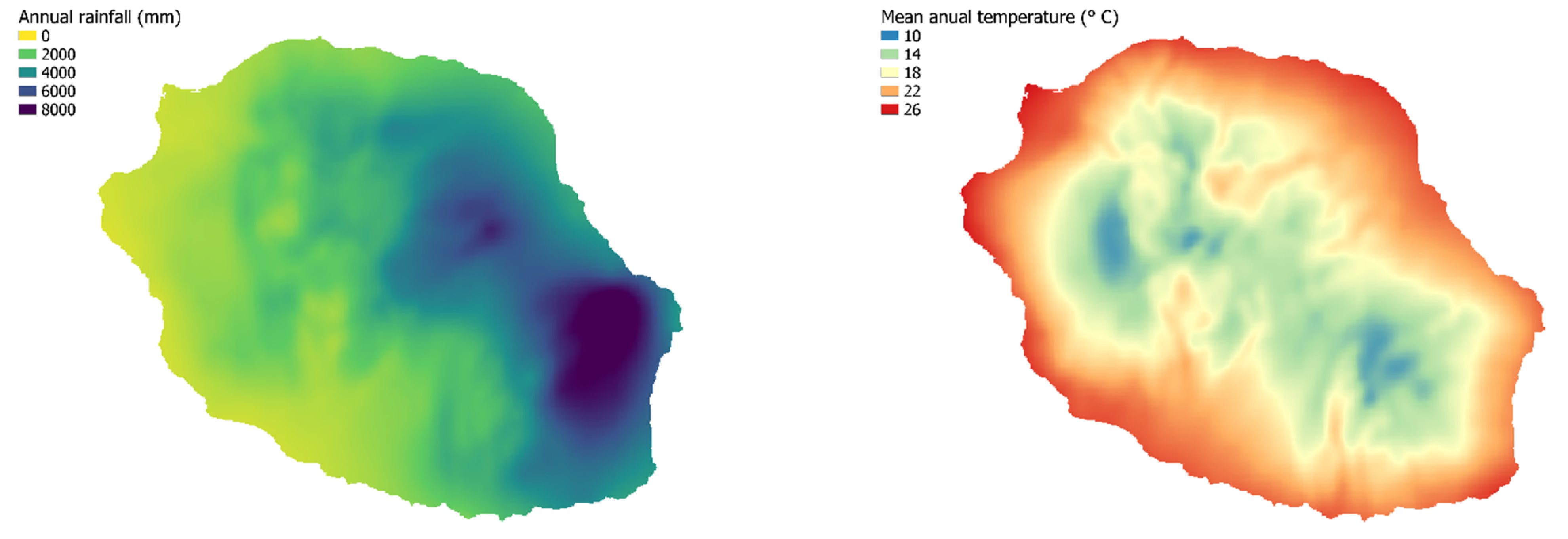
2.1. Study Site
2.2. Sampling Design
2.3. Plot Structure and Composition
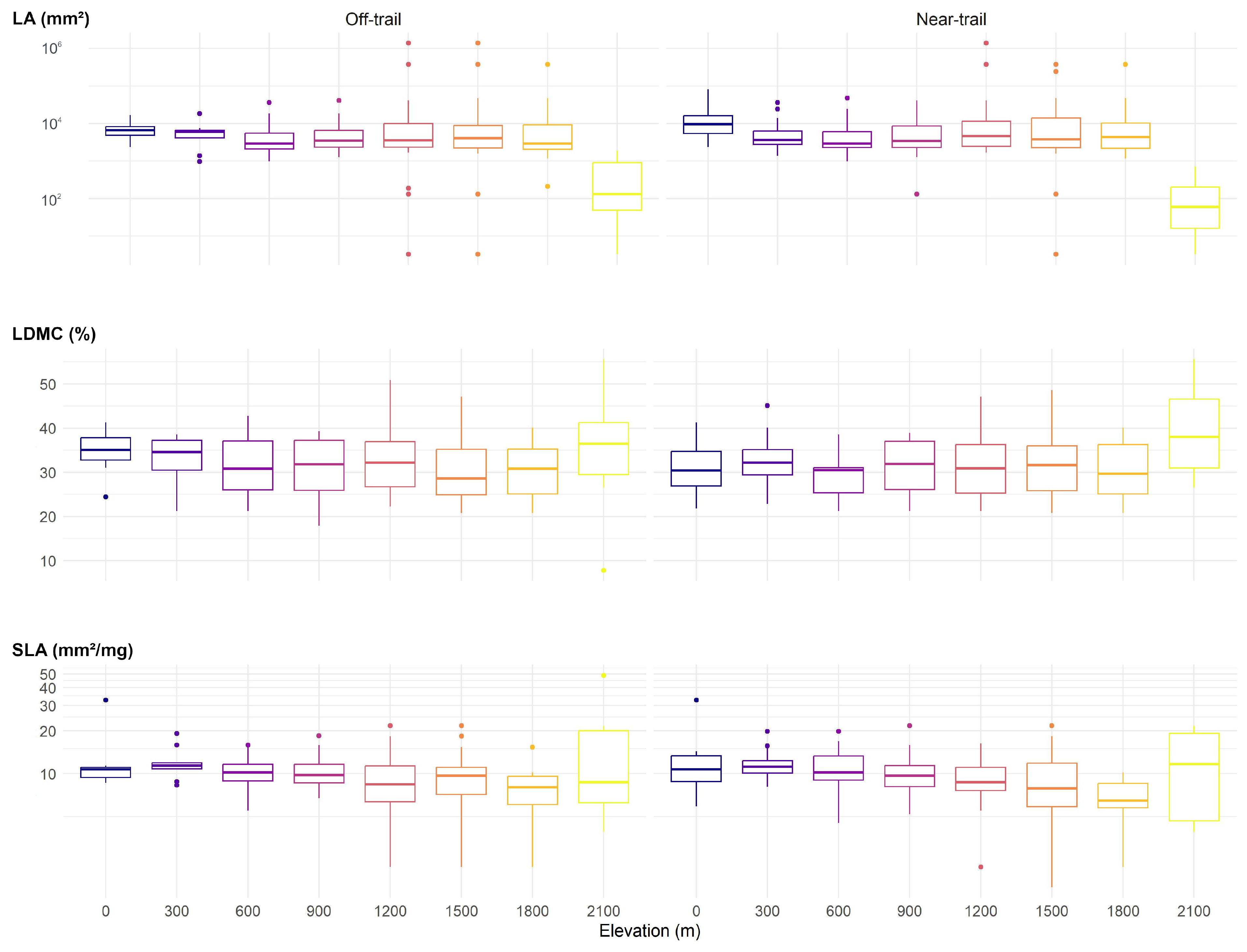
2.4. Leaf Traits
2.5. Data Analyses
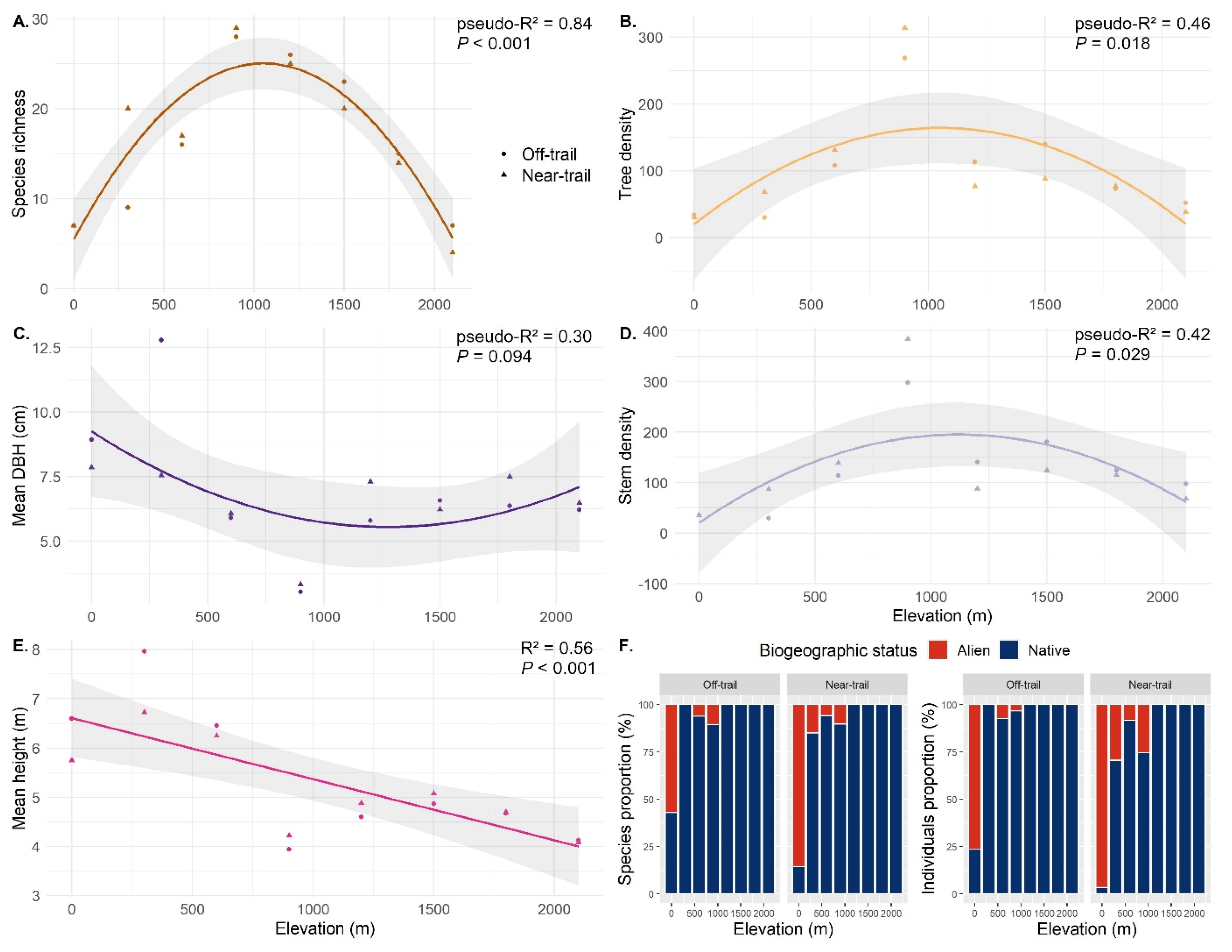
3. Results
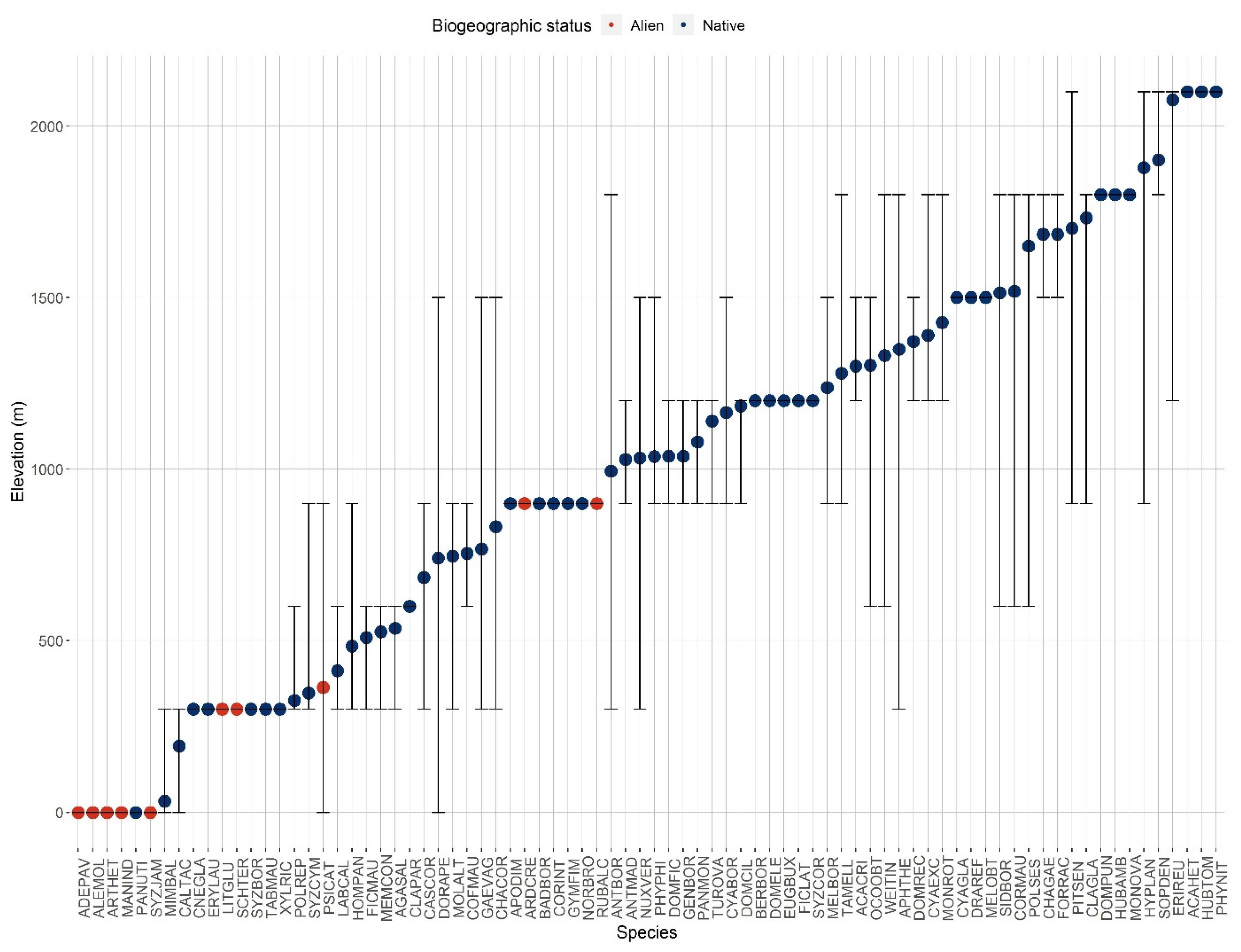
3.1. Plot Inventories
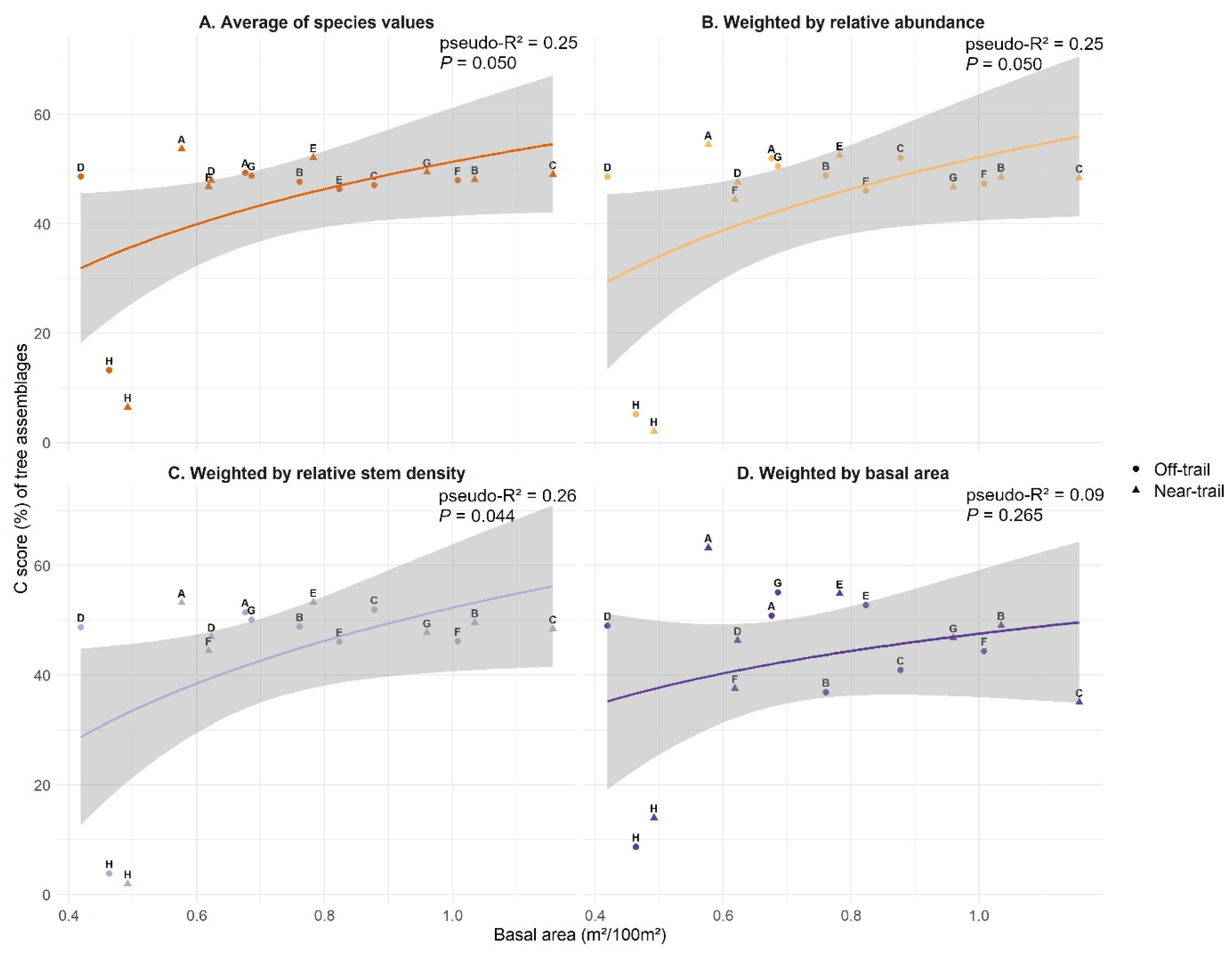
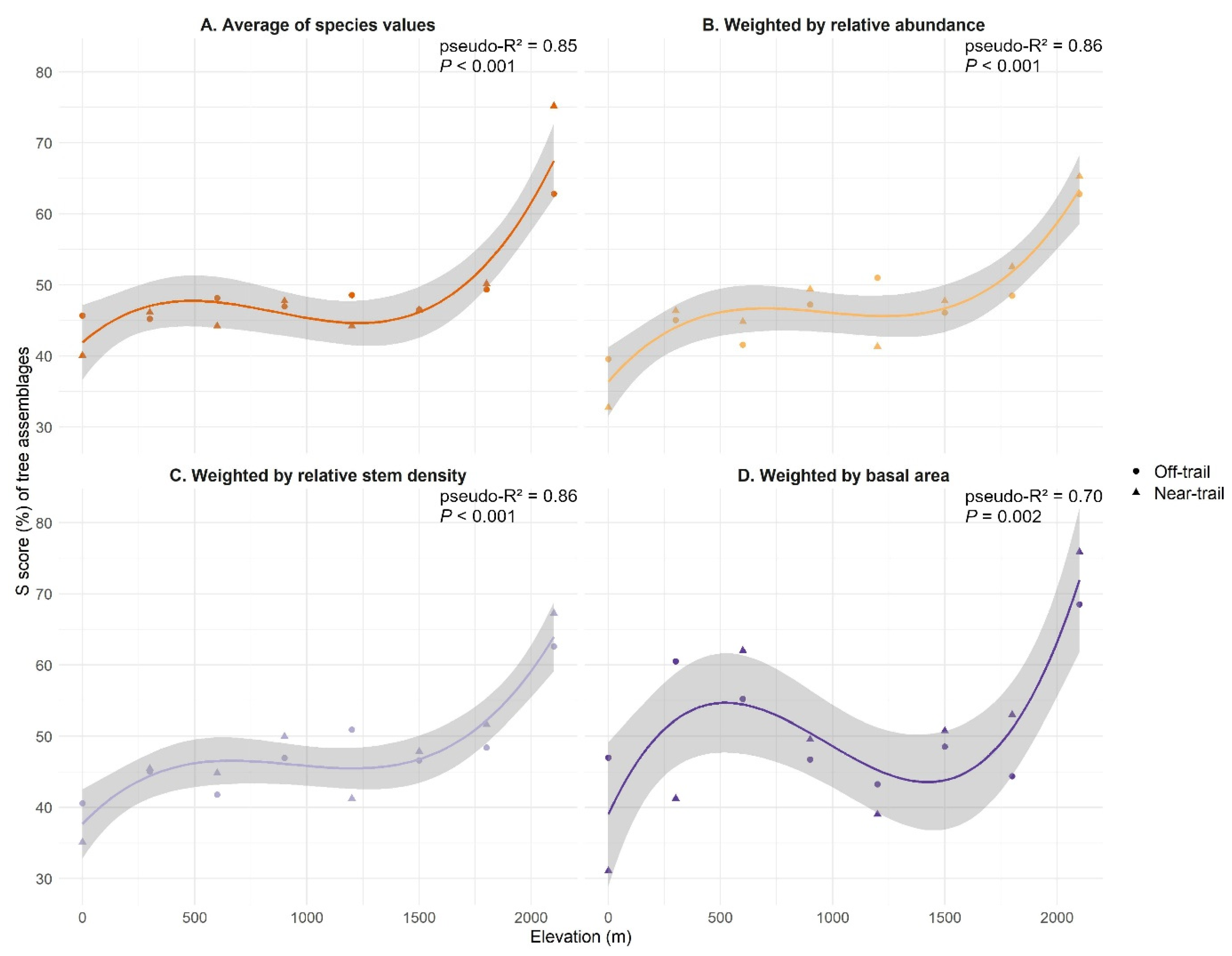
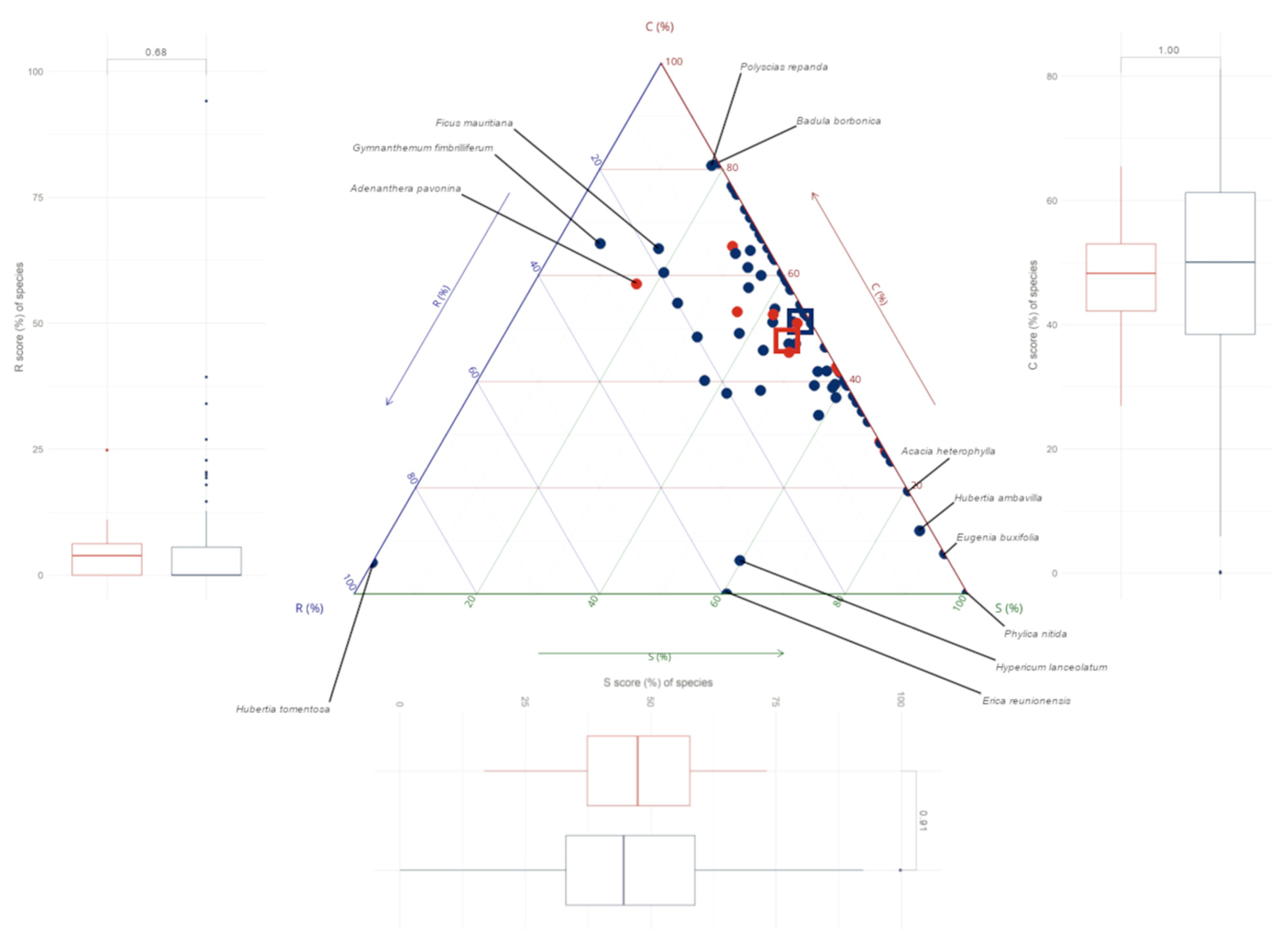
3.2. CSR Strategies of Trees on Réunion
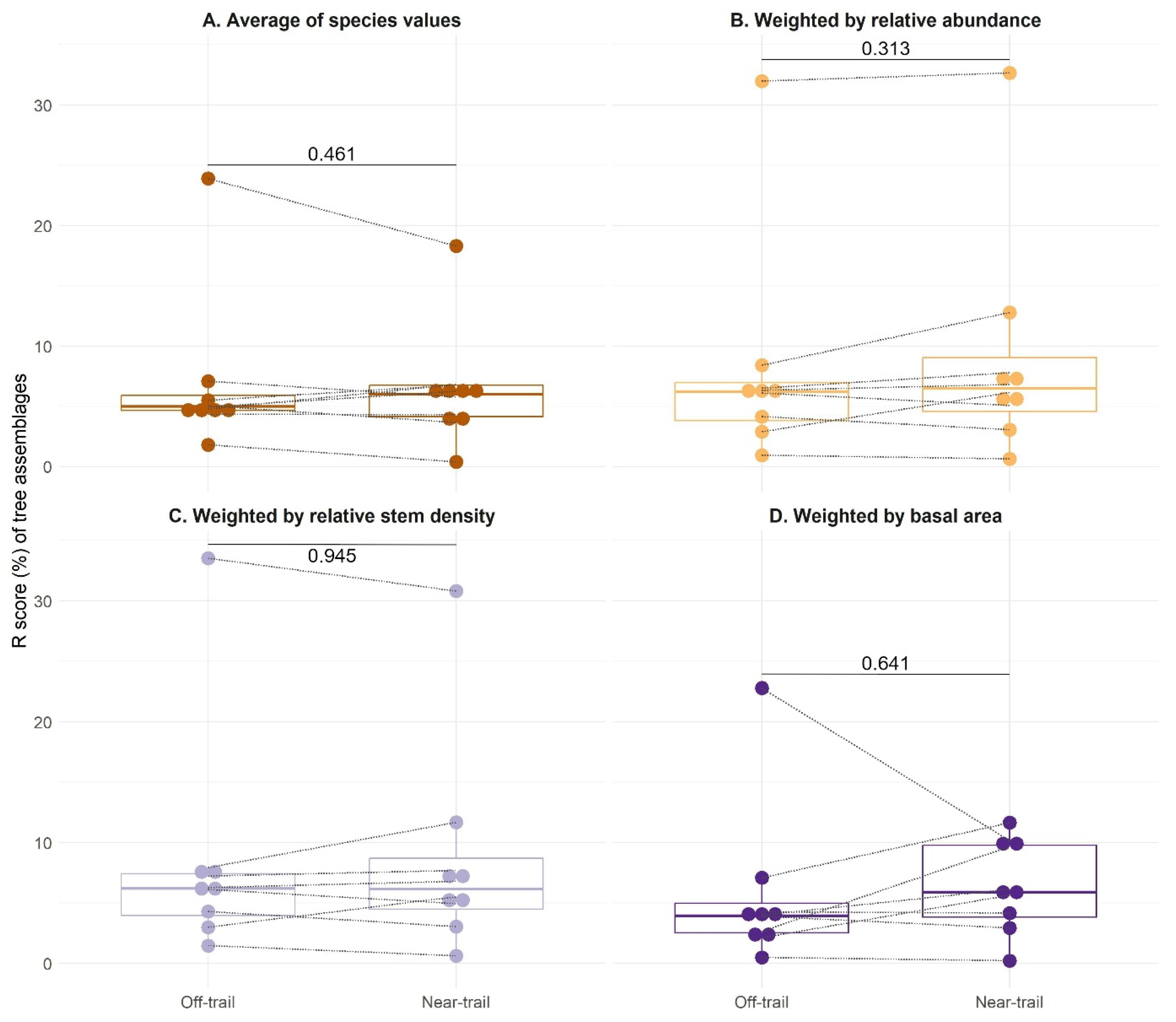
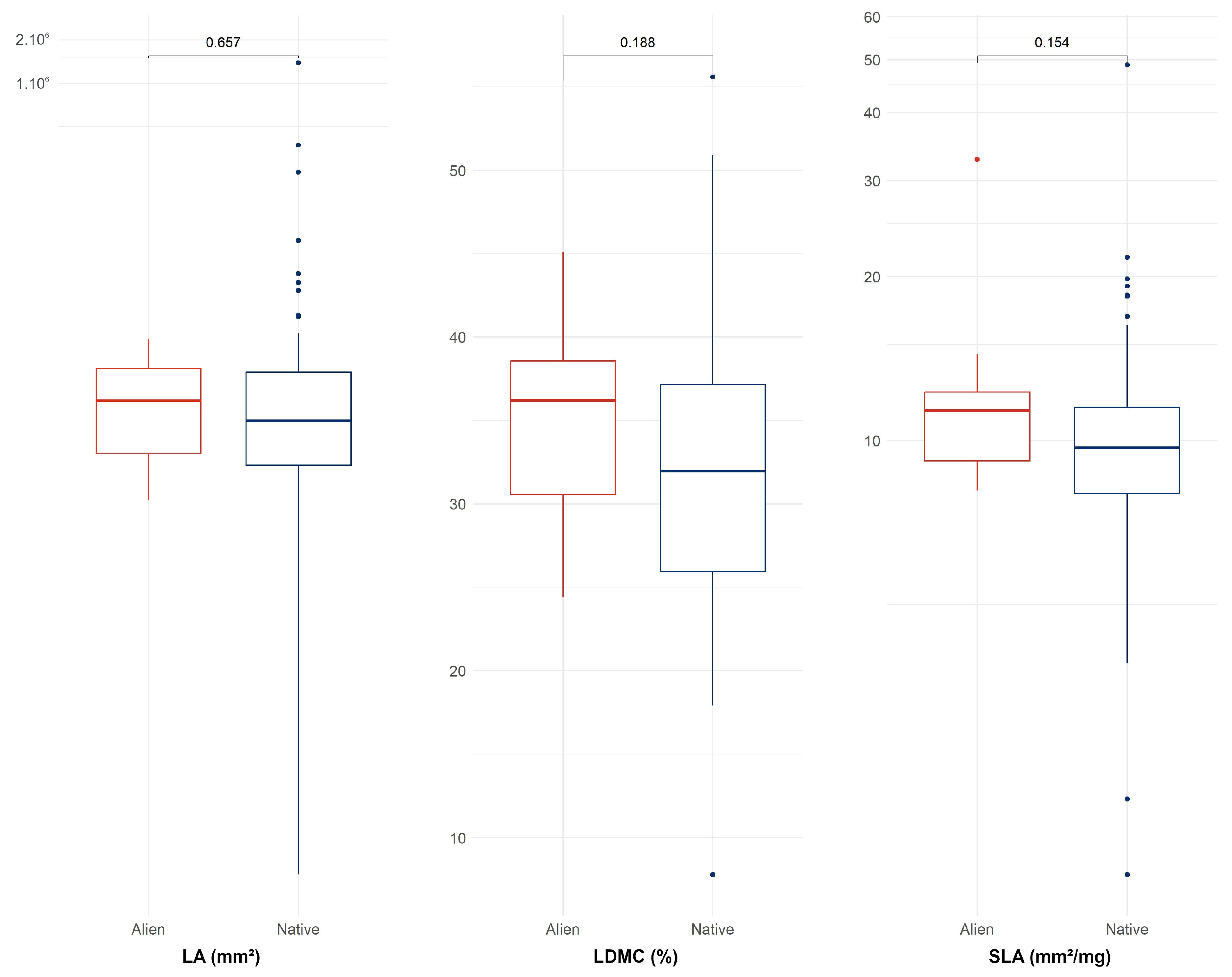
3.3. CSR Strategies of Native and Alien Tree Species on Réunion
4. Discussion
4.1. Assigned Ecological Strategies Were Only Partially Validated for Trees on Réunion
4.2. Native and Alien Trees Use Similar Ecological Strategies on Réunion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | IUCN Status | Protected | Biogeographic Status | Habit | Species ID |
|---|---|---|---|---|---|---|
| Mangifera indica | Anacardiaceae | - | Alien | T | MANIND | |
| Schinus terebinthifolia | Anacardiaceae | - | Alien | T | SCHTER | |
| Xylopia richardii | Annonaceae | LC | X | Endemic (M) | T | XYLRIC |
| Aphloia theiformis | Aphloiaceae | LC | - | Native | T | APHTHE |
| Tabernaemontana mauritiana | Apocynaceae | NT | - | Endemic (M) | S | TABMAU |
| Polyscias cf. sessiliflora | Araliaceae | EN | X | Endemic (R) | S | POLSES |
| Polyscias repanda | Araliaceae | LC | - | Endemic (R) | T | POLREP |
| Acanthophoenix crinita | Arecaceae | VU | - | Endemic (R) | P | ACACRI |
| Cordyline mauritiana | Asparagaceae | LC | - | Endemic (M) | S | CORMAU |
| Dracaena reflexa | Asparagaceae | LC | - | Endemic (IO) | T | DRAREF |
| Gymnanthemum fimbrilliferum | Asteraceae | LC | - | Endemic (R) | S | GYMFIM |
| Hubertia ambavilla | Asteraceae | LC | - | Endemic (M) | S | HUBAMB |
| Hubertia tomentosa | Asteraceae | LC | - | Endemic (R) | S | HUBTOM |
| Memecylon confusum | Bois de balai | LC | - | Endemic (R) | S | MEMCON |
| Calophyllum tacamahaca | Calophyllaceae | LC | - | Endemic (M) | T | CALTAC |
| Cnestis glabra | Connaraceae | LC | - | Endemic (IO) | V | CNEGLA |
| Weinmannia tinctoria | Cunoniaceae | LC | - | Endemic (M) | T | WEITIN |
| Cyathea borbonica | Cyatheaceae | LC | - | Endemic (M) | F | CYABOR |
| Cyathea excelsa | Cyatheaceae | LC | - | Endemic (M) | F | CYAEXC |
| Cyathea glauca | Cyatheaceae | LC | - | Endemic (R) | F | CYAGLA |
| Agarista salicifolia | Ericaceae | LC | - | Native | T | AGASAL |
| Erica reunionensis | Ericaceae | LC | - | Endemic (R) | S | ERIREU |
| Erythroxylum laurifolium | Erythroxylaceae | LC | - | Endemic (M) | S | ERYLAU |
| Forgesia racemosa | Escalloniaceae | LC | - | Endemic (R) | S | FORRAC |
| Mangifera indica | Anacardiaceae | - | Alien | T | MANIND | |
| Aleurites moluccanus | Euphorbiaceae | - | Alien | T | ALEMOL | |
| Claoxylon glandulosum | Euphorbiaceae | LC | - | Endemic (R) | S | CLAGLA |
| Claoxylon parviflorum | Euphorbiaceae | LC | - | Endemic (M) | S | CLAPAR |
| Cordemoya integrifolia | Euphorbiaceae | LC | - | Endemic (M) | T | CORINT |
| Acacia heterophylla | Fabaceae | LC | - | Endemic (R) | T | ACAHET |
| Adenanthera pavonina var. pavonina | Fabaceae | - | Alien | T | ADEPAV | |
| Sophora denudata | Fabaceae | EN | X | Endemic (R) | S | SOPDEN |
| Hypericum lanceolatum | Hypericaceae | LC | - | Endemic (IO) | S | HYPLAN |
| Litsea glutinosa | Lauraceae | - | Alien | T | LITGLU | |
| Ocotea obtusata | Lauraceae | LC | - | Endemic (M) | T | OCOOBT |
| Geniostoma borbonicum | Loganiaceae | LC | - | Endemic (M) | S | GENBOR |
| Dombeya ciliata | Malvaceae | LC | - | Endemic (R) | T | DOMCIL |
| Dombeya elegans | Malvaceae | LC | X | Endemic (R) | S | DOMELE |
| Dombeya ficulnea | Malvaceae | LC | - | Endemic (R) | T | DOMFIC |
| Dombeya punctata | Malvaceae | LC | - | Endemic (R) | T | DOMPUN |
| Dombeya reclinata | Malvaceae | LC | - | Endemic (R) | T | DOMREC |
| Turraea ovata | Meliaceae | VU | - | Endemic (M) | S | TUROVA |
| Apodytes dimidiata | Metteniusaceae | VU | - | Native | T | APODIM |
| Monimia ovalifolia | Monimiaceae | LC | - | Endemic (M) | S | MONOVA |
| Monimia rotundifolia | Monimiaceae | LC | - | Endemic (R) | S | MONROT |
| Tambourissa elliptica | Monimiaceae | LC | - | Endemic (R) | T | TAMELL |
| Artocarpus heterophyllus | Moraceae | - | Alien | T | ARTHET | |
| Ficus lateriflora | Moraceae | LC | - | Endemic (M) | T | FICLAT |
| Ficus mauritiana | Moraceae | LC | - | Endemic (M) | T | FICMAU |
| Eugenia buxifolia | Myrtaceae | LC | - | Endemic (R) | S | EUGBUX |
| Psidium cattleyanum | Myrtaceae | - | Alien | S | PSICAT | |
| Syzygium borbonicum | Myrtaceae | EN | X | Endemic (R) | T | SYZBOR |
| Syzygium cordemoyi | Myrtaceae | LC | - | Endemic (R) | S | SYZCOR |
| Syzygium cymosum | Myrtaceae | LC | - | Endemic (M) | S | SYZCYM |
| Syzygium jambos | Myrtaceae | - | Alien | T | SYZJAM | |
| Noronhia broomeana | Oleaceae | LC | - | Endemic (M) | T | NORBRO |
| Pandanus montanus | Pandanaceae | LC | - | Endemic (R) | S | PANMON |
| Pandanus utilis | Pandanaceae | LC | - | Native | T | PANUTI |
| Antidesma madagascariense | Phyllanthaceae | LC | - | Endemic (IO) | S | ANTMAD |
| Phyllanthus phillyreifolius | Phyllanthaceae | LC | - | Endemic (M) | S | PHYPHI |
| Pittosporum senacia | Pittosporaceae | LC | - | Endemic (IO) | S | PITSEN |
| Ardisia crenata | Primulaceae | - | Alien | S | ARDCRE | |
| Badula borbonica | Primulaceae | LC | X | Endemic (R) | S | BADBOR |
| Phylica nitida | Rhamnaceae | LC | - | Endemic (M) | S | PHYNIT |
| Rubus alceifolius | Rosaceae | - | Alien | V | RUBALC | |
| Antirhea borbonica | Rubiaceae | LC | - | Endemic (IO) | T | ANTBOR |
| Bertiera borbonica | Rubiaceae | DD | X | Endemic (R) | S | BERBOR |
| Chassalia corallioides | Rubiaceae | LC | - | Endemic (R) | S | CHACOR |
| Chassalia gaertneroides | Rubiaceae | LC | - | Endemic (R) | S | CHAGAE |
| Coffea mauritiana | Rubiaceae | LC | - | Endemic (M) | S | COFMAU |
| Gaertnera vaginata | Rubiaceae | LC | - | Endemic (R) | S | GAEVAG |
| Melicope borbonica | Rutaceae | LC | - | Endemic (R) | S | MELBOR |
| Melicope obtusifolia | Rutaceae | LC | - | Endemic (M) | T | MELOBT |
| Casearia coriacea | Salicaceae | LC | - | Endemic (M) | S | CASCOR |
| Homalium paniculatum | Salicaceae | LC | - | Endemic (M) | T | HOMPAN |
| Doratoxylon apetalum | Sapindaceae | LC | - | Endemic (IO) | T | DORAPE |
| Molinaea alternifolia | Sapindaceae | LC | - | Endemic (M) | T | MOLALT |
| Labourdonnaisia calophylloides | Sapotaceae | LC | - | Endemic (M) | T | LABCAL |
| Mimusops balata | Sapotaceae | LC | - | Endemic (M) | T | MIMBAL |
| Sideroxylon borbonicum | Sapotaceae | LC | - | Endemic (R) | T | SIDBOR |
Appendix C

Appendix D

Appendix E. Procedure Used to Test the Sensitivity of Our Comparisons between Native and Alien Species to Their Unbalanced Numbers
Appendix F
| Elevation (m) | A | B | C | Ssim |
|---|---|---|---|---|
| 0 | 7 | 7 | 4 | 0.57 |
| 300 | 20 | 29 | 4 | 0.28 |
| 600 | 17 | 16 | 12 | 0.73 |
| 900 | 29 | 28 | 25 | 0.88 |
| 1200 | 25 | 26 | 18 | 0.71 |
| 1500 | 20 | 23 | 15 | 0.70 |
| 1800 | 14 | 15 | 12 | 0.83 |
| 2100 | 4 | 7 | 4 | 0.73 |
Appendix G

References
- Richardson, D.M.; Pyšek, P.; Carlton, J.T. A Compendium of Essential Concepts and Terminology in Invasion Ecology. In Fifty Years of Invasion Ecology; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2010; pp. 409–420. ISBN 978-1-4443-2998-8. [Google Scholar]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No Saturation in the Accumulation of Alien Species Worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef] [PubMed]
- van Kleunen, M.; Dawson, W.; Essl, F.; Pergl, J.; Winter, M.; Weber, E.; Kreft, H.; Weigelt, P.; Kartesz, J.; Nishino, M.; et al. Global Exchange and Accumulation of Non-Native Plants. Nature 2015, 525, 100–103. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Hoopes, M.F.; Marchetti, M.P. Invasion Ecology; John Wiley & Sons: Hoboken, NJ, USA, 2013; ISBN 1-118-57082-0. [Google Scholar]
- Butchart, S.H.; Walpole, M.; Collen, B.; Van Strien, A.; Scharlemann, J.P.; Almond, R.E.; Baillie, J.E.; Bomhard, B.; Brown, C.; Bruno, J. Global Biodiversity: Indicators of Recent Declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef]
- Pouteau, R.; Brunel, C.; Dawson, W.; Essl, F.; Kreft, H.; Lenzner, B.; Meyer, C.; Pergl, J.; Pyšek, P.; Seebens, H.; et al. Environmental and Socioeconomic Correlates of Extinction Risk in Endemic Species. Divers. Distrib. 2022, 28, 53–64. [Google Scholar] [CrossRef]
- IPBES Thematic Assessment Report on Invasive Alien Species and Their Control. Available online: https://www.ipbes.net/ias (accessed on 25 October 2023).
- Fernández-Palacios, J.M.; Kreft, H.; Irl, S.D.H.; Norder, S.; Ah-Peng, C.; Borges, P.A.V.; Burns, K.C.; de Nascimento, L.; Meyer, J.-Y.; Montes, E.; et al. Scientists’ Warning—The Outstanding Biodiversity of Islands Is in Peril. Glob. Ecol. Conserv. 2021, 31, e01847. [Google Scholar] [CrossRef] [PubMed]
- Kueffer, C.; Daehler, C.C.; Torres-Santana, C.W.; Lavergne, C.; Meyer, J.-Y.; Otto, R.; Silva, L. A Global Comparison of Plant Invasions on Oceanic Islands. Perspect. Plant Ecol. Evol. Syst. 2010, 12, 145–161. [Google Scholar] [CrossRef]
- Meyer, J.-Y. Critical Issues and New Challenges for Research and Management of Invasive Plants in the Pacific Islands. Pac. Conserv. Biol. 2014, 20, 146–164. [Google Scholar] [CrossRef]
- Meyer, J.-Y.; Lavergne, C. Beautés Fatales: Acanthaceae Species as Invasive Alien Plants on Tropical Indo-Pacific Islands. Divers. Distrib. 2004, 10, 333–347. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Dawson, W.; Schlaepfer, D.; Jeschke, J.M.; Fischer, M. Are Invaders Different? A Conceptual Framework of Comparative Approaches for Assessing Determinants of Invasiveness. Ecol. Lett. 2010, 13, 947–958. [Google Scholar] [CrossRef]
- Grinnell, J. The Niche-Relationships of the California Thrasher. Auk 1917, 34, 427–433. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.-L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the Concept of Trait Be Functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Lambdon, P.W.; Lloret, F.; Hulme, P.E. Do Alien Plants on Mediterranean Islands Tend to Invade Different Niches from Native Species? Biol. Invasions 2008, 10, 703–716. [Google Scholar] [CrossRef]
- Pouteau, R.; Hulme, P.E.; Duncan, R.P. Widespread Native and Alien Plant Species Occupy Different Habitats. Ecography 2015, 38, 462–471. [Google Scholar] [CrossRef]
- Gallien, L.; Münkemüller, T.; Albert, C.H.; Boulangeat, I.; Thuiller, W. Predicting Potential Distributions of Invasive Species: Where to Go from Here? Divers. Distrib. 2010, 16, 331–342. [Google Scholar] [CrossRef]
- Zeballos, S.R.; Giorgis, M.A.; Cingolani, A.M.; Cabido, M.; Whitworth-Hulse, J.I.; Gurvich, D.E. Do Alien and Native Tree Species from Central Argentina Differ in Their Water Transport Strategy? Austral Ecol. 2014, 39, 984–991. [Google Scholar] [CrossRef]
- Leishman, M.R.; Thomson, V.P.; Cooke, J. Native and Exotic Invasive Plants Have Fundamentally Similar Carbon Capture Strategies. J. Ecol. 2010, 98, 28–42. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M. The Biogeography of Naturalization in Alien Plants. J. Biogeogr. 2006, 33, 2040–2050. [Google Scholar] [CrossRef]
- Ordonez, A.; Wright, I.J.; Olff, H. Functional Differences between Native and Alien Species: A Global-Scale Comparison. Funct. Ecol. 2010, 24, 1353–1361. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The Worldwide Leaf Economics Spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- da Costa, H.D.J.A.; Gurgel, E.S.C.; do Amaral, D.D.; Vasconcelos, L.V.; Rebelo, L.G.B.; Teodoro, G.S. CSR Ecological Strategies, Functional Traits and Trade-Offs of Woody Species in Amazon Sandplain Forest. Flora 2020, 273, 151710. [Google Scholar] [CrossRef]
- Grime, J.P. Vegetation Classification by Reference to Strategies. Nature 1974, 250, 26–31. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies, Vegetation Processes, and Ecosystem Properties; John Wiley & Sons: Hoboken, NJ, USA, 2006; ISBN 0-470-85040-X. [Google Scholar]
- Grime, J.P.; Pierce, S. The Evolutionary Strategies That Shape Ecosystems; John Wiley & Sons: Hoboken, NJ, USA, 2012; ISBN 1-118-22327-6. [Google Scholar]
- Guo, W.-Y.; van Kleunen, M.; Winter, M.; Weigelt, P.; Stein, A.; Pierce, S.; Pergl, J.; Moser, D.; Maurel, N.; Lenzner, B.; et al. The Role of Adaptive Strategies in Plant Naturalization. Ecol. Lett. 2018, 21, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.-Y.; van Kleunen, M.; Pierce, S.; Dawson, W.; Essl, F.; Kreft, H.; Maurel, N.; Pergl, J.; Seebens, H.; Weigelt, P.; et al. Domestic Gardens Play a Dominant Role in Selecting Alien Species with Adaptive Strategies That Facilitate Naturalization. Glob. Ecol. Biogeogr. 2019, 28, 628–639. [Google Scholar] [CrossRef]
- Caccianiga, M.; Luzzaro, A.; Pierce, S.; Ceriani, R.M.; Cerabolini, B. The Functional Basis of a Primary Succession Resolved by CSR Classification. Oikos 2006, 112, 10–20. [Google Scholar] [CrossRef]
- Hodgson, J.G.; Wilson, P.J.; Hunt, R.; Grime, J.P.; Thompson, K. Allocating CSR Plant Functional Types: A Soft Approach to a Hard Problem. Oikos 1999, 85, 282–294. [Google Scholar] [CrossRef]
- Pierce, S.; Brusa, G.; Vagge, I.; Cerabolini, B.E.L. Allocating CSR Plant Functional Types: The Use of Leaf Economics and Size Traits to Classify Woody and Herbaceous Vascular Plants. Funct. Ecol. 2013, 27, 1002–1010. [Google Scholar] [CrossRef]
- Pierce, S.; Luzzaro, A.; Caccianiga, M.; Ceriani, R.M.; Cerabolini, B. Disturbance Is the Principal A-scale Filter Determining Niche Differentiation, Coexistence and Biodiversity in an Alpine Community. J. Ecol. 2007, 95, 698–706. [Google Scholar] [CrossRef]
- Pierce, S.; Negreiros, D.; Cerabolini, B.E.L.; Kattge, J.; Díaz, S.; Kleyer, M.; Shipley, B.; Wright, S.J.; Soudzilovskaia, N.A.; Onipchenko, V.G.; et al. A Global Method for Calculating Plant CSR Ecological Strategies Applied across Biomes World-Wide. Funct. Ecol. 2017, 31, 444–457. [Google Scholar] [CrossRef]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Colin Prentice, I. The Global Spectrum of Plant Form and Function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef]
- Pierce, S.; Bottinelli, A.; Bassani, I.; Ceriani, R.M.; Cerabolini, B.E.L. How Well Do Seed Production Traits Correlate with Leaf Traits, Whole-Plant Traits and Plant Ecological Strategies? Plant Ecol. 2014, 215, 1351–1359. [Google Scholar] [CrossRef]
- Rosado, B.H.P.; de Mattos, E.A. On the Relative Importance of CSR Ecological Strategies and Integrative Traits to Explain Species Dominance at Local Scales. Funct. Ecol. 2017, 31, 1969–1974. [Google Scholar] [CrossRef]
- Rosenfield, M.F.; Müller, S.C.; Overbeck, G.E. Short Gradient, but Distinct Plant Strategies: The CSR Scheme Applied to Subtropical Forests. J. Veg. Sci. 2019, 30, 984–993. [Google Scholar] [CrossRef]
- Wen, Y.; Chen, C.; He, B.; Lu, X. CSR Ecological Strategies and Functional Traits of the Co-Existing Species along the Succession in the Tropical Lowland Rain Forest. Forests 2022, 13, 1272. [Google Scholar] [CrossRef]
- Dalle Fratte, M.; Bolpagni, R.; Brusa, G.; Caccianiga, M.; Pierce, S.; Zanzottera, M.; Cerabolini, B.E.L. Alien Plant Species Invade by Occupying Similar Functional Spaces to Native Species. Flora 2019, 257, 151419. [Google Scholar] [CrossRef]
- Rojas-Sandoval, J.; Ackerman, J.D.; Marcano-Vega, H.; Willig, M.R. Alien Species Affect the Abundance and Richness of Native Species in Tropical Forests: The Role of Adaptive Strategies. Ecosphere 2022, 13, e4291. [Google Scholar] [CrossRef]
- Macarthur, R.; Levins, R. The Limiting Similarity, Convergence, and Divergence of Coexisting Species. Am. Nat. 1967, 101, 377–385. [Google Scholar] [CrossRef]
- Tecco, P.A.; Urcelay, C.; Diaz, S.; Cabido, M.; Pérez-Harguindeguy, N. Contrasting Functional Trait Syndromes Underlay Woody Alien Success in the Same Ecosystem. Austral Ecol. 2013, 38, 443–451. [Google Scholar] [CrossRef]
- Réchou, A.; Flores, O.; Jumaux, G.; Duflot, V.; Bousquet, O.; Pouppeville, C.; Bonnardot, F. Spatio-Temporal Variability of Rainfall in a High Tropical Island: Patterns and Large-Scale Drivers in Réunion Island. Q. J. R. Meteorol. Soc. 2019, 145, 893–909. [Google Scholar] [CrossRef]
- Gross, N.; Börger, L.; Duncan, R.P.; Hulme, P.E. Functional Differences between Alien and Native Species: Do Biotic Interactions Determine the Functional Structure of Highly Invaded Grasslands? Funct. Ecol. 2013, 27, 1262–1272. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Fernández-Palacios, J.M.; Matthews, T.J.; Borregaard, M.K.; Triantis, K.A. Island Biogeography: Taking the Long View of Nature’s Laboratories. Science 2017, 357, eaam8326. [Google Scholar] [CrossRef] [PubMed]
- Carlquist, S.J. Island Biology; Columbia University Press: New York, NY, USA, 1974; pp. 1–686. ISBN 978-0-231-03562-0. [Google Scholar]
- Whittaker, R.J.; Fernández-Palacios, J.M. Island Biogeography: Ecology, Evolution, and Conservation; Oxford University Press: Oxford, UK, 2007; ISBN 0-19-856611-5. [Google Scholar]
- Barton, K.E.; Fortunel, C. Island Plant Functional Syndromes and Competition with Invasive Species. J. Biogeogr. 2023, 50, 641–653. [Google Scholar] [CrossRef]
- Schrader, J.; Wright, I.J.; Kreft, H.; Westoby, M. A Roadmap to Plant Functional Island Biogeography. Biol. Rev. 2021, 96, 2851–2870. [Google Scholar] [CrossRef]
- DEAL Liste Des Plantes Invasives et Potentiellement Invasives à La Réunion. Available online: https://www.especesinvasives.re/focus/article/liste-especes-flore (accessed on 25 October 2023).
- Strasberg, D.; Rouget, M.; Richardson, D.M.; Baret, S.; Dupont, J.; Cowling, R.M. An Assessment of Habitat Diversity and Transformation on La Réunion Island (Mascarene Islands, Indian Ocean) as a Basis for Identifying Broad-Scale Conservation Priorities. Biodivers. Conserv. 2005, 14, 3015–3032. [Google Scholar] [CrossRef]
- Cadet, T. La Végétation de l’Ile de La Réunion: Étude Phytoécologique et Phytosociologique; Université d’Aix-Marseille: Aix-en Provence, France, 1977. [Google Scholar]
- Liedtke, R.; Barros, A.; Essl, F.; Lembrechts, J.J.; Wedegärtner, R.E.M.; Pauchard, A.; Dullinger, S. Hiking Trails as Conduits for the Spread of Non-Native Species in Mountain Areas. Biol. Invasions 2020, 22, 1121–1134. [Google Scholar] [CrossRef]
- Baret, S.; Strasberg, D. Les Effets de l’ouverture de Sentiers Sur l’invasion Des Zones Protégées de l’île de La Réunion (Archipel Des Mascareignes, Océan Indien) Par Des Espèces Végétales Exotiques. Rev. D’écologie 2005, 60, 325–332. [Google Scholar]
- Boullet, V. Index de La Flore Vasculaire de La Réunion (Trachéophytes): Statuts, Menaces et Protections; Conservatoire Botanique National de Mascarin: Saint-Leu (Réunion), France, 2020. [Google Scholar]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New Handbook for Standardised Measurement of Plant Functional Traits Worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing: Malden, MA, USA, 2004. [Google Scholar]
- Hamilton, N.E.; Ferry, M. Ggtern: Ternary Diagrams Using Ggplot2. J. Stat. Softw. 2018, 87, 1–17. [Google Scholar] [CrossRef]
- Revell, L.J. Phytools: An R Package for Phylogenetic Comparative Biology (and Other Things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Jin, Y.; Qian, H.V. PhyloMaker: An R Package That Can Generate Very Large Phylogenies for Vascular Plants. Ecography 2019, 42, 1353–1359. [Google Scholar] [CrossRef]
- Kunstler, G.; Falster, D.; Coomes, D.A.; Hui, F.; Kooyman, R.M.; Laughlin, D.C.; Poorter, L.; Vanderwel, M.; Vieilledent, G.; Wright, S.J.; et al. Plant Functional Traits Have Globally Consistent Effects on Competition. Nature 2016, 529, 204–207. [Google Scholar] [CrossRef]
- Chesson, P.; Gebauer, R.L.E.; Schwinning, S.; Huntly, N.; Wiegand, K.; Ernest, M.S.K.; Sher, A.; Novoplansky, A.; Weltzin, J.F. Resource Pulses, Species Interactions, and Diversity Maintenance in Arid and Semi-Arid Environments. Oecologia 2004, 141, 236–253. [Google Scholar] [CrossRef] [PubMed]
- Baret, S.; Rouget, M.; Richardson, D.M.; Lavergne, C.; Egoh, B.; Dupont, J.; Strasberg, D. Current Distribution and Potential Extent of the Most Invasive Alien Plant Species on La Réunion (Indian Ocean, Mascarene Islands). Austral Ecol. 2006, 31, 747–758. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Diamond, J.M.; Karr, J.R. Density Compensation in Island Faunas. Ecology 1972, 53, 330–342. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Weber, E.; Fischer, M. A Meta-Analysis of Trait Differences between Invasive and Non-Invasive Plant Species. Ecol. Lett. 2010, 13, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Hulme, P.E.; Bernard-Verdier, M. Comparing Traits of Native and Alien Plants: Can We Do Better? Funct. Ecol. 2018, 32, 117–125. [Google Scholar] [CrossRef]
- Panetta, F.D.; Mckee, J. Recruitment of the Invasive Ornamental, Schinus Terebinthifolius, Is Dependent upon Frugivores. Aust. J. Ecol. 1997, 22, 432–438. [Google Scholar] [CrossRef]
- Yang, Q.-H.; YE, W.-H.; Wang, Z.-M.; Yin, X.-J. Seed Germination Physiology of Ardisia Crenata Var. Bicolor. Seed Sci. Technol. 2009, 37, 291–302. [Google Scholar] [CrossRef]
- Gleditsch, J.M.; Carlo, T.A. Fruit Quantity of Invasive Shrubs Predicts the Abundance of Common Native Avian Frugivores in Central Pennsylvania. Divers. Distrib. 2011, 17, 244–253. [Google Scholar] [CrossRef]
- Spotswood, E.N.; Meyer, J.-Y.; Bartolome, J.W. An Invasive Tree Alters the Structure of Seed Dispersal Networks between Birds and Plants in French Polynesia. J. Biogeogr. 2012, 39, 2007–2020. [Google Scholar] [CrossRef]
- Weiher, E.; Clarke, G.D.P.; Keddy, P.A. Community Assembly Rules, Morphological Dispersion, and the Coexistence of Plant Species. Oikos 1998, 81, 309–322. [Google Scholar] [CrossRef]
- Laughlin, D.C. The Intrinsic Dimensionality of Plant Traits and Its Relevance to Community Assembly. J. Ecol. 2014, 102, 186–193. [Google Scholar] [CrossRef]
- Wang, Y.-J.; Chen, D.; Yan, R.; Yu, F.-H.; van Kleunen, M. Invasive Alien Clonal Plants Are Competitively Superior over Co-Occurring Native Clonal Plants. Perspect. Plant Ecol. Evol. Syst. 2019, 40, 125484. [Google Scholar] [CrossRef]
- Chapla, T.E.; Campos, J.B. Allelopathic Evidence in Exotic Guava (Psidium guajava L.). Braz. Arch. Biol. Technol. 2010, 53, 1359–1362. [Google Scholar] [CrossRef]
- Morgan, E.C.; Overholt, W.A. Potential Allelopathic Effects of Brazilian Pepper (Schinus Terebinthifolius Raddi, Anacardiaceae) Aqueous Extract on Germination and Growth of Selected Florida Native Plants1. J. Torrey Bot. Sci. 2005, 132, 11–15. [Google Scholar] [CrossRef]
- Callaway, R.M.; Ridenour, W.M. Novel Weapons: Invasive Success and the Evolution of Increased Competitive Ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Foster Huenneke, L.; Vitousek, P.M. Seedling and Clonal Recruitment of the Invasive Tree Psidium Cattleianum: Implications for Management of Native Hawaiian Forests. Biol. Conserv. 1990, 53, 199–211. [Google Scholar] [CrossRef]
- Clerissi, C.; Chaïb, S.; Raviglione, D.; Espiau, B.; Bertrand, C.; Meyer, J.-Y. Metabarcoding and Metabolomics Reveal the Effect of the Invasive Alien Tree Miconia Calvescens DC. on Soil Diversity on the Tropical Island of Mo’orea (French Polynesia). Microorganisms 2023, 11, 832. [Google Scholar] [CrossRef]
- Bray, S.R.; Kitajima, K.; Sylvia, D.M. Mycorrhizae Differentially Alter Growth, Physiology, and Competitive Ability of an Invasive Shrub. Ecol. Appl. 2003, 13, 565–574. [Google Scholar] [CrossRef]
- Macdonald, I.A.; Thébaud, C.; Strahm, W.A.; Strasberg, D. Effects of Alien Plant Invasions on Native Vegetation Remnants on La Réunion (Mascarene Islands, Indian Ocean). Environ. Conserv. 1991, 18, 51–61. [Google Scholar] [CrossRef]






| Alien Species | Elevation (m) | Relative Abundance (%) | C:S:R (%) | Ecological Strategy |
|---|---|---|---|---|
| Adenanthera pavonina | 0 | 38 | 58:17:25 | C/CSR |
| Aleurites moluccanus | 0 | 5 | 65:29:06 | C/CS |
| Artocarpus heterophyllus | 0 | 2 | 53:36:11 | CS/CSR |
| Mangifera indica | 0 | 23 | 51:47:02 | CS |
| Syzygium jambos | 0 | 3 | 42:58:00 | CS |
| Litsea glutinosa | 300 | 1 | 27:73:00 | S/CS |
| Schinus terebinthifolia | 300 | 1 | 45:48:06 | CS |
| Ardisia crenata | 900 | 1 | 29:71:00 | S/CS |
| Rubus alceifolius | 900 | 5 | 53:42:05 | CS |
| Psidium cattleyanum | 0; 300; 600; 900 | 16; 18; 8; 10 | 43:57:00 | CS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heymans, L.; Meyer, J.-Y.; Ah-Peng, C.; Ethève, Q.; Flores, O.; Lavergne, C.; Mallet, B.; Parlevliet, H.; Strasberg, D.; Pouteau, R. Do Native and Alien Species Differ in Their Ecological Strategies? A Test with Woody Plants in Tropical Rainforests on Réunion Island (Mascarene Archipelago, Indian Ocean). Plants 2023, 12, 3990. https://doi.org/10.3390/plants12233990
Heymans L, Meyer J-Y, Ah-Peng C, Ethève Q, Flores O, Lavergne C, Mallet B, Parlevliet H, Strasberg D, Pouteau R. Do Native and Alien Species Differ in Their Ecological Strategies? A Test with Woody Plants in Tropical Rainforests on Réunion Island (Mascarene Archipelago, Indian Ocean). Plants. 2023; 12(23):3990. https://doi.org/10.3390/plants12233990
Chicago/Turabian StyleHeymans, Lyse, Jean-Yves Meyer, Claudine Ah-Peng, Quentin Ethève, Olivier Flores, Christophe Lavergne, Bertrand Mallet, Hilde Parlevliet, Dominique Strasberg, and Robin Pouteau. 2023. "Do Native and Alien Species Differ in Their Ecological Strategies? A Test with Woody Plants in Tropical Rainforests on Réunion Island (Mascarene Archipelago, Indian Ocean)" Plants 12, no. 23: 3990. https://doi.org/10.3390/plants12233990
APA StyleHeymans, L., Meyer, J. -Y., Ah-Peng, C., Ethève, Q., Flores, O., Lavergne, C., Mallet, B., Parlevliet, H., Strasberg, D., & Pouteau, R. (2023). Do Native and Alien Species Differ in Their Ecological Strategies? A Test with Woody Plants in Tropical Rainforests on Réunion Island (Mascarene Archipelago, Indian Ocean). Plants, 12(23), 3990. https://doi.org/10.3390/plants12233990