

Selection and Validation of Optimal RT-qPCR Reference Genes for the Normalization of Gene Expression under Different Experimental Conditions in Lindera megaphylla

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Total RNA Extraction and cDNA Synthesis
2.3. Selection of Candidate Reference Genes and Design of RT-qPCR Primers
2.4. RT-PCR and RT-qPCR Data Analysis
2.5. Candidate Reference Gene Expression Stability Analysis
2.6. Validation of Candidate Reference Genes by RT-qPCR
3. Results
3.1. Verification of Amplicon Size, Primers Specificity and PCR Amplification Efficiency
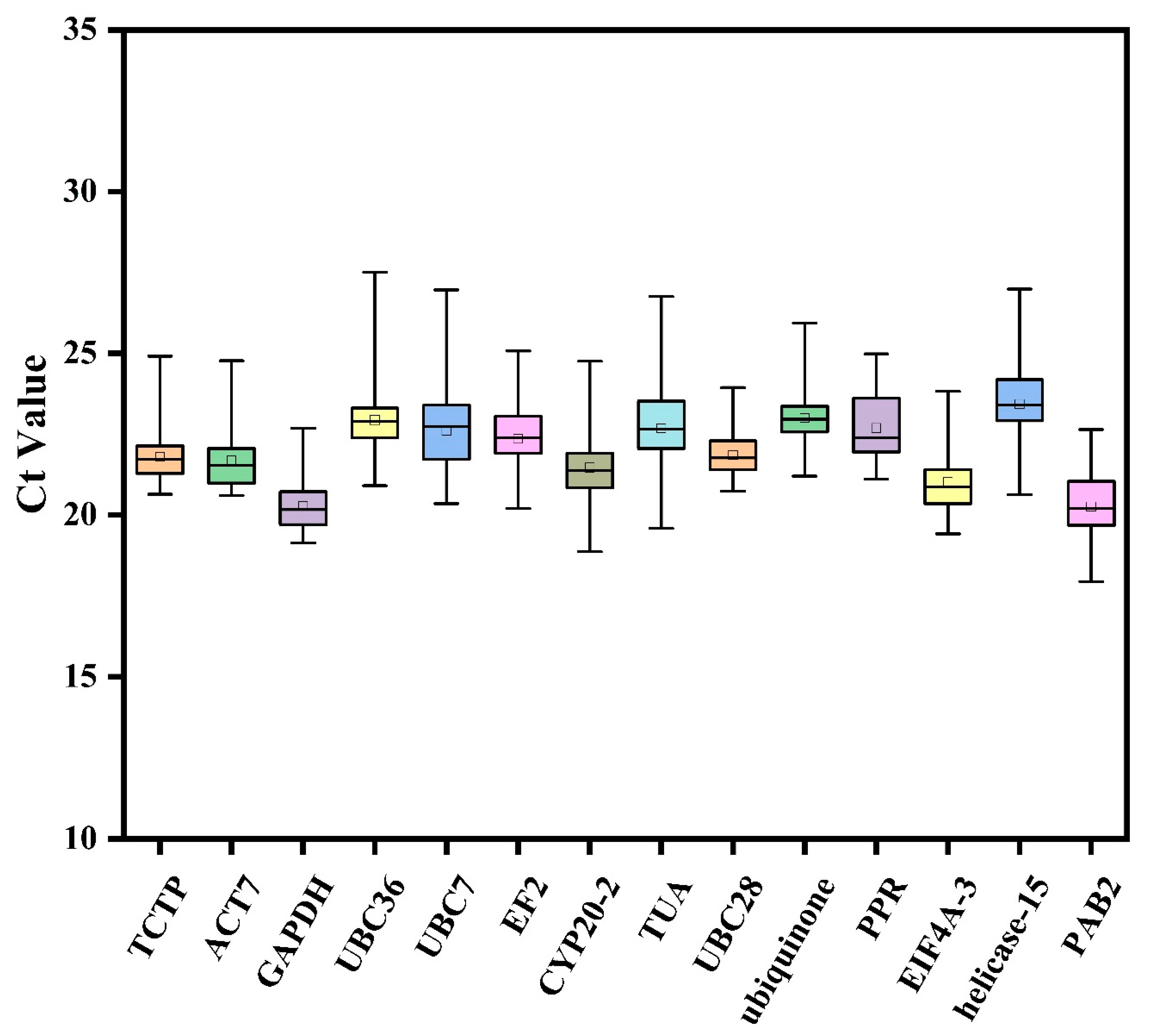
3.2. Transcript Abundance of Candidate Reference Genes
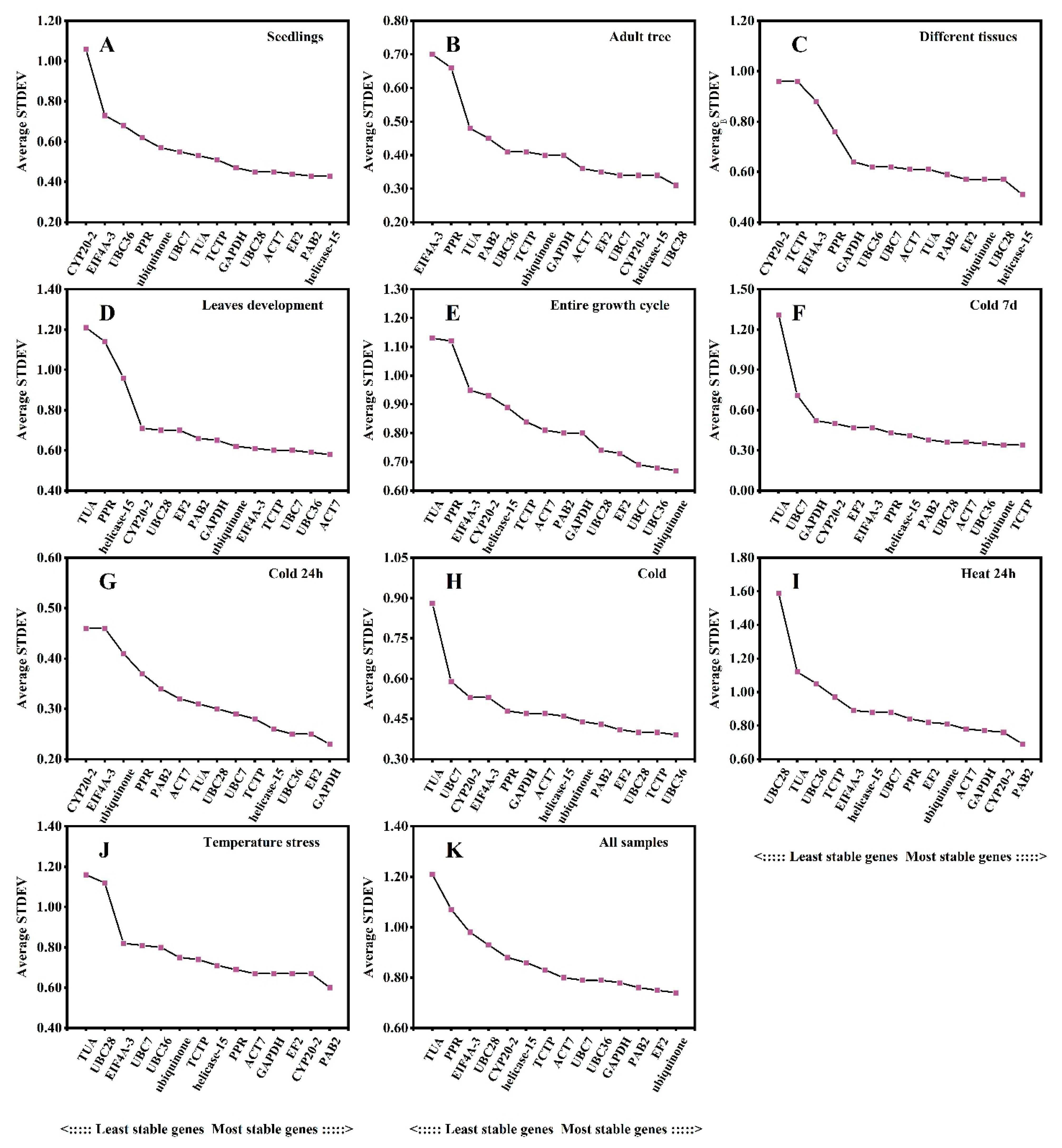
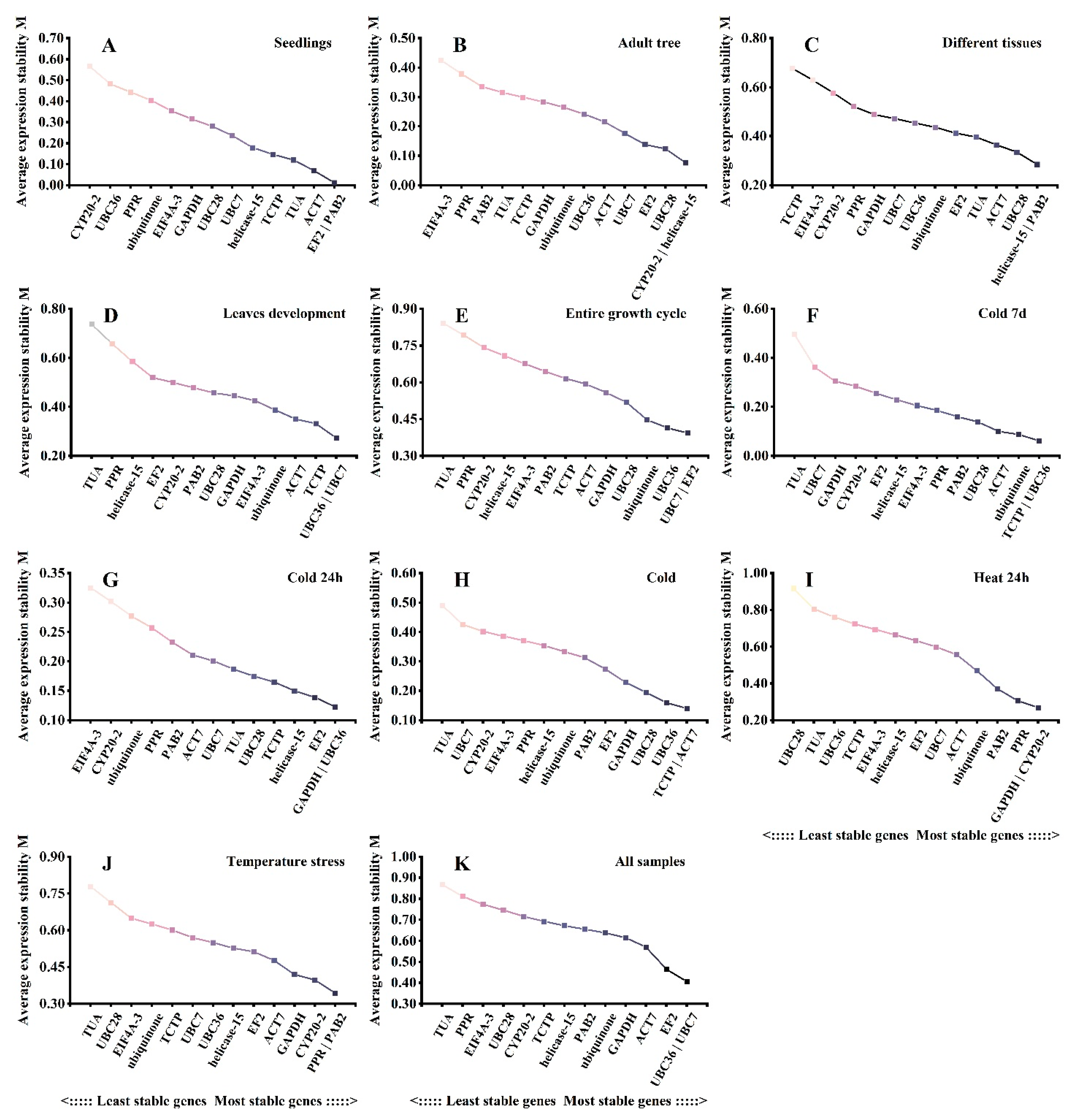
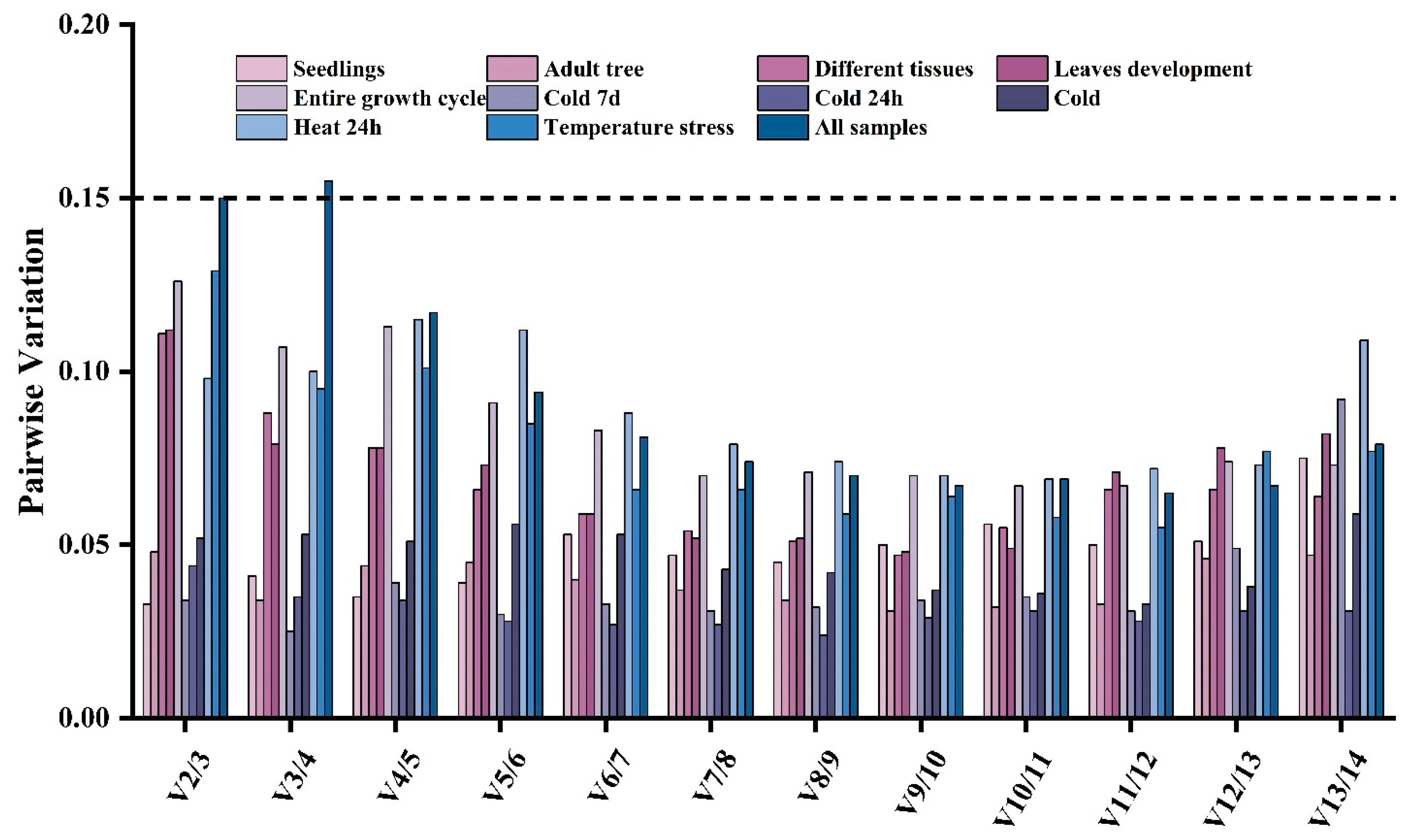
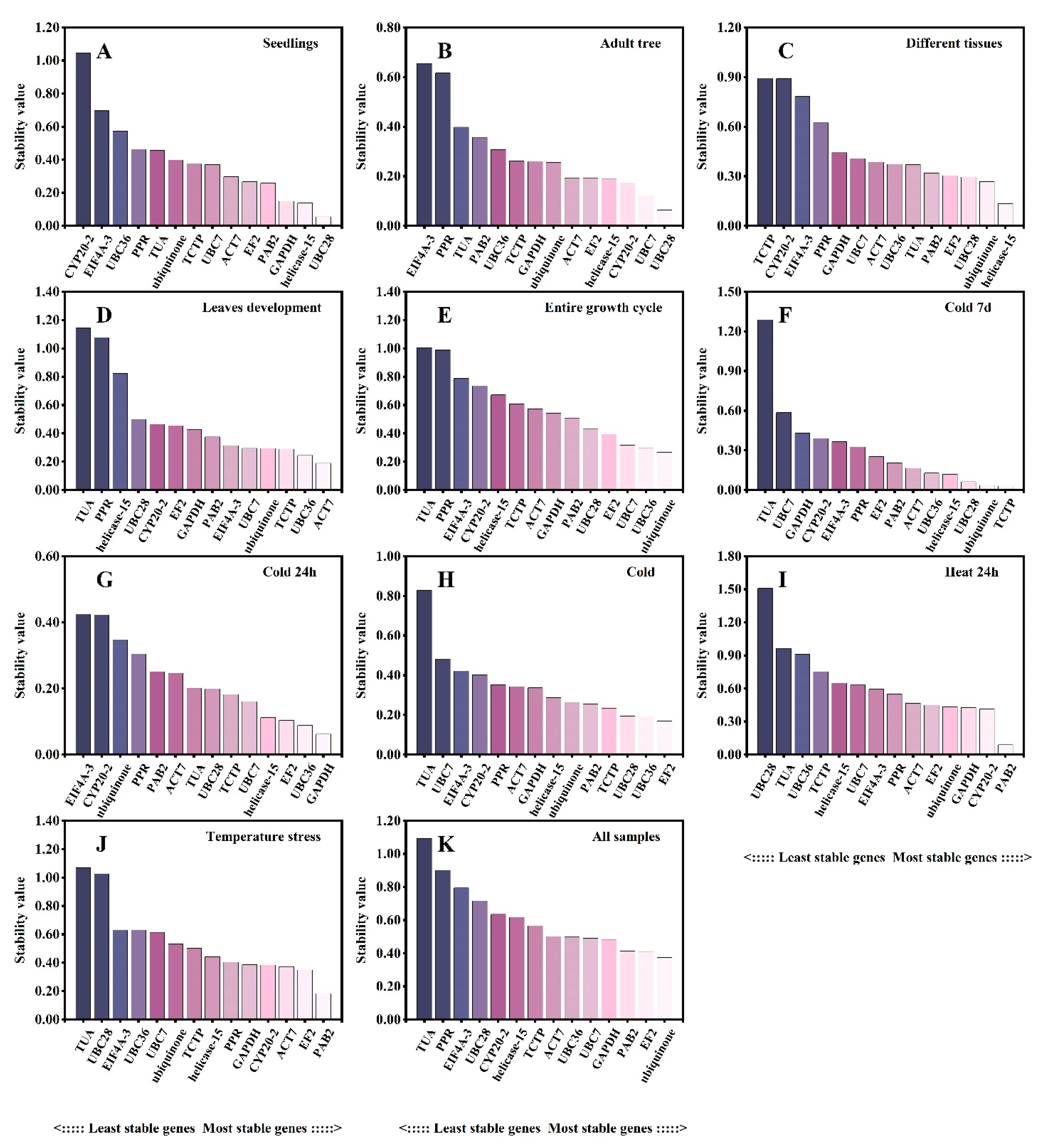
3.3. Estimation of the Stability of the Reference Genes under Different Experimental Conditions
3.4. Delta Ct Method Analysis
3.5. geNorm Analysis
3.6. NormFinder Analysis
3.7. BestKeeper Algorithm
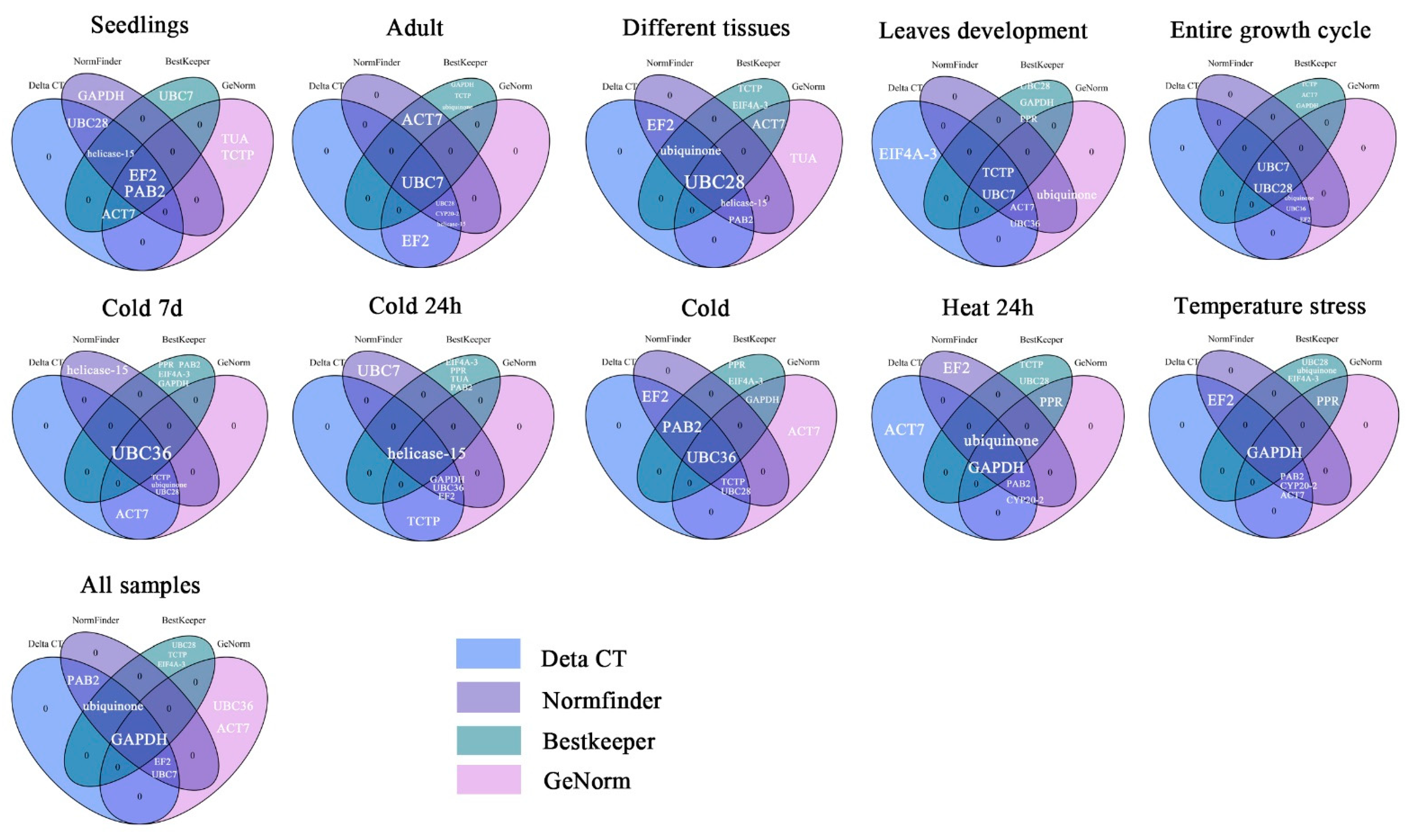
3.8. Comprehensive Stability Analysis Using RefFinder
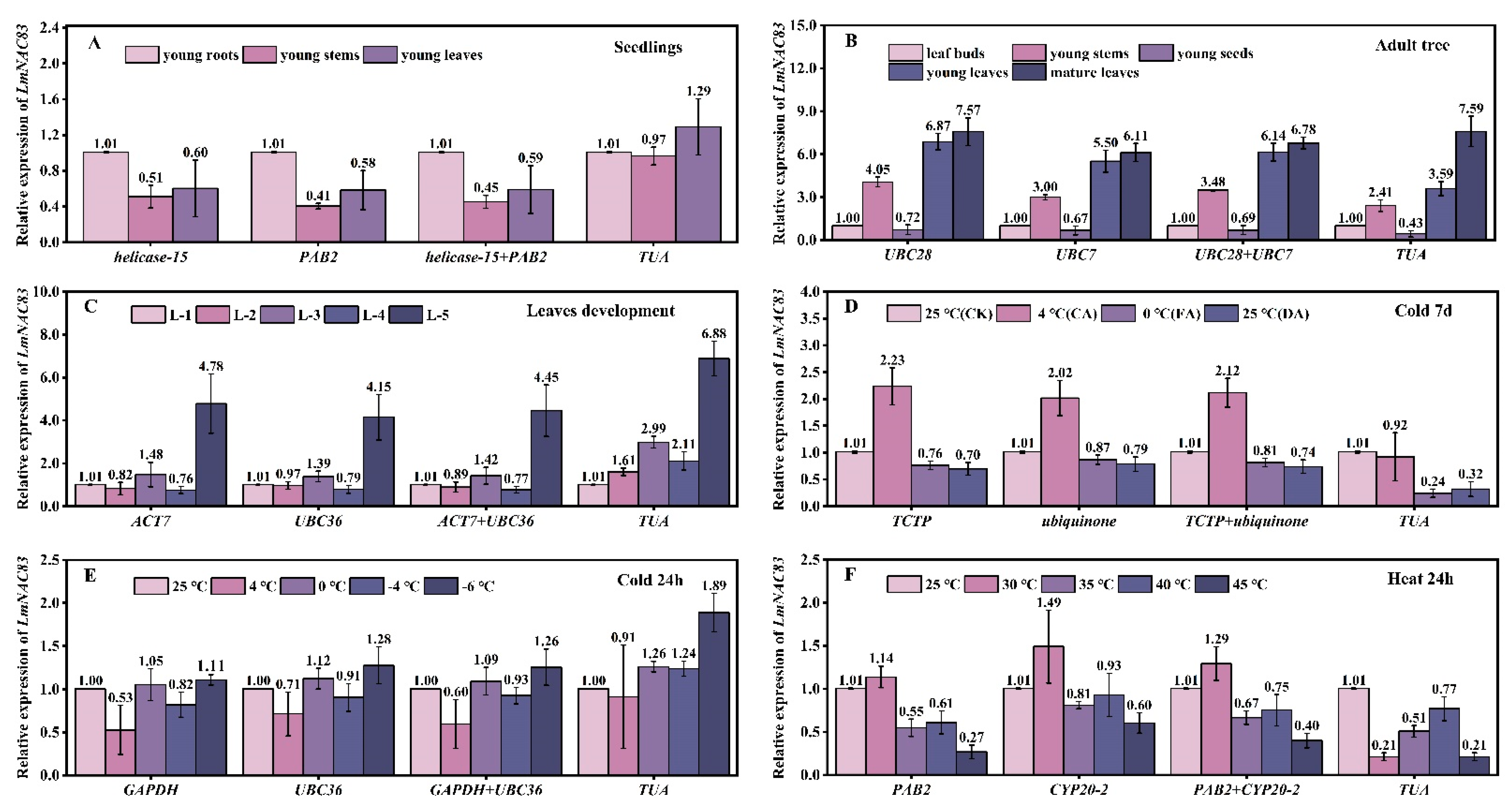
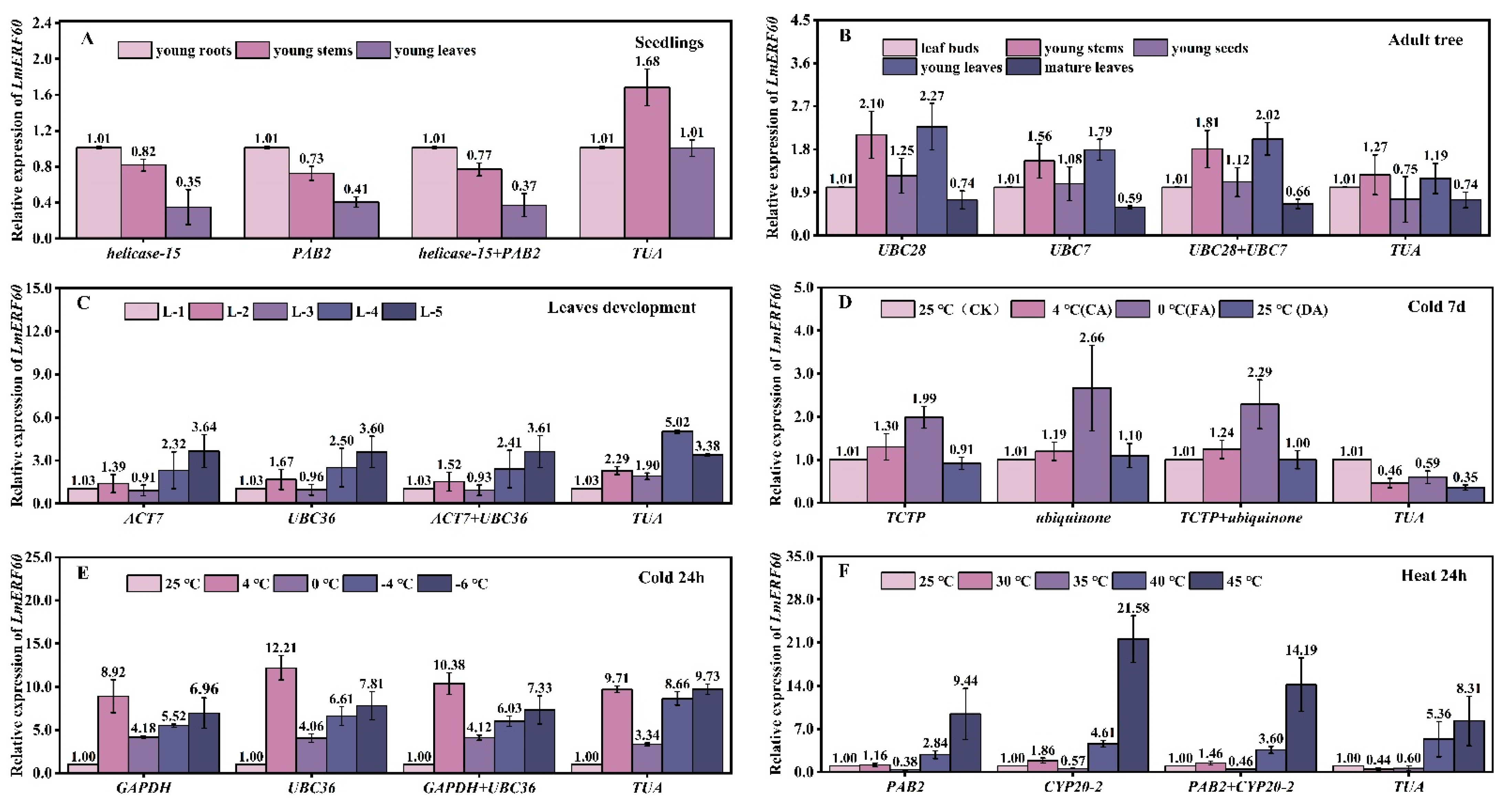
3.9. Validation of Reference Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, K.; Wang, Z.W.; Huang, W.C.; Hu, Y.H. Characterization of two complete chloroplast genomes of Lindera megaphylla (Lauraceae). Mitochondrial DNA B Resour. 2019, 4, 2851–2852. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Xuan, B.; Peng, B.; Li, C.; Chai, X.; Tu, P. The genus Lindera: A source of structurally diverse molecules having pharmacological significance. Phytochem. Rev. 2015, 15, 869–906. [Google Scholar] [CrossRef]
- Lin, H.F.; Huang, H.L.; Liao, J.F.; Shen, C.C.; Huang, R.L. Dicentrine analogue-induced G2/M arrest and apoptosis through inhibition of topoisomerase II activity in human cancer cells. Planta Med. 2015, 81, 830–837. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.L.; Chen, C.C.; Huang, Y.L.; Ou, J.C.; Hu, C.P.; Chen, C.F.; Chang, C. Anti-tumor effects of d-dicentrine from the root of Lindera megaphylla. Planta Med. 1998, 64, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.J.; Lin, L.C.; Chen, K.T.; Chen, C.F. Northalifoline, a new isoquinolone alkaloid from the pedicels of Lindera megaphylla. J. Nat. Prod. 1994, 57, 689–694. [Google Scholar] [CrossRef]
- Wei, S. Effects of Three Plant Volatiles on Negative Air Ions and Air Microorganisms. Master’s Thesis, Zhejiang A&F University, Hangzhou, China, 2021. [Google Scholar] [CrossRef]
- Dong, J.; Zhu, S.; Zhao, H.; Wang, J. The research progress on Lindera megaphylla. J. Henan For. Sci. Technol. 2021, 41, 12–15. [Google Scholar]
- Wu, X.; Yuan, J.; Zhou, M.; Ding, Y.; Liu, M. Research on the overwintering survival of Lindera megaphylla in central China. Anhui Agric. Sci. Bull. 2017, 23, 106–108. [Google Scholar] [CrossRef]
- Yang, J.; Ning, L.; Yang, L.; Wang, J.; Qian, Y.; Chen, Y. Structural characteristics of Lindera megaphylla wood and its volatile organic compounds. J. Zhejiang AF Univ. 2018, 35, 927–934. [Google Scholar] [CrossRef]
- Bian, J.; Cheng, M.; Luo, S.; Chen, S.; Liu, S.; Bai, Z. GC/MS Analysis of volatile substances in essential oilof Lindera megaphylla blade and its application. J. Southwest Univ. (Nat. Sci. Ed.) 2014, 36, 82–88. [Google Scholar] [CrossRef]
- Ding, Q.; Zhang, Y.; Liu, G.; Xu, Z.; Fei, Y. Effects of water stress on photosynthesis characteristics of cinnamomum camphora and Lindera megaphylla Seedlings. J. Southwest For. Univ. 2015, 35, 14–20. [Google Scholar] [CrossRef]
- Teng, K.; Yang, Q.; Dai, G.; Ji, Y. The effect of low temperature stress on physiological and membrtine lipid peroxidatic reaction of Lindera megaphylla seedling. J. Henan Agric. Univ. 2002, 36, 151–154. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Q.; Jiang, Y.; Li, Y.; Zhang, H.; Li, R. Reference genes identification for normalization of qPCR under multiple stresses in Hordeum brevisubulatum. Plant Methods 2018, 14, 110. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, A.; Yang, S.; Huang, L. Reference gene selection for real-time quantitative PCR normalization in Hemarthria compressa and Hemarthria altissima leaf tissue. Mol. Biol. Rep. 2019, 46, 4763–4769. [Google Scholar] [CrossRef] [PubMed]
- Fleige, S.; Pfaffl, M.W. RNA integrity and the effect on the real-time qRT-PCR performance. Mol. Asp. Med. 2006, 27, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Mao, W.; Duan, Z.; Que, Q.; Zhou, W.; Chen, X.; Li, P. Selection and validation of reference genes for measuring gene expression in Toona ciliata under different experimental conditions by quantitative real-time PCR analysis. BMC Plant Biol. 2020, 20, 450. [Google Scholar] [CrossRef]
- Czechowski, T.; Stitt, M.; Altmann, T.; Udvardi, M.K.; Scheible, W.R. Genome-wide identification and testing of superior reference genes for transcript normalization in Arabidopsis. Plant Physiol. 2005, 139, 5–17. [Google Scholar] [CrossRef]
- Chakraborty, S.; Dutta, S.; Biswas, P.; Das, M. Identification of candidate reference genes in tropical bamboos stable across species, tissues, and developmental stages. Biol. Plant. 2019, 63, 253–261. [Google Scholar] [CrossRef]
- Li, F.; Cheng, Y.; Ma, L.; Li, S.; Wang, J. Identification of reference genes provides functional insights into meiotic recombination suppressors in Gerbera hybrida. Hortic. Plant J. 2022, 8, 123–132. [Google Scholar] [CrossRef]
- Qu, R.; Miao, Y.; Cui, Y.; Cao, Y.; Zhou, Y.; Tang, X.; Yang, J.; Wang, F. Selection of reference genes for the quantitative real-time PCR normalization of gene expression in Isatis indigotica fortune. BMC Mol. Biol. 2019, 20, 9. [Google Scholar] [CrossRef]
- Ragni, E.; Vigano, M.; Rebulla, P.; Giordano, R.; Lazzari, L. What is beyond a qRT-PCR study on mesenchymal stem cell differentiation properties: How to choose the most reliable housekeeping genes. J. Cell Mol. Med. 2013, 17, 168–180. [Google Scholar] [CrossRef]
- Chen, M.; Wang, B.; Li, Y.; Zeng, M.; Liu, J.; Ye, X.; Zhu, H.; Wen, Q. Reference gene selection for qRT-PCR analyses of luffa (Luffa cylindrica) plants under abiotic stress conditions. Sci. Rep. 2021, 11, 3161. [Google Scholar] [CrossRef]
- Umadevi, P.; Suraby, E.J.; Anandaraj, M.; Nepolean, T. Identification of stable reference gene for transcript normalization in black pepper-Phytophthora capsici pathosystem. Physiol. Mol. Biol. Plants 2019, 25, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Gai, Y.; Gao, J.; Nie, W.; Bao, Z.; Wang, W.; Xu, X.; Wu, J.; He, Y. Selection and validation of reference genes for quantitative real-time PCR normalization in Psoralea corylifolia (Babchi) under various abiotic stress. J. Plant Physiol. 2022, 274, 153722. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Z.; Luan, Y.; Xu, C.; Tao, J.; Zhao, D. Selection of Stable Reference Genes for Quantitative Real-Time PCR on Herbaceous Peony (Paeonia lactiflora Pall.) in Response to Drought Stress. J. Oceanol. Limnol. 2023, 3, 801–814. [Google Scholar] [CrossRef]
- Yao, Y.; Wang, X.; Chen, B.; Zheng, S.; Wang-Pruski, G.; Chen, X.; Guo, R. Evaluation of Reference Genes Suitable for Gene Expression during Root Enlargement in Cherry Radish Based on Transcriptomic Data. Horticulturae 2023, 9, 20. [Google Scholar] [CrossRef]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative RT-PCR data: A model based variance estimation approach to identify genes suited for normalization applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excelbased tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Dudziak, K.; Sozoniuk, M.; Szczerba, H.; Kuzdralinski, A.; Kowalczyk, K.; Borner, A.; Nowak, M. Identification of stable reference genes for qPCR studies in common wheat (Triticum aestivum L.) seedlings under short-term drought stress. Plant Methods 2020, 16, 58. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xue, J.; Zhu, L.; Hu, H.; Yang, J.; Cui, J.; Xu, J. Selection and optimization of reference genes for microRNA expression normalization by qRT-PCR in chinese cedar (Cryptomeria fortunei) under multiple stresses. Int. J. Mol. Sci. 2021, 22, 7246. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Z.; Ren, M.; Liu, X.; Zhou, X.; Yang, J. Selection of reference genes for RT-qPCR analysis in the hawthorn spider mite, amphitetranychus viennensis (Acarina: Tetranychidae), under acaricide treatments. J. Econ. Entomol. 2022, 115, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, C.; Yang, H.; Lyu, L.; Li, W.; Wu, W. Selection and validation of candidate reference genes for gene expression analysis by RT-qPCR in rubus. Int. J. Mol. Sci. 2021, 22, 10533. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chen, H.; Yuan, J.S.; Kollner, T.G.; Chen, Y.; Guo, Y.; Zhuang, X.; Chen, X.; Zhang, Y.; Fu, J.; et al. The rice terpene synthase gene OsTPS19 functions as an (S)-limonene synthase in planta, and its overexpression leads to enhanced resistance to the blast fungus Magnaporthe oryzae. Plant Biotechnol. J. 2018, 16, 1778–1787. [Google Scholar] [CrossRef]
- Qi, C.; Jiang, H.; Zhao, X.; Mao, K.; Liu, H.; Li, Y.; Hao, Y. The characterization, authentication, and gene expression pattern of the mdcer family in malus domestica. Hortic. Plant J. 2019, 5, 1–9. [Google Scholar] [CrossRef]
- Mergaert, P.; Kereszt, A.; Kondorosi, E. Gene expression in nitrogen-fixing symbiotic nodule cells in Medicago truncatula and other nodulating plants. Plant Cell 2020, 32, 42–68. [Google Scholar] [CrossRef]
- Kiseleva, A.A.; Bragina, M.K.; Muterko, A.F.; Salina, E.A. Functional characterization of genes with daily expression patterns in common wheat. Plant Mol. Biol. 2022, 109, 135–146. [Google Scholar] [CrossRef]
- Lin, L.; Han, X.; Chen, Y.; Wu, Q.; Wang, Y. Identification of appropriate reference genes for normalizing transcript expression by quantitative real-time PCR in Litsea cubeba. Mol. Genet. Genom. 2013, 288, 727–737. [Google Scholar] [CrossRef]
- Zhao, M.; Jin, J.; Gao, T.; Zhang, N.; Jing, T.; Wang, J.; Ban, Q.; Schwab, W.; Song, C. Glucosyltransferase CsUGT78A14 regulates flavonols accumulation and reactive oxygen species scavenging in response to cold stress in Camellia sinensis. Front Plant Sci. 2019, 10, 1675. [Google Scholar] [CrossRef]
- Yang, N.; Liu, J.; Gao, Q.; Gui, S.; Chen, L.; Yang, L.; Huang, J.; Deng, T.; Luo, J.; He, L.; et al. Genome assembly of a tropical maize inbred line provides insights into structural variation and crop improvement. Nat. Genet. 2019, 51, 1052–1059. [Google Scholar] [CrossRef] [PubMed]
- Nicot, N.; Hausman, J.F.; Hoffmann, L.; Evers, D. Housekeeping gene selection for real-time RT-PCR normalization in potato during biotic and abiotic stress. J. Exp. Bot. 2005, 56, 2907–2914. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Xie, F.; Liang, W.; Liang, Y.; Zhang, Z.; Zhao, J.; Hu, G.; Qin, Y. HuNAC20 and HuNAC25, two novel NAC genes from pitaya, confer cold tolerance in transgenic Arabidopsis. Int. J. Mol. Sci. 2022, 23, 2189. [Google Scholar] [CrossRef] [PubMed]
- Mijiti, M.; Wang, Y.; Wang, L.; Habuding, X. Tamarix hispida NAC transcription factor ThNAC4 confers salt and drought stress tolerance to transgenic Tamarix and Arabidopsis. Plants 2022, 11, 2647. [Google Scholar] [CrossRef]
- Srivastava, R.; Kobayashi, Y.; Koyama, H.; Sahoo, L. Cowpea NAC1/NAC2 transcription factors improve growth and tolerance to drought and heat in transgenic cowpea through combined activation of photosynthetic and antioxidant mechanisms. J. Integr. Plant Biol. 2022, 65, 25–44. [Google Scholar] [CrossRef]
- Yu, Y.; Yu, M.; Zhang, S.; Song, T.; Zhang, M.; Zhou, H.; Wang, Y.; Xiang, J.; Zhang, X. Transcriptomic identification of wheat AP2/ERF transcription factors and functional characterization of TaERF-6-3A in response to drought and salinity stresses. Int. J. Mol. Sci. 2022, 23, 3272. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, L.; Hu, H.; Tang, N.; Shi, L.; Xu, F.; Wang, S. Arabidopsis ERF012 is a versatile regulator of plant growth, development and abiotic stress responses. Int. J. Mol. Sci. 2022, 23, 6841. [Google Scholar] [CrossRef]
- De Oliveira, L.F.; Piovezani, A.R.; Ivanov, D.A.; Yoshida, L.; Segal Floh, E.I.S.; Kato, M.J. Selection and validation of reference genes for measuring gene expression in Piper species at different life stages using RT-qPCR analysis. Plant Physiol. Biochem. 2022, 171, 201–212. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, F.; Zhang, X.; Li, Z.; Zhao, Y.; Lohaus, R.; Chang, X.; Dong, W.; Ho, S.Y.W.; Liu, X.; et al. The water lily genome the early evolution of flowering plants. Nature 2020, 577, 79–84. [Google Scholar] [CrossRef]
- Exelby, K.; Herrera-Delgado, E.; Perez, L.G.; Perez-Carrasco, R.; Sagner, A.; Metzis, V.; Sollich, P.; Briscoe, J. Precision of tissue patterning is controlled by dynamical properties of gene regulatory networks. Development 2021, 148, dev197566. [Google Scholar] [CrossRef] [PubMed]
- Pierandrei, S.; Truglio, G.; Ceci, F.; Del Porto, P.; Bruno, S.M.; Castellani, S.; Conese, M.; Ascenzioni, F.; Lucarelli, M. DNA methylation patterns correlate with the expression of SCNN1A, SCNN1B, and SCNN1G (Epithelial Sodium Channel, ENaC) genes. Int. J. Mol. Sci. 2021, 22, 3754. [Google Scholar] [CrossRef] [PubMed]
- Abbas, F.; Ke, Y.; Yu, R.; Yue, Y.; Amanullah, S.; Jahangir, M.M.; Fan, Y. Volatile terpenoids: Multiple functions, biosynthesis, modulation and manipulation by genetic engineering. Planta 2017, 246, 803–816. [Google Scholar] [CrossRef]
- Elbl, P.; Navarro, B.V.; de Oliveira, L.F.; Almeida, J.; Mosini, C.; dos Santos, A.L.W.; Rossi, M.; Floh, E.I.S. Identification and evaluation of reference genes for quantitative analysis of Brazilian pine (Araucaria angustifolia Bertol. Kuntze) gene expression. PLoS ONE 2015, 10, e0136714. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.M.; Guo, C.F.; Peng, J.; Fan, Z.Y.; Hafeez, M.; Ali, D.; Wang, K.; Almarzoug, M.H.A.; Qiu, B.L. Screening and validation of reference genes using in RT-qPCR for gene expression studies in Paederus fuscipes, a medically and agriculturally important insect. J. King Saud Univ. Sci. 2022, 34, 101654. [Google Scholar] [CrossRef]
- Hernandez, F.P.V.; Nunez, M.M.; Rivas, M.R.; Portillo, R.E.V.; Martinez, M.D.B.; Suarez, S.L.; Cardenas, F.D.R. Reference genes for RT-qPCR normalisation in different tissues, developmental stages and stress conditions of amaranth. Plant Biol. 2018, 20, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Jian, B.; Liu, B.; Bi, Y.; Hou, W.; Wu, C.; Han, T. Validation of internal control for gene expression study in soybean by quantitative real-time PCR. BMC Mol. Biol. 2008, 9, 59. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Sample Sets | Tissue Type | Biological Replicates | Sampling Dates | Total Number of Samples |
|---|---|---|---|---|
| Different tissues of oneyear-old seedlings | Roots, stems and leaves | 3 | 1 | 3 |
| Different tissues of adult trees | Leaf buds, young stems, young seeds, young leaves and mature leaves | 3 | 1 | 15 |
| Developmental stages | Leaves | 3 | 15 | 45 |
| Cold stress for 7 days | Leaves | 3 | 4 | 12 |
| Cold stress for 24 h | Leaves | 3 | 5 | 15 |
| Heat stress for 24 h | Leaves | 3 | 5 | 15 |
| Gene Symbol | Gene Name | Primer Sequence (5′ → 3′) | Product Length (bp) | Standard Curve | E (%) | R² |
|---|---|---|---|---|---|---|
| TCTP | translationally controlled tumor protein | F:GTTTCTCACCCTCCAACTTAGG R:CATTTCGCCTCCAGGAACA | 102 | y = −2.4502x + 29.195 | 95.070 | 0.9992 |
| ACT7 | actin-related protein 7 | F:AAGCCAACAGGGAGAAGATG R:CACCCGAGTCCAGAACAATAC | 132 | y = −2.3523x + 28.25 | 103.003 | 0.9971 |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase | F:CGGAGGATGATGTGGTTTCTAC R:GCGACAAGCTTGACAAAGTG | 106 | y = −2.3607x + 27.623 | 98.050 | 0.9994 |
| UBC36 | ubiquitin-conjugating enzyme E2 36 | F:CCCGAAGGTTCGATTTCTCA R:TGAAGAGCAGGACTCCATTTATC | 102 | y = −2.3618x + 29.443 | 101.509 | 0.9978 |
| UBC7 | ubiquitin-conjugating enzyme E2 7 | F:TCATGAGCTTCCCAGCAAATTA R:CGTCCGTCGGGATAAACATTAG | 91 | y = −2.442x + 29.477 | 96.932 | 0.9974 |
| EF2 | elongation factor 2-like | F:GCGGATAAGGGTAGGTTCTTT R:TTCTGGCCAGGAACATAGTTAG | 104 | y = −1.9725x + 27.627 | 96.045 | 0.9919 |
| CYP20-2 | peptidyl-prolyl cis-trans isomerase CYP20-2, chloroplastic | F:AACACCAACGGTAGCCAAT R:TCCAGAACCTGCCCAAATAC | 86 | y = −2.4129x + 27.719 | 101.307 | 0.9963 |
| UBQ | polyubiquitin | F:CCTCGCCGACTACAATATTCA R:CACCTCCAGAGTAATCGTCTTC | 115 | y = −2.2086x + 23.796 | 85.947 | 0.9989 |
| TUA | Alpha-tubulin | F:GCCTTACAACAGTGTGCTTTC R:ATCTAGAGATCGACGGCAGATA | 106 | y = −2.3673x + 27.37 | 101.414 | 0.9972 |
| UBC28 | ubiquitin-conjugating enzyme E2 28-like | F:ACAATTATGGGACCAGCAGATAG R:GGGTGGCTTGAATGGGTAAT | 90 | y = −2.3925x + 28.93 | 101.491 | 0.9972 |
| ICln | chloride conductance regulatory protein ICln | F:TGAGCGACACCGATAGAGAA R:TAAATGCAAGGAGAGGCGTAAG | 103 | y = −2.6401x + 31.012 | 64.420 | 0.9953 |
| ubiquinone | NADH dehydrogenase | F:ATCCGACGGGCGATTAAAG R:TCTAGCCTCTTCTTCCAGATACT | 123 | y = −2.1552x + 28.534 | 107.169 | 0.9975 |
| PPR | pentatricopeptide repeatcontaining protein | F:CTTTAAGCCAGACCAGCAAATG R:TCCTCTTTCAGCCATCTTTCC | 106 | y = −2.3288x + 29.75 | 102.616 | 0.9976 |
| SDE2 | replication stress response regulator SDE2-like | F:TAGACGGGCGGACCAGAT R:GAGGAGGACGGTGCAGGAG | 197 | y = −2.7496x + 30.276 | 86.753 | 0.9912 |
| EIF4A-3 | eukaryotic initiation factor 4A-3like | F:TCTTTGTTGCGGTTGAGCG R:ACCAATCCACCTTTCTTTTCG | 117 | y = −2.8406x + 28.345 | 95.752 | 0.9918 |
| helicase-15 | DEAD-box ATP-dependent RNA helicase 15 | F:CCTGGGAGAATACTGGCACTG R:GGCCTCGTCGTCCACATAA | 249 | y = −3.1364x + 30.269 | 92.361 | 0.9992 |
| PAB2 | polyadenylate-binding protein 2like | F:CCCAAGCTGTTGAGGATCTTA R:CCTTTCAGCTCCATCTCTCTTT | 100 | y = −2.4748x + 31.269 | 91.035 | 0.9919 |
| CYP95 | peptidyl-prolyl cis-trans isomerase CYP95 like | F:GGGTTCAGTCATCGTTACTCTT R:GCGTTCACTTCTTCCTCCATA | 99 | y = −2.4245x + 29.937 | 103.599 | 0.9898 |
| RHA2A | E3 ubiquitin-protein ligase RHA2A | F:CTTTAGCGGGAGCGATGT R:CAAGCACTCTCTGTGGAAGA | 112 | y = −2.3815x + 31.29 | 93.904 | 0.9870 |
| EF1α | Translation elongation factor EF1A | F:AAATGAGGAGGAGCGTGTAAAG R:CGCTGATCATGTTAGGGACATAG | 128 | y = −2.6481x + 30.807 | 83.491 | 0.8779 |
| Ranking | Seedlings | Adult Tree | Different Tissues | Leaf Development | Entire Growth Cycle | Cold 7 d | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene Name | SD | CV (%) | Gene Name | SD | CV (%) | Gene Name | SD | CV (%) | Gene Name | SD | CV (%) | Gene Name | SD | CV (%) | Gene Name | SD | CV (%) | |
| 1 | UBC7 | 0.06 | 0.24 | GAPDH | 0.09 | 0.48 | ACT7 | 0.29 | 1.36 | UBC28 | 0.60 | 2.76 | UBC28 | 0.58 | 2.64 | PPR | 0.06 | 0.27 |
| 2 | helicase-15 | 0.14 | 0.57 | TCTP | 0.18 | 0.83 | UBC28 | 0.31 | 1.40 | GAPDH | 0.66 | 3.24 | TCTP | 0.59 | 2.73 | PAB2 | 0.11 | 0.55 |
| 3 | EF2 | 0.22 | 0.93 | ubiquinone | 0.21 | 0.91 | TCTP | 0.37 | 1.71 | TCTP | 0.71 | 3.27 | UBC7 | 0.69 | 3.01 | EIF4A-3 | 0.14 | 0.68 |
| 4 | PAB2 | 0.22 | 1.01 | UBC7 | 0.24 | 1.08 | EIF4α | 0.37 | 1.80 | UBC7 | 0.73 | 3.21 | ACT7 | 0.69 | 3.20 | GAPDH | 0.21 | 1.04 |
| 5 | ACT7 | 0.27 | 1.28 | ACT7 | 0.25 | 1.20 | ubiquinone | 0.39 | 1.69 | PPR | 0.80 | 3.56 | GAPDH | 0.69 | 3.40 | UBC36 | 0.23 | 1.01 |
| 6 | GAPDH | 0.28 | 1.32 | EIF4A-3 | 0.27 | 1.30 | PTB | 0.39 | 1.85 | ACT7 | 0.80 | 3.67 | ubiquinone | 0.75 | 3.27 | TCTP | 0.25 | 1.15 |
| 7 | UBC28 | 0.3 | 1.35 | UBC28 | 0.30 | 1.37 | TUA | 0.45 | 1.97 | EIF4A-3 | 0.88 | 4.11 | UBC36 | 0.78 | 3.44 | UBC28 | 0.25 | 1.15 |
| 8 | PPR | 0.34 | 1.38 | TUA | 0.35 | 1.54 | helicase-15 | 0.46 | 1.93 | UBC36 | 0.89 | 3.97 | EIF4A-3 | 0.78 | 3.69 | ACT7 | 0.25 | 1.19 |
| 9 | TUA | 0.35 | 1.51 | UBC36 | 0.36 | 1.58 | UBC36 | 0.50 | 2.17 | CYP20-2 | 0.89 | 4.21 | EF2 | 0.89 | 4.04 | ubiquinone | 0.27 | 1.16 |
| 10 | TCTP | 0.37 | 1.73 | EF2 | 0.36 | 1.64 | EF2 | 0.60 | 2.66 | ubiquinone | 0.91 | 3.98 | PPR | 0.95 | 4.17 | CYP20-2 | 0.32 | 1.47 |
| 11 | ubiquinone | 0.46 | 1.93 | CYP20-2 | 0.4 | 1.85 | PPR | 0.69 | 2.92 | EF2 | 1.05 | 4.78 | CYP20-2 | 0.97 | 4.51 | helicase-15 | 0.34 | 1.45 |
| 12 | EIF4A-3 | 0.54 | 2.63 | helicase-15 | 0.42 | 1.79 | UBC7 | 0.69 | 2.97 | PAB2 | 1.06 | 5.34 | PAB2 | 1.02 | 5.05 | EF2 | 0.40 | 1.80 |
| 13 | UBC36 | 0.58 | 2.46 | PAB2 | 0.43 | 2.05 | GAPDH | 0.73 | 3.62 | helicase-15 | 1.38 | 6.02 | helicase-15 | 1.14 | 4.91 | UBC7 | 0.63 | 2.74 |
| 14 | CYP20-2 | 0.80 | 3.40 | PPR | 0.61 | 2.65 | CYP26-2 | 0.96 | 4.33 | TUA | 1.59 | 7.25 | TUA | 1.32 | 5.93 | TUA | 1.10 | 4.79 |
| Ranking | Cold 24 h | Cold | Heat 24 h | Stress Treatment | All Samples | |||||||||||||
| Gene Name | SD | CV (%) | Gene Name | SD | CV (%) | Gene Name | SD | CV (%) | Gene Name | SD | CV (%) | Gene Name | SD | CV (%) | ||||
| 1 | EIF4A-3 | 0.1 | 0.46 | PPR | 0.18 | 0.82 | UBC28 | 0.22 | 1.03 | UBC28 | 0.37 | 1.7 | UBC28 | 0.51 | 2.32 | |||
| 2 | PPR | 0.14 | 0.65 | EIF4A-3 | 0.19 | 0.91 | ubiquinone | 0.54 | 2.35 | PPR | 0.42 | 1.86 | TCTP | 0.56 | 2.57 | |||
| 3 | helicase-15 | 0.3 | 1.29 | PAB2 | 0.29 | 1.42 | TCTP | 0.77 | 3.5 | ubiquinone | 0.45 | 1.94 | GAPDH | 0.6 | 2.98 | |||
| 4 | TUA | 0.32 | 1.41 | UBC36 | 0.3 | 1.29 | PPR | 0.8 | 3.5 | EIF4A-3 | 0.47 | 2.24 | ubiquinone | 0.64 | 2.76 | |||
| 5 | PAB2 | 0.33 | 1.64 | GAPDH | 0.32 | 1.55 | GAPDH | 0.82 | 3.98 | GAPDH | 0.49 | 2.39 | EIF4A-3 | 0.66 | 3.14 | |||
| 6 | GAPDH | 0.35 | 1.69 | ubiquinone | 0.34 | 1.45 | EF2 | 0.83 | 3.55 | TCTP | 0.51 | 2.35 | ACT7 | 0.68 | 3.12 | |||
| 7 | UBC7 | 0.36 | 1.54 | UBC28 | 0.36 | 1.65 | PAB2 | 0.91 | 4.39 | PAB2 | 0.51 | 2.52 | UBC7 | 0.71 | 3.08 | |||
| 8 | ubiquinone | 0.37 | 1.59 | TCTP | 0.37 | 1.71 | EIF4A-3 | 0.91 | 4.44 | CYP20-2 | 0.63 | 2.96 | UBC36 | 0.72 | 3.16 | |||
| 9 | UBC36 | 0.38 | 1.65 | helicase-15 | 0.39 | 1.67 | CYP20-2 | 0.95 | 4.38 | ACT7 | 0.66 | 3.04 | EF2 | 0.81 | 3.62 | |||
| 10 | UBC28 | 0.42 | 1.87 | ACT7 | 0.4 | 1.83 | ACT7 | 1.03 | 4.63 | EF2 | 0.68 | 2.99 | PAB2 | 0.82 | 4.05 | |||
| 11 | EF2 | 0.43 | 1.94 | EF2 | 0.42 | 1.86 | UBC7 | 1.09 | 4.5 | UBC7 | 0.7 | 2.97 | PPR | 0.83 | 3.64 | |||
| 12 | TCTP | 0.51 | 2.3 | UBC7 | 0.43 | 1.84 | helicase-15 | 1.17 | 4.81 | UBC36 | 0.72 | 3.09 | CYP20-2 | 0.85 | 3.94 | |||
| 13 | ACT7 | 0.53 | 2.43 | CYP20-2 | 0.53 | 2.48 | TUA | 1.3 | 5.33 | helicase-15 | 0.73 | 3.08 | helicase-15 | 0.98 | 4.18 | |||
| 14 | CYP20-2 | 0.66 | 3.11 | TUA | 0.67 | 2.93 | UBC36 | 1.36 | 5.64 | TUA | 1.21 | 5.2 | TUA | 1.22 | 5.4 | |||
| Ranking | Seedlings | Adult Tree | Different Tissues | Leaf Development | Entire Growth Cycle | Cold 7 d | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene Name | Geomean of Ranking Values | Gene Name | Geomean of Ranking Values | Gene Name | Geomean of Ranking Values | Gene Name | Geomean of Ranking Values | Gene Name | Geomean of Ranking Values | Gene Name | Geomean of Ranking Values | |
| 1 | PAB2 | 2.21 | UBC28 | 2.21 | helicase-15 | 1.68 | ACT7 | 2.11 | ubiquinone | 2.21 | TCTP | 1.86 |
| 2 | helicase-15 | 2.21 | UBC7 | 2.21 | UBC28 | 2.91 | UBC36 | 2.45 | UBC7 | 2.45 | ubiquinone | 2.71 |
| 3 | EF2 | 2.78 | CYP20-2 | 2.78 | PAB2 | 3.34 | UBC7 | 2.99 | UBC36 | 3.13 | UBC36 | 2.94 |
| 4 | UBC28 | 4.09 | helicase-15 | 4.09 | ACT7 | 3.72 | TCTP | 3.00 | UBC28 | 3.34 | UBC28 | 4.53 |
| 5 | ACT7 | 4.36 | GAPDH | 4.36 | ubiquinone | 3.98 | GAPDH | 5.29 | EF2 | 3.46 | PPR | 4.74 |
| 6 | UBC7 | 4.58 | ACT7 | 4.58 | EF2 | 4.68 | UBC28 | 5.30 | GAPDH | 5.45 | PAB2 | 4.74 |
| 7 | GAPDH | 5.58 | ubiquinone | 5.58 | TUA | 6.19 | ubiquinone | 5.89 | TCTP | 6.00 | ACT7 | 5.57 |
| 8 | TCTP | 7.27 | EF2 | 7.27 | UBC36 | 7.97 | EIF4A-3 | 5.96 | ACT7 | 6.88 | EIF4A-3 | 7.00 |
| 9 | TUA | 7.33 | TCTP | 7.33 | EIF4A-3 | 9.30 | PAB2 | 8.82 | PAB2 | 7.90 | helicase-15 | 7.26 |
| 10 | ubiquinone | 10.22 | UBC36 | 10.22 | UBC7 | 9.46 | CYP20-2 | 9.69 | EIF4A-3 | 10.02 | GAPDH | 9.12 |
| 11 | PPR | 10.38 | TUA | 10.38 | TCTP | 9.53 | EF2 | 10.22 | helicase-15 | 10.94 | EF2 | 9.64 |
| 12 | EIF4A-3 | 11.93 | EIF4A-3 | 11.93 | GAPDH | 10.68 | PPR | 10.72 | CYP20-2 | 11.24 | CYP20-2 | 10.74 |
| 13 | UBC36 | 12.49 | PAB2 | 12.49 | PPR | 11.24 | helicase-15 | 12.24 | PPR | 12.17 | UBC7 | 13.00 |
| 14 | CYP20-2 | 14.00 | PPR | 14.00 | CYP20-2 | 12.98 | TUA | 14.00 | TUA | 14.00 | TUA | 14.00 |
| Ranking | Cold 24 h | Cold | Heat 24 h | Stress Treatment | All Samples | |||||||
| Gene Name | Geomean of Ranking Values | Gene Name | Geomean of Ranking Values | Gene Name | Geomean of Ranking Values | Gene Name | Geomean of Ranking Values | Gene Name | Geomean of Ranking Values | |||
| 1 | GAPDH | 1.57 | UBC36 | 2.21 | PAB2 | 2.30 | PAB2 | 1.57 | ubiquinone | 2.21 | ||
| 2 | UBC36 | 2.45 | TCTP | 3.13 | CYP20-2 | 2.45 | PPR | 2.91 | EF2 | 3.22 | ||
| 3 | helicase-15 | 3.72 | UBC28 | 3.60 | GAPDH | 2.59 | ACT7 | 4.05 | UBC7 | 3.64 | ||
| 4 | EF2 | 4.15 | EF2 | 4.03 | ubiquinone | 3.76 | GAPDH | 4.16 | GAPDH | 3.94 | ||
| 5 | UBC7 | 6.40 | PAB2 | 4.79 | PPR | 4.92 | EF2 | 4.68 | UBC36 | 4.12 | ||
| 6 | TUA | 6.51 | ACT7 | 5.33 | ACT7 | 6.16 | CYP20-2 | 4.68 | PAB2 | 5.01 | ||
| 7 | TCTP | 6.51 | PPR | 5.62 | EF2 | 6.16 | UBC28 | 6.85 | TCTP | 5.83 | ||
| 8 | PPR | 7.18 | GAPDH | 6.32 | UBC28 | 7.24 | ubiquinone | 7.19 | ACT7 | 5.86 | ||
| 9 | EIF4A-3 | 7.24 | ubiquinone | 6.45 | TCTP | 7.95 | helicase-15 | 8.17 | UBC28 | 6.04 | ||
| 10 | UBC28 | 7.36 | EIF4A-3 | 7.50 | UBC7 | 8.89 | TCTP | 8.18 | helicase-15 | 9.58 | ||
| 11 | PAB2 | 8.41 | helicase-15 | 7.94 | EIF4A-3 | 8.94 | EIF4A-3 | 8.92 | EIF4A-3 | 9.64 | ||
| 12 | ACT7 | 9.87 | CYP20-2 | 11.72 | helicase-15 | 9.64 | UBC7 | 10.22 | CYP20-2 | 10.47 | ||
| 13 | ubiquinone | 10.84 | UBC7 | 12.74 | UBC36 | 12.47 | UBC36 | 10.36 | PPR | 12.47 | ||
| 14 | CYP20-2 | 13.24 | TUA | 14.00 | TUA | 13.00 | TUA | 14.00 | TUA | 14.00 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Liu, J.; Chen, P.; Zhang, X.; Wang, K.; Lu, J.; Li, Y. Selection and Validation of Optimal RT-qPCR Reference Genes for the Normalization of Gene Expression under Different Experimental Conditions in Lindera megaphylla. Plants 2023, 12, 2185. https://doi.org/10.3390/plants12112185
Liu H, Liu J, Chen P, Zhang X, Wang K, Lu J, Li Y. Selection and Validation of Optimal RT-qPCR Reference Genes for the Normalization of Gene Expression under Different Experimental Conditions in Lindera megaphylla. Plants. 2023; 12(11):2185. https://doi.org/10.3390/plants12112185
Chicago/Turabian StyleLiu, Hongli, Jing Liu, Peng Chen, Xin Zhang, Ke Wang, Jiuxing Lu, and Yonghua Li. 2023. "Selection and Validation of Optimal RT-qPCR Reference Genes for the Normalization of Gene Expression under Different Experimental Conditions in Lindera megaphylla" Plants 12, no. 11: 2185. https://doi.org/10.3390/plants12112185
APA StyleLiu, H., Liu, J., Chen, P., Zhang, X., Wang, K., Lu, J., & Li, Y. (2023). Selection and Validation of Optimal RT-qPCR Reference Genes for the Normalization of Gene Expression under Different Experimental Conditions in Lindera megaphylla. Plants, 12(11), 2185. https://doi.org/10.3390/plants12112185