1. Introduction
Catharanthus roseus (L.) G. Don, a member of the Apocynaceae family, is a popular flowering plant. It is an indigenous species to Madagascar and is widely distributed throughout the African, American, Asian and southern European regions. In India,
C. roseus has been spread across all the major parts of Gujarat, Madhya Pradesh, Assam, Bihar, Uttar Pradesh, Karnataka and Tamil Nadu [
1]. The plant is well known for both its ornamental and medicinal value. It produces nearly 130 alkaloids, of which vincristine and vinblastine are the two major compounds that are used in the treatment of leukemia and Hodgkin’s lymphoma [
2]. For decades, this plant has been exploited for pharmaceutically active compounds from its native environments and thus is at risk of declining in the wild. Plant tissue culture proves to be an effective biotechnological tool for the rapid propagation of plants under aseptic conditions with a lesser risk of microbial infections [
3]. Several in vitro studies using different explants have been successfully conducted for somatic embryogenesis [
4] and organogenesis in
C. roseus [
5,
6].
In recent times, double haploid (DH) production via anther is a promising option for developing improved plant varieties with high yields of medicinally important bioactive compounds [
7]. In vitro anther culture has been attempted in various plants such as
Actinidia arguta Planch [
8] and
Triticum aestivum L. [
9]. Various factors such as stage of anther, culture conditions, plant growth regulators (PGRs) and genotypic and ploidy status determine the success of DH generation [
10]. These factors necessitate ascertaining the ploidy status of anther-derived callus to generate true-to-type DH lines, which can be performed with a flow cytometric technique. The flow cytometry method (FCM) measures the genome size by examining the nuclei at a relatively faster rate and thus validates the ploidy levels of different plant tissues [
11]. Recent investigations of genome size analysis using FCM have been reported for different plants [
12,
13]. Phytochemical profiling using gas chromatography coupled with mass spectrometry (GC–MS) has emerged as an important procedure for identifying and quantifying therapeutically significant compounds present in medicinal plants. This technique is relatively faster, accurate and needs a minimum volume of extracts to detect a wide range of bioactive compounds such as alkaloids, long-chain hydrocarbons, steroids, sugars, amino acids and nitro compounds [
14]. Major bioactive compounds extracted from different plant parts of
C. roseus such as stem, root and leaf include vincristine, vinblastine, reserpine, ajmalicine, vindolinine and catharine, which possess anti-cancerous, anti-diabetic, anti-fungal and anti-microbial activities [
15]. GC–MS-based profiling has been recently reported for several plants including
Silybum marianum L. [
16] and
Chukrasia velutina [
17], but the information on tissue-culture-raised plants’ phytocompound profiling is relatively much less. The present work, therefore, focuses on investigating the ploidy status of anther-derived callus of
C. roseus using flow cytometry. The elemental composition of both anther and anther calli was studied using a scanning electron microscopy–energy-dispersive X-ray microanalysis (SEM–EDX) technique. The identification of the bioactive compounds present in methanolic extracts of anther and anther-derived calli was conducted for the first time in
C. roseus using GC–MS analysis. This report will help to understand and improve the yield of the important pharmaceutical compounds synthesized from anther-derived callus.
3. Discussion
The present work was conducted to evaluate the callusing potentiality of anthers of
C. roseus under in vitro culture conditions. The type and concentration of PGRs used in media strongly affect callusing ability and are different in different plant species. Initially, the anthers were subject to different concentrations and combinations of PGRs amended in MS medium. The results indicate that a high-to-low ratio of auxin: cytokinin concentrations was proven to be the best in inducing callus with a maximum mean fresh weight, which is very similar to Kou et al.’s [
19] and Rout et al.’s [
20] observations. Likewise, TDZ alone at different concentrations was found to be equally effective in producing callus and subsequent proliferation. Previous reports suggested that TDZ (a cytokinin-like PGR) alone may be used in improving callusing ability in different explants [
21,
22]. A comparison of the elemental distribution on the surfaces of anther and anther-derived callus was performed using SEM–EDX analysis, revealing a nearly similar elemental composition on both samples. EDX analyzes X-rays emitted from samples receiving a high-energy electron beam. This technique facilitates the qualitative and semi-quantitative detection of surface elements of samples and has been extensively used on various plant species such as sesame [
23] and lemongrass [
24].
Medicinal plants are an ingenious source of bioactive compounds that fight against several chronic diseases, and these phytocompounds can be identified and quantified using the GC–MS technique [
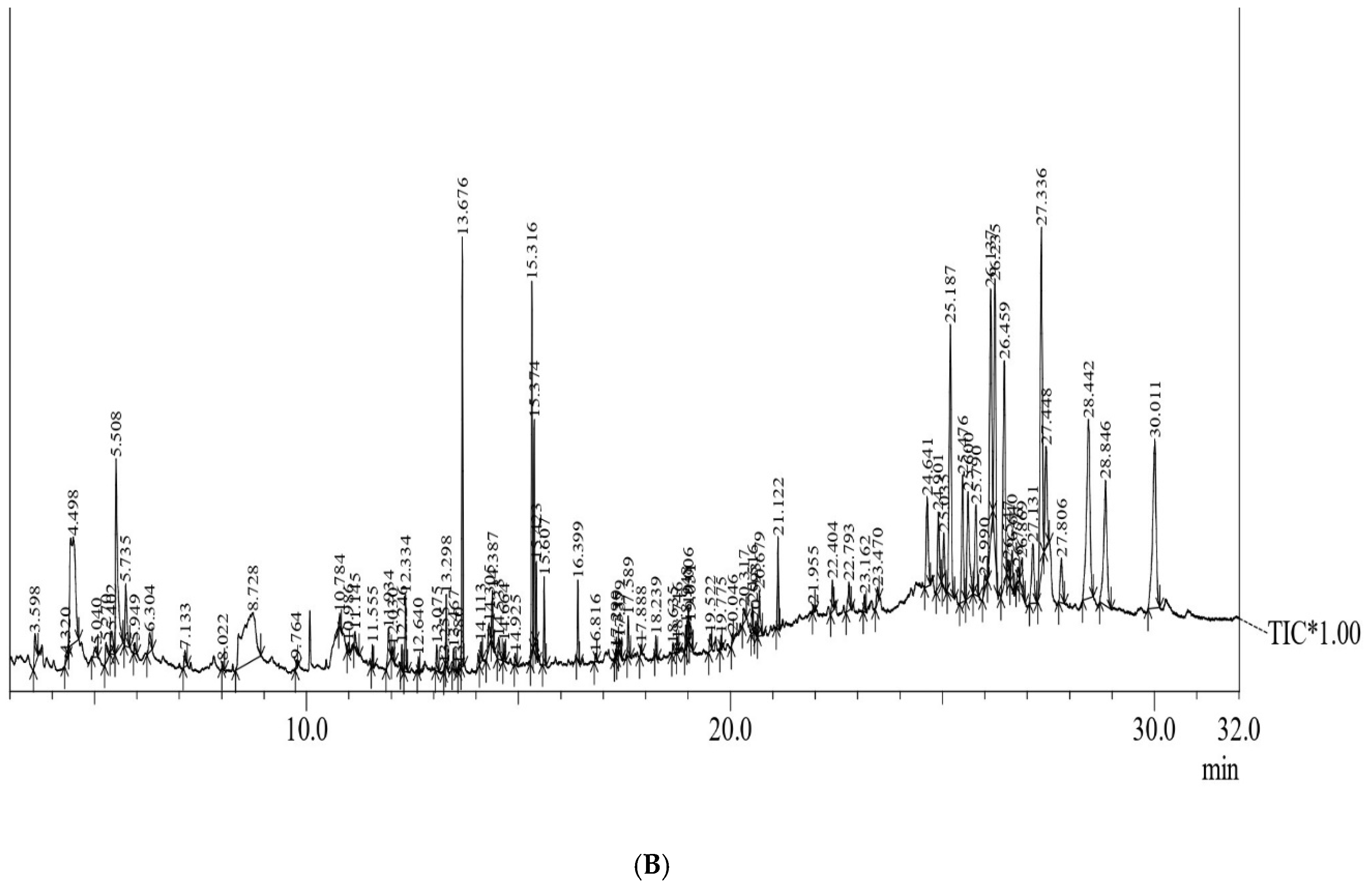
25]. In the current study, phytochemical profiling with GC–MS of methanolic extracts (
Figure 5) of anther and anther-derived callus of
C. roseus has been conducted. The results obtained show the presence of various phytoconstituents, including carbohydrates, alkaloids, phenols, saponins, phytosterols, terpenoids, steroids, etc. A total of 14 bioactives are common in both the extracted samples. However, there are compounds that are exclusive to each sample that confer various biological properties to this plant. The presence of secondary metabolites in callus, which are otherwise not detected in anther tissue, may be due to the fact that certain bioactive compounds accumulate in specific cells or tissues or in a specific growth stage (mostly the stationary phase) of in vitro cultures [
26]. Therefore, developing callus from different tissues to obtain therapeutically active compounds is of high significance.
The major compounds of medicinal value present in the methanolic extract of anthers were squalene (triterpene), alpha-amyrin (triterpene), coronaridine (alkaloid) and cedrol (essential oil), which possess anti-oxidant, gastroprotective and hepatoprotective, anti-cancerous and anti-inflammatory properties, respectively [
27,
28,
29,
30]. Similarly, in anther-derived calli exclusively, 17 compounds are present having diverse medicinal properties, and these compounds are listed in
Table 5. These include stearic acid, linoleic acid, oleic acid, vindolinine, pleiocarpamine, pericyclivine, ajmalicine, 19-epiajmalicine, beta-stigmasterol, cycloartenol, etc. Ajmalicine and vindolinine are well-known alkaloids having anti-cancerous, anti-hypertensive and anti-oxidant properties [
15,
31]. Recently, an alkaloid named pleiocarpamine has been isolated from the stem bark of
Rauvolfia caffra and is reported to possess anti-seizure activity [
32]. Cycloartenol (a triterpenoid) and stigmasterol (a sterol) have also been detected in present studies and are associated with immunosuppressive, anti-hypercholestrolemic and anti-inflammatory activities, respectively [
33,
34]. Compounds such as cycloartenol, ajmalicine, vindolinine, pleiocarpamine and pericyclivine have been reported previously in leaf tissues of
C. roseus [
35,
36]. Some reports of phytocompounds identified from different tissues using GC–MS were noted earlier [
37,
38], but till now, no information on the phytocompounds present in anther or anther-derived callus was available for
C. roseus.
The ploidy status of anther-derived callus was checked using flow cytometry, and the results show that the ploidy of the calli was haploid in nature, confirming the involvement of microspores in developing callus. Similar observations have also been reported for other plant species [
7,
10,
39]. FCM is the widely used approach for determining the ploidy of plants developed through callus, somatic embryos and other in vitro-regenerated pathways [
40]. The origin of diploid plants from anthers may be due to the involvement of other somatic cells such as anther wall, filament or flower septum in developing callus. Spontaneous chromosomal doubling can also be a mechanism in the generation of polyploidy in anther-derived regenerants. In certain cases, mixoploids and aneuploids have also been noted in anther cultures of different plants [
8,
41], but these polyploids were not detected in this experiment. This is the first-ever report of GC–MS analysis of medically significant compounds from anther tissue of
C. roseus, which enriches the phytocompound library of
Catharanthus and may be utilized in the pharmaceutical and industrial sectors.
4. Materials and Methods
4.1. Anther Culture and Growth Conditions
The mature flowers of
C. roseus were collected from the herbal garden, Jamia Hamdard, New Delhi, and the anthers were used as explants for experimentations. The surface sterilization of flowers was performed following the method of Bansal et al. [
3] described earlier. The sterilized anthers were excised from the flowers and aseptically cultured onto agar-solidified basal Murashige and Skoog (MS) medium supplemented with various concentrations and combinations of plant growth regulators (PGRs) and sub-cultured every 3–4 weeks. The cultures were incubated at a temperature of 24 ± 2 °C with 48 μmol/m
2/s
2 illumination (white fluorescent light) for a 16 h photoperiod.
4.2. Callus Induction and Proliferation
The disinfected anthers were inoculated on MS augmented with different concentrations (alone or in combination) of α-naphthalene acetic acid (NAA), kinetin (Kn) and thidiazuron (TDZ) ranging from 0.1 to 1.0 mg/L for callus induction. Callus formation started within 14–16 days of culture and proliferated on the same medium with successive subculturing. The callus induction frequency and the callus fresh weight were recorded after 6 weeks of culture.
4.3. Surface Morphology and Elemental Analysis
The surface morphology and elemental profile of anther and anther-derived callus were determined using energy-dispersive X-ray microanalysis (EDX) combined with scanning electron microscopy (SEM). For this purpose, the samples were primarily fixed with Karnovsky’s fixative and washed with 0.1 M phosphate buffer at 4 °C. Afterward, a series of dehydrations with acetone (30%, 50%, 70%, 90% and 100%) were performed at 15 min intervals, and then critical-point drying was performed at 1100 p.s.i. These samples were then mounted on aluminum stubs and sputter-coated with gold having a 35 nm thick film. Finally, the coated samples were viewed at an accelerating voltage of 20 kV under a scanning electron microscope (Zeiss, Oberkochen, Germany) equipped with EDAX.
4.4. Preparation of Extracts
The methanolic extracts of both samples were prepared according to the protocol of Hussain et al. [
42]. About 1.0 g of anther and anther callus were shade dried and crushed into fine powder using mortar and pestle (
Figure 3A,B). Each sample was then extracted in 5.0 mL methanol in an orbital shaker for 48 h. Afterward, the extracts were filtered through Whatman filter paper no. 1 and evaporated to dryness. The obtained extracts were stored in an airtight container with proper labeling at 4 °C for further use (
Figure 3C).
4.5. GC–MS Analysis
GC–MS analyses of these extracts were conducted on GC–MS QP-2010 equipment (Shimadzu, Japan) at Advanced Instrumentation Research Facility (AIRF), JNU, New Delhi. The program settings were as follows: Helium was used as a carrier gas (1 mL/min), and the initial and final temperatures were programmed at 100 °C and 260 °C, respectively, with a hold time of 18 min. Ion source temperature was 220 °C with an interface temperature of 270 °C and solvent cut time of 2.5 min. Other specifications included: detector gain mode relative to the tuning result, detector gain +0.00 kV, threshold of 1000, start time 3 min, end time 39.98 min, event time 0.3 s, scan speed of 2000, start m/z 40.00 and end m/z 600.00.
4.6. Metabolite Data Processing and Analysis
The bioactive compounds were identified using the mass spectral database of the NIST17 library. The unknown compounds’ spectra were compared with the known phytocompound spectra available in the NIST library, and the name, molecular weight and structure of the compounds were determined.
4.7. Flow Cytometric Analysis
The ploidy status of anther-derived calli was examined using the flow cytometry method as described by Galbraith [
43]. A total of 3 samples of anther-derived callus were randomly chosen, along with a reference standard of diploid leaves of
C. roseus with a known 2C DNA content of 1.51 pg [
44]. Approximately 50 mg of callus was added to a Petri plate having 1.0 mL ice-cold Galbraith’s buffer (nuclei isolation buffer) and finely macerated with the help of a surgical blade. The homogenate was then filtered with a 100 µm nylon mesh to eliminate larger cellular remnants and was finally stained with 50 µg/mL PI RNase (propidium iodide RNase) (Sigma-Aldrich, St. Louis, MO, USA) for 8–10 min. The samples were incubated in the dark at 4 °C for about 40 min and eventually examined on a BD FACS(Calibur) flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA). The relative nuclear DNA of anther-derived callus of
C. roseus was estimated using the below formula [
45]:
4.8. Statistical Analysis
In the tissue-culture experiment, three explants (anthers) per culture tube were inoculated with five replicates of every experimental treatment, and each experiment was repeated twice. The data are expressed as mean ± standard error, and the analysis was performed using one-way analysis of variance (ANOVA). The significance of mean difference was determined using Duncan’s multiple range test (DMRT) at
p < 0.05 using SPSS Ver. 26.0 (SPSS Inc., Chicago, IL, USA) [
46]. The flow cytometric study was repeated thrice with randomly chosen standard (donor plant) and callus samples.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}