
Synergistic Effects of Kaolin and Silicon Nanoparticles for Ameliorating Deficit Irrigation Stress in Maize Plants by Upregulating Antioxidant Defense Systems

 , , ,
, , ,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
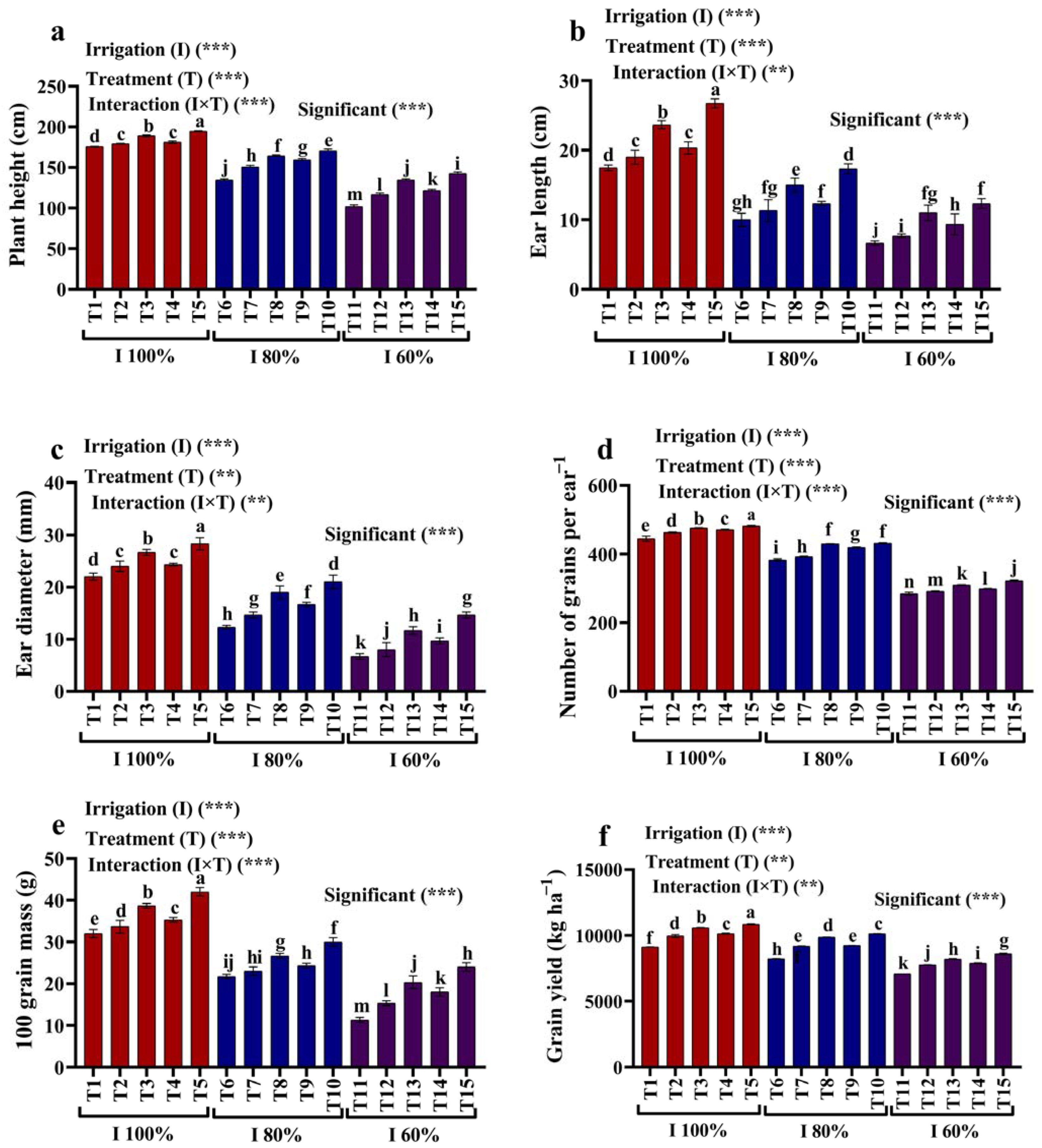
2.1. Growth and Yield Traits
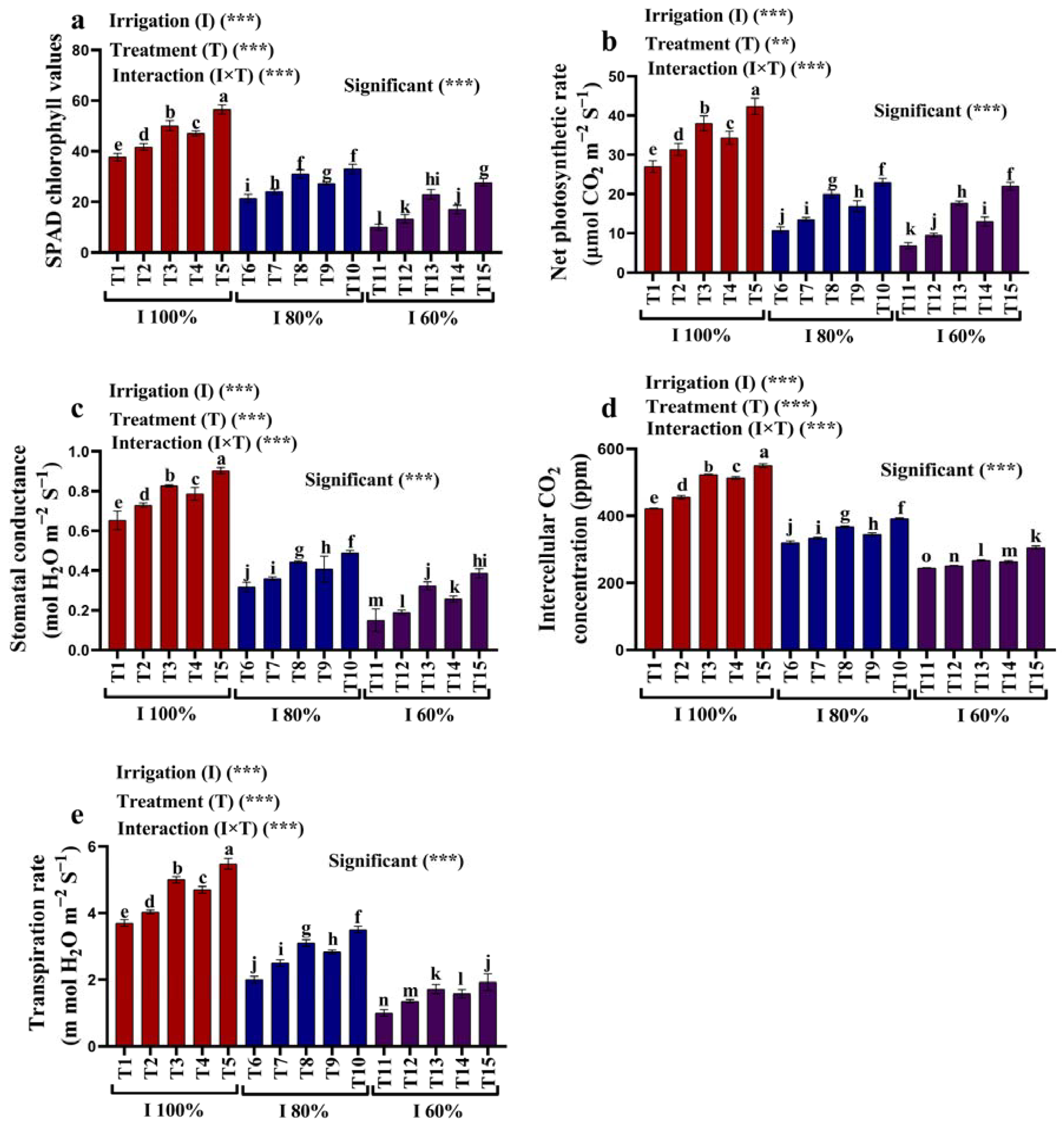
2.2. Photosynthetic Characteristics
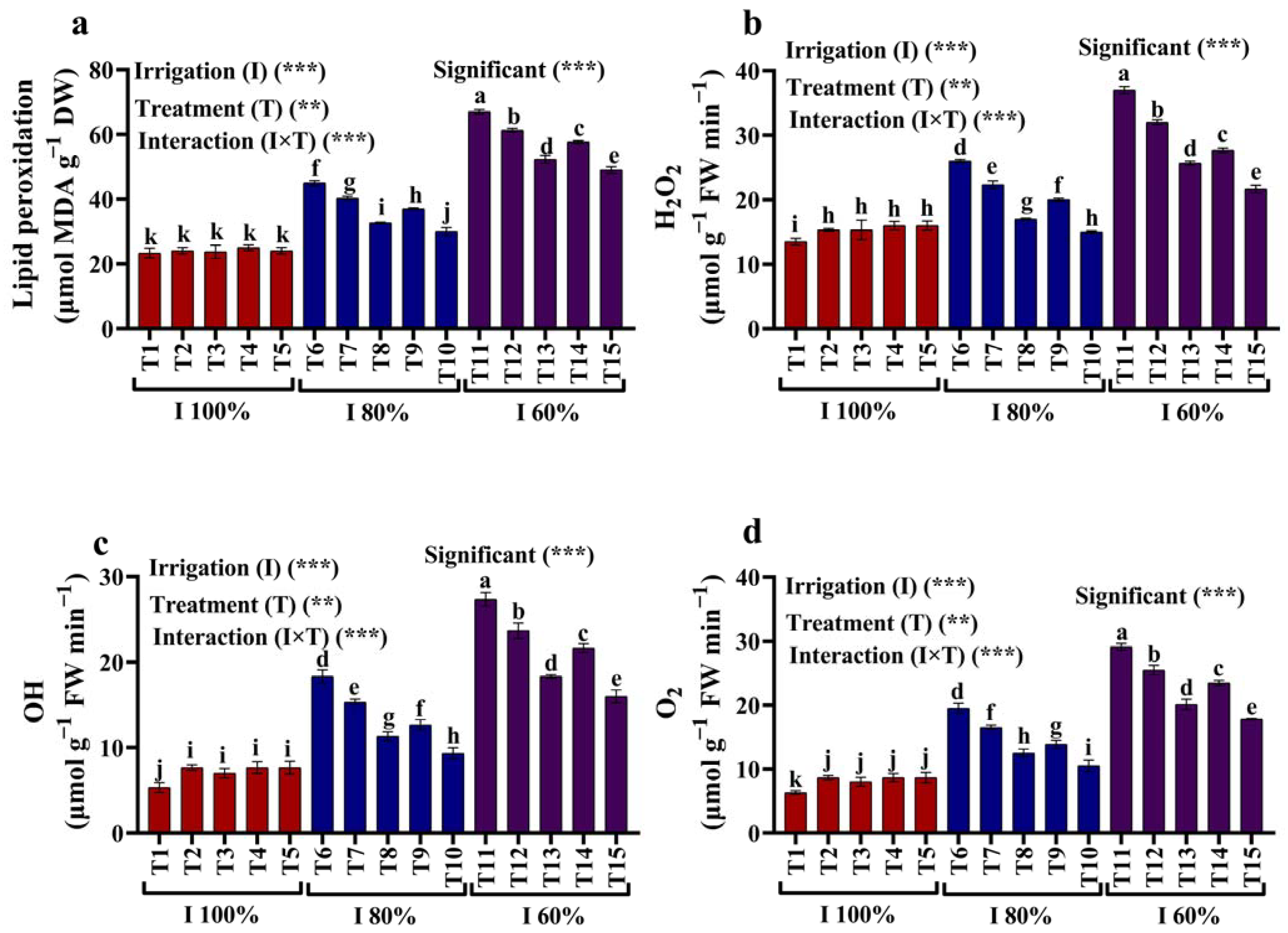
2.3. Lipid Peroxidation Content and ROS Production
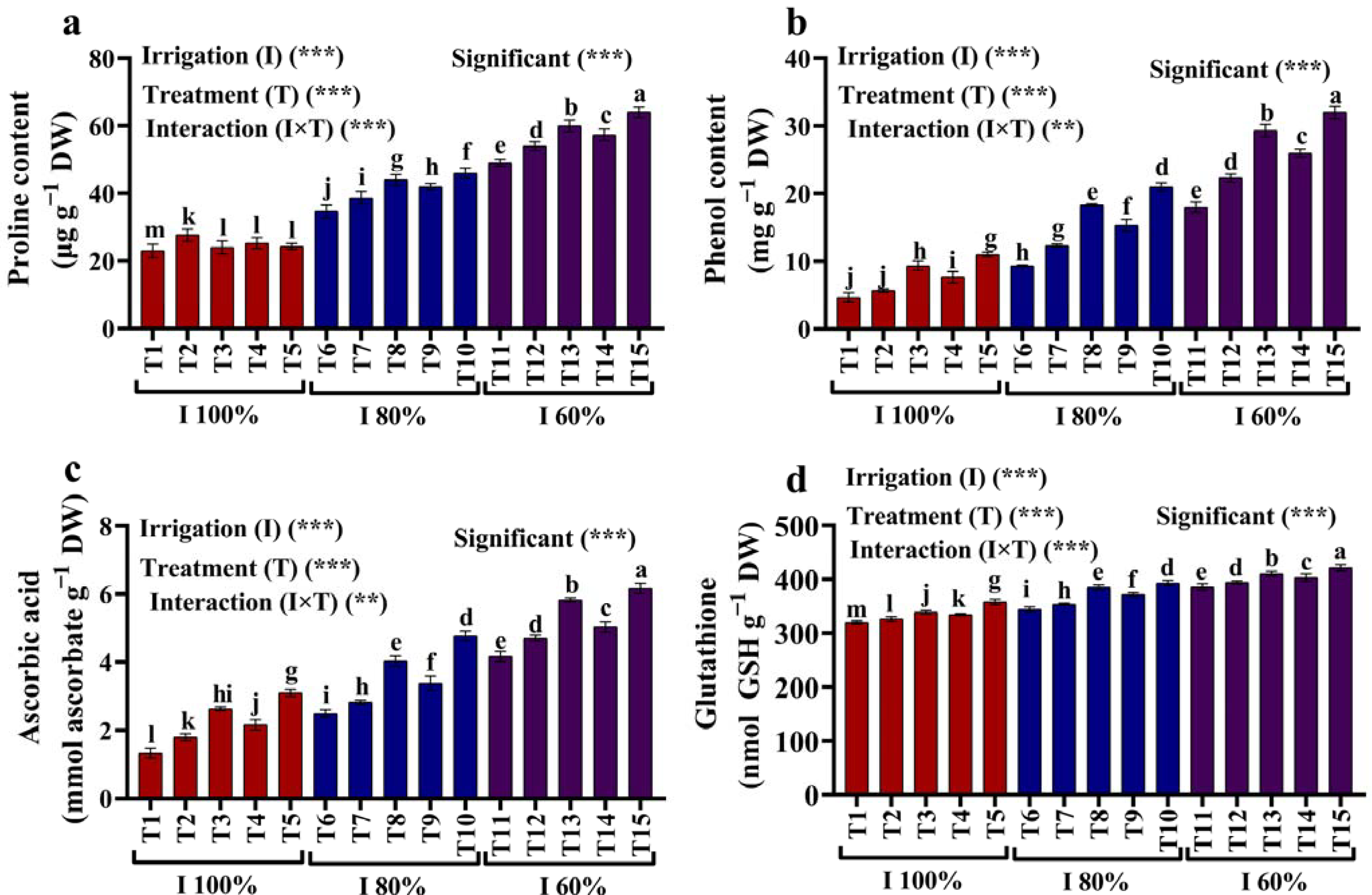
2.4. Proline, Phenol, and Non-Enzymatic Antioxidants
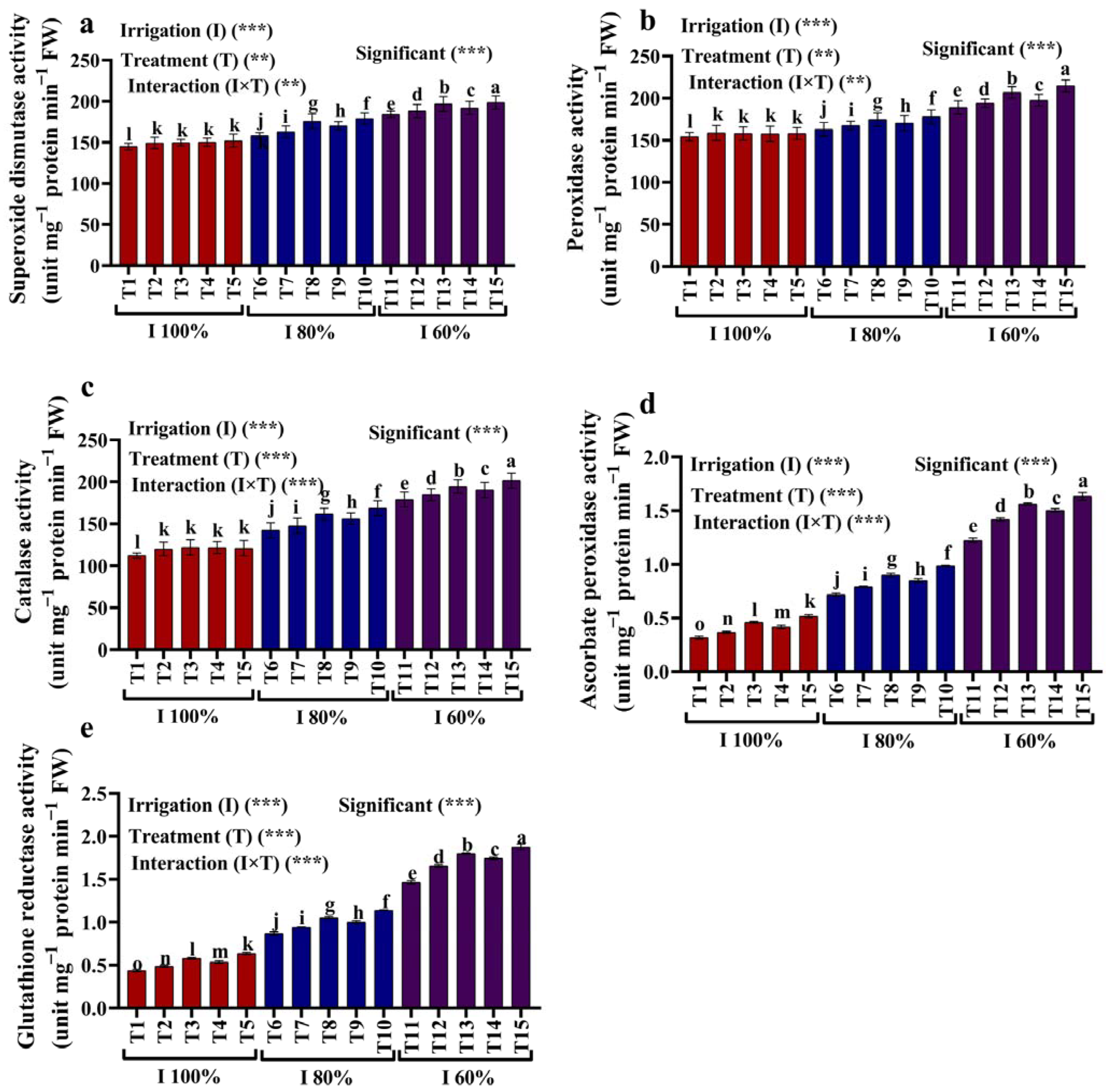
2.5. Antioxidant Enzymes
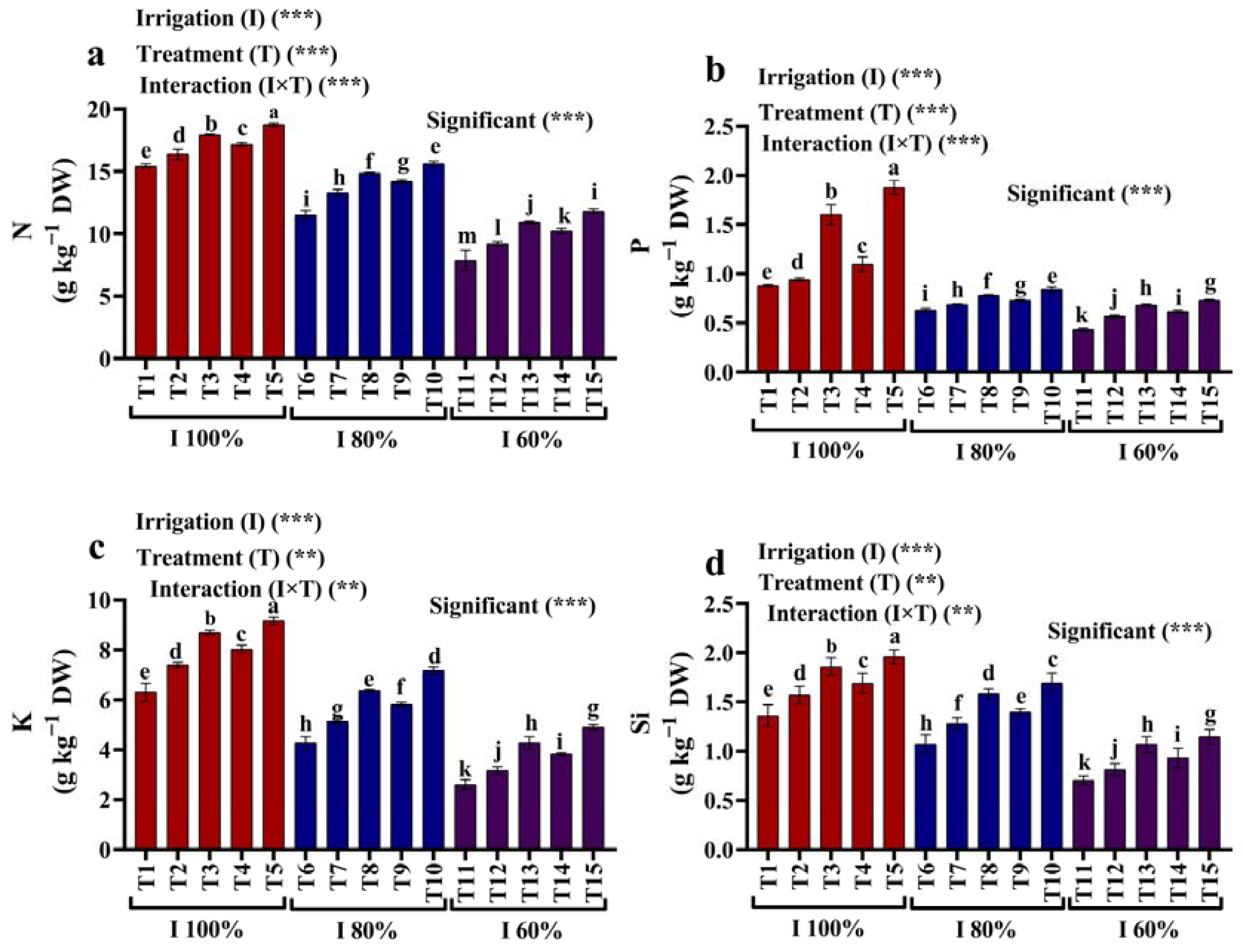
2.6. Mineral Content
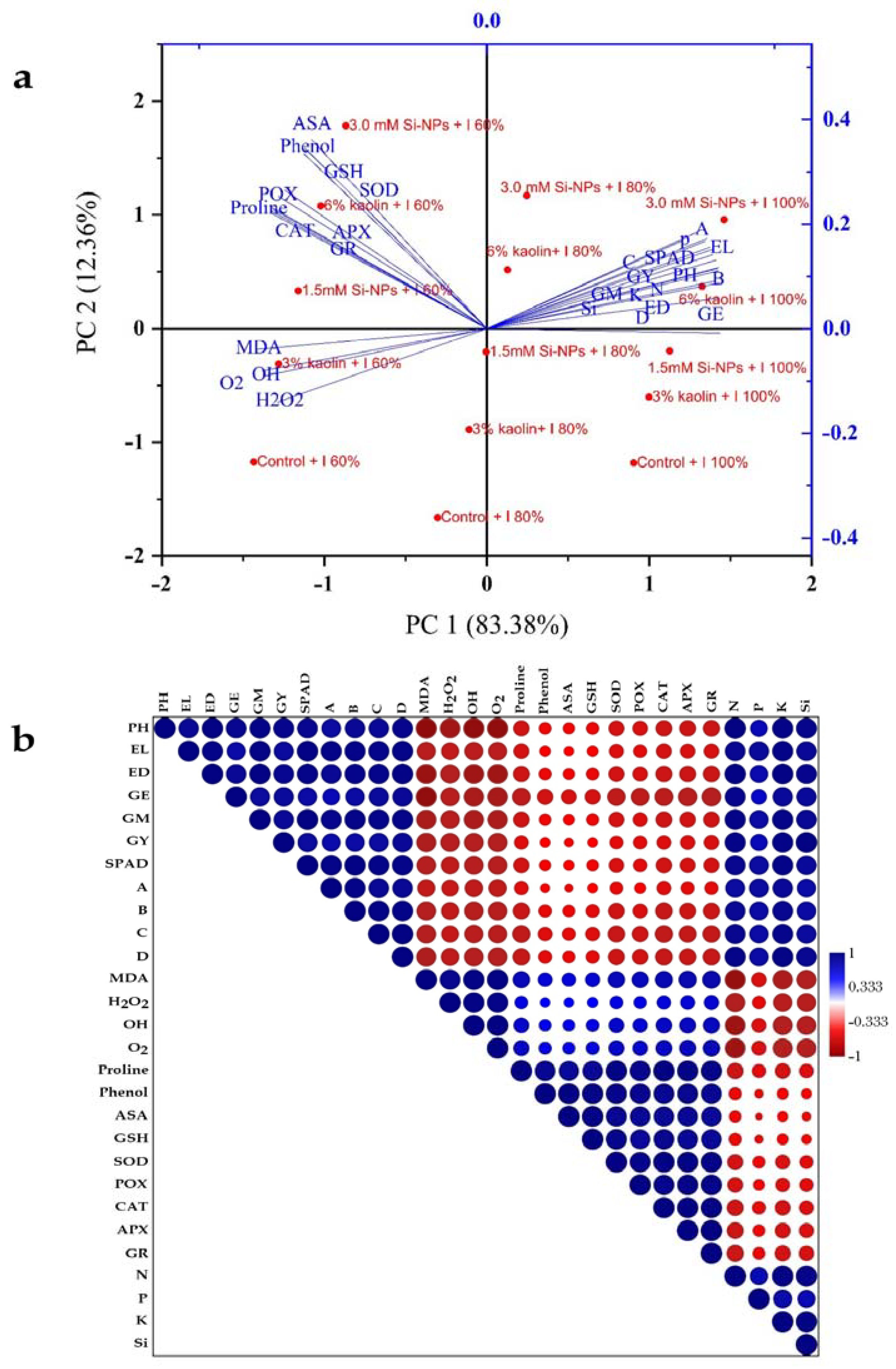
2.7. A Principal Component Analysis and Heat Map PeA.R.S.on Correlation
3. Materials and Methods
- Chemicals:
- Procedure:
- General Properties:
- Characterization:
3.1. Plant Materials and Agricultural Practices
3.2. Irrigation Water Applied (IWA)
| Group I | |
| T1 | Full irrigation with 100% of AW. |
| T2 | Full irrigation with 100% of AW + foliar application of 3% kaolin. |
| T3 | Full irrigation with 100% of AW + foliar application of 6% kaolin. |
| T4 | Full irrigation with 100% of AW + foliar application of 1.5 mM SiO2 NPs. |
| T5 | Full irrigation with 100% of AW + foliar application of 3.0 mM SiO2 NPs. |
| Group 2 | |
| T6 | Irrigation with 80% of AW. |
| T7 | Irrigation with 80% of AW + foliar application of 3% kaolin. |
| T8 | Irrigation with 80% of AW + foliar application of 6% kaolin. |
| T9 | Irrigation with 80% of AW + foliar application of 1.5 mM SiO2 NPs. |
| T10 | Irrigation with 80% of AW + foliar application of 3.0 mM SiO2 NPs. |
| Group 3 | |
| T11 | Irrigation with 60% of AW. |
| T12 | Irrigation with 60% of AW + foliar application of 3% kaolin. |
| T13 | Irrigation with 60% of AW + foliar application of 6% kaolin. |
| T14 | Irrigation with 60% of AW + foliar application of 1.5 mM SiO2 NPs. |
| T15 | Irrigation with 60% of AW + foliar application of 3.0 mM SiO2 NPs. |
3.3. Measurements
3.3.1. Growth and Yield Traits
3.3.2. Photosynthetic Characteristics
3.3.3. ROS Indicators
Lipid Peroxidation
Hydrogen Peroxide
Superoxide Anion
Hydroxyl Radical
3.3.4. Total Proline and Phenol Content and Non-Antioxidant Enzymes
3.3.5. Determination of Antioxidant Enzymes
3.3.6. Determination of Mineral Contents in Grains
3.4. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pawlak, K.; Kołodziejczak, M. The role of agriculture in ensuring food security in developing countries: Considerations in the context of the problem of sustainable food production. Sustainability 2020, 12, 5488. [Google Scholar] [CrossRef]
- Rosa, L.; Chiarelli, D.D.; Rulli, M.C.; Dell’Angelo, J.; D’Odorico, P. Global agricultural economic water scarcity. Sci. Adv. 2020, 6, eaaz6031. [Google Scholar] [CrossRef] [PubMed]
- Wudil, A.H.; Usman, M.; Rosak-Szyrocka, J.; Pilař, L.; Boye, M. Reversing Years for Global Food Security: A Review of the Food Security Situation in Sub-Saharan Africa (SSA). Int. J. Environ. Res. Public Health 2022, 19, 14836. [Google Scholar] [CrossRef]
- Aires, E.S.; Ferraz, A.K.L.; Carvalho, B.L.; Teixeira, F.P.; Putti, F.F.; de Souza, E.P.; Rodrigues, J.D.; Ono, E.O. Foliar application of salicylic acid to mitigate water stress in tomato. Plants 2022, 11, 1775. [Google Scholar] [CrossRef]
- Kapoor, D.; Bhardwaj, S.; Landi, M.; Sharma, A.; Ramakrishnan, M.; Sharma, A. The impact of drought in plant metabolism: How to exploit tolerance mechanisms to increase crop production. Appl. Sci. 2020, 10, 5692. [Google Scholar] [CrossRef]
- Dawood, M.F.; Abu-Elsaoud, A.M.; Sofy, M.R.; Mohamed, H.I.; Soliman, M.H. Appraisal of kinetin spraying strategy to alleviate the harmful effects of UVC stress on tomato plants. Environ. Sci. Pollut. Res. 2022, 29, 52378–52398. [Google Scholar] [CrossRef] [PubMed]
- El-Sheshtawy, H.S.; Mahdy, H.M.; Sofy, A.R.; Sofy, M.R. Production of biosurfactant by Bacillus megaterium and its correlation with lipid peroxidation of Lactuca sativa. Egypt. J. Pet. 2022, 31, 1–6. [Google Scholar] [CrossRef]
- Dawood, M.F.; Sofy, M.R.; Mohamed, H.I.; Sofy, A.R.; Abdel-kader, H.A. Hydrogen sulfide modulates salinity stress in common bean plants by maintaining osmolytes and regulating nitric oxide levels and antioxidant enzyme expression. J. Soil Sci. Plant Nutr. 2022, 22, 3708–3726. [Google Scholar] [CrossRef]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant survival under drought stress: Implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef]
- Al-Mokadem, A.Z.; Alnaggar, A.E.-A.M.; Mancy, A.G.; Sofy, A.R.; Sofy, M.R.; Mohamed, A.K.S.; Abou Ghazala, M.M.; El-Zabalawy, K.M.; Salem, N.F.; Elnosary, M.E. Foliar Application of Chitosan and Phosphorus Alleviate the Potato virus Y-Induced Resistance by Modulation of the Reactive Oxygen Species, Antioxidant Defense System Activity and Gene Expression in Potato. Agronomy 2022, 12, 3064. [Google Scholar] [CrossRef]
- Hanif, S.; Saleem, M.F.; Sarwar, M.; Irshad, M.; Shakoor, A.; Wahid, M.A.; Khan, H.Z. Biochemically triggered heat and drought stress tolerance in rice by proline application. J. Plant Growth Regul. 2021, 40, 305–312. [Google Scholar] [CrossRef]
- Krishnan, S.R.; Nallakaruppan, M.; Chengoden, R.; Koppu, S.; Iyapparaja, M.; Sadhasivam, J.; Sethuraman, S. Smart Water Resource Management Using Artificial Intelligence—A Review. Sustainability 2022, 14, 13384. [Google Scholar] [CrossRef]
- García-Vila, M.; Fereres, E.; Mateos, L.; Orgaz, F.; Steduto, P. Deficit irrigation optimization of cotton with AquaCrop. Agron. J. 2009, 101, 477–487. [Google Scholar] [CrossRef]
- Dinis, L.-T.; Bernardo, S.; Matos, C.; Malheiro, A.; Flores, R.; Alves, S.; Costa, C.; Rocha, S.; Correia, C.; Luzio, A. Overview of Kaolin Outcomes from vine to wine: Cerceal white variety case study. Agronomy 2020, 10, 1422. [Google Scholar] [CrossRef]
- Sharma, S.; Singh, S.; Bahuguna, A.; Yadav, B.; Barthwal, A.; Nandan, R.; Khatana, R.; Pandey, A.; Thakur, R.; Singh, H. Nanotechnology: An Efficient Tool in Plant Nutrition Management. Ecosystem Services: Types, Management and Benefits; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2022. [Google Scholar]
- Mohamed, H.I.; Abd-Elsalam, K.A.; Tmam, A.M.; Sofy, M.R. Silver-based nanomaterials for plant diseases management: Today and future perspectives. In Silver Nanomaterials for Agri-Food Applications; Elsevier: Amsterdam, The Netherlands, 2021; pp. 495–526. [Google Scholar]
- Albalawi, M.A.; Abdelaziz, A.M.; Attia, M.S.; Saied, E.; Elganzory, H.H.; Hashem, A.H. Mycosynthesis of Silica Nanoparticles Using Aspergillus niger: Control of Alternaria solani Causing Early Blight Disease, Induction of Innate Immunity and Reducing of Oxidative Stress in Eggplant. Antioxidants 2022, 11, 2323. [Google Scholar] [CrossRef] [PubMed]
- Hajizadeh, H.S.; Azizi, S.; Rasouli, F.; Okatan, V. Modulation of physiological and biochemical traits of two genotypes of Rosa damascena Mill. by SiO2-NPs under In vitro drought stress. BMC Plant Biol. 2022, 22, 538. [Google Scholar] [CrossRef]
- Fouda, H.; Sofy, M. Effect of biological synthesis of nanoparticles from Penicillium chrysogenum as well as traditional salt and chemical nanoparticles of zinc on canola plant oil productivity and metabolic. Egypt. J. Chem. 2022, 65, 507–516. [Google Scholar] [CrossRef]
- Babu, S.; Singh, R.; Yadav, D.; Rathore, S.S.; Raj, R.; Avasthe, R.; Yadav, S.; Das, A.; Yadav, V.; Yadav, B. Nanofertilizers for agricultural and environmental sustainability. Chemosphere 2022, 292, 133451. [Google Scholar] [CrossRef]
- Akhtar, N.; Ilyas, N.; Hayat, R.; Yasmin, H.; Noureldeen, A.; Ahmad, P. Synergistic effects of plant growth promoting rhizobacteria and silicon dioxide nano-particles for amelioration of drought stress in wheat. Plant Physiol. Biochem. 2021, 166, 160–176. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Hosseini, M.S.; Hoveizeh, N.F.; Kadkhodaei, S.; Vaculík, M. Comparative morphological, physiological and molecular analyses of drought-stressed strawberry plants affected by SiO2 and SiO2-NPs foliar spray. Sci. Hortic. 2023, 309, 111686. [Google Scholar] [CrossRef]
- Rai-Kalal, P.; Tomar, R.S.; Jajoo, A. Seed nanopriming by silicon oxide improves drought stress alleviation potential in wheat plants. Funct. Plant Biol. 2021, 48, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Sutulienė, R.; Ragelienė, L.; Samuolienė, G.; Brazaitytė, A.; Urbutis, M.; Miliauskienė, J. The response of antioxidant system of drought-stressed green pea (Pisum sativum L.) affected by watering and foliar spray with silica nanoparticles. Horticulturae 2022, 8, 35. [Google Scholar] [CrossRef]
- Elisa, D.-H.; Marcela, G.-M.; Alejandra, G.-U.J.; Elena, D.-H.M. The nutraceutical value of maize (Zea mays L.) landraces and the determinants of its variability: A review. J. Cereal Sci. 2022, 103, 103399. [Google Scholar] [CrossRef]
- Sabagh, A.; Çiğ, F.; Seydoşoğlu, S.; Battaglia, M.L.; Javed, T.; Iqbal, M.A.; Awad, M. Salinity stress in maize: Effects of stress and recent developments of tolerance for improvement. Cereal Grains 2021, 1, 213. [Google Scholar]
- Aman, M. Genetic Variability, Heritability and Association of Quantitative Traits in Maize (Zea mays L.) Genotypes. Bioinformatics 2021, 25, 534–536. [Google Scholar] [CrossRef]
- Teixeira, M.; Feijão, E.; Catarino, L.; Matos, A.R.; Figueiredo, A.; Marques da Silva, J. Exploring local maize diversity for increased agricultural sustainability: New insights into drought stress response and recovery of Guinea-Bissau landraces. Sustainability 2021, 13, 5441. [Google Scholar] [CrossRef]
- Jafarzadeh, M.; Rahman, I.; Sipaut, C. Synthesis of silica nanoparticles by modified sol–gel process: The effect of mixing modes of the reactants and drying techniques. Sol-Gel Sci. Technol. 2009, 50, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Page, A.L.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties, 2nd ed.; Amen Society of Agronomy, Inc.: Madison, WI, USA, 1982. [Google Scholar]
- Klute, A.; Dirksen, C. Hydraulic conductivity and diffusivity: Laboratory methods. Methods Soil Anal. Part 1 Phys. Mineral. Methods 1986, 5, 687–734. [Google Scholar]
- Badawy, I.H.; Hmed, A.A.; Sofy, M.R.; Al-Mokadem, A.Z. Alleviation of cadmium and nickel toxicity and phyto-stimulation of tomato plant l. by endophytic micrococcus luteus and enterobacter cloacae. Plants 2022, 11, 2018. [Google Scholar] [CrossRef]
- Hernández, J.A.; Almansa, M.S. Short-term effects of salt stress on antioxidant systems and leaf water relations of pea leaves. Physiol. Plant. 2002, 115, 251–257. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Jabs, T.; Dietrich, R.A.; Dangl, J.L. Initiation of runaway cell death in an Arabidopsis mutant by extracellular superoxide. Science 1996, 273, 1853–1856. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.; Aruoma, O.I. The deoxyribose method: A simple “test-tube” assay for determination of rate constants for reactions of hydroxyl radicals. Anal. Biochem. 1987, 165, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.; Waldren, R.A.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ziouti, A.; El Modafar, C.; Fleuriet, A.; El Boustani, E. Les polyphenols, marqueurs potentiels de la resistance du palmier dattier (Phoenix dactylifera L.) au Fausarium oxysporum f. sp. albedinis. Bull. Liaison-Groupe Polyphen. 1992, 16, 346. [Google Scholar]
- Jagota, S.; Dani, H. A new colorimetric technique for the estimation of vitamin C using Folin phenol reagent. Analyt Biochem 1982, 127, 178–182. [Google Scholar] [CrossRef]
- Owens, C.; Belcher, R. A colorimetric micro-method for the determination of glutathione. J. Biochem. 1965, 94, 705. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Taniguchi, N.; Maeda, K.; Kitano, M. On ‘Generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase’ by Yasuhisa Kono. Arch. Biochem. Biophys. 2022, 726, 109115. [Google Scholar] [CrossRef]
- Thomas, R.L.; Jen, J.J.; Morr, C.V. Changes in soluble and bound peroxidase—IAA oxidase during tomato fruit development. J. Food Sci. 1982, 47, 158–161. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Jiang, M.; Zhang, J. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Experim. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bremner, J. Determination of nitrogen in soil by the Kjeldahl method. J. Agric. Sci. 1960, 55, 11–33. [Google Scholar] [CrossRef]
- Houba, V.; Uittenbogaard, J.; Pellen, P. Wageningen evaluating programmes for analytical laboratories (WEPAL), organization and purpose. Commun. Soil Sci. Plant Anal. 1996, 27, 421–431. [Google Scholar] [CrossRef]
- Eraslan, F.; Inal, A.; Pilbeam, D.J.; Gunes, A. Interactive effects of salicylic acid and silicon on oxidative damage and antioxidant activity in spinach (Spinacia oleracea L. cv. Matador) grown under boron toxicity and salinity. Plant Growth Regul. 2008, 55, 207–219. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons: Hoboken, NJ, USA, 1984. [Google Scholar]
- Ahmadi, S.H.; Solgi, S.; Sepaskhah, A.R. Quinoa: A super or pseudo-super crop? Evidences from evapotranspiration, root growth, crop coefficients, and water productivity in a hot and semi-arid area under three planting densities. Agric. Water Manag. 2019, 225, 105784. [Google Scholar] [CrossRef]
- Haider, I.; Raza, M.A.S.; Iqbal, R.; Aslam, M.U.; Habib-ur-Rahman, M.; Raja, S.; Khan, M.T.; Aslam, M.M.; Waqas, M.; Ahmad, S. Potential effects of biochar application on mitigating the drought stress implications on wheat (Triticum aestivum L.) under various growth stages. J. Saudi Chem. Soc. 2020, 24, 974–981. [Google Scholar] [CrossRef]
- Humplík, J.F.; Bergougnoux, V.; Van Volkenburgh, E. To stimulate or inhibit? That is the question for the function of abscisic acid. Trends Plant Sci. 2017, 22, 830–841. [Google Scholar] [CrossRef]
- Zhao, H.; Qu, S.; Guo, S.; Zhao, H.; Liang, S.; Xu, M. Virtual water scarcity risk to global trade under climate change. J. Clean. Prod. 2019, 230, 1013–1026. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Selwey, W.A.; Ibrahim, A.A.; Shady, M.; Alsadon, A.A. Foliar applications of ZnO and SiO2 nanoparticles mitigate water deficit and enhance potato yield and quality traits. Agronomy 2023, 13, 466. [Google Scholar] [CrossRef]
- Wang, X.; Vignjevic, M.; Liu, F.; Jacobsen, S.; Jiang, D.; Wollenweber, B. Drought priming at vegetative growth stages improves tolerance to drought and heat stresses occurring during grain filling in spring wheat. Plant Growth Regul. 2015, 75, 677–687. [Google Scholar] [CrossRef]
- Hassanpour, H.; Eydi, A.; Hekmati, M. Electromagnetic field improved nanoparticle impact on antioxidant activity and secondary metabolite production in Anthemis gilanica seedlings. Int. J. Agron. 2021, 2021, 8730234. [Google Scholar] [CrossRef]
- Sofy, M.R.; Mancy, A.G.; Alnaggar, A.E.A.M.; Refaey, E.E.; Mohamed, H.I.; Elnosary, M.E.; Sofy, A.R. A polishing the harmful effects of Broad Bean Mottle Virus infecting broad bean plants by enhancing the immunity using different potassium concentrations. Not. Bot. Horti Agrobot. Cluj-Napoca 2022, 50, 12654. [Google Scholar] [CrossRef]
- El–Bially, M.E.; Saudy, H.S.; Hashem, F.A.; El–Gabry, Y.A.; Shahin, M.G. Salicylic acid as a tolerance inducer of drought stress on sunflower grown in sandy soil. Gesunde Pflanz. 2022, 74, 603–613. [Google Scholar] [CrossRef]
- Jańczak-Pieniążek, M.; Migut, D.; Piechowiak, T. The Effect of Exogenous Copper–Quercetin Complex on Wheat (Triticum aestivum L.) Seedlings Growth under Drought Stress. Chem. Proc. 2022, 10, 18. [Google Scholar]
- Time, A.; Garrido, M.; Acevedo, E. Water relations and growth response to drought stress of Prosopis tamarugo Phil. A review. J. Soil Sci. Plant Nutr. 2018, 18, 329–343. [Google Scholar] [CrossRef]
- Agha, M.S.; Haroun, S.A.; Abbas, M.A.; Sofy, M.R.; Mowafy, A.M. Growth and Metabolic Response of Glycine max to the Plant Growth-Promoting Enterobacter Delta PSK and Bradyrhizobium japonicum Under Salinity Stress. J. Plant Growth Regul. 2023, 1–15. [Google Scholar] [CrossRef]
- El-Ramady, H.; Verma, K.K.; Rajput, V.D.; Minkina, T.; Elbehery, F.; Elbasiony, H.; Elsakhawy, T.; Omara, A.E.-D.; Amer, M. Sources of silicon and nano-silicon in soils and plants. In Silicon and Nano-Silicon in Environmental Stress Management and Crop Quality Improvement; Elsevier: Amsterdam, The Netherlands, 2022; pp. 1–15. [Google Scholar]
- Bhattacharjee, R.; Kumar, L.; Mukerjee, N.; Anand, U.; Dhasmana, A.; Preetam, S.; Bhaumik, S.; Sihi, S.; Pal, S.; Khare, T. The emergence of metal oxide nanoparticles (NPs) as a phytomedicine: A two-facet role in plant growth, nano-toxicity and anti-phyto-microbial activity. Biomed. Pharmacother. 2022, 155, 113658. [Google Scholar] [CrossRef]
- Kataria, S.; Jain, M.; Rastogi, A.; Živčák, M.; Brestic, M.; Liu, S.; Tripathi, D.K. Role of nanoparticles on photosynthesis: Avenues and applications. In Nanomaterials in Plants, Algae and Microorganisms; Elsevier: Amsterdam, The Netherlands, 2019; pp. 103–127. [Google Scholar]
- Alagoz, S.M.; Arvanaghi, H.R.; Dolatabadi, N.; Khalaki, M.A.; Moameri, M.; Lajayer, B.A.; van Hullebusch, E.D. Impact on nutritional status of plants treated with nanoparticles. In Toxicity of Nanoparticles in Plants; Elsevier: Amsterdam, The Netherlands, 2022; pp. 333–358. [Google Scholar]
- Behboudi, F.; Tahmasebi Sarvestani, Z.; Kassaee, M.Z.; Modares Sanavi, S.; Sorooshzadeh, A. Improving growth and yield of wheat under drought stress via application of SiO2 nanoparticles. J. Agric. Sci. Technol. 2018, 20, 1479–1492. [Google Scholar]
- Song, Y.; Li, J.; Liu, M.; Meng, Z.; Liu, K.; Sui, N. Nitrogen increases drought tolerance in maize seedlings. Funct. Plant Biol. 2019, 46, 350–359. [Google Scholar] [CrossRef]
- AlKahtani, M.D.; Hafez, Y.M.; Attia, K.; Rashwan, E.; Husnain, L.A.; AlGwaiz, H.I.; Abdelaal, K.A. Evaluation of silicon and proline application on the oxidative machinery in drought-stressed sugar beet. Antioxidants 2021, 10, 398. [Google Scholar] [CrossRef]
- Li, J.; Meng, B.; Chai, H.; Yang, X.; Song, W.; Li, S.; Lu, A.; Zhang, T.; Sun, W. Arbuscular mycorrhizal fungi alleviate drought stress in C3 (Leymus chinensis) and C4 (Hemarthria altissima) grasses via altering antioxidant enzyme activities and photosynthesis. Front. Plant Sci. 2019, 10, 499. [Google Scholar] [CrossRef] [Green Version]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic stress and reactive oxygen species: Generation, signaling, and defense mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Salehi, H.; Rahman, M.A.; Zahid, Z.; Madadkar Haghjou, M.; Najafi-Kakavand, S.; Charagh, S.; Osman, H.S.; Albaqami, M.; Zhuang, Y. Plant hormones and neurotransmitter interactions mediate antioxidant defenses under induced oxidative stress in plants. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.F.; Sofy, M.R.; Mohamed, H.I.; Sofy, A.R.; Abdel-Kader, H.A. N-or/and P-deprived Coccomyxa chodatii SAG 216–2 extracts instigated mercury tolerance of germinated wheat seedlings. Plant Soil 2023, 483, 225–253. [Google Scholar] [CrossRef]
- Siddique, A.; Kandpal, G.; Kumar, P. Proline accumulation and its defensive role under diverse stress condition in plants: An overview. J. Pure Appl. Microbiol. 2018, 12, 1655–1659. [Google Scholar] [CrossRef]
- Wu, H.-H.; Zou, Y.-N.; Rahman, M.M.; Ni, Q.-D.; Wu, Q.-S. Mycorrhizas alter sucrose and proline metabolism in trifoliate orange exposed to drought stress. Sci. Rep. 2017, 7, 42389. [Google Scholar] [CrossRef] [Green Version]
- Abdelhamid, M.T.; Sekara, A.; Pessarakli, M.; Alarcón, J.; Brestic, M.; El-Ramady, H.; Gad, N.; Mohamed, H.I.; Fares, W.M.; Heba, S.S. New approaches for improving salt stress tolerance in rice. Rice Research for Quality Improvement: Genomics and Genetic Engineering. Breed. Techniq. Abiotic Stress Toleran. 2020, 247–268. [Google Scholar]
- Król, A.; Amarowicz, R.; Weidner, S. Changes in the composition of phenolic compounds and antioxidant properties of grapevine roots and leaves (Vitis vinifera L.) under continuous of long-term drought stress. Acta Physiol. Plant. 2014, 36, 1491–1499. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, S.; Sharma, D.; Singh, S.; Ramamurthy, P.C.; Verma, T.; Pujari, M.; Singh, J.; Kapoor, D.; Prasad, R. Physiological and molecular insights into the role of silicon in improving plant performance under abiotic stresses. Plant Soil 2022, 1–19. [Google Scholar] [CrossRef]
- Albqmi, M.; Selim, S.; Al-Sanea, M.M.; Alnusaire, T.S.; Almuhayawi, M.S.; Jaouni, S.K.A.; Hussein, S.; Warrad, M.; Sofy, M.R.; AbdElgawad, H. Interactive Effect of Arbuscular Mycorrhizal Fungi (AMF) and Olive Solid Waste on Wheat under Arsenite Toxicity. Plants 2023, 12, 1100. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.; Cheng, H.; Chen, L.; Zhu, H.; Mao, P.; Wang, M. Influence of exogenous ascorbic acid and glutathione priming on mitochondrial structural and functional systems to alleviate aging damage in oat seeds. BMC Plant Biol. 2020, 20, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Ziada, L.M.; Haroun, S.A.; Gamel, R.M.E.; Abbas, M.A.; Sofy, M.R.; El-Banna, H.Y. Secondary metabolites, hormonal homeostasis, and antioxidant enzymes of Moringa oleifera in response to white or violet Light Emitting Diodes (LEDs) combined with cytokinins under tissue culture conditions. Not. Bot. Horti Agrobot. Cluj-Napoca 2022, 50, 12879. [Google Scholar] [CrossRef]
- Mansoor, S.; Ali Wani, O.; Lone, J.K.; Manhas, S.; Kour, N.; Alam, P.; Ahmad, A.; Ahmad, P. Reactive oxygen species in plants: From source to sink. Antioxidants 2022, 11, 225. [Google Scholar] [CrossRef]
- Li, J.; Ran, X.; Zhou, M.; Wang, K.; Wang, H.; Wang, Y. Oxidative stress and antioxidant mechanisms of obligate anaerobes involved in biological waste treatment processes: A review. Sci. Total Environ. 2022, 838, 156454. [Google Scholar] [CrossRef]
- Ghasemi, A.; Farzaneh, S.; Moharramnejad, S.; Sharifi, R.S.; Youesf, A.F.; Telesinski, A.; Kalaji, H.M.; Mojski, J. Impact of 24-epibrassinolide, spermine, and silicon on plant growth, antioxidant defense systems, and osmolyte accumulation of maize under water stress. Sci. Rep. 2022, 12, 14648. [Google Scholar] [CrossRef]
- Mushtaq, N.; Iqbal, S.; Hayat, F.; Raziq, A.; Ayaz, A.; Zaman, W. Melatonin in Micro-Tom Tomato: Improved Drought Tolerance via the Regulation of the Photosynthetic Apparatus, Membrane Stability, Osmoprotectants, and Root System. Life 2022, 12, 1922. [Google Scholar] [CrossRef]
- Meitha, K.; Pramesti, Y.; Suhandono, S. Reactive oxygen species and antioxidants in postharvest vegetables and fruits. Int. J. Food Sci. 2020, 2020, 8817778. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Rahman, M.M.; Ansary, M.M.U.; Keya, S.S.; Abdelrahman, M.; Miah, M.G.; Phan Tran, L.-S. Silicon in mitigation of abiotic stress-induced oxidative damage in plants. Crit. Rev. Biotechnol. 2021, 41, 918–934. [Google Scholar] [CrossRef]
- Ulrich, K.; Jakob, U. The role of thiols in antioxidant systems. Free Radic. Biol. Med. 2019, 140, 14–27. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Al-Huqail, A.A.; Alqahtani, M.A.; Ahmad, P. Silicon is dependent on hydrogen sulphide to improve boron toxicity tolerance in pepper plants by regulating the AsA-GSH cycle and glyoxalase system. Chemosphere 2020, 257, 127241. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.F.; Zaid, A.; Latef, A.A.H.A. Salicylic acid spraying-induced resilience strategies against the damaging impacts of drought and/or salinity stress in two varieties of Vicia faba L. seedlings. J. Plant Growth Regul. 2022, 41, 1919–1942. [Google Scholar] [CrossRef]
- Alam, P.; Arshad, M.; Al-Kheraif, A.A.; Azzam, M.A.; Al Balawi, T. Silicon Nanoparticle-Induced Regulation of Carbohydrate Metabolism, Photosynthesis, and ROS Homeostasis in Solanum lycopersicum Subjected to Salinity Stress. ACS Omega 2022, 7, 31834–31844. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Chung, Y.S.; Lee, E.; Tripathi, P.; Heo, S.; Kim, K.-H. Root response to drought stress in rice (Oryza sativa L.). Int. J. Mol. Sci. 2020, 21, 1513. [Google Scholar] [CrossRef] [Green Version]
- Le, V.N.; Rui, Y.; Gui, X.; Li, X.; Liu, S.; Han, Y. Uptake, transport, distribution and bio-effects of SiO2 nanoparticles in Bt-transgenic cotton. J. Nanobiotechnol. 2014, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Alghamdi, S.A.; Alharby, H.F.; Bamagoos, A.A.; Zaki, S.-n.S.; Abu El-Hassan, A.M.; Desoky, E.-S.M.; Mohamed, I.A.; Rady, M.M. Rebalancing Nutrients, Reinforcing Antioxidant and Osmoregulatory Capacity, and Improving Yield Quality in Drought-Stressed Phaseolus vulgaris by Foliar Application of a Bee-Honey Solution. Plants 2022, 12, 63. [Google Scholar] [CrossRef]
- Guo, K.; Hu, A.; Wang, K.; Wang, L.; Fu, D.; Hao, Y.; Wang, Y.; Ali, A.; Adeel, M.; Rui, Y. Effects of spraying nano-materials on the absorption of metal (loid) s in cucumber. IET Nanobiotechnol. 2019, 13, 712–719. [Google Scholar] [CrossRef]
- Amer, M.; El-Emary, F.A. Impact of foliar with nano-silica in mitigation of salt stress on some soil properties, crop-water productivity and anatomical structure of maize and faba bean. Environ. Biodivers. Soil Secur. 2018, 2, 25–38. [Google Scholar]
- Ali, M.; Afzal, S.; Parveen, A.; Kamran, M.; Javed, M.R.; Abbasi, G.H.; Malik, Z.; Riaz, M.; Ahmad, S.; Chattha, M.S. Silicon mediated improvement in the growth and ion homeostasis by decreasing Na+ uptake in maize (Zea mays L.) cultivars exposed to salinity stress. Plant Physiol. Biochem. 2021, 158, 208–218. [Google Scholar] [CrossRef]
- Liu, B.; Soundararajan, P.; Manivannan, A. Mechanisms of silicon-mediated amelioration of salt stress in plants. Plants 2019, 8, 307. [Google Scholar] [CrossRef] [Green Version]
- Ocvirk, D.; Špoljarević, M.; Kristić, M.; Hancock, J.T.; Teklić, T.; Lisjak, M. The effects of seed priming with sodium hydrosulphide on drought tolerance of sunflower (Helianthus annuus L.) in germination and early growth. Ann. Appl. Biol. 2021, 178, 400–413. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Property | Soil Layer (cm) | |||
|---|---|---|---|---|
| 0–15 | 15–30 | 30–45 | 45–60 | |
| Particle size distribution: | ||||
| Coarse sand (%) | 4.78 | 4.81 | 5.11 | 5.39 |
| Fine sand (%) | 74.35 | 75.14 | 75.58 | 75.99 |
| Silt (%) | 6.55 | 6.52 | 6.44 | 5.98 |
| Clay (%) | 14.32 | 13.53 | 12.87 | 12.64 |
| Texture class | Sandy loam | Sandy loam | Sandy loam | Sandy loam |
| Field capacity , %) | 14.45 | 14.14 | 13.96 | 13.84 |
| Permanent wilting point (, %) | 6.11 | 5.99 | 5.84 | 5.71 |
| Available water (AW, %) | 8.34 | 8.15 | 8.12 | 8.13 |
| Bulk density (Mg m−3) | 1.46 | 1.65 | 1.71 | 1.74 |
| Total porosity (%) | 44.91 | 37.74 | 35.47 | 34.34 |
| pH (1:2.5 soil/water suspension) | 7.88 | 7.87 | 7.91 | 7.94 |
| ECe (soil paste extract, dSm−1) | 1.65 | 1.63 | 1.50 | 1.35 |
| Organic carbon (g kg−1) | 2.69 | 2.66 | 2.54 | 2.44 |
| Organic matter (g kg−1) | 4.63 | 4.58 | 4.37 | 4.20 |
| CaCO3 content (g kg−1) | 21.10 | 22.35 | 23.15 | 23.87 |
| Soluble cations (mmolc L−1): | ||||
| Ca2+ | 2.15 | 2.09 | 1.97 | 1.85 |
| Mg2+ | 2.77 | 2.69 | 2.59 | 2.37 |
| Na+ | 10.91 | 10.86 | 9.85 | 8.71 |
| K+ | 0.67 | 0.65 | 0.63 | 0.59 |
| Soluble anions (mmolc L−1): | ||||
| CO32− | 0.00 | 0.00 | 0.00 | 0.00 |
| HCO3− | 2.97 | 2.91 | 2.87 | 2.75 |
| Cl− | 8.98 | 8.85 | 7.79 | 6.82 |
| SO42− | 4.55 | 4.53 | 4.38 | 3.95 |
| Month | Temperature (°C) | Wind Speed (m s−1) | Relative Humidity (%) | Surface Pressure (kPa) | ||
|---|---|---|---|---|---|---|
| Max | Min | Max | Min | |||
| 2021 | ||||||
| May | 42.01 | 11.83 | 10.33 | 0.40 | 50.69 | 99.77 |
| June | 42.12 | 15.25 | 10.20 | 0.38 | 41.50 | 99.48 |
| July | 41.68 | 18.37 | 8.44 | 0.44 | 42.88 | 99.12 |
| August | 42.68 | 20.30 | 8.48 | 0.80 | 45.38 | 99.15 |
| 2022 | ||||||
| May | 41.67 | 14.90 | 9.29 | 0.32 | 36.69 | 99.64 |
| June | 41.45 | 15.86 | 8.85 | 1.73 | 41.38 | 99.62 |
| July | 43.00 | 20.12 | 9.15 | 1.01 | 41.12 | 99.16 |
| August | 44.01 | 20.37 | 8.41 | 0.49 | 42.88 | 99.26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Mokadem, A.Z.; Sheta, M.H.; Mancy, A.G.; Hussein, H.-A.A.; Kenawy, S.K.M.; Sofy, A.R.; Abu-Shahba, M.S.; Mahdy, H.M.; Sofy, M.R.; Al Bakry, A.F.; et al. Synergistic Effects of Kaolin and Silicon Nanoparticles for Ameliorating Deficit Irrigation Stress in Maize Plants by Upregulating Antioxidant Defense Systems. Plants 2023, 12, 2221. https://doi.org/10.3390/plants12112221
Al-Mokadem AZ, Sheta MH, Mancy AG, Hussein H-AA, Kenawy SKM, Sofy AR, Abu-Shahba MS, Mahdy HM, Sofy MR, Al Bakry AF, et al. Synergistic Effects of Kaolin and Silicon Nanoparticles for Ameliorating Deficit Irrigation Stress in Maize Plants by Upregulating Antioxidant Defense Systems. Plants. 2023; 12(11):2221. https://doi.org/10.3390/plants12112221
Chicago/Turabian StyleAl-Mokadem, Alshymaa Z., Mohamed H. Sheta, Ahmed G. Mancy, Hebat-Allah A. Hussein, Sahar K. M. Kenawy, Ahmed R. Sofy, Mahmoud S. Abu-Shahba, Hesham M. Mahdy, Mahmoud R. Sofy, Alaa Fathy Al Bakry, and et al. 2023. "Synergistic Effects of Kaolin and Silicon Nanoparticles for Ameliorating Deficit Irrigation Stress in Maize Plants by Upregulating Antioxidant Defense Systems" Plants 12, no. 11: 2221. https://doi.org/10.3390/plants12112221
APA StyleAl-Mokadem, A. Z., Sheta, M. H., Mancy, A. G., Hussein, H. -A. A., Kenawy, S. K. M., Sofy, A. R., Abu-Shahba, M. S., Mahdy, H. M., Sofy, M. R., Al Bakry, A. F., & Agha, M. S. (2023). Synergistic Effects of Kaolin and Silicon Nanoparticles for Ameliorating Deficit Irrigation Stress in Maize Plants by Upregulating Antioxidant Defense Systems. Plants, 12(11), 2221. https://doi.org/10.3390/plants12112221