
Population Genetics, Genetic Structure, and Inbreeding of Commiphora gileadensis (L.) C. Chr Inferred from SSR Markers in Some Mountainous Sites of Makkah Province


Abstract
:1. Introduction
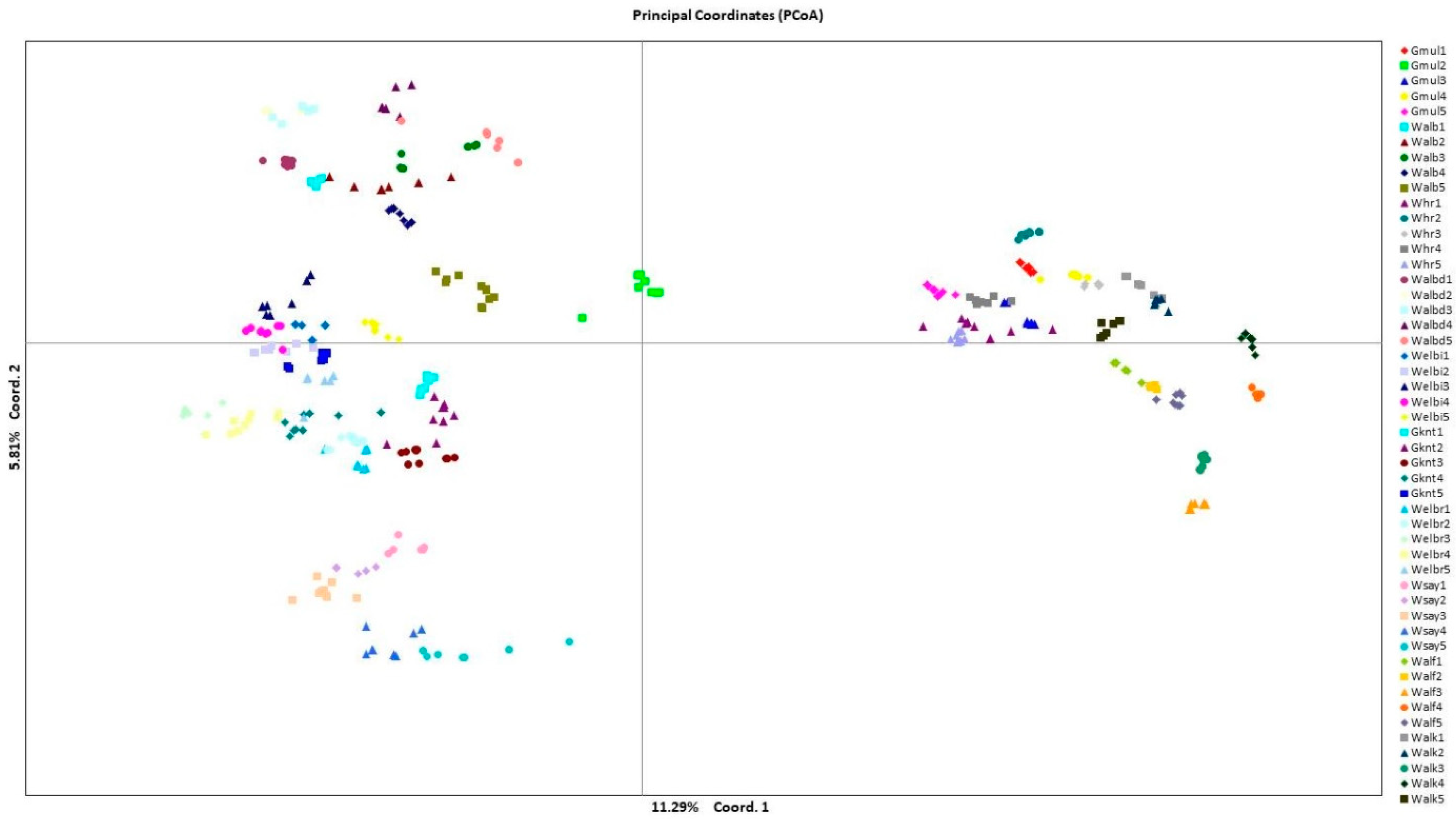
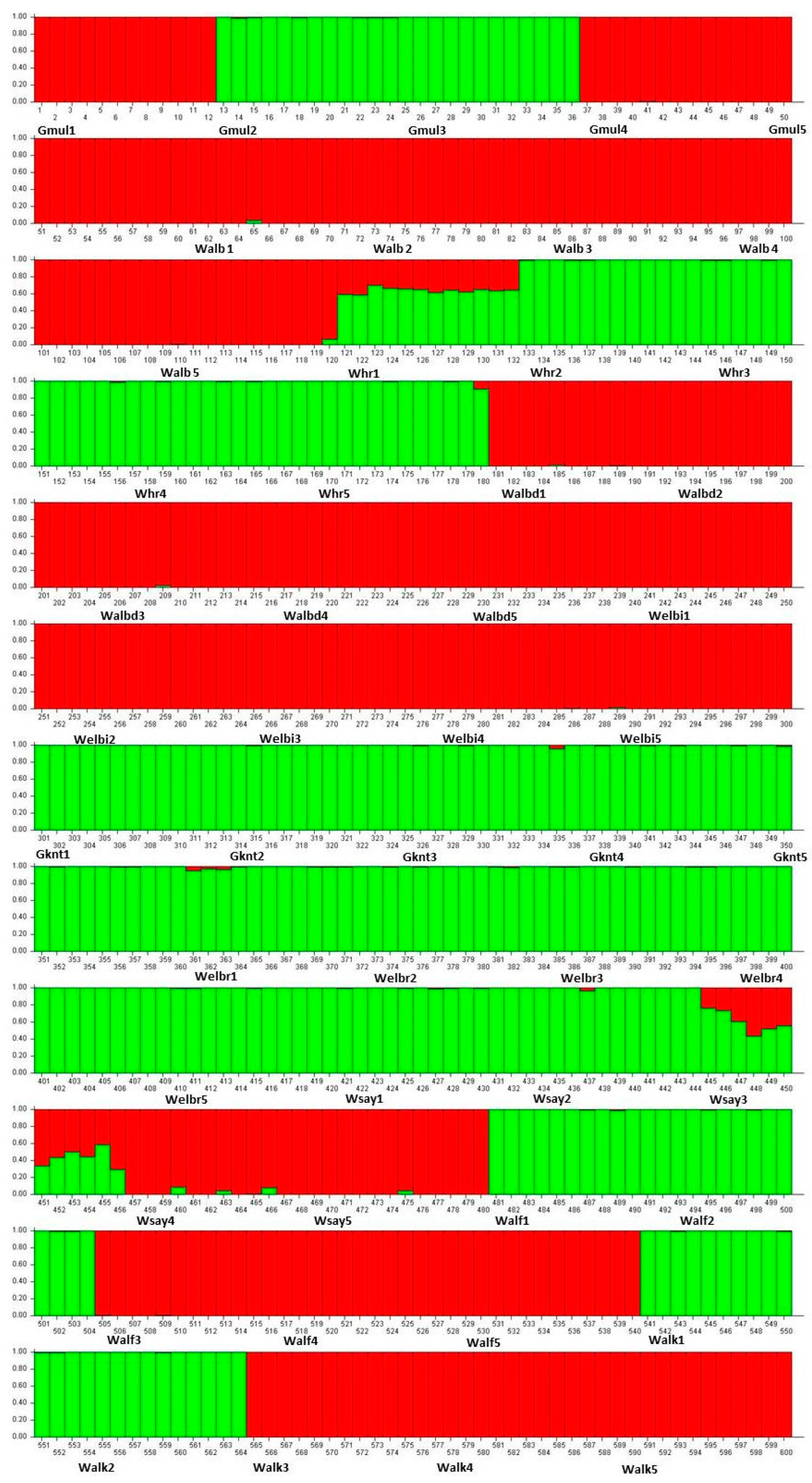
2. Results
3. Discussion
4. Materials and Methods
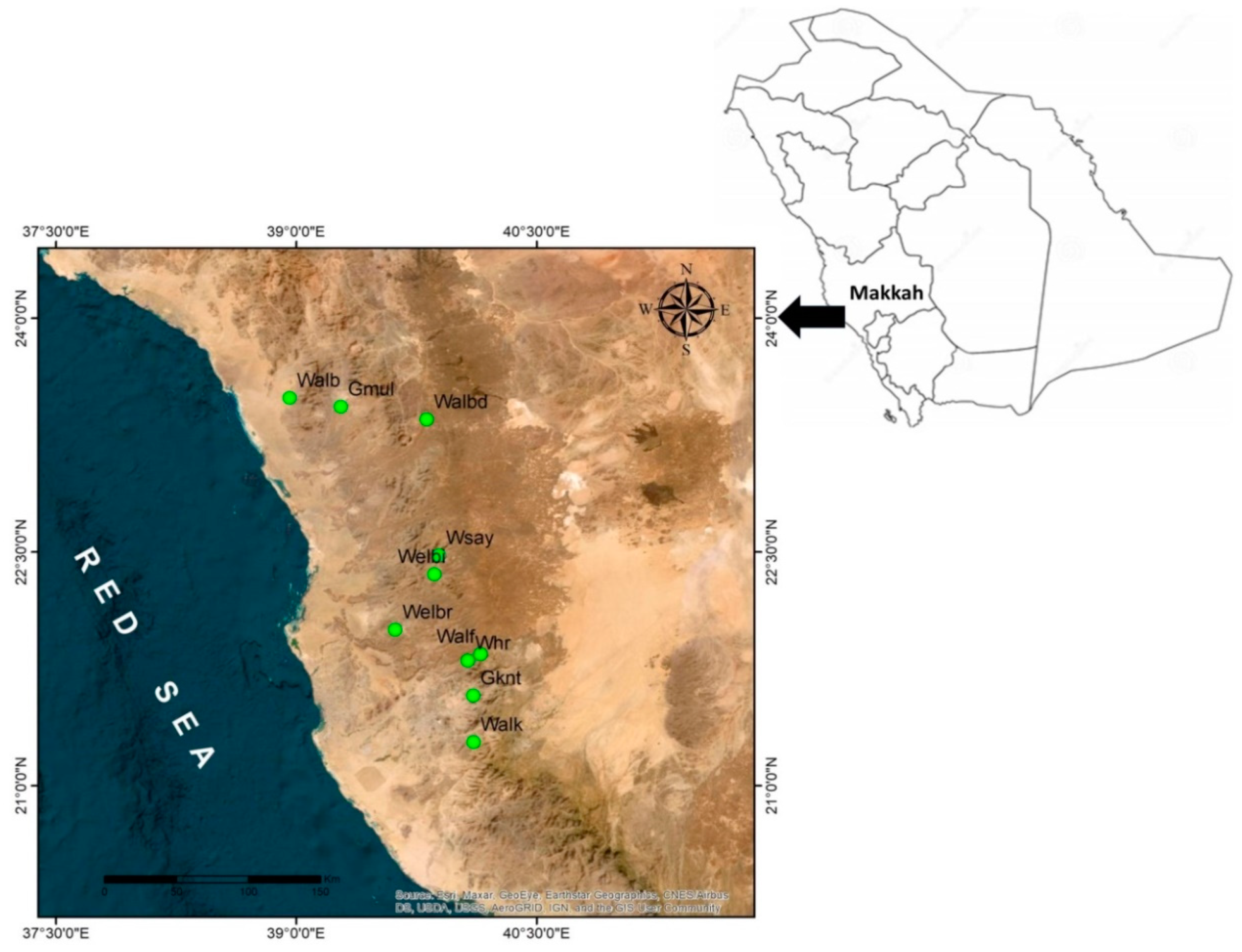
4.1. Plant Materials
4.2. Genomic DNA Extraction and PCR Tests
4.3. Population Genetic Analysis
5. Conclusions and Recommendations
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mathur, M.; Mathur, P.; Purohit, H. Ecological niche modelling of a critically endangered species Commiphora wightii (Arn.) Bhandari using bioclimatic and non-bioclimatic variables. Ecol. Process 2023, 12, 8. [Google Scholar] [CrossRef]
- Miller, A.G.; Morris, M. Plants of Dhofar, the Southern Region of Oman: Traditional, Economic and Medicinal Uses; The Office of the Advisor for Conservation of the Environment, Diwan of Royal Court Sultanate of Oman: Muscat, Oman, 1988. [Google Scholar]
- Wood, J.R.I. A Handbook of the Yemen Flora. With Color Illustrations by Hugo Haig-Thomas; Royal Botanic Gardens: Kew, UK, 1997. [Google Scholar]
- Shen, T.; Li, G.H.; Wang, X.N.; Lou, H.X. The genus Commiphora: A review of its traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2012, 142, 319–330. [Google Scholar] [CrossRef]
- Mahr, D. Commiphora: An Introduction to the Genus. Cactus Succul. J. 2012, 84, 140–154. [Google Scholar] [CrossRef]
- González-Tejero, M.R.; Casares-Porcel, M.; Sánchez-Rojas, C.P.; Ramiro-Gutiérrez, J.M.; Molero-Mesa, J.; Pieroni, A.; Giusti, M.E.; Censorii, E.; De Pasquale, C.; Della, A.; et al. Medicinal plants in the Mediterranean area: Synthesis of the results of the project Rubia. J. Ethnopharmacol. 2008, 116, 341–357. [Google Scholar] [CrossRef]
- Tarawneh, Q.Y.; Chowdhury, S. Trends of Climate Change in Saudi Arabia: Implications on Water Resources. Climate 2018, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Issar, A.S. The impact of global warming on the water resources of the Middle East: Past, present and future. In Climate Changes and Water Resources in the Middle East and North Africa; Zereini, F., Hötzl, H., Eds.; Springer: Heidelberg, Germany, 2008. [Google Scholar]
- Soultan, A.; Wikelski, M.; Safi, K. Risk of biodiversity collapse under climate change in the Afro-Arabian region. Sci. Rep. 2019, 9, 955. [Google Scholar] [CrossRef] [Green Version]
- Luijten, S.H.; Dierick, A.; Gerard, J.; Oostermeijer, B.; Raijmann, L.E.J.; Den Nijs, H.C.M. Population size, genetic variation, and reproductive success in a rapidly declining, self-incompatible perennial (Arnica montana) in the Netherlands. Conserv. Biol. 2000, 14, 1776–1787. [Google Scholar]
- Hansson, B.; Westerberg, L. On the correlation between heterozygosity and fitness in natural populations. Mol. Ecol. 2002, 11, 2467–2474. [Google Scholar] [CrossRef] [Green Version]
- Bastiaan, S.; Hamish, G.S. Effects of genetic drift and gene flow on the selective maintenance of genetic variation. Genetics 2003, 194, 235–244. [Google Scholar]
- Shalabi, L.F.; Otaif, F.S. Commiphora Jacq (Burseraceae) in Saudi Arabia, Botanical, Phytochemical and Ethnobotanical Notes. Ecologies 2022, 3, 38–57. [Google Scholar] [CrossRef]
- Hatmaker, E.A.; Staton, M.E.; Dattilo, A.J.; Hadziabdic, D.; Rinehart, T.A.; Schilling, E.E.; Trigiano, R.N.; Wadl, P.A. Population Structure and Genetic Diversity within the endangered species Pityopsis ruthii (Asteraceae). Front. Plant Sci. 2018, 9, 943. [Google Scholar] [CrossRef] [Green Version]
- Raju, A.J.S.; Lakshmi, P.V.; Ramana, K.V.; Chandra, P.H. Entomophily, ornithophily and anemochory in the self-incompatible Boswellia ovalifoliolata Bal. & Henry (Burseraceae), an endemic and endangered medicinally important tree species. J. Threat. Taxa 2012, 4, 2673–2684. [Google Scholar]
- Keller, L.F.; Waller, D.M. Inbreeding effects in wild populations. Trends Ecol. Evol. 2002, 17, 230–241. [Google Scholar] [CrossRef]
- Vilas, C.; San Miguel, E.; Amaro, R.; García, C. Relative contribution of inbreeding depression and eroded adaptive diversity to extinction risk in small populations of shore campion. Conserv. Biol. 2005, 20, 229–238. [Google Scholar] [CrossRef]
- Kadam, U.S.; Lossie, A.C.; Schulz, B.; Irudayaraj, J. Gene expression analysis using conventional and imaging methods. In DNA and RNA Nanobiotechnologies in Medicine: Diagnosis and Treatment of Diseases; Springer: Berlin/Heidelberg, Germany, 2013; pp. 141–162. [Google Scholar]
- Kim, K.S.; Sappington, T.W. Microsatellite Data Analysis for Population Genetics. In Microsatellites. Methods in Molecular Biology (Methods and Protocols); Kantartzi, S., Ed.; Humana Press: Totowa, NJ, USA, 2013; Volume 1006, pp. 271–295. [Google Scholar]
- Upadhyay, A.; Kadam, U.S.; Chacko, P.; Karibasappa, G.S. Microsatellite and RAPD analysis of grape (Vitis spp.) accessions and identification of duplicates/misnomers in germplasm collection. Indian J. Hortic. 2010, 67, 8–15. [Google Scholar]
- Upadhyay, A.; Kadam, U.S.; Chacko, P.M.; Aher, L.; Karibasappa, G.S. Microsatellite analysis to differentiate clones of Thompson seedless grapevine. Indian J. Hortic. 2010, 67, 260–263. [Google Scholar]
- Hinge, V.R.; Shaikh, I.M.; Chavhan, R.L.; Deshmukh, A.S.; Shelake, R.M.; Ghuge, S.A.; Dethe, A.M.; Suprasanna, P.; Kadam, U.S. Assessment of genetic diversity and volatile content of commercially grown banana (Musa spp.) cultivars. Sci. Rep. 2022, 12, 1–6. [Google Scholar] [CrossRef]
- Chavhan, R.L.; Sable, S.; Narwade, A.V.; Hinge, V.R.; Kalbande, B.B.; Mukherjee, A.K.; Chakrabarty, P.K.; Kadam, U.S. Multiplex molecular marker-assisted analysis of significant pathogens of cotton (Gossypium sp.). Biocatal. Agric. Biotechnol. 2023, 47, 102557. [Google Scholar] [CrossRef]
- Szczecińska, M.; Sramko, G.; Wołosz, K.; Sawicki, J. Genetic Diversity and Population Structure of the Rare and Endangered Plant Species Pulsatilla patens (L.) Mill in East Central Europe. PLoS ONE 2016, 11, e0151730. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.L.; Wang, H.C.; Yu, Z.X.; Schinnerl, J.; Tang, R.; Geng, Y.P.; Chen, G. Genetic diversity and structure of the endemic and endangered species Aristolochia delavayi growing along the Jinsha River. Plant Divers 2021, 43, 225–233. [Google Scholar] [CrossRef]
- Chen, L.; Pan, T.; Qian, H.; Zhang, M.; Yang, G.; Wang, X. Genetic Diversity and Population Structure Revealed by SSR Markers on Endemic Species Osmanthus serrulatus Rehder from Southwestern Sichuan Basin, China. Forests 2021, 12, 1365. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Misiewicz, T.M.; Barbosa, C.E.; Fine, P.V. Microsatellite primers for an Amazonian lowland tropical tree, Protium subserratum (Burseraceae). Am. J. Bot. 2012, 99, e465-7. [Google Scholar] [CrossRef] [Green Version]
- Maradani, B.S.; Gudasalamani, R.; Setty, S.; Chandrasekaran, R. Development of microsatellite markers for the resin-yielding, non-timber forest product species Boswellia serrata (Burseraceae). Appl. Plant Sci. 2018, 6, e01180. [Google Scholar] [CrossRef] [Green Version]
- Koffi, K.G.; Heuertz, M.; Jans, R.; Hardy, O.J.; Vendramin, G.G.; Duminil, J. Characterization of new microsatellite loci isolated from Santiria trimera (Burseraceae). Am. J. Bot. 2012, 99, e334-6. [Google Scholar] [CrossRef]
- Rimlinger, A.; Marie, L.; Avana, M.L.; Bouka, G.U.; Zekraoui, L.; Mariac, C.; Carrière, S.M.; Duminil, J. New microsatellite markers for Dacryodes edulis (Burseraceae), an indigenous fruit tree species from Central Africa. Mol. Biol. Rep. 2020, 47, 2391–2396. [Google Scholar] [CrossRef]
- Mansour, H.; Sliwinska, E. Genetic Diversity and Inbreeding Level of Cotoneaster orbicularis Schltdl. in The Sinai Mountains Revealed by Microsatellite Markers and Flow Cytometry. Egypt. J. Bot. 2017, 2, 351–361. [Google Scholar] [CrossRef]
- Mansour, H.; Alsamadany, H.; Al-Hasawi, Z.M. Genetic diversity and genetic structure of Salvadora persica L., rare plant species in Rabigh province, Saudi Arabia: Implications for conservation. J. Taibah Univ. Sci. 2020, 14, 881–888. [Google Scholar] [CrossRef]
- Mansour, H.; Alsamadany, H.; Al-Hasawi, Z.M. Molecular Assessment of Genetic Diversity and Genetic Structure of Rhanterium epapposum Oliv. in Scarce Populations in Some Regions of Western Saudi Arabia. Plants 2022, 11, 1560. [Google Scholar] [CrossRef]
- Jimenez, A.; Mansour, H.; Keller, B.; Conti, E. Low genetic diversity and a high level of inbreeding in the Sinai primrose (Primula boveana), a species on the brink of extinction. Plant Syst. Evol. 2014, 300, 1199–1208. [Google Scholar] [CrossRef] [Green Version]
- Blomqvist, D.; Pauliny, A.; Larsson, M.; Flodin, L.A. Trapped in the extinction vortex? Strong genetic effects in a declining vertebrate population. BMC Evol. Biol. 2010, 10, 33. [Google Scholar] [CrossRef] [Green Version]
- Jacquemyn, H.; Roldán-Ruiz, I.; Honnay, O. Evidence for demographic bottlenecks and limited gene flow leading to low genetic diversity in a rare thistle. Conserv. Genet. 2010, 11, 1979–1987. [Google Scholar] [CrossRef]
- Smyser, T.J.; Duchamp, J.E.; Johnson, S.A.; Larkin, J.L.; Rhodes, O.E., Jr. Consequences of metapopulation collapse: Comparison of genetic attributes between two Allegheny woodrat metapopulations. Conserv. Genet. 2012, 13, 849–858. [Google Scholar] [CrossRef]
- Al-Gharaibeh, M.M.; Hamasha, H.R.; Rosche, C.; Lachmuth, S.; Wesche, K.; Hensen, I. Environmental gradients shape the genetic structure of two medicinal Salvia species in Jordan. Plant Biol. 2017, 19, 227–238. [Google Scholar]
- Al-Rowaily, S.L.; El-Bana, M.I.; Al-Bakre, D.A.; Assaeed, A.M.; Hegazy, A.K.; Ali, M.B. Effects of open grazing and livestock exclusion on floristic composition and diversity in natural ecosystem of Western Saudi Arabia. Saudi. J. Biol. Sci. 2015, 22, 430–437. [Google Scholar] [CrossRef] [Green Version]
- Harter, T.; Davis, H.; Mathews, M.; Meyer, R. Shallow ground water quality on dairy farms with irrigated forage crops. J. Contam. Hydrol. 2022, 55, 287–315. [Google Scholar] [CrossRef] [PubMed]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef]
- Spieth, P.T. Gene flow and genetic differentiation. Genetics 1974, 78, 961–965. [Google Scholar] [CrossRef]
- Arif, I.A.; Khan, H.A.; Shobrak, M.; Al Homaidan, A.A.; Al Sadoon, M.; Al Farhan, A.H.; Bahkali, A.H. Interpretation of electrophoretograms of seven microsatellite loci to determine the genetic diversity of the Arabian Oryx. Genet. Mol. Res. 2010, 9, 259–265. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx v.6.5: Genetic analysis in Excel. Population genetic software for teaching and research. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Slatkin, M. A measure of population subdivision based on microsatellite allele frequencies. Genetics 1995, 139, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.; Stephens, M.; Rosenberg, N.; Donnelly, P. Association mapping in structured populations. Am. J. Hum. Genet. 2000, 67, 170–180. [Google Scholar] [CrossRef] [Green Version]
- Earl, E.A.; Von Holdt, B.M. Structure Harvester: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Res. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Michalakis, Y.; Excoffier, L. A generic estimation of population subdivision using distances between alleles with special reference for microsatellite loci. Genetics 1996, 142, 1061–1064. [Google Scholar] [CrossRef]
- Barton, N.H.; Slatkin, M. A Quasi-equilibrium theory of the distribution of rare alleles in a subdivided population. Heredity 1986, 56, 409–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, A.; Uchida, K.; Ozeki, M.; Iwasaki, T.; Nakahama, N.; Suka, T. Conservation of endangered and rare plants requires strategies additional to deer-proof fencing for conservation of sub-alpine plant diversity. Appl. Veg. Sci. 2021, 24, e12553. [Google Scholar] [CrossRef]
- Oakley, C.G.; Lundemo, S.; Ågren, J.; Schemske, D.W. Heterosis is common and inbreeding depression absent in natural populations of Arabidopsis thaliana. J. Evol. Biol. 2019, 32, 592–603. [Google Scholar] [CrossRef]
- Holsinger, K.E.; Gottlieb, L.D. Conservation of rare and endangered plants: Principles and prospects. In Genetics and Conservation of Rare Plants; Falk, D.A., Holsinger, K.E., Eds.; Oxford University Press: New York, NY, USA, 1991; pp. 195–208. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Population | Na | Ne | I | No. of Private Alleles | Ho | He | P | Fis |
|---|---|---|---|---|---|---|---|---|
| Gmul 1 | 1.200 | 1.103 | 0.092 | 0.066 | 0.073 | 0.058 | 18.00 | −0.221 |
| Gmul 2 | 1.320 | 1.144 | 0.139 | 0.057 | 0.083 | 0.085 | 26.00 | 0.136 |
| Gmul 3 | 1.440 | 1.281 | 0.209 | 0.087 | 0.143 | 0.131 | 32.00 | −0.073 |
| Gmul 4 | 1.380 | 1.166 | 0.145 | 0.066 | 0.078 | 0.084 | 26.00 | 0.163 |
| Gmul 5 | 1.340 | 1.172 | 0.155 | 0.052 | 0.083 | 0.097 | 30.00 | 0.064 |
| Walb 1 | 1.140 | 1.062 | 0.066 | 0.000 | 0.040 | 0.042 | 14.00 | −0.031 |
| Walb 2 | 1.300 | 1.163 | 0.151 | 0.028 | 0.042 | 0.098 | 28.00 | 0.583 |
| Walb 3 | 1.240 | 1.144 | 0.122 | 0.020 | 0.047 | 0.079 | 20.00 | 0.317 |
| Walb 4 | 1.220 | 1.080 | 0.085 | 0.000 | 0.013 | 0.052 | 22.00 | 0.691 |
| Walb 5 | 1.480 | 1.229 | 0.217 | 0.000 | 0.112 | 0.138 | 40.00 | 0.152 |
| Whr 1 | 1.340 | 1.180 | 0.162 | 0.057 | 0.133 | 0.104 | 30.00 | −0.123 |
| Whr 2 | 1.340 | 1.161 | 0.140 | 0.064 | 0.057 | 0.084 | 26.00 | 0.335 |
| Whr 3 | 1.180 | 1.061 | 0.067 | 0.059 | 0.050 | 0.041 | 18.00 | 0.009 |
| Whr 4 | 1.280 | 1.124 | 0.117 | 0.039 | 0.065 | 0.071 | 22.00 | 0.162 |
| Whr 5 | 1.160 | 1.059 | 0.068 | 0.052 | 0.035 | 0.042 | 16.00 | 0.234 |
| Walbd1 | 1.120 | 1.056 | 0.054 | 0.028 | 0.043 | 0.034 | 10.00 | −0.147 |
| Walbd2 | 1.100 | 1.037 | 0.040 | 0.000 | 0.018 | 0.025 | 10.00 | 0.092 |
| Walbd3 | 1.240 | 1.133 | 0.120 | 0.000 | 0.037 | 0.079 | 22.00 | 0.438 |
| Walbd4 | 1.120 | 1.037 | 0.047 | 0.000 | 0.007 | 0.028 | 12.00 | 0.796 |
| Walbd5 | 1.400 | 1.129 | 0.148 | 0.028 | 0.031 | 0.089 | 40.00 | 0.767 |
| Welbi1 | 1.080 | 1.039 | 0.039 | 0.028 | 0.010 | 0.025 | 8.00 | 0.429 |
| Welbi2 | 1.240 | 1.079 | 0.091 | 0.000 | 0.032 | 0.053 | 20.00 | 0.305 |
| Welbi3 | 1.260 | 1.152 | 0.139 | 0.000 | 0.037 | 0.092 | 26.00 | 0.528 |
| Welbi4 | 1.260 | 1.112 | 0.107 | 0.020 | 0.038 | 0.066 | 20.00 | 0.393 |
| Welbi5 | 1.240 | 1.118 | 0.115 | 0.000 | 0.087 | 0.072 | 20.00 | −0.204 |
| Gknt 1 | 1.420 | 1.256 | 0.189 | 0.070 | 0.142 | 0.113 | 30.00 | −0.245 |
| Gknt 2 | 1.420 | 1.187 | 0.162 | 0.044 | 0.072 | 0.093 | 30.00 | 0.330 |
| Gknt 3 | 1.340 | 1.227 | 0.160 | 0.062 | 0.055 | 0.098 | 20.00 | 0.415 |
| Gknt 4 | 1.420 | 1.241 | 0.195 | 0.020 | 0.077 | 0.121 | 30.00 | 0.347 |
| Gknt 5 | 1.360 | 1.208 | 0.155 | 0.070 | 0.080 | 0.091 | 24.00 | 0.160 |
| Welbr1 | 1.240 | 1.141 | 0.108 | 0.034 | 0.057 | 0.065 | 16.00 | 0.032 |
| Welbr2 | 1.180 | 1.079 | 0.080 | 0.000 | 0.030 | 0.049 | 14.00 | 0.428 |
| Welbr3 | 1.240 | 1.149 | 0.129 | 0.000 | 0.000 | 0.087 | 24.00 | 1.000 |
| Welbr4 | 1.280 | 1.166 | 0.147 | 0.044 | 0.052 | 0.097 | 26.00 | 0.417 |
| Welbr5 | 1.200 | 1.110 | 0.105 | 0.000 | 0.025 | 0.069 | 20.00 | 0.619 |
| Wsay1 | 1.160 | 1.077 | 0.073 | 0.048 | 0.050 | 0.047 | 16.00 | 0.007 |
| Wsay2 | 1.120 | 1.071 | 0.062 | 0.000 | 0.020 | 0.041 | 12.00 | 0.667 |
| Wsay3 | 1.260 | 1.121 | 0.118 | 0.028 | 0.055 | 0.075 | 24.00 | 0.440 |
| Wsay4 | 1.160 | 1.090 | 0.083 | 0.020 | 0.033 | 0.055 | 16.00 | 0.357 |
| Wsay5 | 1.180 | 1.119 | 0.102 | 0.020 | 0.042 | 0.069 | 18.00 | 0.434 |
| Walf 1 | 1.240 | 1.135 | 0.113 | 0.028 | 0.088 | 0.070 | 18.00 | −0.233 |
| Walf 2 | 1.100 | 1.051 | 0.048 | 0.000 | 0.037 | 0.031 | 10.00 | 0.089 |
| Walf 3 | 1.240 | 1.124 | 0.113 | 0.044 | 0.077 | 0.070 | 20.00 | −0.099 |
| Walf 4 | 1.140 | 1.084 | 0.065 | 0.020 | 0.027 | 0.042 | 10.00 | 0.409 |
| Walf 5 | 1.220 | 1.130 | 0.117 | 0.039 | 0.095 | 0.078 | 22.00 | −0.139 |
| Walk 1 | 1.240 | 1.120 | 0.106 | 0.020 | 0.073 | 0.065 | 18.00 | −0.139 |
| Walk 2 | 1.140 | 1.062 | 0.056 | 0.000 | 0.022 | 0.034 | 10.00 | 0.412 |
| Walk 3 | 1.180 | 1.101 | 0.095 | 0.000 | 0.072 | 0.062 | 18.00 | −0.158 |
| Walk 4 | 1.140 | 1.052 | 0.050 | 0.028 | 0.015 | 0.031 | 10.00 | 0.416 |
| Walk 5 | 1.220 | 1.136 | 0.114 | 0.069 | 0.070 | 0.073 | 18.00 | −0.024 |
| Overall mean | 1.245 | 1.125 | 0.112 | 0.029 | 0.055 | 0.070 | 20.60 | 0.238 |
| Population Acronym | Population Site | Longitude (E) | Latitude (N) | Altitude (m.) | Total No. of Individuals |
|---|---|---|---|---|---|
| Gmul 1 | Gebel Al Muliesaa | 39°16′43″ | 23°26′03″ | 216 | 22 |
| Gmul 2 | 30 | ||||
| Gmul 3 | 19 | ||||
| Gmul 4 | 20 | ||||
| Gmul 5 | 15 | ||||
| Walb 1 | Wadi Albathna | 38°57′32″ | 23°29′22″ | 371 | 19 |
| Walb 2 | 29 | ||||
| Walb 3 | 38 | ||||
| Walb 4 | 33 | ||||
| Walb 5 | 30 | ||||
| Whr 1 | Wadi Houra | 40°04′18″ | 21°48′16″ | 612 | 16 |
| Whr 2 | 26 | ||||
| Whr 3 | 30 | ||||
| Whr 4 | 35 | ||||
| Whr 5 | 22 | ||||
| Walbd1 | Wadi Albaidaa | 39°48′54″ | 23°21′07″ | 600 | 48 |
| Walbd2 | 36 | ||||
| Walbd3 | 30 | ||||
| Walbd4 | 42 | ||||
| Walbd5 | 33 | ||||
| Welbi1 | Wadi Elebiedia | 39°51′43″ | 22°21′31″ | 551 | 44 |
| Welbi2 | 40 | ||||
| Welbi3 | 33 | ||||
| Welbi4 | 49 | ||||
| Welbi5 | 31 | ||||
| Gknt 1 | Gebel Kniethl | 40°06′19″ | 21°34′45″ | 762 | 38 |
| Gknt 2 | 31 | ||||
| Gknt 3 | 22 | ||||
| Gknt 4 | 37 | ||||
| Gknt 5 | 35 | ||||
| Welbr | Wadi Elbarasa | 39°37′07″ | 22°00′09″ | 320 | 44 |
| Welbr | 22 | ||||
| Welbr | 29 | ||||
| Welbr | 26 | ||||
| Welbr | 35 | ||||
| Wsay1 | Wadi Sayaa | 39°53′32″ | 22°28′42″ | 629 | 39 |
| Wsay2 | 36 | ||||
| Wsay3 | 30 | ||||
| Wsay4 | 22 | ||||
| Wsay5 | 28 | ||||
| Walf 1 | Wadi Alfawara | 40°09′02″ | 21°50′42″ | 721 | 32 |
| Walf 2 | 40 | ||||
| Walf 3 | 33 | ||||
| Walf 4 | 23 | ||||
| Walf 5 | 18 | ||||
| Walk 1 | Wadi Alkharar | 40°06′26″ | 21°16′48″ | 747 | 34 |
| Walk 2 | 33 | ||||
| Walk 3 | 21 | ||||
| Walk 4 | 32 | ||||
| Walk 5 | 29 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mansour, H.; Alamer, K.H.; Al-Hasawi, Z.M. Population Genetics, Genetic Structure, and Inbreeding of Commiphora gileadensis (L.) C. Chr Inferred from SSR Markers in Some Mountainous Sites of Makkah Province. Plants 2023, 12, 2506. https://doi.org/10.3390/plants12132506
Mansour H, Alamer KH, Al-Hasawi ZM. Population Genetics, Genetic Structure, and Inbreeding of Commiphora gileadensis (L.) C. Chr Inferred from SSR Markers in Some Mountainous Sites of Makkah Province. Plants. 2023; 12(13):2506. https://doi.org/10.3390/plants12132506
Chicago/Turabian StyleMansour, Hassan, Khalid H. Alamer, and Zaki M. Al-Hasawi. 2023. "Population Genetics, Genetic Structure, and Inbreeding of Commiphora gileadensis (L.) C. Chr Inferred from SSR Markers in Some Mountainous Sites of Makkah Province" Plants 12, no. 13: 2506. https://doi.org/10.3390/plants12132506
APA StyleMansour, H., Alamer, K. H., & Al-Hasawi, Z. M. (2023). Population Genetics, Genetic Structure, and Inbreeding of Commiphora gileadensis (L.) C. Chr Inferred from SSR Markers in Some Mountainous Sites of Makkah Province. Plants, 12(13), 2506. https://doi.org/10.3390/plants12132506