
Analysis of Virus-Derived siRNAs in Strawberry Plants Co-Infected with Multiple Viruses and Their Genotypes
 ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
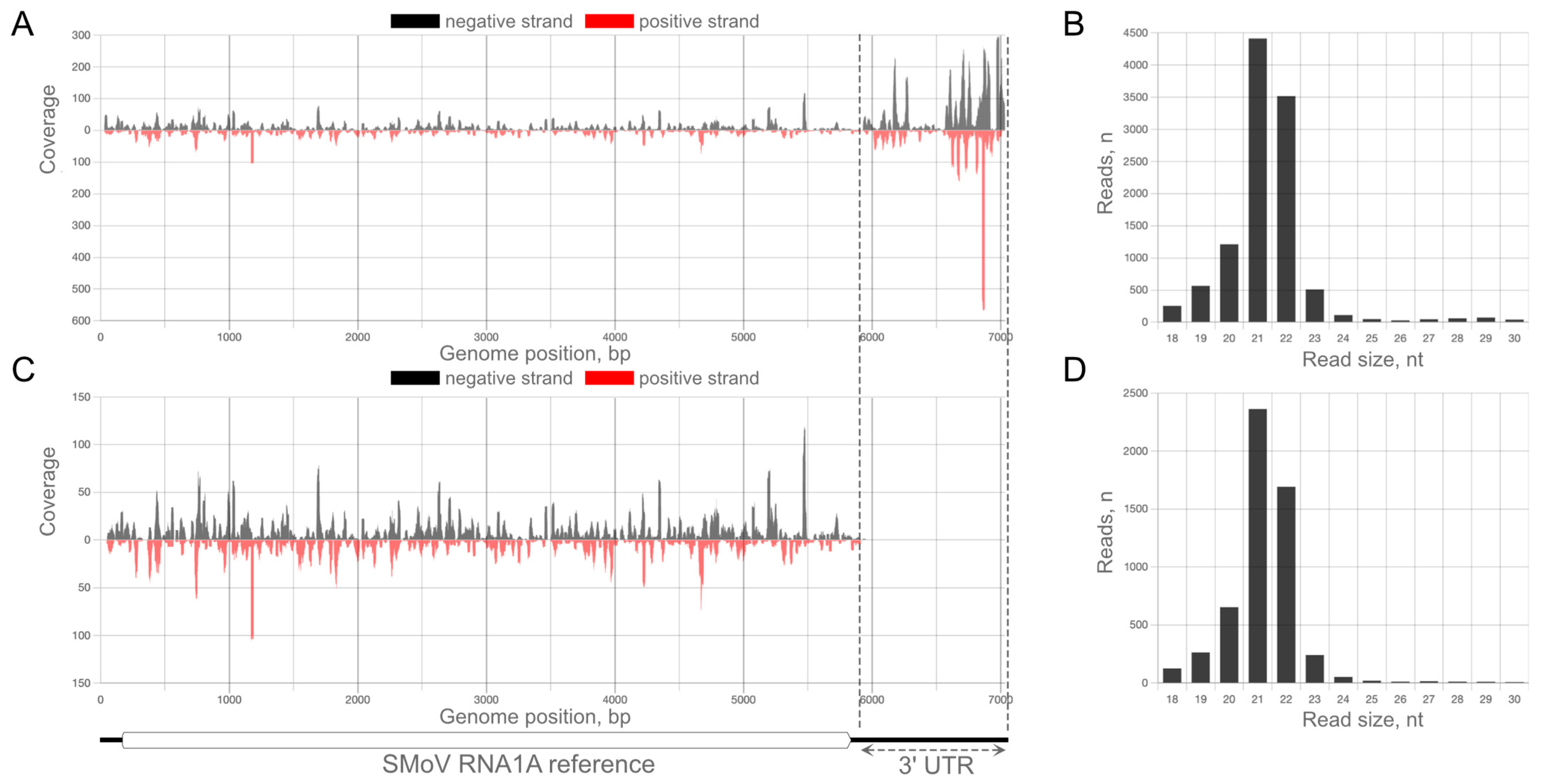
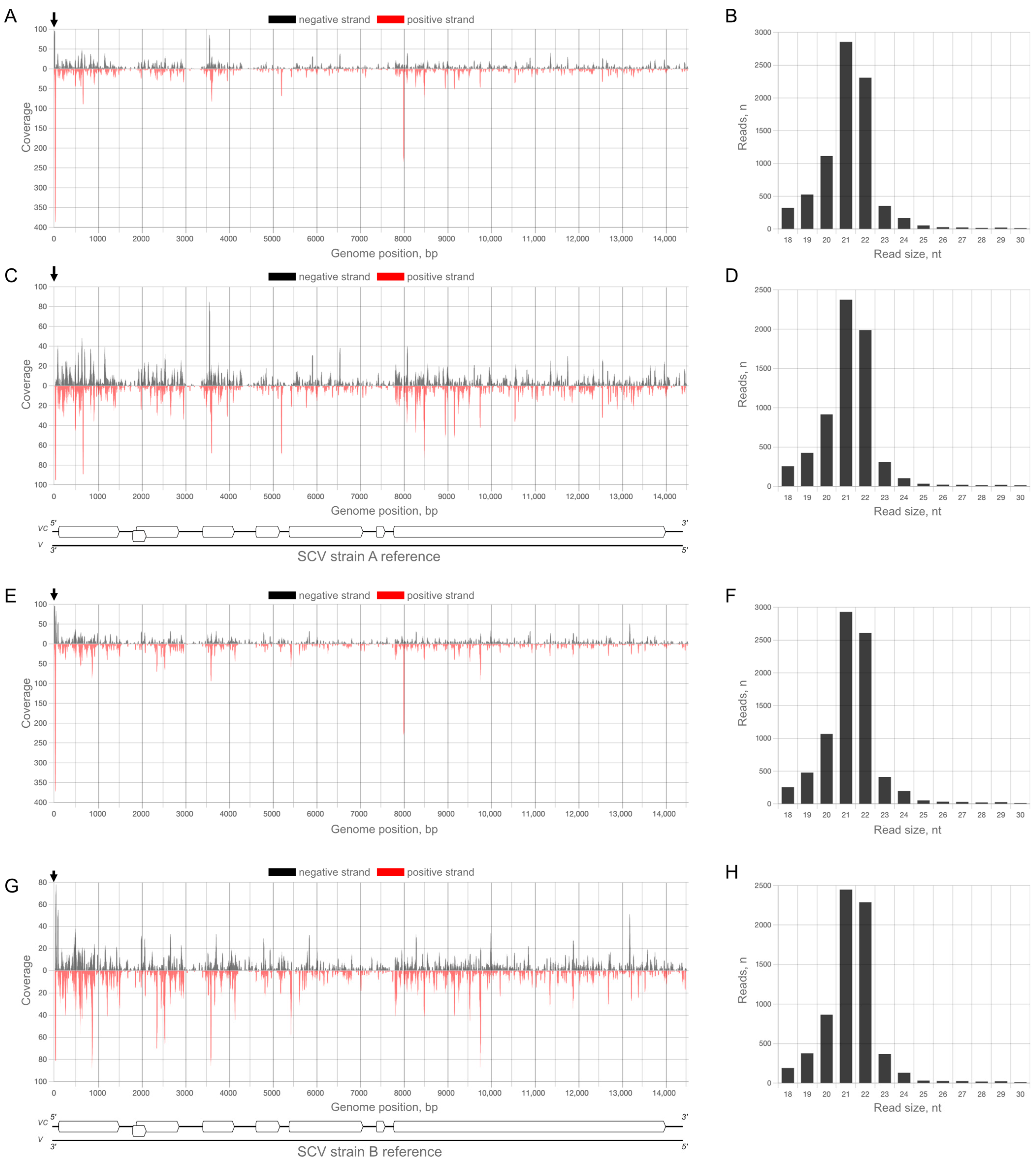
2.1. Analysis of vsiRNA Associated with the Original Complex of Viral Strains
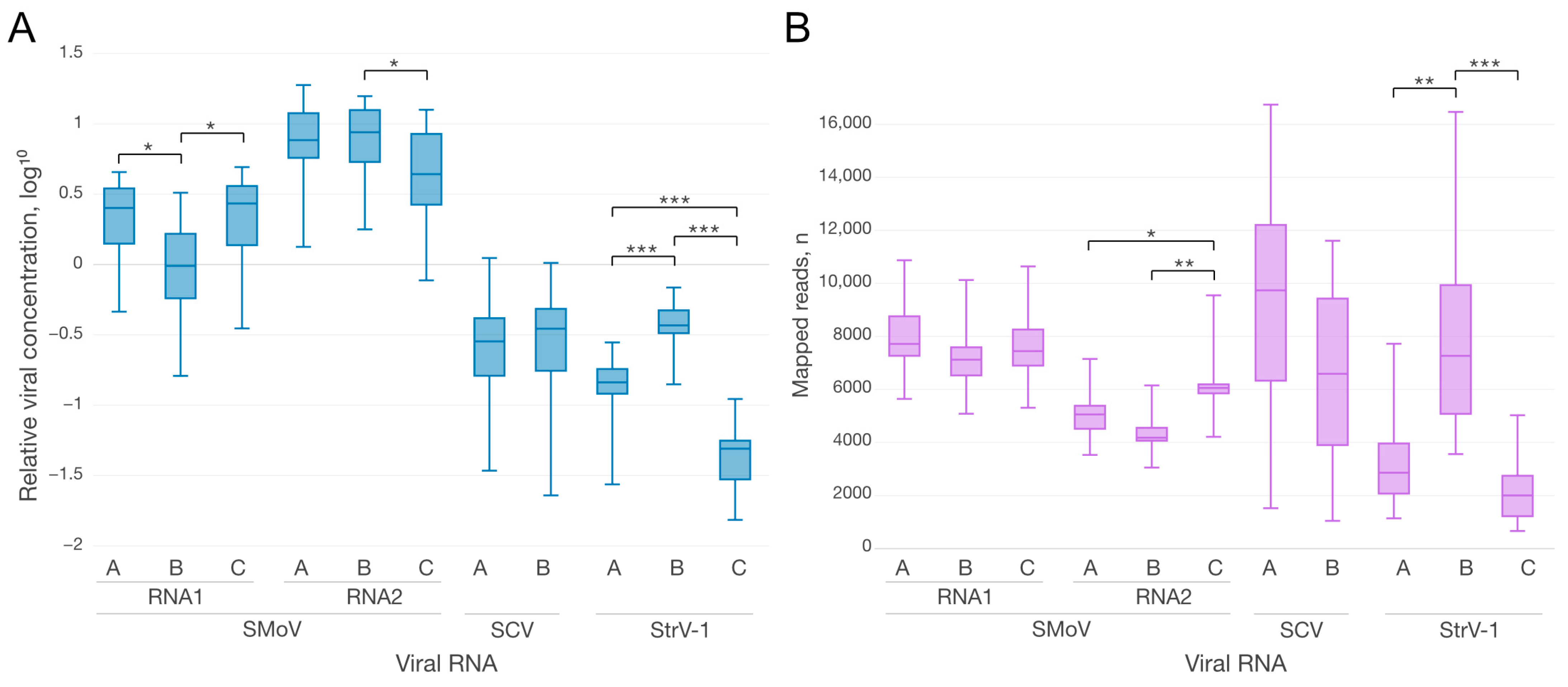
2.2. Comparison of Viral Concentrations and vsiRNA Abundances
2.3. The Abundance of vsiRNAs against Individual Strains of SMoV Does Not Correspond to Their Relative Concentration
2.4. Analysis of vsiRNA against StrV-1 Strains in StrV-1-Infected Strawberry
3. Summary and Conclusions
4. Materials and Methods
4.1. Plant Materials
4.2. Total and Small RNA Isolation
4.3. cDNA Synthesis and Two-Step Reverse-Transcription qPCR (RT-qPCR)
4.4. High-Throughput Sequencing and Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ding, S.-W.; Voinnet, O. Antiviral Immunity Directed by Small RNAs. Cell 2007, 130, 413–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnár, A.; Csorba, T.; Lakatos, L.; Várallyay, É.; Lacomme, C.; Burgyán, J. Plant Virus-Derived Small Interfering RNAs Originate Predominantly from Highly Structured Single-Stranded Viral RNAs. J. Virol. 2005, 79, 7812–7818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pooggin, M.M. Small RNA-Omics for Plant Virus Identification, Virome Reconstruction, and Antiviral Defense Characterization. Front. Microbiol. 2018, 9, 2779. [Google Scholar] [CrossRef] [PubMed]
- Brodersen, P.; Voinnet, O. The Diversity of RNA Silencing Pathways in Plants. Trends Genet. 2006, 22, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Sijen, T.; Fleenor, J.; Simmer, F.; Thijssen, K.L.; Parrish, S.; Timmons, L.; Plasterk, R.H.A.; Fire, A. On the Role of RNA Amplification in DsRNA-Triggered Gene Silencing. Cell 2001, 107, 465–476. [Google Scholar] [CrossRef] [Green Version]
- Parent, J.-S.; Bouteiller, N.; Elmayan, T.; Vaucheret, H. Respective Contributions of Arabidopsis DCL2 and DCL4 to RNA Silencing. Plant J. 2015, 81, 223–232. [Google Scholar] [CrossRef]
- Mohamed, A.; Jin, Z.; Osman, T.; Shi, N.; Tör, M.; Jackson, S.; Hong, Y. Hotspot SiRNA Confers Plant Resistance against Viral Infection. Biology 2022, 11, 714. [Google Scholar] [CrossRef]
- Rego-Machado, C.M.; Nakasu, E.Y.T.; Silva, J.M.F.; Lucinda, N.; Nagata, T.; Inoue-Nagata, A.K. SiRNA Biogenesis and Advances in Topically Applied DsRNA for Controlling Virus Infections in Tomato Plants. Sci. Rep. 2020, 10, 22277. [Google Scholar] [CrossRef]
- Rêgo-Machado, C.M.; Inoue-Nagata, A.K.; Nakasu, E.Y.T. Topical Application of DsRNA for Plant Virus Control: A Review. Trop. Plant Pathol. 2023, 48, 11–22. [Google Scholar] [CrossRef]
- Voloudakis, A.E.; Kaldis, A.; Patil, B.L. RNA-Based Vaccination of Plants for Control of Viruses. Annu. Rev. Virol. 2022, 9, 521–548. [Google Scholar] [CrossRef]
- Syller, J. Facilitative and Antagonistic Interactions between Plant Viruses in Mixed Infections: Plant Virus Interactions in Mixed Infections. Mol. Plant Pathol. 2012, 13, 204–216. [Google Scholar] [CrossRef]
- Villamor, D.E.V.; Ho, T.; Al Rwahnih, M.; Martin, R.R.; Tzanetakis, I.E. High Throughput Sequencing For Plant Virus Detection and Discovery. Phytopathology 2019, 109, 716–725. [Google Scholar] [CrossRef]
- Moreno, A.B.; López-Moya, J.J. When Viruses Play Team Sports: Mixed Infections in Plants. Phytopathology 2020, 110, 29–48. [Google Scholar] [CrossRef]
- Villamor, D.E.V.; Keller, K.E.; Martin, R.R.; Tzanetakis, I.E. Comparison of High Throughput Sequencing to Standard Protocols for Virus Detection in Berry Crops. Plant Dis. 2022, 106, 518–525. [Google Scholar] [CrossRef]
- Xu, Y.; Ghanim, M.; Liu, Y. Editorial: Mixed Infections of Plant Viruses in Nature and the Impact on Agriculture. Front. Microbiol. 2022, 13, 922607. [Google Scholar] [CrossRef]
- Cuevas, J.M.; Willemsen, A.; Hillung, J.; Zwart, M.P.; Elena, S.F. Temporal Dynamics of Intrahost Molecular Evolution for a Plant RNA Virus. Mol. Biol. Evol. 2015, 32, 1132–1147. [Google Scholar] [CrossRef] [Green Version]
- Hasiów-Jaroszewska, B.; Boezen, D.; Zwart, M.P. Metagenomic Studies of Viruses in Weeds and Wild Plants: A Powerful Approach to Characterise Variable Virus Communities. Viruses 2021, 13, 1939. [Google Scholar] [CrossRef]
- Makau, D.N.; Lycett, S.; Michalska-Smith, M.; Paploski, I.A.D.; Cheeran, M.C.-J.; Craft, M.E.; Kao, R.R.; Schroeder, D.C.; Doeschl-Wilson, A.; VanderWaal, K. Ecological and Evolutionary Dynamics of Multi-Strain RNA Viruses. Nat. Ecol. Evol. 2022, 6, 1414–1422. [Google Scholar] [CrossRef]
- Schreiber, S.J.; Ke, R.; Loverdo, C.; Park, M.; Ahsan, P.; Lloyd-Smith, J.O. Cross-Scale Dynamics and the Evolutionary Emergence of Infectious Diseases. Virus Evol. 2021, 7, veaa105. [Google Scholar] [CrossRef]
- Nuismer, S.L.; Basinski, A.J.; Schreiner, C.; Whitlock, A.; Remien, C.H. Reservoir Population Ecology, Viral Evolution and the Risk of Emerging Infectious Disease. Proc. R. Soc. B Biol. Sci. 2022, 289, 20221080. [Google Scholar] [CrossRef]
- Syller, J.; Grupa, A. Antagonistic Within-Host Interactions between Plant Viruses: Molecular Basis and Impact on Viral and Host Fitness: Antagonistic Interactions between Plant Viruses. Mol. Plant Pathol. 2016, 17, 769–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, L.J.S.; Bokil, V.A.; Cunniffe, N.J.; Hamelin, F.M.; Hilker, F.M.; Jeger, M.J. Modelling Vector Transmission and Epidemiology of Co-Infecting Plant Viruses. Viruses 2019, 11, 1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeger, M.J. The Epidemiology of Plant Virus Disease: Towards a New Synthesis. Plants 2020, 9, 1768. [Google Scholar] [CrossRef] [PubMed]
- Piche, L.M.; Singh, R.P.; Nie, X.; Gudmestad, N.C. Diversity Among Potato Virus Y Isolates Obtained from Potatoes Grown in the United States. Phytopathology 2004, 94, 1368–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folimonova, S.Y.; Harper, S.J.; Leonard, M.T.; Triplett, E.W.; Shilts, T. Superinfection Exclusion by Citrus Tristeza Virus Does Not Correlate with the Production of Viral Small RNAs. Virology 2014, 468–470, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Bergua, M.; Kang, S.-H.; Folimonova, S.Y. Understanding Superinfection Exclusion by Complex Populations of Citrus Tristeza Virus. Virology 2016, 499, 331–339. [Google Scholar] [CrossRef]
- Della Bartola, M.; Byrne, S.; Mullins, E. Characterization of Potato Virus Y Isolates and Assessment of Nanopore Sequencing to Detect and Genotype Potato Viruses. Viruses 2020, 12, 478. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-F.; Zhang, S.; Guo, Q.; Sun, R.; Wei, T.; Qu, F. A New Mechanistic Model for Viral Cross Protection and Superinfection Exclusion. Front. Plant Sci. 2018, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Ziebell, H.; Carr, J.P. Cross-Protection. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2010; Volume 76, pp. 211–264. ISBN 978-0-12-374525-5. [Google Scholar]
- Folimonova, S.Y. Superinfection Exclusion Is an Active Virus-Controlled Function That Requires a Specific Viral Protein. J. Virol. 2012, 86, 5554–5561. [Google Scholar] [CrossRef] [Green Version]
- Simón-Mateo, C.; García, J.A. Antiviral Strategies in Plants Based on RNA Silencing. Biochim. Biophys. Acta BBA—Gene Regul. Mech. 2011, 1809, 722–731. [Google Scholar] [CrossRef] [Green Version]
- Kung, Y.-J.; Lin, P.-C.; Yeh, S.-D.; Hong, S.-F.; Chua, N.-H.; Liu, L.-Y.; Lin, C.-P.; Huang, Y.-H.; Wu, H.-W.; Chen, C.-C.; et al. Genetic Analyses of the FRNK Motif Function of Turnip Mosaic Virus Uncover Multiple and Potentially Interactive Pathways of Cross-Protection. Mol. Plant-Microbe Interact. 2014, 27, 944–955. [Google Scholar] [CrossRef] [Green Version]
- Mesel, F.; Zhao, M.; García, B.; Simón-Mateo, C.; García, J.A. Targeting of Genomic and Negative-sense Strands of Viral RNA Contributes to Antiviral Resistance Mediated by Artificial miRNAs and Promotes the Emergence of Complex Viral Populations. Mol. Plant Pathol. 2022, 23, 1640–1657. [Google Scholar] [CrossRef]
- Maas, J.L. (Ed.) Compendium of Strawberry Diseases, 2nd ed.; The American Phytopathological Society: St. Paul, MN, USA, 1998. [Google Scholar]
- Martin, R.R.; Tzanetakis, I.E. Characterization and Recent Advances in Detection of Strawberry Viruses. Plant Dis. 2006, 90, 384–396. [Google Scholar] [CrossRef] [Green Version]
- Tzanetakis, I.E.; Martin, R.R. Expanding Field of Strawberry Viruses Which Are Important in North America. Int. J. Fruit Sci. 2013, 13, 184–195. [Google Scholar] [CrossRef]
- Bonneau, P.; Hogue, R.; Tellier, S.; Fournier, V. Evaluation of Various Sources of Viral Infection in Strawberry Fields of Quebec, Canada. J. Econ. Entomol. 2019, 112, 2577–2583. [Google Scholar] [CrossRef]
- Diaz-Lara, A.; Stevens, K.A.; Klaassen, V.; Hwang, M.S.; Al Rwahnih, M. Sequencing a Strawberry Germplasm Collection Reveals New Viral Genetic Diversity and the Basis for New RT-QPCR Assays. Viruses 2021, 13, 1442. [Google Scholar] [CrossRef]
- Torrico, A.K.; Salazar, S.M.; Kirschbaum, D.S.; Conci, V.C. Yield Losses of Asymptomatic Strawberry Plants Infected with Strawberry Mild Yellow Edge Virus. Eur. J. Plant Pathol. 2018, 150, 983–990. [Google Scholar] [CrossRef]
- Valentova, L.; Rejlova, M.; Franova, J.; Cmejla, R. Symptomless Infection by Strawberry Virus 1 (StrV-1) Leads to Losses in Strawberry Yields. Plant Pathol. 2022, 71, 1220–1228. [Google Scholar] [CrossRef]
- Fránová, J.; Přibylová, J.; Koloniuk, I. Molecular and Biological Characterization of a New Strawberry Cytorhabdovirus. Viruses 2019, 11, 982. [Google Scholar] [CrossRef] [Green Version]
- Fránová, J.; Lenz, O.; Přibylová, J.; Čmejla, R.; Valentová, L.; Koloniuk, I. High Incidence of Strawberry Polerovirus 1 in the Czech Republic and Its Vectors, Genetic Variability and Recombination. Viruses 2021, 13, 2487. [Google Scholar] [CrossRef]
- Koloniuk, I.; Fránová, J.; Sarkisova, T.; Přibylová, J. Complete Genome Sequences of Two Divergent Isolates of Strawberry Crinkle Virus Coinfecting a Single Strawberry Plant. Arch. Virol. 2018, 163, 2539–2542. [Google Scholar] [CrossRef] [PubMed]
- Koloniuk, I.; Matyášová, A.; Brázdová, S.; Veselá, J.; Přibylová, J.; Fránová, J.; Elena, S.F. Transmission of Diverse Variants of Strawberry Viruses Is Governed by a Vector Species. Viruses 2022, 14, 1362. [Google Scholar] [CrossRef] [PubMed]
- Bhagwat, B.; Dickison, V.; Ding, X.; Walker, M.; Bernardy, M.; Bouthillier, M.; Creelman, A.; DeYoung, R.; Li, Y.; Nie, X.; et al. Genome Sequence Analysis of Five Canadian Isolates of Strawberry Mottle Virus Reveals Extensive Intra-Species Diversity and a Longer RNA2 with Increased Coding Capacity Compared to a Previously Characterized European Isolate. Arch. Virol. 2016, 161, 1657–1663. [Google Scholar] [CrossRef] [PubMed]
- Cieślińska, M. Genetic Diversity of Seven Strawberry Mottle Virus Isolates in Poland. Plant Pathol. J. 2019, 35, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; He, C.; Wu, M.; Gao, D.; Dong, Z.; Hou, S.; Feng, Z.; Wang, H. Incidence, Genomic Diversity, and Evolution of Strawberry Mottle Virus in China. Biocell 2021, 45, 1137–1151. [Google Scholar] [CrossRef]
- Shanks, C.H. Seasonal Populations of the Strawberry Aphid and Transmission of Strawberry Viruses in the Field in Relation to Virus Control in Western Washington1. J. Econ. Entomol. 1965, 58, 316–322. [Google Scholar] [CrossRef]
- Shao, Y.; Chan, C.Y.; Maliyekkel, A.; Lawrence, C.E.; Roninson, I.B.; Ding, Y. Effect of Target Secondary Structure on RNAi Efficiency. RNA 2007, 13, 1631–1640. [Google Scholar] [CrossRef] [Green Version]
- Simon, A.E.; Miller, W.A. 3′ Cap-Independent Translation Enhancers of Plant Viruses. Annu. Rev. Microbiol. 2013, 67, 21–42. [Google Scholar] [CrossRef] [Green Version]
- Bera, S.; Ilyas, M.; Mikkelsen, A.A.; Simon, A.E. Conserved Structure Associated with Different 3′CITEs Is Important for Translation of Umbraviruses. Viruses 2023, 15, 638. [Google Scholar] [CrossRef]
- Miozzi, L.; Pantaleo, V.; Burgyán, J.; Accotto, G.P.; Noris, E. Analysis of Small RNAs Derived from Tomato Yellow Leaf Curl Sardinia Virus Reveals a Cross Reaction between the Major Viral Hotspot and the Plant Host Genome. Virus Res. 2013, 178, 287–296. [Google Scholar] [CrossRef]
- Murad, L.; Bielawski, J.P.; Matyasek, R.; Kovarík, A.; Nichols, R.A.; Leitch, A.R.; Lichtenstein, C.P. The Origin and Evolution of Geminivirus-Related DNA Sequences in Nicotiana. Heredity 2004, 92, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Kircher, M.; Sawyer, S.; Meyer, M. Double Indexing Overcomes Inaccuracies in Multiplex Sequencing on the Illumina Platform. Nucleic Acids Res. 2012, 40, e3. [Google Scholar] [CrossRef] [Green Version]
- Kutnjak, D.; Tamisier, L.; Adams, I.; Boonham, N.; Candresse, T.; Chiumenti, M.; De Jonghe, K.; Kreuze, J.F.; Lefebvre, M.; Silva, G.; et al. A Primer on the Analysis of High-Throughput Sequencing Data for Detection of Plant Viruses. Microorganisms 2021, 9, 841. [Google Scholar] [CrossRef]
- Zhao, D.; Song, G. High-Throughput Sequencing as an Effective Approach in Profiling Small RNAs Derived from a Hairpin RNA Expression Vector in Woody Plants. Plant Sci. 2014, 228, 39–47. [Google Scholar] [CrossRef]
- Fuentes, A.; Carlos, N.; Ruiz, Y.; Callard, D.; Sánchez, Y.; Ochagavía, M.E.; Seguin, J.; Malpica-López, N.; Hohn, T.; Lecca, M.R.; et al. Field Trial and Molecular Characterization of RNAi-Transgenic Tomato Plants That Exhibit Resistance to Tomato Yellow Leaf Curl Geminivirus. Mol. Plant-Microbe Interact. 2016, 29, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Simon, A.E.; Roossinck, M.J.; Havelda, Z. Plant Virus Satellite and Defective Interfering RNAs: New Paradigms for a New Century. Annu. Rev. Phytopathol. 2004, 42, 415–437. [Google Scholar] [CrossRef] [Green Version]
- Várallyay, É.; Oláh, E.; Havelda, Z. Independent Parallel Functions of P19 Plant Viral Suppressor of RNA Silencing Required for Effective Suppressor Activity. Nucleic Acids Res. 2014, 42, 599–608. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; He, C.; Gao, D.; Xu, T.; Xing, F.; Yan, J.; Zhan, B.; Li, S.; Wang, H. Identification of Silencing Suppressor Protein Encoded by Strawberry Mottle Virus. Front. Plant Sci. 2022, 13, 786489. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2014. [Google Scholar]
- Zhang, Y.; Peng, X.; Liu, Y.; Li, Y.; Luo, Y.; Wang, X.; Tang, H. Evaluation of Suitable Reference Genes for QRT-PCR Normalization in Strawberry (Fragaria × Ananassa) under Different Experimental Conditions. BMC Mol. Biol. 2018, 19, 8. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Strain | Plant | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1B | 2A | 3K | ||||||||||
| 1B_1 | 1B_2 | 1B_3 | 1B_4 | 2A_2 | 2A_3 | 2A_4 | 3K_1 | 3K_2 | 3K_3 | 3K_4 | ||
| SMoV | RNA1A | 1087 | 772 | 732 | 722 | 799 | 750 | 606 | 846 | 906 | 564 | 927 |
| RNA1B | 1013 | 712 | 686 | 656 | 716 | 649 | 537 | 751 | 767 | 508 | 818 | |
| RNA1C | 1064 | 744 | 709 | 671 | 777 | 724 | 581 | 823 | 829 | 531 | 884 | |
| RNA2A | 715 | 507 | 506 | 487 | 472 | 431 | 381 | 533 | 544 | 353 | 549 | |
| RNA2B | 615 | 406 | 416 | 418 | 407 | 418 | 309 | 441 | 470 | 305 | 486 | |
| RNA2C | 955 | 589 | 599 | 626 | 607 | 581 | 463 | 606 | 612 | 421 | 672 | |
| SCV | A | 781 | 152 | 255 | 701 | 1319 | 1091 | 565 | 974 | 1122 | 1389 | 1674 |
| B | 813 | 105 | 137 | 346 | 1081 | 1072 | 434 | 659 | 720 | 657 | 1161 | |
| StrV-1 | A | 265 | 171 | 114 | 138 | 772 | 770 | 301 | 286 | 394 | 244 | 398 |
| B | 588 | 427 | 420 | 356 | 1300 | 1647 | 834 | 727 | 1115 | 642 | 872 | |
| C | 116 | 129 | 66 | 111 | 503 | 423 | 143 | 237 | 276 | 200 | 273 | |
| Plant | Viruses | Obtained Via | Libraries |
|---|---|---|---|
| 1B | SCV-A, -B, SMoV RNA1ABC, RNA2ABC, StrV-1-A, -B, -C | Stolon propagation | 1B1, 1B2, 1B3, 1B4 |
| 2A | --‘’-- | --‘’-- | 2A2, 2A3, 2A4 |
| 3K | --‘’-- | --‘’-- | 3K1, 3K2, 3K3, 3K4 |
| Fv_StrV-1_A | StrV-1-A | Aphid transmission | S98, S95 |
| Fv_StrV-1_B | StrV-1-B | --‘’-- | S107, S108, S99 |
| Fv_StrV-1_AB | StrV-1-A, -B | Wedge grafting petiole | S101 |
| Fv-SMoV_BC_AC-1 | SMoV RNA1BC, RNA2AC | Aphid transmission | S1, S2 |
| Fv-SMoV_BC_AC-2 | --‘’-- | --‘’-- | S3, S4, S92 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koloniuk, I.; Matyášová, A.; Brázdová, S.; Veselá, J.; Přibylová, J.; Várallyay, E.; Fránová, J. Analysis of Virus-Derived siRNAs in Strawberry Plants Co-Infected with Multiple Viruses and Their Genotypes. Plants 2023, 12, 2564. https://doi.org/10.3390/plants12132564
Koloniuk I, Matyášová A, Brázdová S, Veselá J, Přibylová J, Várallyay E, Fránová J. Analysis of Virus-Derived siRNAs in Strawberry Plants Co-Infected with Multiple Viruses and Their Genotypes. Plants. 2023; 12(13):2564. https://doi.org/10.3390/plants12132564
Chicago/Turabian StyleKoloniuk, Igor, Alena Matyášová, Sára Brázdová, Jana Veselá, Jaroslava Přibylová, Eva Várallyay, and Jana Fránová. 2023. "Analysis of Virus-Derived siRNAs in Strawberry Plants Co-Infected with Multiple Viruses and Their Genotypes" Plants 12, no. 13: 2564. https://doi.org/10.3390/plants12132564
APA StyleKoloniuk, I., Matyášová, A., Brázdová, S., Veselá, J., Přibylová, J., Várallyay, E., & Fránová, J. (2023). Analysis of Virus-Derived siRNAs in Strawberry Plants Co-Infected with Multiple Viruses and Their Genotypes. Plants, 12(13), 2564. https://doi.org/10.3390/plants12132564