
Seed Germination Ecology of the Medicinal Plant Peganum harmala (Zygophyllaceae)

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Seed Characteristics
2.2. Imbibition Test
2.3. Effects of Light and Temperature on Germination
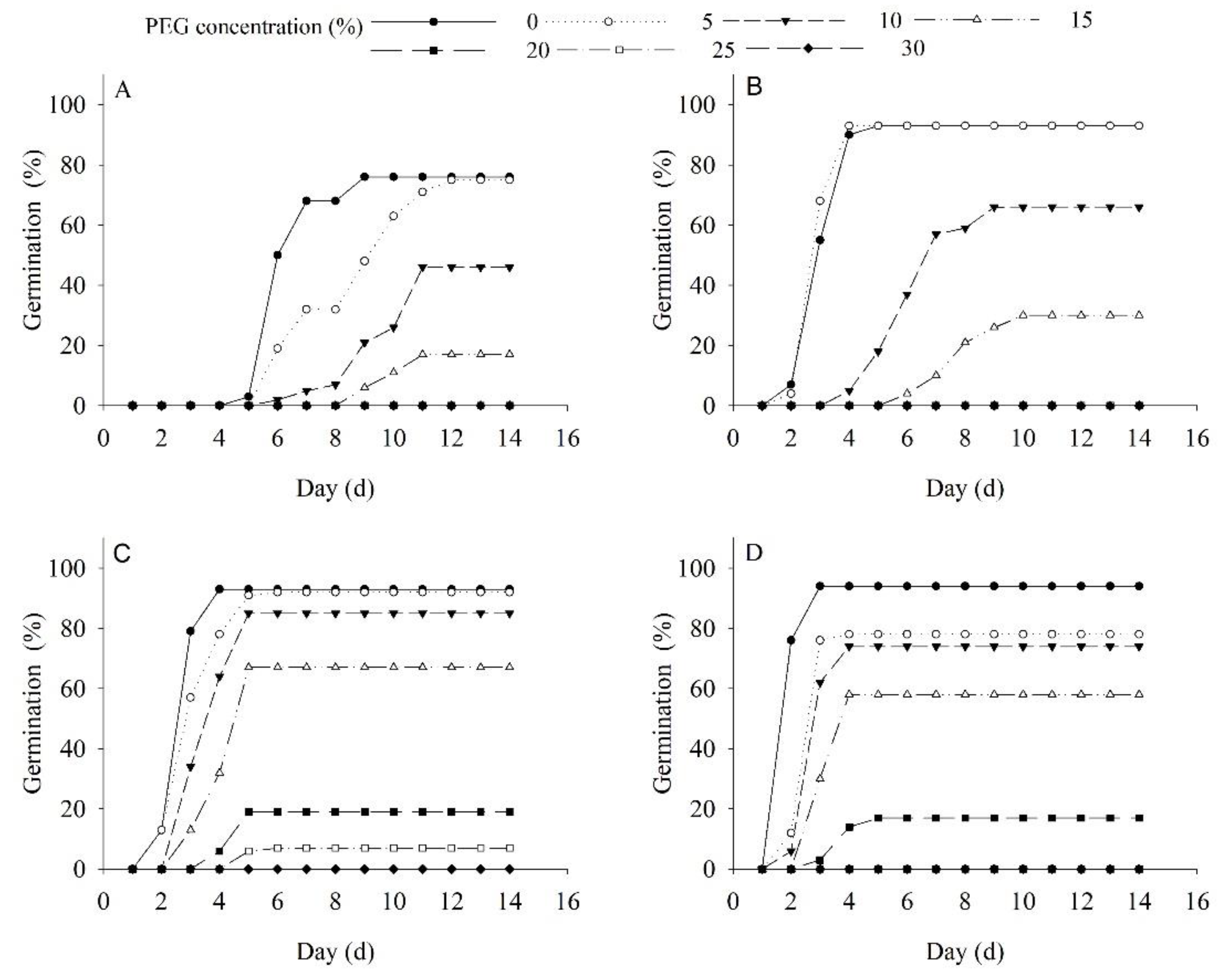
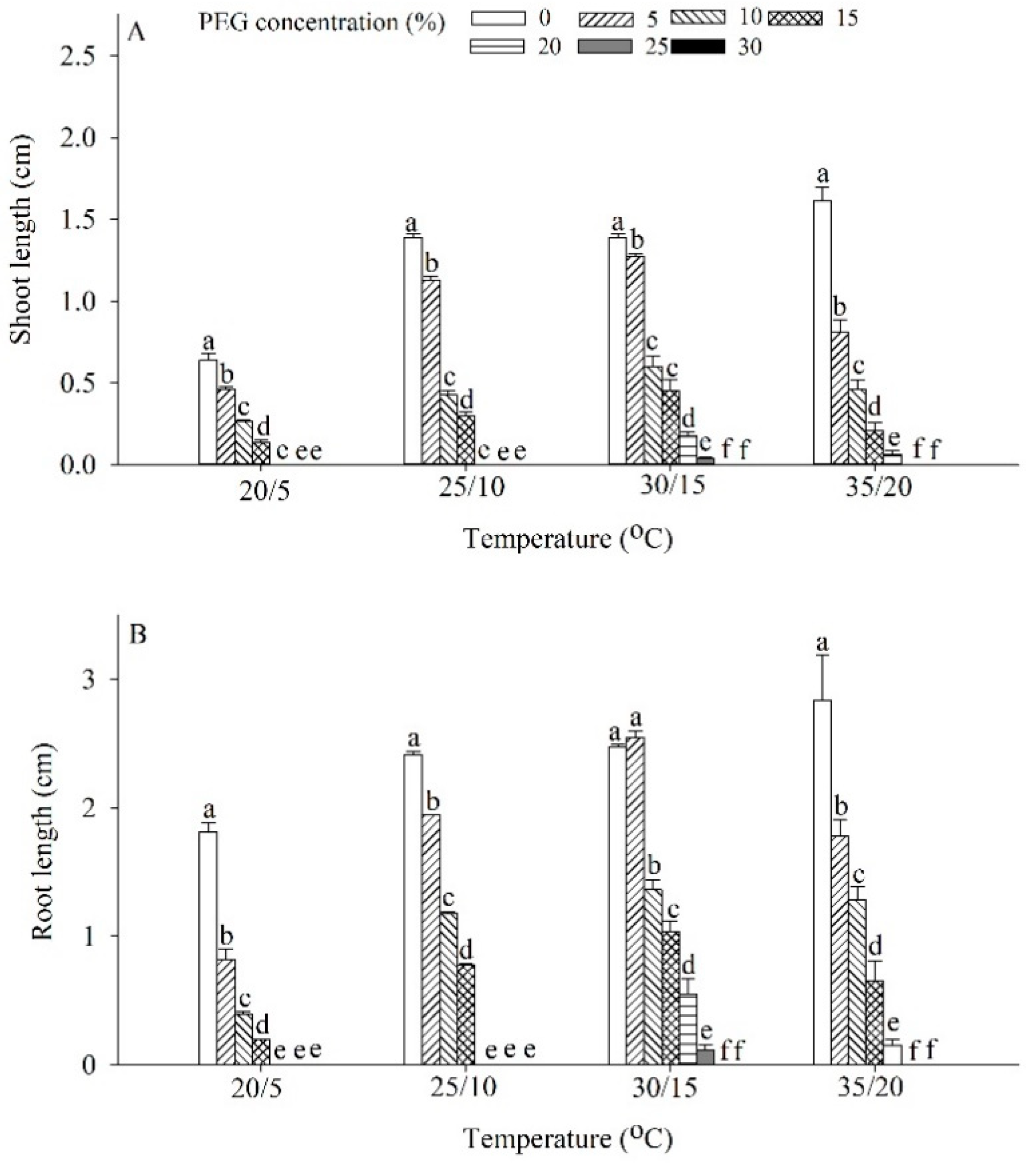
2.4. Effects of Drought and Temperature on Germination and Seedling Growth
2.5. Effects of Salinity on Germination and Seedling Growth
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Study Species
5.2. Seed Collection
5.3. Seed Characteristics
5.4. Imbibition Test
5.5. Effects of Light and Temperature on Germination
5.6. Effects of Drought and Temperature on Germination and Seedling Growth
5.7. Effects of Salinity on Germination and Seedling Growth
5.8. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nwozo, O.S.; Effiong, E.M.; Aja, P.M.; Awuchi, C.G. Antioxidant, phytochemical, and therapeutic properties of medicinal plants: A review. Int. J. Food Prop. 2023, 26, 359–388. [Google Scholar] [CrossRef]
- Bhatt, A.; Caron, M.M.; Chen, X.X.; Yu, D.S.; Niu, Y.L. Effect of temperature, light and storage on seed germination of Salvia plebeia R.Br., Leonurus japonicus Houtt., Mosla scabra (Thunb.) C.Y.Wu & H.W.Li and Perilla frutescens (L.) Britton. J. Appl. Res. Med. Arom. Plant. 2022, 31, 100402. [Google Scholar]
- Sharma, B.; Yadav, D.K. Metabolomics and network pharmacology in the exploration of the multi-targeted therapeutic approach of traditional medicinal plants. Plants 2022, 11, 3243. [Google Scholar] [CrossRef] [PubMed]
- Rashid, S.; Sameti, M.; Alqarni, M.H.; Bar, F.M.A. In vivo investigation of the inhibitory effect of Peganum harmala L. and its major alkaloids on ethylene glycol-induced urolithiasis in rats. J. Ethnopharmacol. 2023, 300, 115752. [Google Scholar] [CrossRef]
- Flora of China Editorial Committee. Flora of China; Science Press and Missouri Botanical Garden Press: Beijing, China, 2008; Volume 11. [Google Scholar]
- Baranova, E.N.; Gulevich, A.A.; Polyakov, V.Y. Effect of NaCl, Na2SO4, and mannitol on utilization of storage starch and formation of plastids in the cotyledons and roots of alfalfa seedlings. Russ. J. Plant Physiol. 2007, 54, 50–57. [Google Scholar] [CrossRef]
- Baranova, E.N.; Gulevich, A.A.; Polyakov, V.Y. Effects of NaCl, Na2SO4, and mannitol on storage lipid mobilization in the cotyledons and roots of purple alfalfa seedlings. Russ. J. Plant Physiol. 2006, 53, 779–784. [Google Scholar] [CrossRef]
- El-Keblawy, A. Light and temperature requirements during germination of potential perennial grasses for rehabilitation of degraded sandy Arabian deserts. Land Degrad. Dev. 2017, 28, 1687–1695. [Google Scholar] [CrossRef]
- El-Keblawy, A.; Al-Shamsi, N.; Maso, K.A. Effect of maternal habitat, temperature and light on germination and salt tolerance of Suaeda vermiculata, a habitat-indifferent halophyte of arid Arabian deserts. Seed Sci. Res. 2018, 28, 140–147. [Google Scholar] [CrossRef]
- Rasool, S.G.; Hameed, A.; Khan, M.A.; Gul, B. Seeds of Halopeplis perfoliata display plastic responses to various abiotic factors during germination. Flora 2017, 236, 76–83. [Google Scholar] [CrossRef]
- Huang, Z.Y.; Zhang, X.S.; Zheng, G.H.; Gutterman, Y. Influence of light, temperature, salinity and storage on seed germination of Haloxylon ammodendron. J. Arid Environ. 2003, 55, 453–464. [Google Scholar] [CrossRef]
- Yan, A.; Chen, Z. The control of seed dormancy and germination by temperature, light and nitrate. Bot. Rev. 2020, 86, 39–75. [Google Scholar] [CrossRef]
- Rasheed, A.; Hameed, A.; Gul, B.; Khan, M.A. Perianth and abiotic factors regulate seed germination of Haloxylon stocksii—A cash crop candidate for degraded saline lands. Land Degrad. Dev. 2019, 30, 1468–1478. [Google Scholar] [CrossRef]
- Gul, B.; Ansari, R.; Flowers, T.J.; Khan, M.A. Germination strategies of halophyte seeds under salinity. Environ. Exp. Bot. 2013, 92, 4–18. [Google Scholar] [CrossRef]
- Koutsovoulou, K.; Daws, W.I.; Thanos, C.A. Campanulaceae: A family with small seeds that require light for germination. Ann. Bot. 2014, 113, 135–143. [Google Scholar] [CrossRef]
- Wang, Y.X.; Li, Z.J.; Jiao, P.P. Study the effects of temperature and light on seeds germination characteristics. J. Tarim Univ. 2009, 21, 15–19. (In Chinese) [Google Scholar]
- Bewley, J.D.; Black, M. Seeds: Physiology of Development and Germination, 2nd ed.; Plenum Press: New York, NY, USA; London, UK, 1994. [Google Scholar]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 201–523. [Google Scholar] [CrossRef] [PubMed]
- Baskin, J.M.; Davis, B.H.; Baskin, C.C.; Gleason, S.M.; Cordell, S. Physical dormancy in seeds of Dodonaea viscosa (Sapindales, Sapindaceae) from Hawaii. Seed Sci. Res. 2004, 14, 81–90. [Google Scholar] [CrossRef]
- El-Sharkawi, H.M.; Springuel, I. Germination of some crop plant seeds under reduced water potential. Seed Sci. Technol. 1977, 5, 677–688. [Google Scholar]
- Hu, R.H.; Liu, H.; Liu, F.H. Seed germination of hemp (Cannabis sativa L.) cultivars responds differently to the stress of salt type and concentration. Ind. Crop. Prod. 2018, 123, 254–261. [Google Scholar] [CrossRef]
- Ahmed, M.Z.; Khan, M.A. Tolerance and recovery responses of playa halophytes to light, salinity and temperature stresses during seed germination. Flora 2010, 205, 764–771. [Google Scholar] [CrossRef]
- Hou, M.C.; Ma, M. Effect of PEG-simulated drought stress on seed germination of three medicinal Liquorice (Glycyrrhiza) species. Legume Res. 2022, 45, 1388–1393. [Google Scholar] [CrossRef]
- Tang, D.F.; Wei, F.; Qin, S.X.; Khan, A.; Kashif, M.H.; Zhou, R.Y. Polyethylene glycol induced drought stress strongly influences seed germination, root morphology and cytoplasm of different kenaf genotypes. Ind. Crop Prod. 2019, 137, 180–186. [Google Scholar] [CrossRef]
- Fischer, R.A.; Turner, N.C. Plant productivity in arid and semi-arid zones. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1978, 29, 277–317. [Google Scholar] [CrossRef]
- Windauer, L.B.; Martinez, J.; Rapoport, D.; Wassner, D.; Benech-Arnold, R. Germination responses to temperature and water potential in Jatropha Curcas Seeds: A hydrotime model explains the difference between dormancy expression and dormancy induction at different incubation temperatures. Ann. Bot. 2012, 109, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.L.; Shi, J.J.; Liao, J.; Qi, X.L.; Wang, J.J.; Zhu, L.J.; Wang, S.M. Study on seed characteristics, germination condition and the tolerant ability to salt and drought of Peganum harmala. J. Anhui Agric. Sci. 2010, 38, 3448–3450. (In Chinese) [Google Scholar]
- Basahi, M.A. Interactions in germination responses to salinity, temperature and light in the desert shrub African Rue (Peganum harmala L.). Appl. Ecol. Environ. Res. 2022, 12, 85–98. [Google Scholar] [CrossRef]
- Ertugrul, O.; Yilar, M.; Kir, H.; Komekci, C. Some physical, chemical, and germination properties of Peganum harmala L. seeds. J. Food Process. Eng. 2022, 45, e13967. [Google Scholar] [CrossRef]
- Xu, H.Q.; Feng, Y.M.; He, M.Z.; Wang, L.; Hu, T.G.; Cheng, B.R.; Nie, K.H. Effect of different temperature and light on seed germination of Peganum harmala L. Seed 2011, 30, 74–76. (In Chinese) [Google Scholar]
- Karakas, S. Biochemical responses and salt removal potential of Peganum harmala L. (Wild Rue) under different NaCl conditions. Appl. Ecol. Environ. Res. 2020, 18, 4353–4369. [Google Scholar] [CrossRef]
- Karam, M.A.; Abd-Elgawad, M.E.; Ali, R.M. Differential gene expression of salt-stressed Peganum harmala L. J. Gen. Eng. Biotech. 2016, 14, 319–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elallem, K.A.; Ben Bakrim, W.; Ennoury, A.; Metougui, M.L.; Yasri, A.; Boularbah, A. Germination parameters and responses of antioxidant enzymatic activities of two medicinal plants (Peganum harmala L. and Origanum majorana L.) under heavy metal stress. J. Soil Sci. Plant Nutr. 2022, 22, 3942–3957. [Google Scholar] [CrossRef]
- Jiang, N.; Chen, L.; Li, J.M.; Li, W.Y.; Jiang, S.L. Lethal and sublethal toxicity of beta-carboline alkaloids from Peganum harmala (L.) against aedes albopictus larvae (Diptera: Culicidae). Toxics 2023, 11, 341. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zan, Y.H.; Yang, H.G.; Yang, M.Y.; Liu, F.S.; Li, S.G.; Peng, X.H.; Lin, B.; Li, Z.L.; Li, D.H.; et al. Anti-tumor alkaloids from Peganum harmala. Phytochemistry 2022, 197, 113107. [Google Scholar] [CrossRef] [PubMed]
- Effect of water deficit on germination, growth and biochemical responses of four potentially invasive ornamental grass species. Plants 2023, 12, 1260. [CrossRef]
- Guaraldo, M.M.D.; Pereira, T.M.; dos Santos, H.O.; de Oliveira, T.L.; Pereira, W.V.S.; Von Pinho, E.V.D. Priming with sodium nitroprusside and hydrogen peroxide increases cotton seed tolerance to salinity and water deficit during seed germination and seedling development. Environ. Exp. Bot. 2023, 209, 105294. [Google Scholar] [CrossRef]
- Moghaddam, M.; Babaei, K.; Pooya, E.S. Germination and growth response of flax (Linum usitatissimum) to salinity stress by different salt types and concentrations. J. Plant Nutr. 2018, 41, 563–573. [Google Scholar] [CrossRef]
- Hadjadj, S.; Sekerifa, B.B.; Khellafi, H.; Krama, K.; Rahmani, S.; El Hadj-Khelil, A.O. Salinity and type of salt effects on seed germination characteristics of nmedicinal plant Zygophyllum album L. (Zygophyllaceae) native to the Algerian Sahara. J. Appl. Res. Med. Arom. Plant. 2022, 31, 100412. [Google Scholar]
- Munns, R.; Gilliham, M. Salinity tolerance of crops–what is the cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Baranova, E.N.; Gulevich, A.A. Asymmetry of plant cell divisions under salt stress. Symmetry 2021, 13, 1811. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature | Regression Equation | Correlation Coefficient | Critical Value PEG (%) | Limit Value PEG (%) |
|---|---|---|---|---|
| 20/5 °C | y = −914.2857x2 − 237.1429x + 80.2288 | 0.9592 | 9.37% | 19.37% |
| 25/10 °C | y =−1971.4286x2 − 103.7143x + 96.3429 | 0.9745 | 14.48% | 19.63% |
| 30/15 °C | y = −1564.2857x2 + 9.9286x + 95.1071 | 0.9377 | 17.30% | 24.98% |
| 35/20 °C | y =−1092.8571x2 − 109.0714x + 92.1786 | 0.9578 | 15.28% | 24.48% |
| Salinity Type | Regression Equation | Correlation Coefficient | Critical Value (mmol L−1) | Limit Value (mmol L−1) |
|---|---|---|---|---|
| NaCl | y = −0.0004x2 − 0.2022x + 97.8060 | 0.9847 | 177.97 | 299.21 |
| Na2SO4 | y = −4720x + 97.6000 | 0.9673 | 100.85 | 206.78 |
| NaHCO3 | y = −0.4030x + 97.8250 | 0.9646 | 106.27 | 230.34 |
| Na2CO3 | y = −0.8937x + 98.6000 | 0.9649 | 54.38 | 110.32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Yan, N.; Tanveer, M.; Zhao, Z.; Jiang, L.; Wang, H. Seed Germination Ecology of the Medicinal Plant Peganum harmala (Zygophyllaceae). Plants 2023, 12, 2660. https://doi.org/10.3390/plants12142660
Li S, Yan N, Tanveer M, Zhao Z, Jiang L, Wang H. Seed Germination Ecology of the Medicinal Plant Peganum harmala (Zygophyllaceae). Plants. 2023; 12(14):2660. https://doi.org/10.3390/plants12142660
Chicago/Turabian StyleLi, Shifeng, Ning Yan, Mohsin Tanveer, Zhenyong Zhao, Li Jiang, and Hongling Wang. 2023. "Seed Germination Ecology of the Medicinal Plant Peganum harmala (Zygophyllaceae)" Plants 12, no. 14: 2660. https://doi.org/10.3390/plants12142660
APA StyleLi, S., Yan, N., Tanveer, M., Zhao, Z., Jiang, L., & Wang, H. (2023). Seed Germination Ecology of the Medicinal Plant Peganum harmala (Zygophyllaceae). Plants, 12(14), 2660. https://doi.org/10.3390/plants12142660