
Endosperm Persistence in Arabidopsis Results in Seed Coat Fractures and Loss of Seed Longevity
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
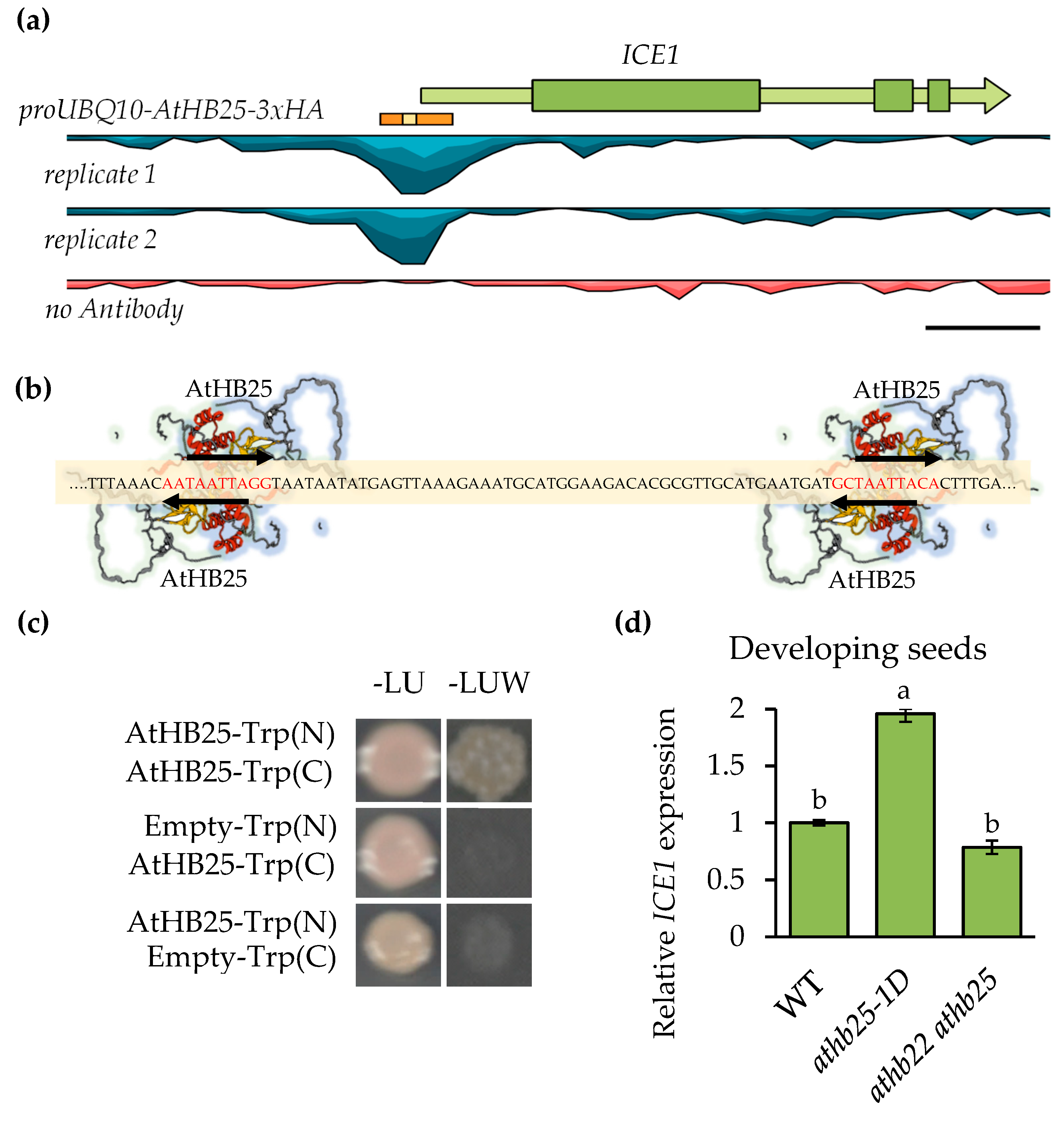
2.1. AtHB25 Regulates ICE1 Expression via Direct Promoter Binding
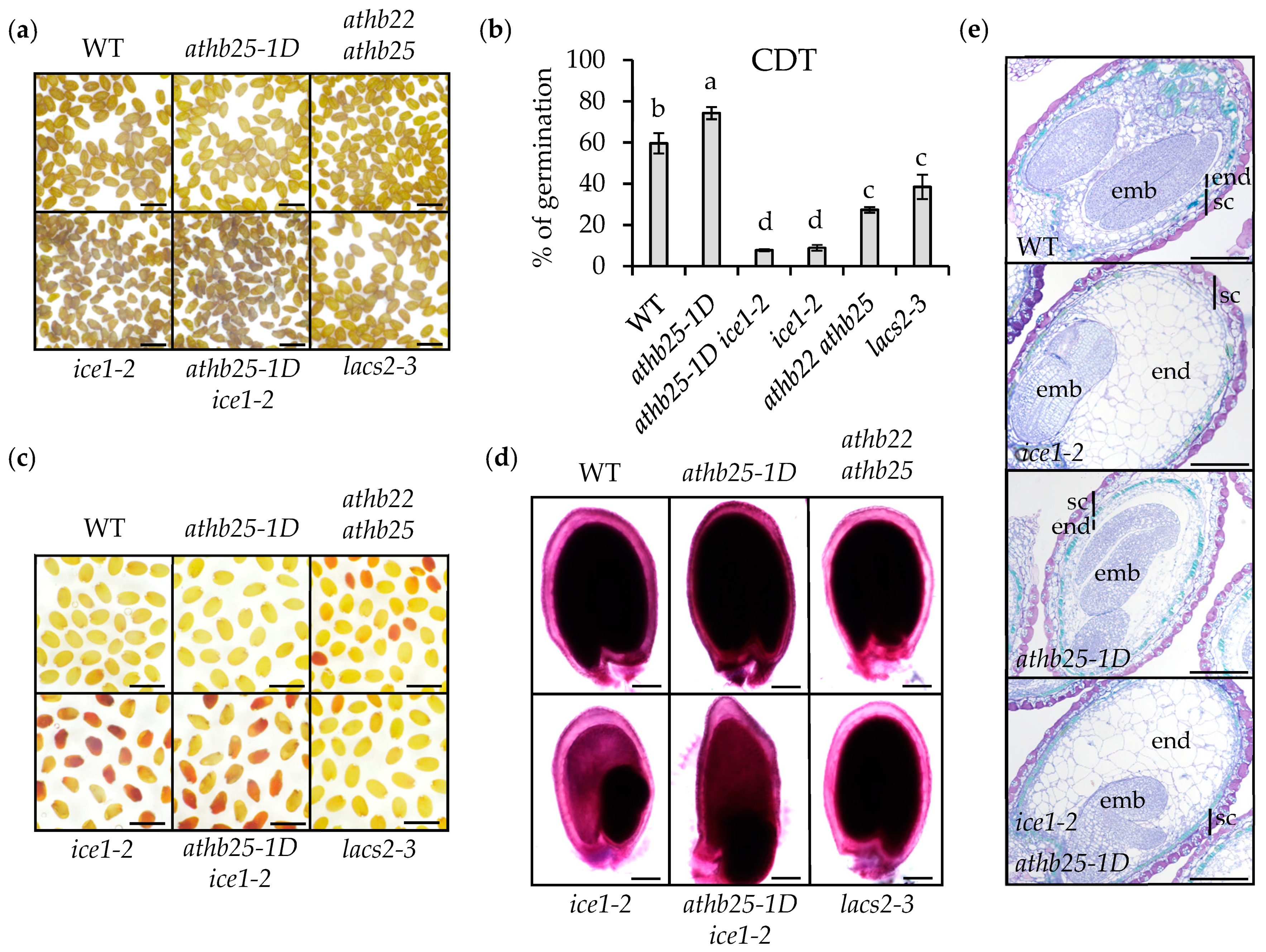
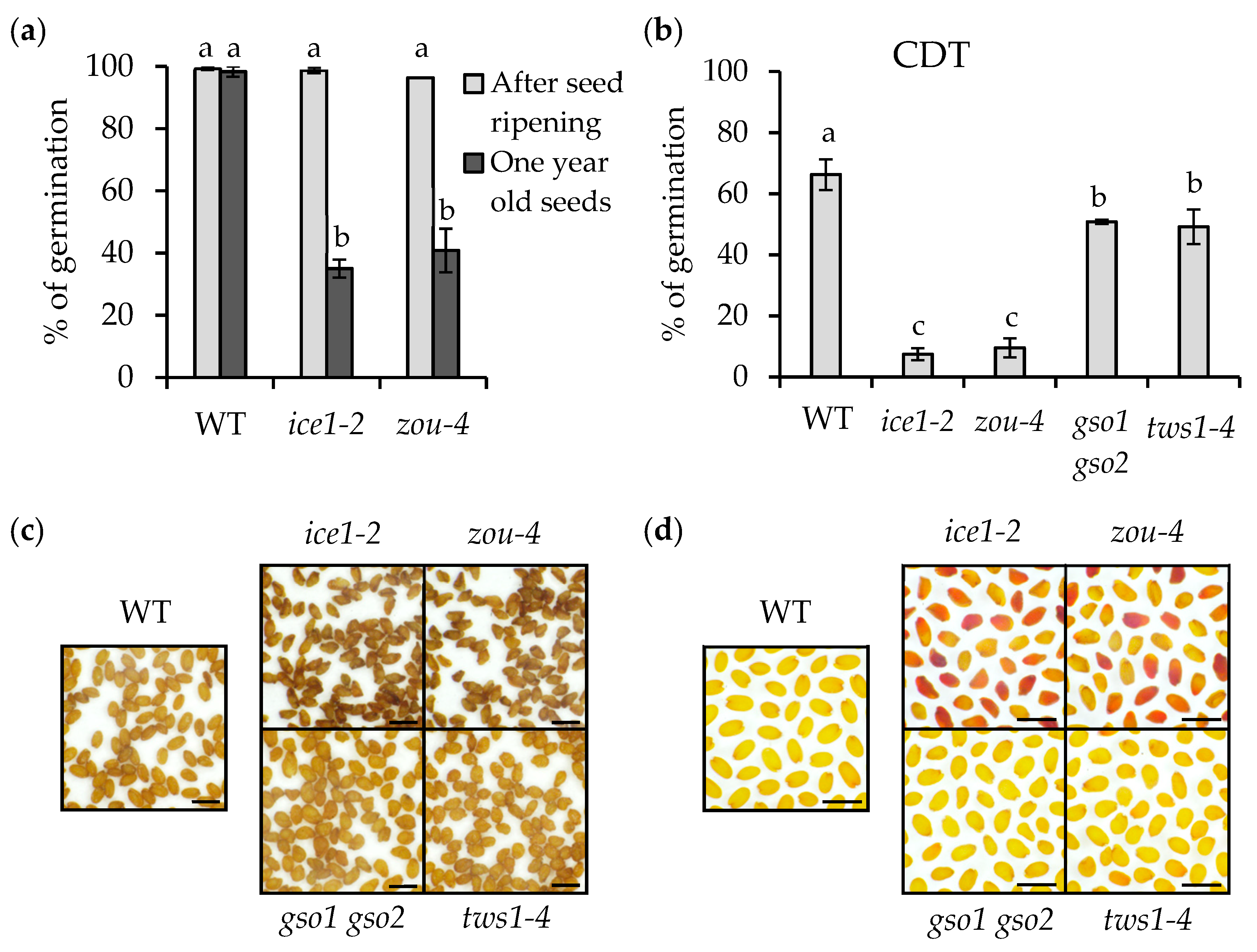
2.2. ICE1 Function Is Crucial for Seed Longevity
2.3. Endosperm Elimination Is Critical for Seed Longevity
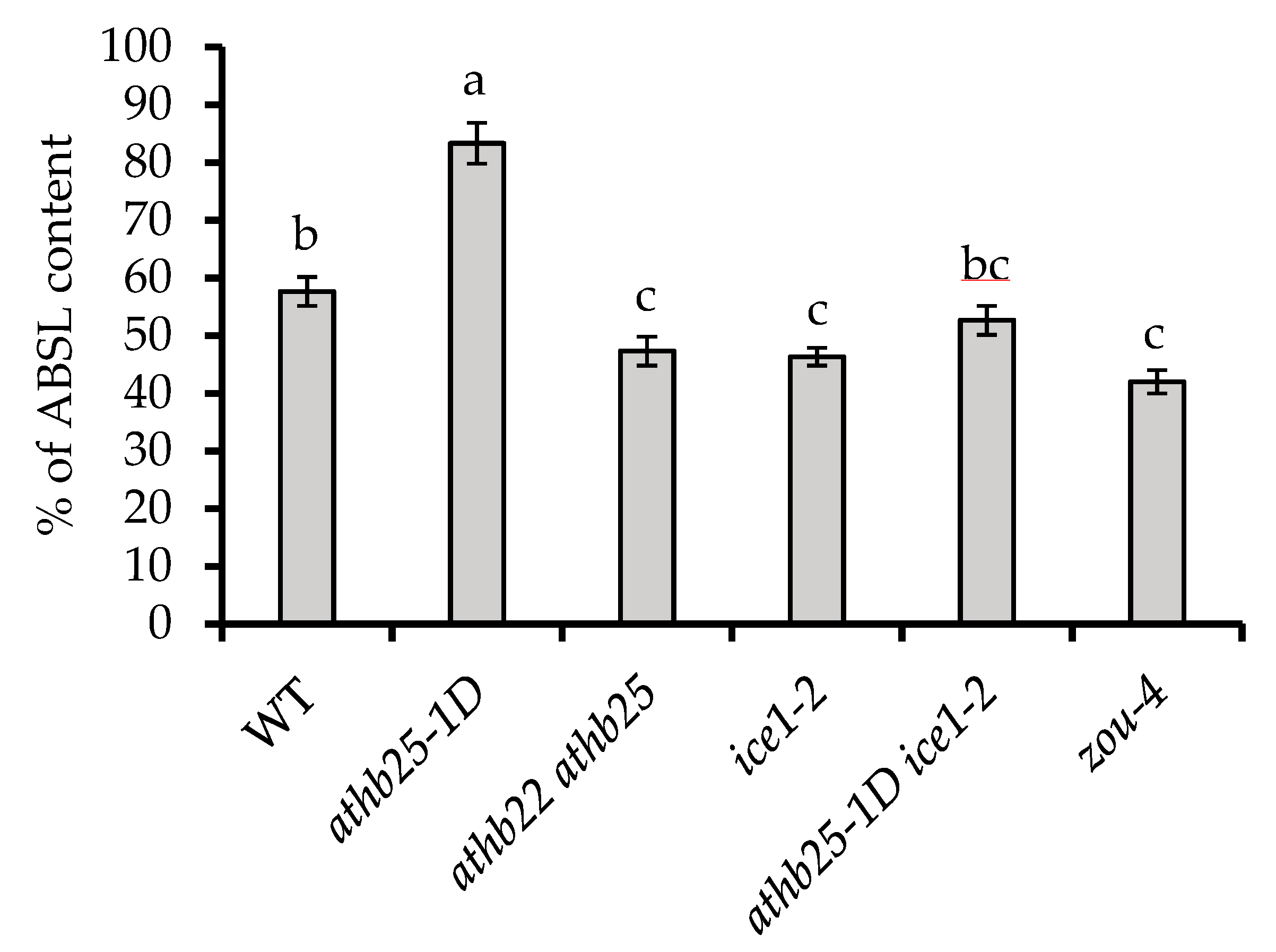
2.4. AtHB25 and ICE1 Regulate Polyphenolic Content in Seeds
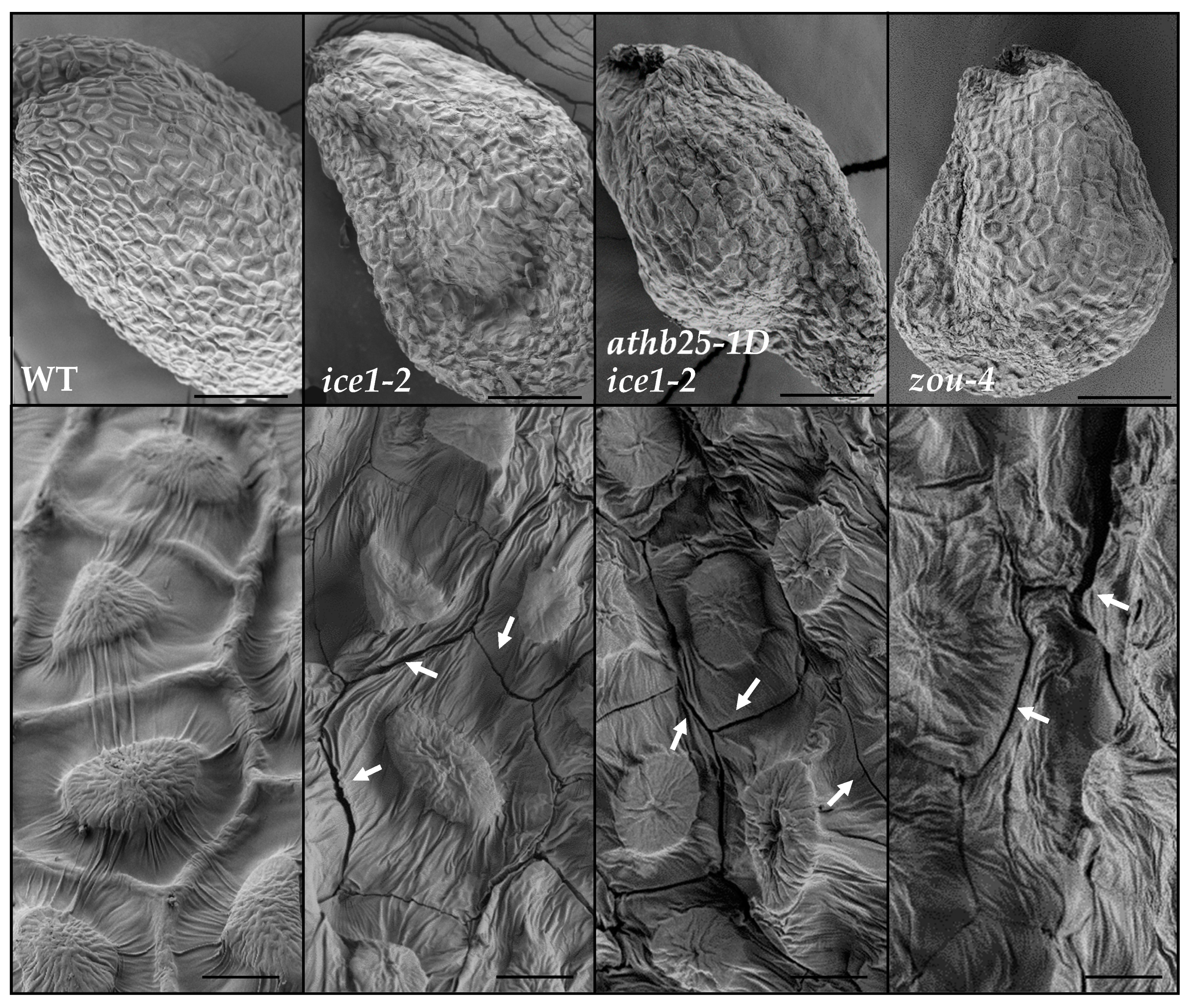
2.5. Endosperm Persistence Leads to Fissures in the Seed Coat
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Seed-Aging Treatments, Seed Dormancy Test, and Seed Germination
4.3. Seed Coat Analyses and Polyphenolic Analysis
4.4. Scanning Electron Microscopy
4.5. Gene Expression Analyses
4.6. Toluidine Blue Staining
4.7. Split-Trp
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berjak, P.; Pammenter, N.W. From Avicennia to Zizania: Seed Recalcitrance in Perspective. Ann. Bot. 2008, 101, 213–228. [Google Scholar] [CrossRef]
- Hoekstra, F.A.; Golovina, E.A.; Buitink, J. Mechanisms of Plant Desiccation Tolerance. Trends Plant Sci. 2001, 6, 431–438. [Google Scholar] [CrossRef]
- Haughn, G.; Chaudhury, A. Genetic Analysis of Seed Coat Development in Arabidopsis. Trends Plant Sci. 2005, 10, 472–477. [Google Scholar] [CrossRef]
- Debeaujon, I.; Léon-Kloosterziel, K.M.; Koornneef, M. Influence of the Testa on Seed Dormancy, Germination, and Longevity in Arabidopsis. Plant Physiol. 2000, 122, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Western, T.L. The Sticky Tale of Seed Coat Mucilages: Production, Genetics, and Role in Seed Germination and Dispersal. Seed Sci. Res. 2012, 22, 1–25. [Google Scholar] [CrossRef]
- Renard, J.; Martínez-Almonacid, I.; Sonntag, A.; Molina, I.; Moya-Cuevas, J.; Bissoli, G.; Muñoz-Bertomeu, J.; Faus, I.; Niñoles, R.; Shigeto, J.; et al. PRX2 and PRX25, Peroxidases Regulated by COG1, Are Involved in Seed Longevity in Arabidopsis. Plant Cell Environ. 2020, 43, 315–326. [Google Scholar] [CrossRef]
- Renard, J.; Martínez-Almonacid, I.; Queralta Castillo, I.; Sonntag, A.; Hashim, A.; Bissoli, G.; Campos, L.; Muñoz-Bertomeu, J.; Niñoles, R.; Roach, T.; et al. Apoplastic Lipid Barriers Regulated by Conserved Homeobox Transcription Factors Extend Seed Longevity in Multiple Plant Species. New Phytol. 2021, 231, 679–694. [Google Scholar] [CrossRef] [PubMed]
- Loubéry, S.; De Giorgi, J.; Utz-Pugin, A.; Demonsais, L.; Lopez-Molina, L. A Maternally Deposited Endosperm Cuticle Contributes to the Physiological Defects of Transparent Testa Seeds. Plant Physiol. 2018, 177, 1218–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkel-Shirley, B. Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanaoka, M.M.; Pillitteri, L.J.; Fujii, H.; Yoshida, Y.; Bogenschutz, N.L.; Takabayashi, J.; Zhu, J.-K.; Torii, K.U. SCREAM/ICE1 and SCREAM2 Specify Three Cell-State Transitional Steps Leading to Arabidopsis Stomatal Differentiation. Plant Cell 2008, 20, 1775–1785. [Google Scholar] [CrossRef] [Green Version]
- Chinnusamy, V.; Ohta, M.; Kanrar, S.; Lee, B.; Hong, X.; Agarwal, M.; Zhu, J.-K. ICE1: A Regulator of Cold-Induced Transcriptome and Freezing Tolerance in Arabidopsis. Genes Dev. 2003, 17, 1043–1054. [Google Scholar] [CrossRef] [Green Version]
- Le, B.H.; Cheng, C.; Bui, A.Q.; Wagmaister, J.A.; Henry, K.F.; Pelletier, J.; Kwong, L.; Belmonte, M.; Kirkbride, R.; Horvath, S.; et al. Global Analysis of Gene Activity during Arabidopsis Seed Development and Identification of Seed-Specific Transcription Factors. Proc. Natl. Acad. Sci. USA 2010, 107, 8063–8070. [Google Scholar] [CrossRef]
- Yang, S.; Johnston, N.; Talideh, E.; Mitchell, S.; Jeffree, C.; Goodrich, J.; Ingram, G. The Endosperm-Specific ZHOUPI Gene of Arabidopsis Thaliana Regulates Endosperm Breakdown and Embryonic Epidermal Development. Development 2008, 135, 3501–3509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denay, G.; Creff, A.; Moussu, S.; Wagnon, P.; Thévenin, J.; Gérentes, M.-F.; Chambrier, P.; Dubreucq, B.; Ingram, G. Endosperm Breakdown in Arabidopsis Requires Heterodimers of the Basic Helix-Loop-Helix Proteins ZHOUPI and INDUCER OF CBP EXPRESSION 1. Development 2014, 141, 1222–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Creff, A.; Brocard, L.; Joubès, J.; Taconnat, L.; Doll, N.M.; Marsollier, A.-C.; Pascal, S.; Galletti, R.; Boeuf, S.; Moussu, S.; et al. A Stress-Response-Related Inter-Compartmental Signalling Pathway Regulates Embryonic Cuticle Integrity in Arabidopsis. PLoS Genet. 2019, 15, e1007847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doll, N.M.; Royek, S.; Fujita, S.; Okuda, S.; Chamot, S.; Stintzi, A.; Widiez, T.; Hothorn, M.; Schaller, A.; Geldner, N.; et al. A Two-Way Molecular Dialogue between Embryo and Endosperm Is Required for Seed Development. Science 2020, 367, 431–435. [Google Scholar] [CrossRef]
- MacGregor, D.R.; Zhang, N.; Iwasaki, M.; Chen, M.; Dave, A.; Lopez-Molina, L.; Penfield, S. ICE1 and ZOU Determine the Depth of Primary Seed Dormancy in Arabidopsis Independently of Their Role in Endosperm Development. Plant J. 2019, 98, 277–290. [Google Scholar] [CrossRef] [Green Version]
- Bueso, E.; Muñoz-Bertomeu, J.; Campos, F.; Brunaud, V.; Martínez, L.; Sayas, E.; Ballester, P.; Yenush, L.; Serrano, R. ARABIDOPSIS THALIANA HOMEOBOX25 Uncovers a Role for Gibberellins in Seed Longevity. Plant Physiol. 2014, 164, 999–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bueso, E.; Muñoz-Bertomeu, J.; Campos, F.; Martínez, C.; Tello, C.; Martínez-Almonacid, I.; Ballester, P.; Simón-Moya, M.; Brunaud, V.; Yenush, L.; et al. Arabidopsis COGWHEEL1 Links Light Perception and Gibberellins with Seed Tolerance to Deterioration. Plant J. 2016, 87, 583–596. [Google Scholar] [CrossRef] [Green Version]
- Tan, Q.K.-G.; Irish, V.F. The Arabidopsis Zinc Finger-Homeodomain Genes Encode Proteins with Unique Biochemical Properties That Are Coordinately Expressed during Floral Development. Plant Physiol. 2006, 140, 1095–1108. [Google Scholar] [CrossRef] [Green Version]
- Tafelmeyer, P.; Johnsson, N.; Johnsson, K. Transforming a (β/α)8-Barrel Enzyme into a Split-Protein Sensor through Directed Evolution. Chem. Biol. 2004, 11, 681–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively Expanding the Structural Coverage of Protein-Sequence Space with High-Accuracy Models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- Bernards, M.A. Demystifying Suberin. Can. J. Bot. 2002, 80, 227–240. [Google Scholar] [CrossRef]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin Biosynthesis and Structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [Green Version]
- Liang, M.; Davis, E.; Gardner, D.; Cai, X.; Wu, Y. Involvement of AtLAC15 in Lignin Synthesis in Seeds and in Root Elongation of Arabidopsis. Planta 2006, 224, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Vilar, F.C.; Siqueira-Soares, R.d.C.; Finger-Teixeira, A.; Oliveira, D.M.d.; Ferro, A.P.; Rocha, G.J.d.; Ferrarese, M.d.L.L.; Santos, W.D.d.; Ferrarese-Filho, O. The Acetyl Bromide Method Is Faster, Simpler and Presents Best Recovery of Lignin in Different Herbaceous Tissues than Klason and Thioglycolic Acid Methods. PLoS ONE 2014, 9, e110000. [Google Scholar] [CrossRef] [PubMed]
- Xing, Q.; Creff, A.; Waters, A.; Tanaka, H.; Goodrich, J.; Ingram, G.C. ZHOUPI Controls Embryonic Cuticle Formation via a Signalling Pathway Involving the Subtilisin Protease ABNORMAL LEAF-SHAPE1 and the Receptor Kinases GASSHO1 and GASSHO2. Development 2013, 140, 770–779. [Google Scholar] [CrossRef] [Green Version]
- Tausch, S.; Leipold, M.; Reisch, C.; Poschlod, P. Dormancy and Endosperm Presence Influence the Ex Situ Conservation Potential in Central European Calcareous Grassland Plants. AoB Plants 2019, 11, plz035. [Google Scholar] [CrossRef] [PubMed]
- Merritt, D.J.; Martyn, A.J.; Ainsley, P.; Young, R.E.; Seed, L.U.; Thorpe, M.; Hay, F.R.; Commander, L.E.; Shackelford, N.; Offord, C.A.; et al. A Continental-Scale Study of Seed Lifespan in Experimental Storage Examining Seed, Plant, and Environmental Traits Associated with Longevity. Biodivers. Conserv. 2014, 23, 1081–1104. [Google Scholar] [CrossRef]
- Probert, R.J.; Daws, M.I.; Hay, F.R. Ecological Correlates of Ex Situ Seed Longevity: A Comparative Study on 195 Species. Ann. Bot. 2009, 104, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Galland, M.; He, D.; Lounifi, I.; Arc, E.; Clément, G.; Balzergue, S.; Huguet, S.; Cueff, G.; Godin, B.; Collet, B.; et al. An Integrated “Multi-Omics” Comparison of Embryo and Endosperm Tissue-Specific Features and Their Impact on Rice Seed Quality. Front. Plant Sci. 2017, 8, 1984. [Google Scholar] [CrossRef] [Green Version]
- Fourquin, C.; Beauzamy, L.; Chamot, S.; Creff, A.; Goodrich, J.; Boudaoud, A.; Ingram, G. Mechanical Stress Mediated by Both Endosperm Softening and Embryo Growth Underlies Endosperm Elimination in Arabidopsis Seeds. Development 2016, 143, 3300–3305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina, I.; Ohlrogge, J.B.; Pollard, M. Deposition and Localization of Lipid Polyester in Developing Seeds of Brassica Napus and Arabidopsis Thaliana. Plant J. 2008, 53, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Sato, H.; Bente, H.; Santos-González, J.; Köhler, C. Endosperm Cellularization Failure Induces a Dehydration-Stress Response Leading to Embryo Arrest. Plant Cell 2023, 35, 874–888. [Google Scholar] [CrossRef] [PubMed]
- De Giorgi, J.; Fuchs, C.; Iwasaki, M.; Kim, W.; Piskurewicz, U.; Gully, K.; Utz-Pugin, A.; Mène-Saffrané, L.; Waridel, P.; Nawrath, C.; et al. The Arabidopsis Mature Endosperm Promotes Seedling Cuticle Formation via Release of Sulfated Peptides. Dev. Cell 2021, 56, 3066–3081.e5. [Google Scholar] [CrossRef] [PubMed]
- Schnurr, J. The Acyl-CoA Synthetase Encoded by LACS2 Is Essential for Normal Cuticle Development in Arabidopsis. Plant Cell Online 2004, 16, 629–642. [Google Scholar] [CrossRef] [Green Version]
- Demonsais, L.; Utz-Pugin, A.; Loubéry, S.; Lopez-Molina, L. Identification of Tannic Cell Walls at the Outer Surface of the Endosperm upon Arabidopsis Seed Coat Rupture. Plant J. 2020, 104, 567–580. [Google Scholar] [CrossRef]
- Pourcel, L.; Routaboul, J.M.; Kerhoas, L.; Caboche, M.; Lepiniec, L.; Debeaujon, I. TRANSPARENT TESTA10 Encodes a Laccase-Like Enzyme Involved in Oxidative Polymerization of Flavonoids in Arabidopsis Seed Coat. Plant Cell 2005, 17, 2966–2980. [Google Scholar] [CrossRef] [Green Version]
- Bissoli, G.; Bono, M.; Martínez-Almonacid, I.; Moreno-Peris, E.; Renard, J.; Espinosa, A.; Naranjo, M.Á.; Yenush, L.; Fita, A.; Serrano, R.; et al. Seed Coat Lignification Level Is Crucial in Capsicum spp. Seed Longevity. Physiol. Plant. 2021, 174, e13600. [Google Scholar] [CrossRef] [PubMed]
- Bessire, M.; Chassot, C.; Jacquat, A.-C.; Humphry, M.; Borel, S.; Petétot, J.M.-C.; Métraux, J.-P.; Nawrath, C. A Permeable Cuticle in Arabidopsis Leads to a Strong Resistance to Botrytis Cinerea. EMBO J. 2007, 26, 2158–2168. [Google Scholar] [CrossRef] [PubMed]
- Tsuwamoto, R.; Fukuoka, H.; Takahata, Y. GASSHO1 and GASSHO2 Encoding a Putative Leucine-Rich Repeat Transmembrane-Type Receptor Kinase Are Essential for the Normal Development of the Epidermal Surface in Arabidopsis Embryos. Plant J. 2008, 54, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Rains, M.K.; Gardiyehewa de Silva, N.D.; Molina, I. Reconstructing the Suberin Pathway in Poplar by Chemical and Transcriptomic Analysis of Bark Tissues. Tree Physiol. 2018, 38, 340–361. [Google Scholar] [CrossRef] [Green Version]
- Oñate-Sánchez, L.; Vicente-Carbajosa, J. DNA-Free RNA Isolation Protocols for Arabidopsis Thaliana, Including Seeds and Siliques. BMC Res. Notes 2008, 1, 93. [Google Scholar] [CrossRef] [Green Version]
- Czechowski, T.; Stitt, M.; Altmann, T.; Udvardi, M.K.; Scheible, W.-R. Genome-Wide Identification and Testing of Superior Reference Genes for Transcript Normalization in Arabidopsis. Plant Physiol. 2005, 139, 5–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Kubo, C.; Nomura, Y.; Matsuo, N.; Banno, H. Identification of Proteins That Interact with a Plant Nuclear Protein Using the Yeast Split-Trp Sensor. Plant Biotechnol. 2014, 31, 289–291. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Renard, J.; Bissoli, G.; Planes, M.D.; Gadea, J.; Naranjo, M.Á.; Serrano, R.; Ingram, G.; Bueso, E. Endosperm Persistence in Arabidopsis Results in Seed Coat Fractures and Loss of Seed Longevity. Plants 2023, 12, 2726. https://doi.org/10.3390/plants12142726
Renard J, Bissoli G, Planes MD, Gadea J, Naranjo MÁ, Serrano R, Ingram G, Bueso E. Endosperm Persistence in Arabidopsis Results in Seed Coat Fractures and Loss of Seed Longevity. Plants. 2023; 12(14):2726. https://doi.org/10.3390/plants12142726
Chicago/Turabian StyleRenard, Joan, Gaetano Bissoli, María Dolores Planes, José Gadea, Miguel Ángel Naranjo, Ramón Serrano, Gwyneth Ingram, and Eduardo Bueso. 2023. "Endosperm Persistence in Arabidopsis Results in Seed Coat Fractures and Loss of Seed Longevity" Plants 12, no. 14: 2726. https://doi.org/10.3390/plants12142726
APA StyleRenard, J., Bissoli, G., Planes, M. D., Gadea, J., Naranjo, M. Á., Serrano, R., Ingram, G., & Bueso, E. (2023). Endosperm Persistence in Arabidopsis Results in Seed Coat Fractures and Loss of Seed Longevity. Plants, 12(14), 2726. https://doi.org/10.3390/plants12142726