

Assessing the Effectiveness of Eco-Friendly Management Approaches for Controlling Wheat Yellow Rust and Their Impact on Antioxidant Enzymes

Abstract
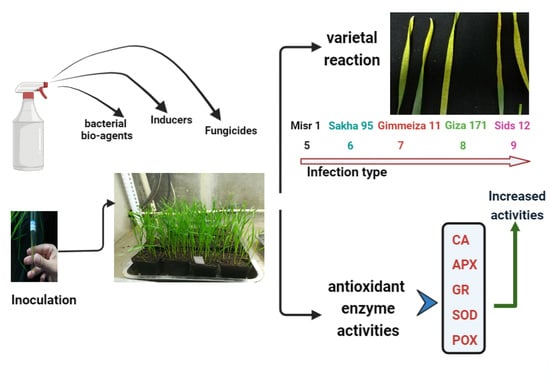
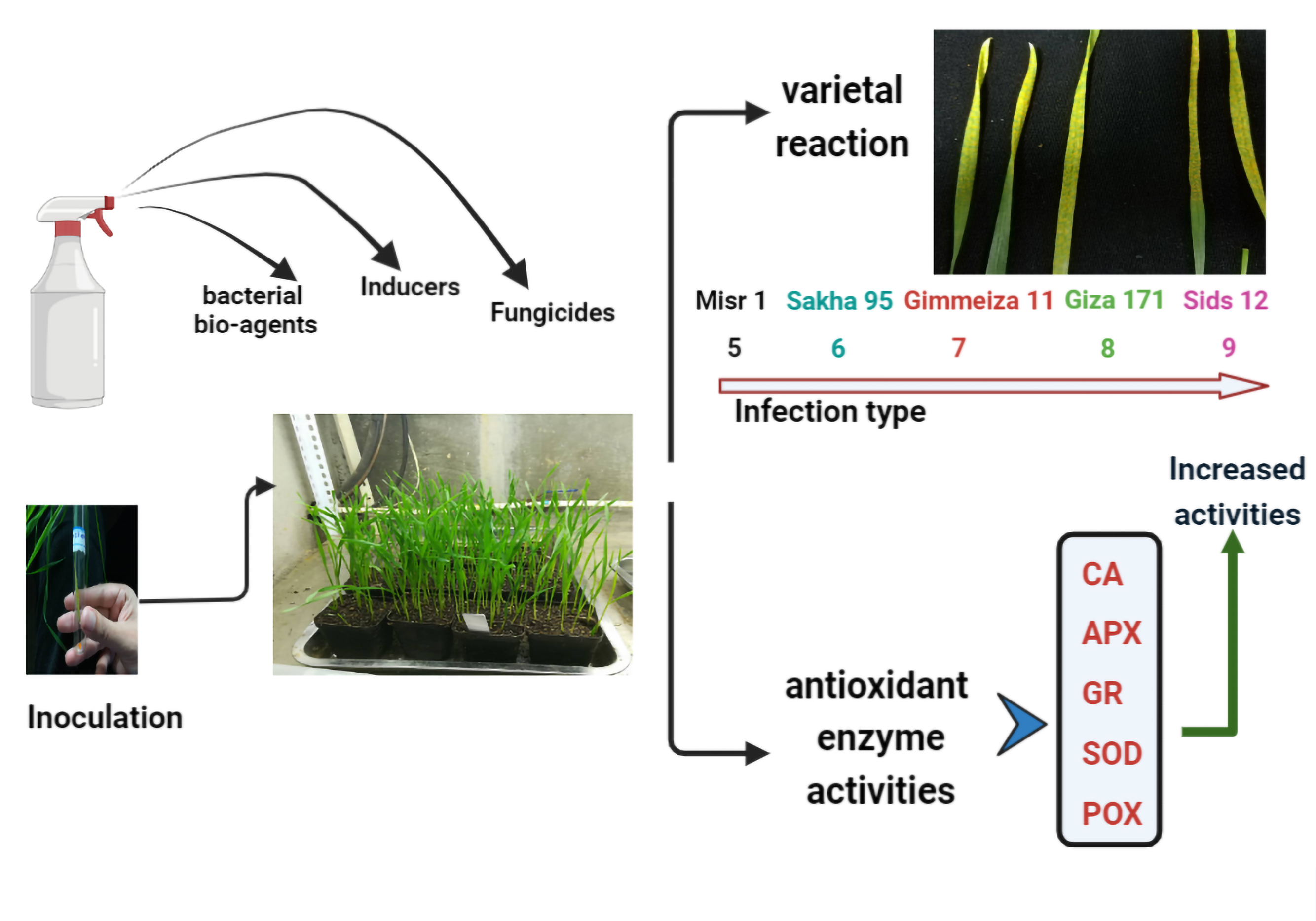
:
1. Introduction
2. Results
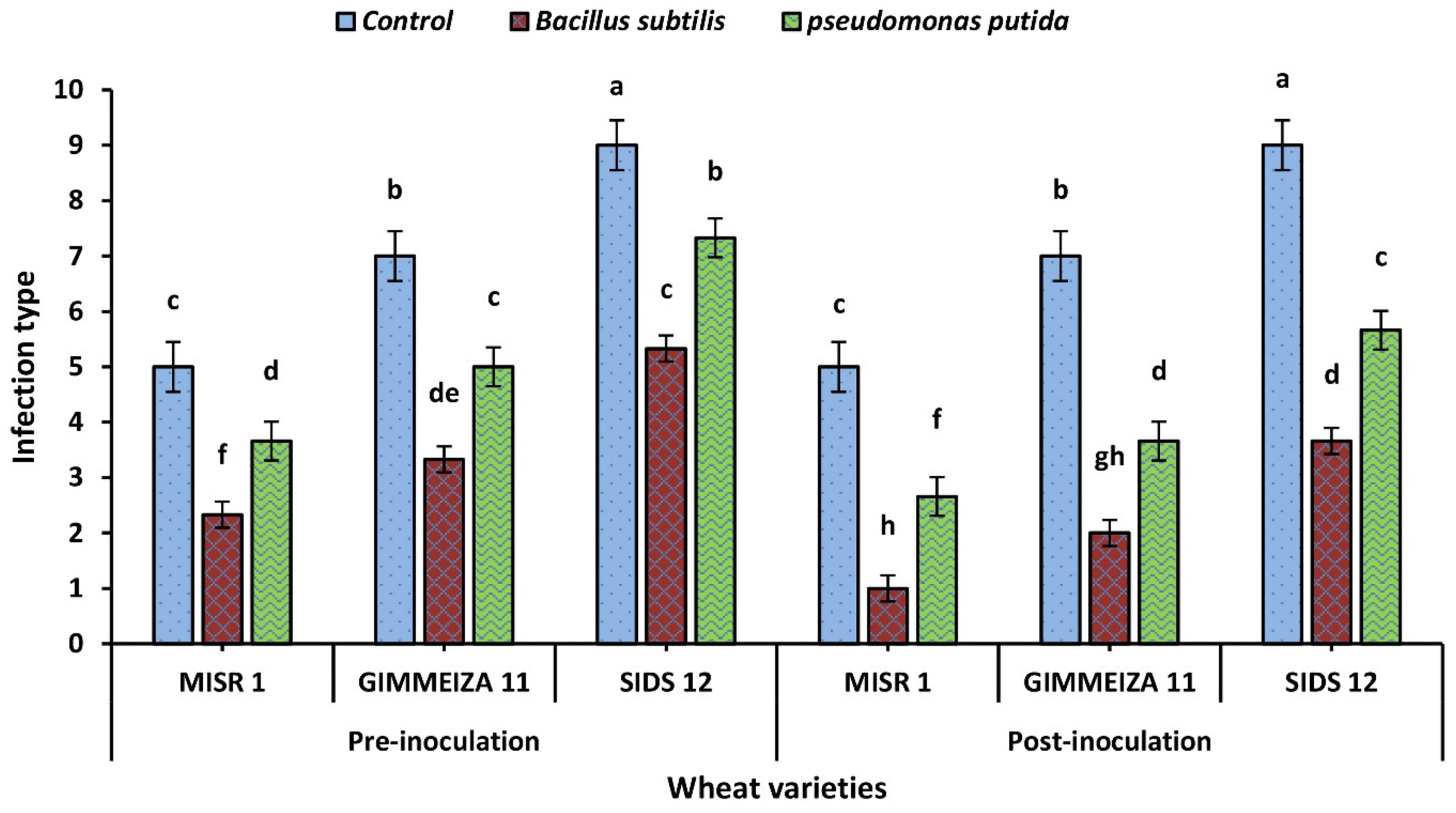
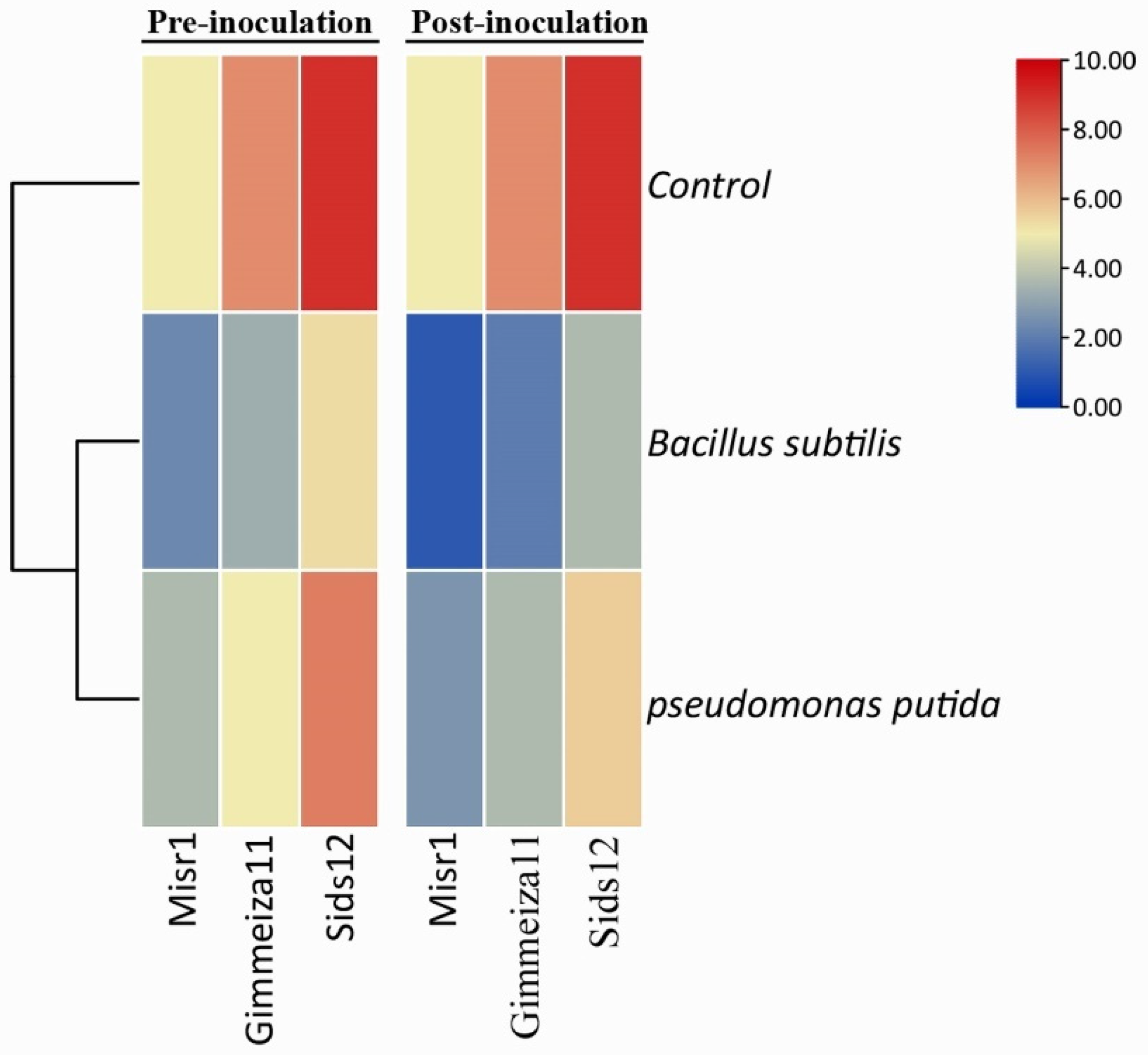
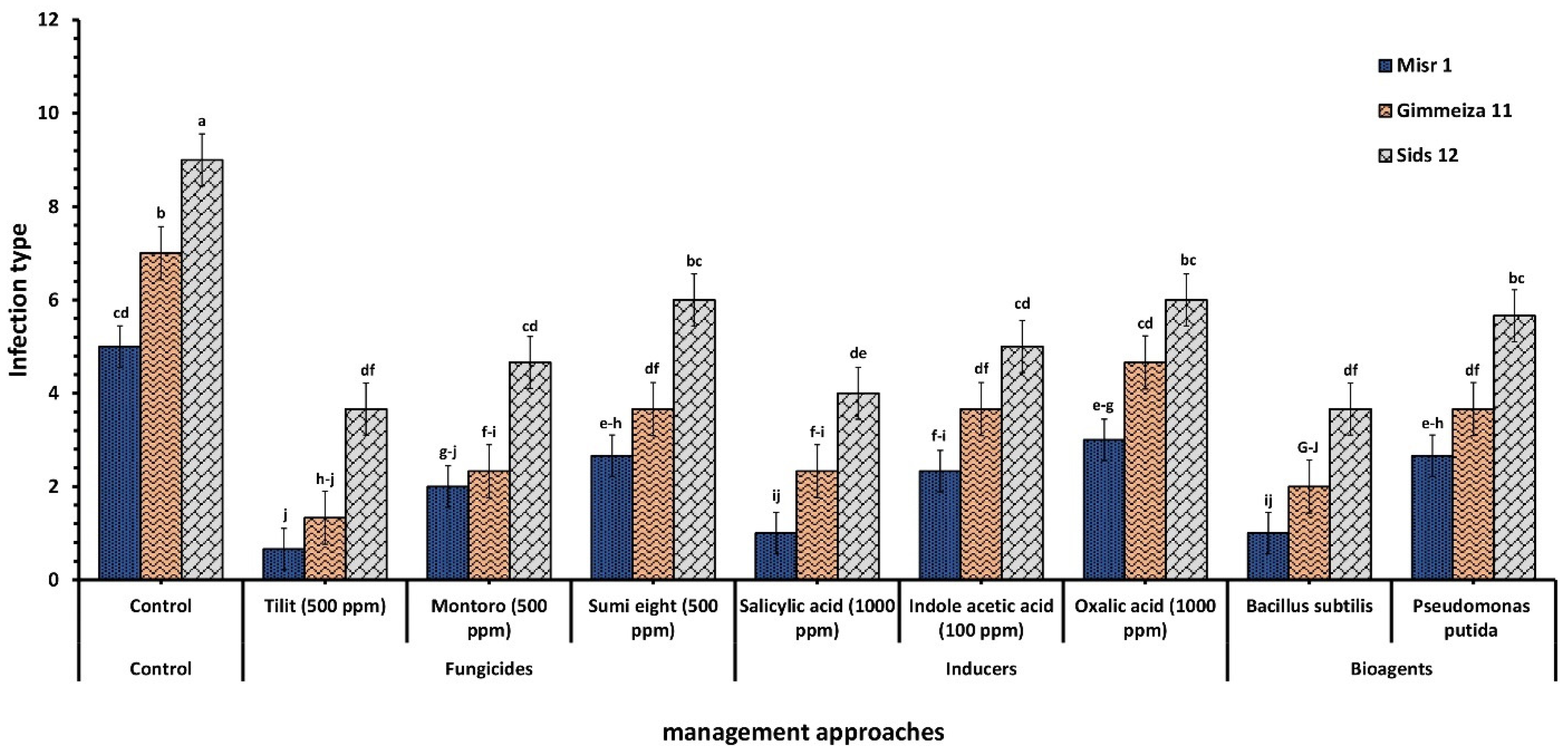
2.1. Wheat Stripe Rust Infection Type upon Application of Two Tested Bacterial Bioagents
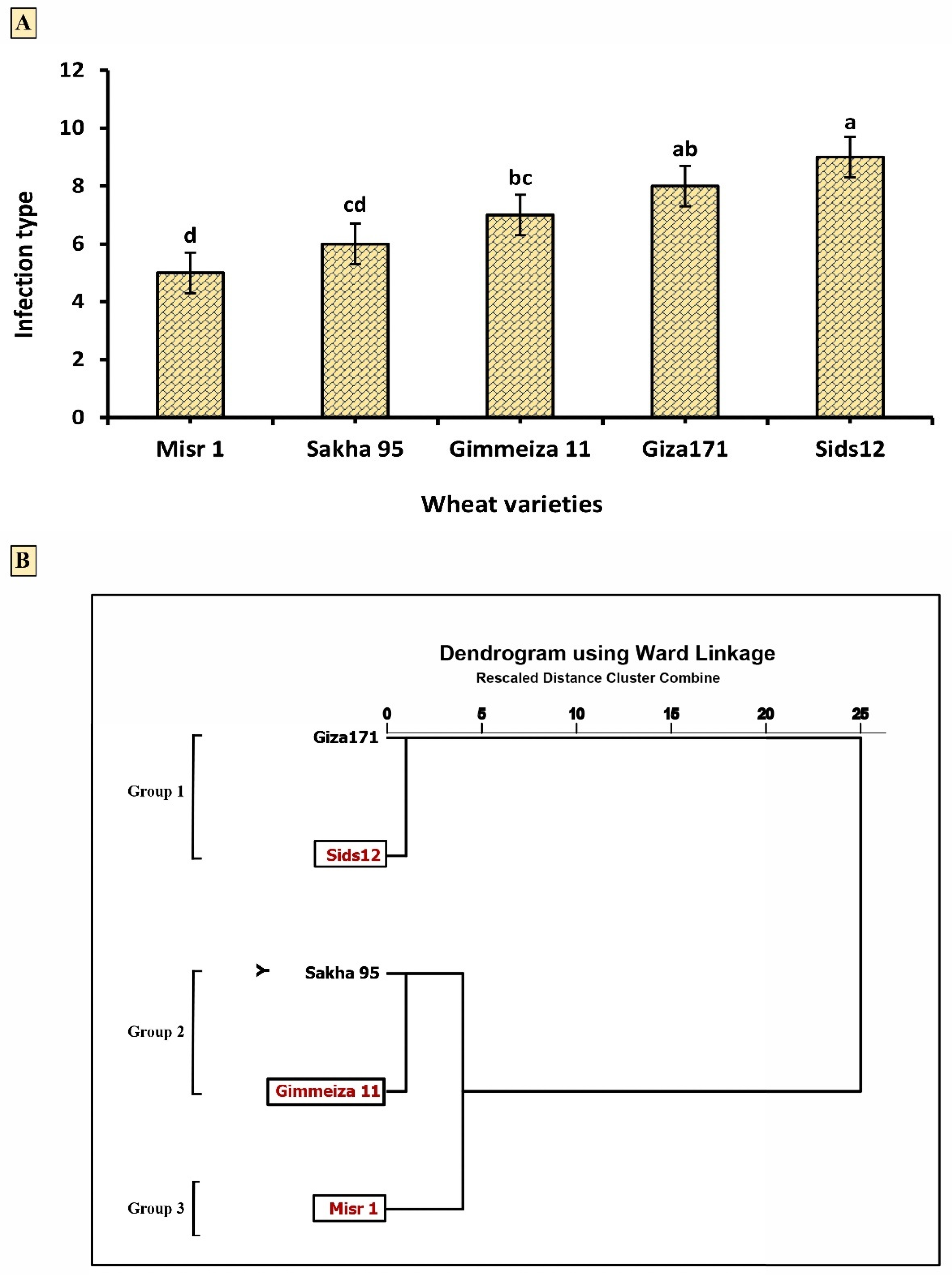
2.2. Wheat Varietal Reaction in Response to Stripe Rust Infection
2.3. Wheat Stripe Rust Infection Type upon Application of Three Tested Chemical Inducers
2.4. Wheat Stripe Rust Infection Type upon Application of Three Tested Fungicides
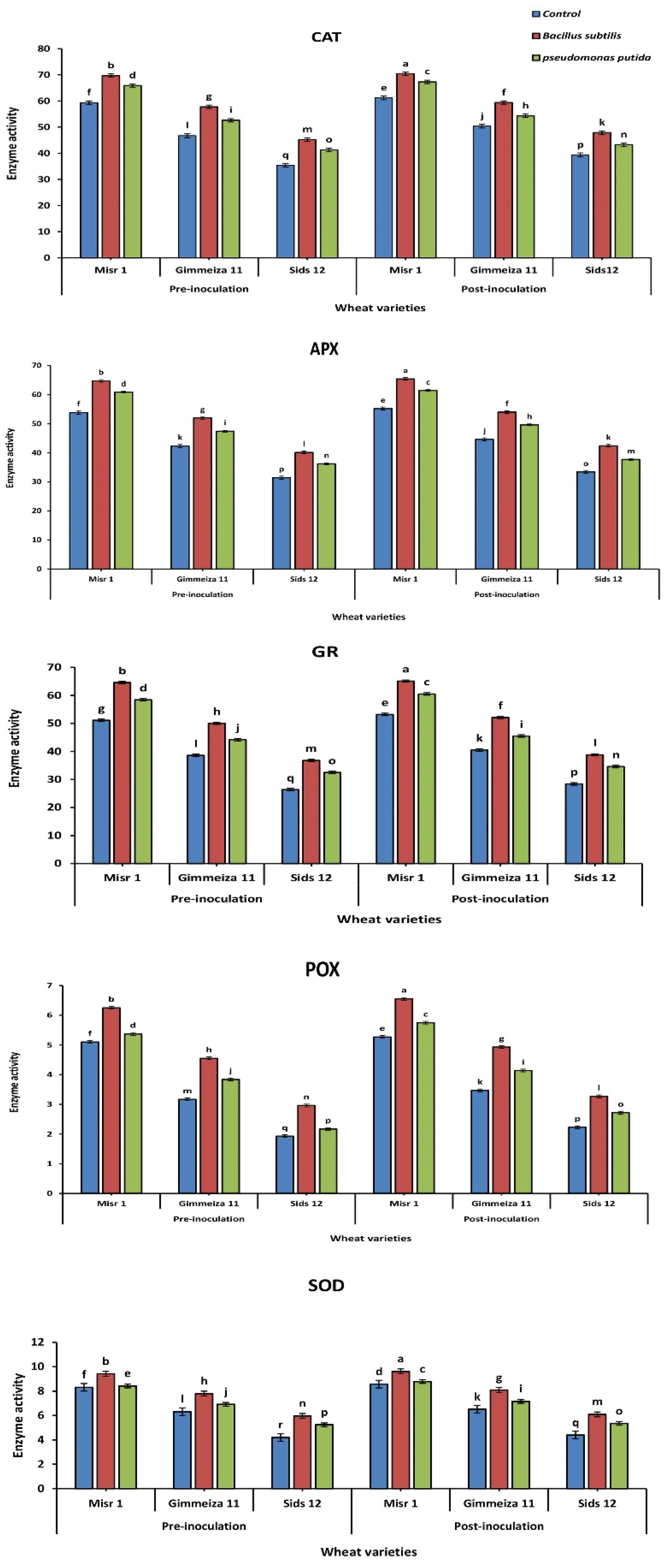
2.5. Antioxidant Enzyme Activity in Wheat-Stripe Rust Interaction
2.5.1. Catalase Activity
2.5.2. Ascorbate Peroxidase (APX) Activity
2.5.3. Glutathione Reductase (GR) Activity
2.5.4. Superoxide Dismutase (SOD) Activity
2.5.5. Peroxidase (POX) Activity
3. Discussion
3.1. Wheat Stripe Rust Infection Type upon Application of Two Tested Bacterial Bioagents
3.2. Wheat Varietal Reaction in Response to Stripe Rust Infection
3.3. Wheat Stripe Rust Infection Type upon Application of Three Tested Chemical Inducers
3.4. Wheat Stripe Rust Infection Type upon Application of Three Tested Fungicides
3.5. Antioxidant Enzyme Activity in Wheat-Stripe Rust Interaction
4. Materials and Methods
4.1. Seedling and Varietal Reaction
4.2. Inoculation of Pst
4.3. Disease Management
4.3.1. Biological Control
4.3.2. Chemical Inducers
4.3.3. Chemical Fungicides
4.4. Determination of Antioxidants Activities
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beddow, J.M.; Pardey, P.G.; Chai, Y.; Hurley, T.M.; Kriticos, D.J.; Braun, H.-J.; Park, R.F.; Cuddy, W.S.; Yonow, T. Research investment implications of shifts in the global geography of wheat stripe rust. Nat. Plants 2015, 1, 15132. [Google Scholar] [CrossRef]
- Figueroa, M.; Hammond-Kosack, K.E.; Solomon, P.S. A review of wheat diseases-a field perspective. Mol. Plant Pathol. 2018, 19, 1523–1536. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wellings, C.; Chen, X.; Kang, Z.; Liu, T. Wheat stripe (yellow) rust caused by Puccinia striiformis f. sp. tritici. Mol. Plant Pathol. 2014, 15, 433–446. [Google Scholar] [CrossRef]
- Thirugnana Sambandham, V.; Shankar, P.; Mukhopadhaya, S. Early Onset Yellow Rust Detection Guided by Remote Sensing Indices. Agriculture 2022, 12, 1206. [Google Scholar] [CrossRef]
- Omara, R.I.; Mazrou, Y.S.; Elsayed, A.; Moawad, N.; Nehela, Y.; Shahin, A.A. MISSR: A Mentoring Interactive System for Stripe Rust. Agronomy 2022, 12, 2416. [Google Scholar] [CrossRef]
- Shahin, A.; Ashmawy, M.; El-Orabey, W.; Esmail, S. Yield losses in wheat caused by stripe rust (Puccinia striiformis) in Egypt. Am. J. Life Sci. 2020, 8, 127–134. [Google Scholar] [CrossRef]
- Mishra, A.; Tiwari, K.; Prakasha, T.; Prasad, S.S. Use of Host Resistance for Management Wheat Rusts. In Innovative Approaches in Diagnosis and Management of Crop Diseases; Apple Academic Press: New York, NY, USA, 2021; pp. 269–301. [Google Scholar]
- Shahin, A.A. Occurrence of new races and virulence changes of the wheat stripe rust pathogen (Puccinia striiformis f. sp. tritici) in Egypt. Arch. Phytopathol. Plant Prot. 2020, 53, 552–569. [Google Scholar] [CrossRef]
- Jørgensen, L.N.; Hovmøller, M.S.; Hansen, J.G.; Lassen, P.; Clark, B.; Bayles, R.; Rodemann, B.; Flath, K.; Jahn, M.; Goral, T. IPM strategies and their dilemmas including an introduction to www.eurowheat.org. J. Integr. Agric. 2014, 13, 265–281. [Google Scholar] [CrossRef]
- Viljanen-Rollinson, S.; Marroni, M.; Butler, R. Wheat stripe rust control using fungicides in New Zealand. N. Z. Plant Prot. 2006, 59, 155–159. [Google Scholar] [CrossRef]
- Khan, M.A.; Raheel, M.; Khan, S.A.; Abid, A.D.; Shahzad, S.; Siddiqui, H.Z.; Atif, M.; Hanif, A. Eco-friendly management of wheat stripe rust through application of Bacillus subtilis in combination with plant defense activators. J. King Saud Univ.-Sci. 2023, 35, 102587. [Google Scholar] [CrossRef]
- Chen, X.; Kang, Z. Integrated Control of Stripe Rust; Springer: Dordrecht, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Zeng, Q.; Zhao, J.; Wu, J.; Zhan, G.; Han, D.; Kang, Z. Wheat stripe rust and integration of sustainable control strategies in China. Front. Agric. Sci. Eng. 2022, 9, 37–51. [Google Scholar] [CrossRef]
- Kiani, T.; Mehboob, F.; Hyder, M.Z.; Zainy, Z.; Xu, L.; Huang, L.; Farrakh, S. Control of stripe rust of wheat using indigenous endophytic bacteria at seedling and adult plant stage. Sci. Rep. 2021, 11, 14473. [Google Scholar] [CrossRef]
- Bale, J.; Van Lenteren, J.; Bigler, F. Biological control and sustainable food production. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 761–776. [Google Scholar] [CrossRef] [Green Version]
- Hermans, S.M.; Buckley, H.L.; Case, B.S.; Curran-Cournane, F.; Taylor, M.; Lear, G. Using soil bacterial communities to predict physico-chemical variables and soil quality. Microbiome 2020, 8, 79. [Google Scholar] [CrossRef]
- Archana, H.; Darshan, K.; Lakshmi, M.A.; Ghoshal, T.; Bashayal, B.M.; Aggarwal, R. Biopesticides: A key player in agro-environmental sustainability. In Trends of Applied Microbiology for Sustainable Economy; Elsevier: Amsterdam, The Netherlands, 2022; pp. 613–653. [Google Scholar]
- Aboulila, A.A. Efficiency of plant growth regulators as inducers for improve systemic acquired resistance against stripe rust disease caused by Puccinia striiformis f. sp. tritici in wheat through up-regulation of PR-1 and PR-4 genes expression. Physiol. Mol. Plant Pathol. 2022, 121, 101882. [Google Scholar] [CrossRef]
- Heil, M.; Bostock, R.M. Induced systemic resistance (ISR) against pathogens in the context of induced plant defences. Ann. Bot. 2002, 89, 503–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hembade, V.L.; Yashveer, S.; Taunk, J.; Sangwan, S.; Tokas, J.; Singh, V.; Redhu, N.S.; Grewal, S.; Malhotra, S.; Kumar, M. Chitosan-Salicylic acid and Zinc sulphate nano-formulations defend against yellow rust in wheat by activating pathogenesis-related genes and enzymes. Plant Physiol. Biochem. 2022, 192, 129–140. [Google Scholar] [CrossRef]
- Alexandersson, E.; Mulugeta, T.; Lankinen, Å.; Liljeroth, E.; Andreasson, E. Plant resistance inducers against pathogens in Solanaceae species-from molecular mechanisms to field application. Int. J. Mol. Sci. 2016, 17, 1673. [Google Scholar] [CrossRef] [Green Version]
- Chen, X. Pathogens which threaten food security: Puccinia striiformis, the wheat stripe rust pathogen. Food Secur. 2020, 12, 239–251. [Google Scholar] [CrossRef]
- Li, H.; Zhao, J.; Feng, H.; Huang, L.; Kang, Z. Biological control of wheat stripe rust by an endophytic Bacillus subtilis strain E1R-j in greenhouse and field trials. Crop Prot. 2013, 43, 201–206. [Google Scholar] [CrossRef]
- Pang, F.; Wang, T.; Zhao, C.; Tao, A.; Yu, Z.; Huang, S.; Yu, G. Novel bacterial endophytes isolated from winter wheat plants as biocontrol agent against stripe rust of wheat. BioControl 2016, 61, 207–219. [Google Scholar] [CrossRef]
- Reiss, A.; Jørgensen, L.N. Biological control of yellow rust of wheat (Puccinia striiformis) with Serenade® ASO (Bacillus subtilis strain QST713). Crop Prot. 2017, 93, 1–8. [Google Scholar] [CrossRef]
- Omara, R.I.; Essa, T.A.; Khalil, A.A.; Elsharkawy, M.M. A case study of non-traditional treatments for the control of wheat stem rust disease. Egypt. J. Biol. Pest Control 2020, 30, 83. [Google Scholar] [CrossRef]
- Dannies, M.; Tugizimana, F.; Steenkamp, P.A.; Piater, L.A.; Dubery, I.A.; Terefe, T.; Mhlongo, M.I. Metabolomic evaluation of PGPR defence priming in wheat (Triticum aestivum L.) cultivars infected with Puccinia striiformis f. sp. tritici (stripe rust). Front. Plant Sci. 2023, 14, 1261. [Google Scholar]
- Loeffler, W.; Tschen, J.S.M.; Vanittanakom, N.; Kugler, M.; Knorpp, E.; Hsieh, T.F.; Wu, T.G. Antifungal effects of bacilysin and fengymycin from Bacillus subtilis F-29-3 a comparison with activities of other Bacillus antibiotics. J. Phytopathol. 1986, 115, 204–213. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, J.; Chen, H.; Fan, Y.; Shi, Z. Antifungal activity of eugenol against Botrytis cinerea. Trop. Plant Pathol. 2010, 35, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Abo-Elyousr, K.A.; Abdel-Rahim, I.R.; Almasoudi, N.M.; Alghamdi, S.A. Native endophytic Pseudomonas putida as a biocontrol agent against common bean rust caused by Uromyces appendiculatus. J. Fungi 2021, 7, 745. [Google Scholar] [CrossRef]
- Abebe, W. Wheat leaf rust disease management: A Review. Int. J. Novel Res. Interdiscip. Stud. 2021, 8, 1–14. [Google Scholar]
- Tjamos, S.E.; Flemetakis, E.; Paplomatas, E.J.; Katinakis, P. Induction of resistance to Verticillium dahliae in Arabidopsis thaliana by the biocontrol agent K-165 and pathogenesis-related proteins gene expression. Mol. Plant-Microbe Interact. 2005, 18, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Shatrupa Ray, S.R.; Vivek Singh, V.S.; Kartikay Bisen, K.B.; Chetan Keswani, C.K.; Surendra Singh, S.S.; Singh, H. Endophytomicrobiont: A multifaceted beneficial interaction. In Advances in PGPR Research; CABI: Wallingford, UK, 2017; pp. 218–233. [Google Scholar]
- Sivasakthi, S.; Usharani, G.; Saranraj, P. Biocontrol potentiality of plant growth promoting bacteria (PGPR)-Pseudomonas fluorescens and Bacillus subtilis: A review. Afr. J. Agric. Res. 2014, 9, 1265–1277. [Google Scholar]
- Zehra, A.; Raytekar, N.A.; Meena, M.; Swapnil, P. Efficiency of microbial bio-agents as elicitors in plant defense mechanism under biotic stress: A review. Curr. Res. Microb. Sci. 2021, 2, 100054. [Google Scholar] [CrossRef] [PubMed]
- Urszula, W.; Barbara, M.; Monika, B.; Zofia, K. Biological control of winter wheat pathogens by bacteria. Acta Fytotech. Zootech. 2004, 7, 349–350. [Google Scholar]
- El-Kazzaz, M.K.; Ghoniem, K.E.; Ashmawy, M.A.; Omar, G.E.; Hafez, Y.M. Suppression of wheat strip rust disease caused by Puccinia striiformis f. sp. tritici by eco-friendly bio-control agents correlated with yield improvement. Fresenius Environ. Bull. 2020, 29, 8385–8393. [Google Scholar]
- Hafez, Y.; Emeran, A.; Esmail, S.; Mazrou, Y.; Abdrabbo, D.; Abdelaal, K. Alternative treatments improve physiological characters, yield and tolerance of wheat plants under leaf rust disease stress. Fresenius Environ. Bull. 2020, 29, 4738–4748. [Google Scholar]
- Esmail, S.M.; Draz, I.S.; Saleem, M.H.; Mumtaz, S.; Elsharkawy, M.M. Penicillium simplicissimum and Trichoderma asperellum counteract the challenge of Puccinia striiformis f. sp. tritici in wheat plants. Egypt. J. Biol. Pest Control 2022, 32, 116. [Google Scholar] [CrossRef]
- Singh, V.K.; Mathuria, R.; Gogoi, R.; Aggarwal, R. Impact of different fungicides and bioagents, and fungicidal spray timing on wheat stripe rust development and grain yield. Indian Phytopathol. 2016, 69, 357–362. [Google Scholar]
- Abu El-Naga, S.; Khalifa, M.; Sherif, S.; Youssef, W.; El-Daoudi, Y.; Shafik, I. Virulence of wheat stripe rust pathotypes identified in Egypt during 1999/2000 and sources of resistance. In Proceedings of the Meeting the Challenge of Yellow Rust in Cereal Crops, Proceedings of the First Regional Conference on Yellow Rust in the Central and West Asia and North Africa Region, Karaj, Iran, 8–14 May 2001; pp. 62–68, International Center for Agricultural Research in the Dry Areas (ICARDA), 2002. [Google Scholar]
- Omara, R.; El-Naggar, D.; El-Malik, A.; Ketta, H. Losses assessment in some Egyptian wheat cultivars caused by stripe rust pathogen (Puccinia striiformis). Egypt. J. Phytopathol. 2016, 44, 191–203. [Google Scholar] [CrossRef]
- Esmail, S.M.; Omara, R.I.; Abdelaal, K.A.; Hafez, Y.M. Histological and biochemical aspects of compatible and incompatible wheat-Puccinia striiformis interactions. Physiol. Mol. Plant Pathol. 2019, 106, 120–128. [Google Scholar] [CrossRef]
- Abdelaal, K.A.; Hafez, Y.; Badr, M.; Youseef, W.; Esmail, S.M. Biochemical, Histological and Molecular Changes in Susceptible and Resistant Wheat Cultivars Inoculated with Stripe Rust Fungus Puccinia striiformis f. sp. tritici. Egypt. J. Biol. Pest Control 2014, 24, 421–429. [Google Scholar]
- Singh, R.; Huerta-Espino, J.; Rajaram, S. Achieving near-immunity to leaf and stripe rusts in wheat by combining slow rusting resistance genes. Acta Phytopathol. Entomol. Hung. 2000, 35, 133–139. [Google Scholar]
- Zheng, S.; Li, Y.; Lu, L.; Liu, Z.; Zhang, C.; Ao, D.; Li, L.; Zhang, C.; Liu, R.; Luo, C. Evaluating the contribution of Yr genes to stripe rust resistance breeding through marker-assisted detection in wheat. Euphytica 2017, 213, 50. [Google Scholar] [CrossRef]
- Abu Aly, A.; Omara, R.; El-Malik, A.; Nagwa, I. Evaluation of new sources of resistance to wheat stripe rust (Puccinia striiformis f. sp. tritici), under Egyptian field conditions. J. Plant Prot. Pathol. 2017, 8, 181–188. [Google Scholar] [CrossRef]
- Coram, T.E.; Wang, M.; Chen, X. Transcriptome analysis of the wheat-Puccinia striiformis f. sp. tritici interaction. Mol. Plant Pathol. 2008, 9, 157–169. [Google Scholar] [CrossRef]
- Liu, R.; Lu, J.; Zhou, M.; Zheng, S.; Liu, Z.; Zhang, C.; Du, M.; Wang, M.; Li, Y.; Wu, Y. Developing stripe rust resistant wheat (Triticum aestivum L.) lines with gene pyramiding strategy and marker-assisted selection. Genet. Resour. Crop Evol. 2020, 67, 381–391. [Google Scholar] [CrossRef]
- Khan, N.; Ali, S.; Shahid, M.A.; Kharabian-Masouleh, A. Advances in detection of stress tolerance in plants through metabolomics approaches. Plant Omics 2017, 10, 153–163. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhang, H.; Yao, J.; Han, Q.; Wang, X.; Huang, L.; Kang, Z. Cytological and molecular characterization of non-host resistance in Arabidopsis thaliana against wheat stripe rust. Plant Physiol. Biochem. 2013, 62, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Bano, A. Interactive effects of Ag-nanoparticles, salicylic acid, and plant growth promoting rhizobacteria on the physiology of wheat infected with yellow rust. J. Plant Pathol. 2020, 102, 1215–1225. [Google Scholar] [CrossRef]
- Elsharkawy, M.M.; Omara, R.I.; Mostafa, Y.S.; Alamri, S.A.; Hashem, M.; Alrumman, S.A.; Ahmad, A.A. Mechanism of wheat leaf rust control using chitosan nanoparticles and salicylic acid. J. Fungi 2022, 8, 304. [Google Scholar] [CrossRef] [PubMed]
- Fayadh, A.H.; Al-Maaroof, E.M.; Fattah, F.A. Induced resistance to wheat yellow rust by chemical inducers. J. Biol. Agric. Healthc. 2013, 3, 56–64. [Google Scholar]
- Corredor-Moreno, P.; Minter, F.; Davey, P.E.; Wegel, E.; Kular, B.; Brett, P.; Lewis, C.M.; Morgan, Y.M.; Macías Pérez, L.A.; Korolev, A.V. The branched-chain amino acid aminotransferase TaBCAT1 modulates amino acid metabolism and positively regulates wheat rust susceptibility. Plant Cell 2021, 33, 1728–1747. [Google Scholar] [CrossRef]
- Khalil, M.S.A.; Abdel-Kader, M.M.; El-Mougy, N.S.; El-Gamal, N.G. Foliar application with organic acids for suppressing the severity of wheat powdery mildew disease caused by Blumeria graminis f. sp. tritici under field conditions. Res. Crops 2021, 22, 319–326. [Google Scholar]
- Devi, B.; Singh, G.; Dash, A.K.; Gupta, S. Chemically induced systemic acquired resistance in the inhibition of French bean rust. Curr. Plant Biol. 2020, 23, 100151. [Google Scholar] [CrossRef]
- Han, Q.-M.; Kang, Z.-S.; Wei, G.-R. Studies on Fulicur and Caramba for controlling wheat stripe rust disease. Plant Prot.-Beijing 2003, 29, 61–63. [Google Scholar]
- Chen, X. Epidemiology and control of stripe rust [Puccinia striiformis f. sp. tritici] on wheat. Can. J. Plant Pathol. 2005, 27, 314–337. [Google Scholar] [CrossRef]
- Wan, A.M.; Chen, X.M.; He, Z. Wheat stripe rust in China. Aust. J. Agric. Res. 2007, 58, 605–619. [Google Scholar] [CrossRef]
- Han, Q.; Kang, Z.; Buchenauer, H.; Huang, L.; Zhao, J. Cytological and immunocytochemical studies on the effects of the fungicide tebuconazole on the interaction of wheat with stripe rust. J. Plant Pathol. 2006, 88, 263–271. [Google Scholar]
- Hamada, M.S.; Yin, Y.; Ma, Z. Sensitivity to iprodione, difenoconazole and fludioxonil of Rhizoctonia cerealis isolates collected from wheat in China. Crop Prot. 2011, 30, 1028–1033. [Google Scholar] [CrossRef]
- Wang, F.; Cao, D.; Shi, L.; He, S.; Li, X.; Fang, H.; Yu, Y. Competitive adsorption and mobility of propiconazole and difenoconazole on five different soils. Bull. Environ. Contam. Toxicol. 2020, 105, 927–933. [Google Scholar] [CrossRef]
- Carmona, M.; Sautua, F.; Pérez-Hérnandez, O.; Reis, E.M. Role of fungicide applications on the integrated management of wheat stripe rust. Front. Plant Sci. 2020, 11, 733. [Google Scholar] [CrossRef]
- Cook, N.M.; Chng, S.; Woodman, T.L.; Warren, R.; Oliver, R.P.; Saunders, D.G. High frequency of fungicide resistance-associated mutations in the wheat yellow rust pathogen Puccinia striiformis f. sp. tritici. Pest Manag. Sci. 2021, 77, 3358–3371. [Google Scholar] [CrossRef] [PubMed]
- Steffens, J.J.; Pell, E.J.; Tien, M. Mechanisms of fungicide resistance in phytopathogenic fungi. Curr. Opin. Biotechnol. 1996, 7, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, L.N.; Matzen, N.; Hansen, J.G.; Semaskiene, R.; Korbas, M.; Danielewicz, J.; Glazek, M.; Maumene, C.; Rodemann, B.; Weigand, S. Four azoles’ profile in the control of Septoria, yellow rust and brown rust in wheat across Europe. Crop Prot. 2018, 105, 16–27. [Google Scholar] [CrossRef]
- Godeau, C.; Morin-Crini, N.; Staelens, J.-N.; Martel, B.; Rocchi, S.; Chanet, G.; Fourmentin, M.; Crini, G. Adsorption of a triazole antifungal agent, difenoconazole, on soils from a cereal farm: Protective effect of hemp felt. Environ. Technol. Innov. 2021, 22, 101394. [Google Scholar] [CrossRef]
- Zhang, Z.; Jiang, W.; Jian, Q.; Song, W.; Zheng, Z.; Wang, D.; Liu, X. Residues and dissipation kinetics of triazole fungicides difenoconazole and propiconazole in wheat and soil in Chinese fields. Food Chem. 2015, 168, 396–403. [Google Scholar] [CrossRef]
- Chen, X. Integration of cultivar resistance and fungicide application for control of wheat stripe rust. Can. J. Plant Pathol. 2014, 36, 311–326. [Google Scholar] [CrossRef]
- Chen, X.; Evans, C.; Garner, J.; Liu, Y. Control of stripe rust of spring wheat with foliar fungicide. In Plant Disease; USDA: Washington, DC, USA, 2013; p. 7. [Google Scholar]
- Ali, Y.; Abbas, T.; Aatif, H.M.; Ahmad, S.; Khan, A.A.; Hanif, C.M. Impact of foliar applications of different fungicides on wheat stripe rust epidemics and grain yield. Pak. J. Phytopathol. 2022, 34, 135–141. [Google Scholar] [CrossRef]
- Bekana, N.B. Efficacy evaluation of different foliar fungicides for the management of wheat strip rust (Puccinia striiformis) in West Shoa Zone, Oromia, Ethiopia. J. Appl. Sci. Environ. Manag. 2019, 23, 1977–1983. [Google Scholar] [CrossRef] [Green Version]
- Jindal, M.M.; Sharma, I.; Bains, N.S. Losses due to stripe rust caused by Puccinia striiformis in different varieties of wheat. J. Cereal Res. 2012, 4, 33–36. [Google Scholar]
- Basandrai, A.K.; Sharma, B.; Basandrai, D. Efficacy of triazole fungicides for the integrated management of yellow rust, leaf rust and powdery mildew of wheat. Plant Dis. Res. 2013, 28, 135–139. [Google Scholar]
- Kang, Z.; Li, X.; Wan, A.; Wang, M.; Chen, X. Differential sensitivity among Puccinia striiformis f. sp. tritici isolates to propiconazole and pyraclostrobin fungicides. Can. J. Plant Pathol. 2019, 41, 415–434. [Google Scholar] [CrossRef]
- Basandrai, A.K.; Mehta, A.; Rathee, V.; Basandrai, D.; Sharma, B. Efficacy of fungicides in managing yellow rust of wheat. J. Cereal Res. 2020, 12, 103–108. [Google Scholar] [CrossRef]
- Covarelli, L.; Orfei, M. Chemical control of foliar disease of winter bread wheat. Inf. Fitopatlolgico 2005, 55, 27–32. [Google Scholar]
- Walters, D.R.; Avrova, A.; Bingham, I.J.; Burnett, F.J.; Fountaine, J.; Havis, N.D.; Hoad, S.P.; Hughes, G.; Looseley, M.; Oxley, S.J. Control of foliar diseases in barley: Towards an integrated approach. Eur. J. Plant Pathol. 2012, 133, 33–73. [Google Scholar] [CrossRef]
- Rodríguez-García, M.F.; González-González, M.; Huerta-Espino, J.; Solano-Hernández, S. Fungicides evaluation against yellow rust (Puccinia striiformis f. sp. hordei) in six barley cultivars. Rev. Mex. Fitopatol. 2021, 39, 414–434. [Google Scholar]
- Boualem, B.; Mohamed, B.; Moulay, B. Effect of application timing of artea and amistar xtra on the yield of wheat (Triticum aestivum L.) under foliar disease in the East-Algerian. Int. J. Agric. Res. 2017, 12, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Omara, R.; Kamel, S.; Hafez, Y.; Morsy, S. Role of Non-traditional Control Treatments in Inducing Resistance Against Wheat Leaf Rust Caused by Puccinia triticina. Egypt. J. Biol. Pest Control 2015, 25, 609–618. [Google Scholar]
- Hanifei, M.; Dehghani, H.; Choukan, R. The role of antioxidant enzymes and phenolic compounds in disease resistance to Fusarium oxysporum f. sp. Melonis race 1.2. Int. J. Agron. Plant Prod. 2013, 4, 1985–1996. [Google Scholar]
- Rizhsky, L.; Hallak-Herr, E.; Van Breusegem, F.; Rachmilevitch, S.; Barr, J.E.; Rodermel, S.; Inzé, D.; Mittler, R. Double antisense plants lacking ascorbate peroxidase and catalase are less sensitive to oxidative stress than single antisense plants lacking ascorbate peroxidase or catalase. Plant J. 2002, 32, 329–342. [Google Scholar] [CrossRef] [Green Version]
- Liau, C.Y.; Lin, C.S. Detection of chitinolytic enzymes in Ipomoea batatas leaf extract by activity staining after gel electrophoresis. J. Chin. Chem. Soc. 2008, 55, 678–681. [Google Scholar] [CrossRef]
- Khalifa, N.A.; Abou-Zeid, N.; Noher, A.M.; Abbas, M.; Sobhy, H. Enzyme activity and biochemical changes associated with induction of systemic resistance of faba bean against damping off disease. Egypt. J. Biol. Pest Control 2016, 26, 395–404. [Google Scholar]
- Trchounian, A.; Petrosyan, M.; Sahakyan, N. Plant cell redox homeostasis and reactive oxygen species. In Redox State as a Central Regulator of Plant-Cell Stress Responses; Springer: Cham, Switzerland, 2016; pp. 25–50. [Google Scholar]
- Milavec, M.; Ravnikar, M.; Kovač, M. Peroxidases and photosynthetic pigments in susceptible potato infected with potato virus YNTN. Plant Physiol. Biochem. 2001, 39, 891–898. [Google Scholar] [CrossRef]
- Hameed, F.E.; Abood, J.K.; Temur, H.A. The change of peroxidase activity in three cucumber cultivars during development of powdery mildew infection. J. Babylon. Univ. Pure Appl. Sci. 2011, 3, 877–883. [Google Scholar]
- El-Komy, M.H. Comparative analysis of defense responses in chocolate spot-resistant and-susceptible faba bean (Vicia faba) cultivars following infection by the necrotrophic fungus Botrytis fabae. Plant Pathol. J. 2014, 30, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Yang, D.; Yang, B.; Gao, Z.; Li, M.; Jiang, Y.; Hu, M. β-Aminobutyric acid induces resistance of mango fruit to postharvest anthracnose caused by Colletotrichum gloeosporioides and enhances activity of fruit defense mechanisms. Sci. Hortic. 2013, 160, 78–84. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox Signaling in Plants. Antioxid Redox Signal. 2013, 18, 2087–2090. [Google Scholar] [CrossRef]
- Martinez, V.; Mestre, T.C.; Rubio, F.; Girones-Vilaplana, A.; Moreno, D.A.; Mittler, R.; Rivero, R.M. Accumulation of flavonols over hydroxycinnamic acids favors oxidative damage protection under abiotic stress. Front. Plant Sci. 2016, 7, 838. [Google Scholar] [CrossRef] [Green Version]
- Abdelaal, K.A. Effect of salicylic acid and abscisic acid on morpho-physiological and anatomical characters of faba bean plants (Vicia faba L.) under drought stress. J. Plant Prod. 2015, 6, 1771–1788. [Google Scholar] [CrossRef] [Green Version]
- Gupta, G.; Parihar, S.S.; Ahirwar, N.K.; Snehi, S.K.; Singh, V. Plant growth promoting rhizobacteria (PGPR): Current and future prospects for development of sustainable agriculture. J. Microb. Biochem. Technol. 2015, 7, 96–102. [Google Scholar]
- Liu, R.; Li, J.; Zhang, L.; Feng, T.; Zhang, Z.; Zhang, B. Fungicide difenoconazole induced biochemical and developmental toxicity in wheat (Triticum aestivum L.). Plants 2021, 10, 2304. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Khaledi, N.; Taheri, P.; Falahati-Rastegar, M. Evaluation of resistance and the role of some defense responses in wheat cultivars to Fusarium head blight. J. Plant Prot. Res. 2017, 57, 398–408. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Wang, X.; Zhang, Q.; Fu, Y.; Feng, C.; Wang, B.; Huang, L.; Kang, Z. Monodehydroascorbate reductase gene, regulated by the wheat PN-2013 miRNA, contributes to adult wheat plant resistance to stripe rust through ROS metabolism. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2014, 1839, 1–12. [Google Scholar] [CrossRef]
- Lagudah, E.S.; Krattinger, S.G.; Herrera-Foessel, S.; Singh, R.P.; Huerta-Espino, J.; Spielmeyer, W.; Brown-Guedira, G.; Selter, L.L.; Keller, B. Gene-specific markers for the wheat gene Lr34/Yr18/Pm38 which confers resistance to multiple fungal pathogens. Theor. Appl. Genet. 2009, 119, 889–898. [Google Scholar] [CrossRef] [Green Version]
- Elsharkawy, M.M.; Shimizu, M.; Takahashi, H.; Ozaki, K.; Hyakumachi, M. Induction of systemic resistance against Cucumber mosaic virus in Arabidopsis thaliana by Trichoderma asperellum SKT-1. Plant Pathol. J. 2013, 29, 193. [Google Scholar] [CrossRef] [Green Version]
- Levy, A.; Guenoune-Gelbart, D.; Epel, B.L. β-1,3-Glucanases: Plasmodesmal gate keepers for intercellular communication. Plant Signal. Behav. 2007, 2, 404–407. [Google Scholar] [CrossRef] [Green Version]
- Wan, A.; Zhao, Z.; Chen, X.; He, Z.; Jin, S.; Jia, Q.; Yao, G.; Yang, J.; Wang, B.; Li, G. Wheat stripe rust epidemic and virulence of Puccinia striiformis f. sp. tritici in China in 2002. Plant Dis. 2004, 88, 896–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeal, F.; Konzak, C.; Smith, E.; Tate, W.; Russell, T. A Uniform System for Recording and Processing Cereal Research Data; Agricultural Research Service Bulletin, United States Department of Agriculture: Washington, DC, USA, 1971; pp. 34–121.
- Vitória, A.P.; Lea, P.J.; Azevedo, R.A. Antioxidant enzymes responses to cadmium in radish tissues. Phytochemistry 2001, 57, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Chance, B.; Maehly, A. Methods in Enzymol; Colowick, S.P., Kaplan, N.O., Eds.; Academic Press, Inc.: New York, NY, USA, 1957; Volume 4, p. 273. [Google Scholar]
- Thomas, R.L.; Jen, J.J.; Morr, C.V. Changes in soluble and bound peroxidase-IAA oxidase during tomato fruit development. J. Food Sci. 1982, 47, 158–161. [Google Scholar] [CrossRef]
- Fielding, J.; Hall, J. A biolchemical and cytochemical study of peroxidase activity in roots of Pisum sativum: I. a comparison of DAB-peroxidase and guaiacol-peroxidase with particular emphasis on the properties of cell wall activity. J. Exp. Bot. 1978, 29, 969–981. [Google Scholar] [CrossRef]
- Sairam, R.K.; Rao, K.V.; Srivastava, G. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P. Ultraviolet-B-and ozone-induced biochemical changes in antioxidant enzymes of Arabidopsis thaliana. Plant Physiol. 1996, 110, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Inducers | Con. ppm | Pre-Inoculation | Post-Inoculation | ||||
|---|---|---|---|---|---|---|---|
| Misr1 | Gimmeiza11 | Sids12 | Misr1 | Gimmeiza11 | Sids12 | ||
| Control | 0 | 5.00 j–l | 7.00 d–f | 9.00 a | 5.00 e–h | 7.00 b,c | 9.00 a |
| Salicylic acid | 250 | 3.66 n–p | 4.66 k–m | 6.66 e–g | 2.33 m,n | 3.33 j–m | 5.33 d–g |
| 500 | 3.00 p,q | 4.33 l–n | 6.33 f–h | 1.66 n,o | 3.00 k–m | 5.00 e–h | |
| 1000 | 2.33 q | 3.66 n–p | 5.66 h–j | 1.00 o,p | 2.33 m,n | 4.00 h–k | |
| Indole acetic acid | 25 | 4.00 m–o | 5.33 i–k | 7.66 c,d | 3.00 k–m | 4.33 g–j | 6.00 c–e |
| 50 | 3.66 n–p | 5.00 j–l | 7.33 c–e | 2.66 l–n | 4.00 h–k | 5.66 d–f | |
| 100 | 3.33 o,p | 4.66 k–m | 6.66 e–g | 2.33 m,n | 3.66 i–l | 5.00 e–h | |
| oxalic acid | 250 | 4.66 k–m | 6.33 f–h | 8.66 a,b | 4.00 h–k | 5.33 d–g | 7.33 b |
| 500 | 4.33 l–n | 6.00 g–i | 8.00 b,c | 3.66 i–l | 5.00 e–h | 6.33 b–d | |
| 1000 | 3.66 n–p | 5.33 i–k | 7.66 c,d | 3.00 k–m | 4.66 f–i | 6.00 c–e | |
| L.S.D. | |||||||
| Varieties | 0.273 | 0.326 | |||||
| Concentration | 0.316 | 0.377 | |||||
| Inducers | 0.273 | 0.326 | |||||
| Varieties × Concentration | 0.547 | 0.653 | |||||
| Varieties × Inducers | 0.474 | 0.566 | |||||
| Concentration × Inducers | 0.547 | 0.653 | |||||
| Varieties × Concentration × inducers | 0.948 | 1.132 | |||||
| Tested Fungicides | Con. ppm | Pre-Inoculation | Post-Inoculation | ||||
|---|---|---|---|---|---|---|---|
| Misr1 | Gimmeiza11 | Sids12 | Misr1 | Gimmeiza11 | Sids12 | ||
| Control | 0 | 5.00 g–i | 7.00 c–e | 9.00 a | 5.00 d–f | 7.00 b | 9.00 a |
| Tilit | 125 | 3.00 l–n | 4.00 i–l | 6.33 d–f | 1.66 m,o | 2.66 j–m | 4.33 e–h |
| 250 | 2.66 m,n | 3.33 k–n | 6.00 e–g | 1.33 n,o | 2.00 l–n | 4.00 f–i | |
| 500 | 2.33 n | 3.00 l–n | 5.33 f–h | 0.66 o | 1.33 n,o | 3.66 g–j | |
| Montoro | 125 | 3.66 j–l | 5.00 g–i | 7.33 b–d | 2.66 j–m | 3.33 h–k | 5.33 c–e |
| 250 | 3.33 k–n | 4.33 h–k | 7.00 c–e | 2.33 k–n | 3.00 i–l | 5.00 d–f | |
| 500 | 3.00 l–n | 4.00 i–l | 6.33 d–f | 2.00 l–n | 2.33 k–n | 4.66 e–g | |
| Sumi 8 | 125 | 4.66 h–j | 6.33 d–f | 8.33 a,b | 3.33 h–k | 4.66 e–g | 6.66 c |
| 250 | 4.00 i–l | 5.33 f–h | 8.00 a–c | 3.00 i–l | 4.00 f–i | 6.33 b,c | |
| 500 | 3.66 j–l | 4.66 h–j | 7.33 b–d | 2.66 j–m | 3.66 g–j | 6.00 b–d | |
| L.S.D. | |||||||
| Varieties | 0.368 | 0.289 | |||||
| Concent. | 0.425 | 0.334 | |||||
| Fungicides | 0.368 | 0.289 | |||||
| Varieties × Concent. | 0.736 | 0.579 | |||||
| Varieties × Fungicides | 0.638 | 0.501 | |||||
| Concent. × Fungicide | 0.736 | 0.579 | |||||
| Varieties × Concent. × Fungicides | 1.276 | 1.003 | |||||
| (a) | |||||||
| Fungicides | Concentration (ppm) | Pre-Inoculation | Post-Inoculation | ||||
| Misr1 | Gimmeiza11 | Sids12 | Misr1 | Gimmeiza11 | Sids12 | ||
| Control | 0 | 59.5 h | 46.4 r | 35.4 z | 61.5 j | 50.4 q | 39.4 x |
| Tilit | 125 | 67.4 b | 55.7 k | 43.4 t | 68.7 c | 58.4 l | 47.4 s |
| 250 | 69.2 a | 57.6 j | 45.1 s | 70 b | 60.1 k | 49.1 r | |
| 500 | 69.7 a | 58.5 i | 45.6 s | 70.6 a | 60.5 k | 49.6 r | |
| Montoro | 125 | 63.9 e | 52.4 n | 40.2 v | 65.8 f | 55.1 n | 44.2 u |
| 250 | 65.4 d | 53.6 m | 41.9 u | 67.1 e | 56.9 m | 46.2 t | |
| 500 | 66.5 c | 54.5 l | 42.3 u | 67.7 d | 57.3 m | 46.3 t | |
| Sumi 8 | 125 | 60.4 g | 48.4 q | 36.9 y | 62.9 i | 51.9 p | 40.9 w |
| 250 | 62.2 f | 49.6 p | 38.3 x | 64.2 h | 53.6 o | 42.7 v | |
| 500 | 62.7 f | 50.8 o | 39.1 w | 64.9 g | 54.1 o | 43.1 v | |
| L.S.D. | |||||||
| Varieties | 0.223 | 0.423 | |||||
| Fungicides | 0.376 | 0.257 | |||||
| varieties × Fungicides | 0.652 | 0.590 | |||||
| (b) | |||||||
| Chemical Inducers | Concentration (ppm) | Pre-Inoculation | Post-Inoculation | ||||
| Misr1 | Gimmeiza11 | Sids12 | Misr1 | Gimmeiza11 | Sids12 | ||
| Control | 0 | 59.0 i | 45.6 s | 35 z | 60.2 g | 48.0 n | 37.1 u |
| Salicylic acid | 250 | 67.2 c | 55.7 l | 43 u | 68.3 b | 56.4 i | 44.4 p |
| 500 | 69.6 b | 56.2 k | 43.4 u | 70.0 a | 58.1 h | 44.8 p | |
| 1000 | 69.6 a | 57.4 j | 44.5 t | 70.4 a | 58.5 h | 46.5 o | |
| Indole acetic acid | 25 | 63.7 e | 52.2 o | 39.2 w | 65.9 d | 53.9 k | 41.2 r |
| 50 | 65.9 d | 52.6 n | 41.3 v | 66.3 d | 54.3 k | 43.3 q | |
| 100 | 66.1 d | 53.8 m | 41.3 v | 67.3 c | 55.3 j | 43.3 q | |
| Oxalic acid | 250 | 60.2 h | 47.8 r | 36.1 y | 62.2 f | 50.1 m | 38.1 t |
| 500 | 62.0 g | 49.7 q | 37.8 x | 63.9 e | 51.8 l | 38.5 t | |
| 1000 | 62.5 f | 50.2 p | 38.2 x | 64.2 e | 52.2 l | 40.2 s | |
| L.S.D. | |||||||
| Varieties | 0.114 | 0.124 | |||||
| Inducers | 0.198 | 0.215 | |||||
| Varieties × Inducers | 0.396 | 0.431 | |||||
| (a) | |||||||
| Fungicides | Concentration (ppm) | Pre-Inoculation | Post-Inoculation | ||||
| Misr1 | Gimmeiza11 | Sids12 | Misr1 | Gimmeiza11 | Sids12 | ||
| Control | 0 | 53.8 j | 41.6 r | 31.4 y | 55.5 i | 44.6 q | 33.4 y |
| Tilit | 125 | 63.1 c | 51.2 l | 39.2 t | 64.3 b | 53 k | 42.3 s |
| 250 | 63.6 b | 51.4 l | 39.5 t | 64.4 b | 53.4 k | 42.4 s | |
| 500 | 64.8 a | 52.5 k | 40.6 s | 65.5 a | 54.6 j | 43.5 r | |
| Montoro | 125 | 59.5 f | 47.3 o | 36.1 v | 60.3 e | 49.7 m | 38.6 u |
| 250 | 60 e | 47.9 n | 36.5 v | 61.4 d | 50.1 m | 39.1 u | |
| 500 | 61.2 d | 49 m | 37.5 u | 62.2 c | 51.2 l | 40.2 t | |
| Sumi 8 | 125 | 55.9 i | 44.2 q | 33.1 x | 57 h | 46 p | 35 x |
| 250 | 56.4 h | 44.3 q | 33.4 x | 57.7 g | 46.8 o | 35.8 w | |
| 500 | 57.6 g | 45.5 p | 34.4 w | 58.9 f | 47.9 n | 36.9 v | |
| L.S.D. | |||||||
| Varieties | 0.119 | 0.185 | |||||
| Fungicides | 0.272 | 0.271 | |||||
| Varieties × Fungicides | 0.461 | 0.479 | |||||
| (b) | |||||||
| Chemical Inducers | Concentration (ppm) | Pre-Inoculation | Post-Inoculation | ||||
| Misr1 | Gimmeiza11 | Sids12 | Misr1 | Gimmeiza11 | Sids12 | ||
| Control | 0 | 52.8 g | 40.9 o | 30.6 y | 54.5 i | 43.1 q | 31.9 y |
| Salicylic acid | 250 | 62.2 b | 49.1 i | 37.7 r | 62.9 c | 51.3 k | 39.1 t |
| 500 | 64.4 a | 51.3 h | 39.3 q | 64.4 b | 53.0 j | 40.5 s | |
| 1000 | 64.6 a | 51.4 h | 39.8 p | 65.3 a | 53.3 j | 41.1 r | |
| Indole acetic acid | 25 | 58.6 d | 45.6 l | 34.7 u | 59.3 e | 48.2 n | 36.0 v |
| 50 | 60.8 c | 47.4 k | 36.2 t | 61.4 d | 49.6 m | 37.7 u | |
| 100 | 61.0 c | 47.9 j | 36.7 s | 61.7 d | 50.3 l | 38.1 u | |
| Oxalic acid | 250 | 55.0 f | 42.1 n | 31.6 x | 55.7 h | 45.2 p | 32.9 x |
| 500 | 57.2 e | 44.2 m | 33.3 w | 57.5 g | 46.8 o | 34.6 w | |
| 1000 | 57.4 e | 44.4 m | 33.7 v | 58.1 f | 47.2 o | 35.0 w | |
| L.S.D. | |||||||
| Varieties | 0.231 | 0.097 | |||||
| Inducers | 0.172 | 0.284 | |||||
| Varieties × Inducers | 0.361 | 0.476 | |||||
| (a) | |||||||
| Fungicides | Concentration (ppm) | Pre-Inoculation | Post-Inoculation | ||||
| Misr1 | Gimmeiza11 | Sids12 | Misr1 | Gimmeiza11 | Sids12 | ||
| Control | 0 | 51.4 j | 38.6 r | 26.4 z | 53.4 g | 40.5 p | 28.4 y |
| Tilit | 125 | 61.7 c | 47.8 l | 34.9 u | 62.8 b | 49.7 j | 37 r |
| 250 | 64.2 b | 50.1 k | 36.8 t | 65.1 a | 51.7 i | 39.2 q | |
| 500 | 64.6 a | 50.4 k | 37.3 s | 65.5 a | 52.3 h | 39.4 q | |
| Montoro | 125 | 57.3 f | 44 n | 31.3 w | 58.9 d | 45.9 l | 33.4 u |
| 250 | 59.1 e | 46.3 m | 33.5 v | 61.2 c | 48.2 k | 34.9 t | |
| 500 | 60.2 C | 46.5 m | 33.7 v | 61.5 c | 48.4 k | 35.8 s | |
| Sumi 8 | 125 | 52.9 i | 40.1 q | 27.7 y | 54.9 f | 42 o | 29.8 x |
| 250 | 55.4 h | 42 p | 29.9 x | 57.2 e | 43.9 n | 31.7 w | |
| 500 | 55.8 g | 42.7 o | 30.1 x | 57.5 e | 44.6 m | 32.2 v | |
| L.S.D. | |||||||
| Varieties | 0.048 | 0.179 | |||||
| Fungicides | 0.173 | 0.192 | |||||
| Varieties × Fungicides | 0.289 | 0.361 | |||||
| (b) | |||||||
| Chemical Inducers | Concentration (ppm) | Pre-Inoculation | Post-Inoculation | ||||
| Misr1 | Gimmeiza11 | Sids12 | Misr1 | Gimmeiza11 | Sids12 | ||
| Control | 0 | 51.0 g | 37.5 n | 25.5 w | 53.0 h | 40.1 o | 27.2 x |
| Salicylic acid | 250 | 61.3 b | 46.9 i | 33.9 q | 62.1 b | 49.2 j | 35.9 r |
| 500 | 64.1 a | 49.5 h | 35.9 p | 64.4 a | 51.5 i | 37.1 q | |
| 1000 | 64.3 a | 49.6 h | 36.3 o | 64.6 a | 51.7 i | 38.0 p | |
| Indole acetic acid | 25 | 58.2 d | 44.1 k | 30.3 s | 58.2 d | 45.3 l | 32.4 t |
| 50 | 58.4 d | 44.2 k | 32.5 r | 60.5 c | 45.5 l | 34.6 s | |
| 100 | 59.9 c | 45.6 j | 32.7 r | 60.8 c | 47.9 k | 35.0 s | |
| Oxalic acid | 250 | 52.5 f | 38.5 m | 27.0 v | 54.3 g | 41.1 n | 28.8 w |
| 500 | 55.2 e | 41.4 l | 27.9 u | 56.3 f | 43.7 m | 29.3 v | |
| 1000 | 55.4 e | 41.5 l | 29.1 t | 56.9 e | 44 m | 31.1 u | |
| L.S.D. | |||||||
| Varieties | 0.112 | 0.108 | |||||
| Inducers | 0.194 | 0.188 | |||||
| Varieties × Inducers | 0.337 | 0.376 | |||||
| (a) | |||||||
| Fungicides | Concentration (ppm) | Pre-Inoculation | Post-Inoculation | ||||
| Misr1 | Gimmeiza11 | Sids12 | Misr1 | Gimmeiza11 | Sids12 | ||
| Control | 0 | 8.38 h | 6.31 o | 4.20 v | 8.57 h | 6.51 q | 4.40 z |
| Tilit | 125 | 9.56 b | 7.99 j | 5.84 q | 9.76 b | 8.10 k | 6.01 t |
| 250 | 9.59 b | 8.00 j | 5.89 q | 9.78 b | 8.19 j | 6.09 s | |
| 500 | 9.74 a | 8.21 i | 6.10 p | 9.93 a | 8.40 i | 6.30 r | |
| Montoro | 125 | 8.98 e | 7.35 l | 5.21 s | 9.24 e | 7.51 m | 5.38 w |
| 250 | 9.13 k | 7.39 l | 5.26 s | 9.32 d | 7.56 m | 5.46 v | |
| 500 | 9.28 c | 7.58 k | 5.47 r | 9.47 c | 7.77 l | 5.67 u | |
| Sumi 8 | 125 | 8.68 g | 6.68 n | 4.54 u | 8.85 g | 6.85 p | 4.77 y |
| 250 | 8.69 g | 6.73 n | 4.59 u | 8.87 g | 6.93 o | 4.82 y | |
| 500 | 8.83 f | 6.94 m | 4.83 t | 9.03 f | 7.16 n | 5.03 x | |
| L.S.D. | |||||||
| Varieties | 0.022 | 0.031 | |||||
| Fungicides | 0.029 | 0.045 | |||||
| Varieties × Fungicides | 0.053 | 0.067 | |||||
| (b) | |||||||
| Chemical Inducers | Concentration (ppm) | Pre-Inoculation | Post-Inoculation | ||||
| Misr1 | Gimmeiza11 | Sids12 | Misr1 | Gimmeiza11 | Sids12 | ||
| Control | 0 | 7.56 h | 6.00 p | 3.99 y | 7.93 h | 6.14 q | 4.35 z |
| Salicylic acid | 250 | 8.64 b | 7.24 j | 5.46 s | 8.98 b | 7.40 j | 5.60 s |
| 500 | 8.94 a | 7.26 j | 5.84 r | 9.26 a | 7.74 i | 5.92 r | |
| 1000 | 8.95 a | 7.41 i | 5.89 q | 9.29 a | 7.76 i | 5.95 r | |
| Indole acetic acid | 25 | 8.21 e | 6.77 m | 4.83 u | 8.53 e | 6.86 m | 5.07 v |
| 50 | 8.42 d | 6.92 l | 5.24 t | 8.78 d | 7.15 l | 5.35 u | |
| 100 | 8.49 c | 7.00 k | 5.28 t | 8.83 c | 7.22 k | 5.43 t | |
| Oxalic acid | 250 | 7.81 g | 6.25 o | 4.24 x | 8.08 g | 6.32 p | 4.53 y |
| 500 | 7.94 f | 6.48 n | 4.34 w | 8.36 f | 6.63 o | 4.81 x | |
| 1000 | 7.98 f | 6.51 n | 4.62 v | 8.38 f | 6.68 n | 4.89 w | |
| LSD. | |||||||
| Varieties | 0.025 | 0.021 | |||||
| Inducers | 0.029 | 0.024 | |||||
| Varieties × Inducers | 0.051 | 0.042 | |||||
| (a) | |||||||
| Fungicides | Concentration (ppm) | Pre-Inoculation | Post-Inoculation | ||||
| Misr1 | Gimmeiza11 | Sids12 | Misr1 | Gimmeiza11 | Sids12 | ||
| Control | 0 | 4.97 i | 3.17 p | 1.93 x | 5.27 i | 3.47 r | 2.23 y |
| Tilit | 125 | 6.23 c | 4.43 k | 2.80 s | 6.53 c | 4.73 l | 3.07 t |
| 250 | 6.51 b | 4.74 j | 2.92 r | 6.81 b | 4.96 k | 3.29 s | |
| 500 | 6.59 a | 4.79 j | 3.01 q | 6.89 a | 5.09 j | 3.31 s | |
| Montoro | 125 | 5.69 f | 3.89 m | 2.41 u | 5.99 f | 4.19 o | 2.71 v |
| 250 | 5.94 e | 4.20 l | 2.63 t | 6.27 e | 4.43 n | 2.93 u | |
| 500 | 6.05 d | 4.25 l | 2.65 t | 6.35 d | 4.55 m | 2.95 u | |
| Sumi 8 | 125 | 5.15 h | 3.35 o | 2.05 w | 5.45 h | 3.65 q | 2.35 x |
| 250 | 5.49 g | 3.69 n | 2.23 v | 5.79 g | 3.96 p | 2.53 w | |
| 500 | 5.49 g | 3.71 n | 2.29 v | 5.81 g | 4.01 p | 2.59 w | |
| L.S.D. | |||||||
| Varieties | 0.023 | 0.037 | |||||
| Fungicides | 0.032 | 0.035 | |||||
| Varieties × Fungicides | 0.058 | 0.068 | |||||
| (b) | |||||||
| Chemical Inducers | Concentration (ppm) | Pre-Inoculation | Post-Inoculation | ||||
| Misr1 | Gimmeiza11 | Sids12 | Misr1 | Gimmeiza11 | Sids12 | ||
| Control | 0 | 4.49 h | 3.02 q | 1.49 x | 4.87 h | 3.37 q | 1.87 x |
| Salicylic acid | 250 | 5.54 b | 4.10 k | 2.54 s | 5.92 c | 4.42 j | 2.92 s |
| 500 | 5.79 a | 4.25 j | 2.79 r | 6.16 b | 4.67 i | 3.16 r | |
| 1000 | 5.84 a | 4.34 i | 2.84 r | 6.22 a | 4.72 i | 3.22 r | |
| Indole acetic acid | 25 | 5.12 d | 3.59 m | 2.12 u | 5.47 e | 4.06 m | 2.47 u |
| 50 | 5.37 c | 3.84 l | 2.17 u | 5.75 d | 4.14 l | 2.72 t | |
| 100 | 5.39 c | 3.89 l | 2.39 t | 5.77 d | 4.27 k | 2.77 t | |
| Oxalic acid | 250 | 4.67 g | 3.14 p | 1.64 w | 5.08 g | 3.52 p | 2.08 w |
| 500 | 4.85 f | 3.33 o | 1.65 w | 5.12 g | 3.73 o | 2.27 v | |
| 1000 | 4.94 e | 3.44 n | 1.94 v | 5.32 f | 3.82 n | 2.32 v | |
| L.S.D. | |||||||
| Varieties | 0.027 | 0.028 | |||||
| Inducers | 0.031 | 0.033 | |||||
| varieties × Inducers | 0.054 | 0.057 | |||||
| SN | Common Name | Trade Name | Active Ingredient |
|---|---|---|---|
| 1 | Propiconazole | Tilt | 25 EC |
| 2 | Difenoconazole + Propiconazole | Montoro | 30 EC |
| 3 | Diniconazole | Sumi eight | 5 EC |
| Infection Type | Infection Class | Symptoms |
|---|---|---|
| 0 | Immune | No visible symptoms |
| 1 | High resistant | Necrotic flecks without sporulation |
| 2 | Resistant | Necrosis without sporulation |
| 3 | Moderately resistant | Necrosis with trace sporulation |
| 4 | Light–moderate | Light sporulation surrounded by necrosis |
| 5 | Moderate | Intermediate sporulation with necrosis/chlorosis |
| 6 | High–moderate | Moderate sporulation surrounded by chlorosis |
| 7 | Moderately susceptible | Moderate sporulation with moderate chlorosis |
| 8 | Susceptible | Sufficient sporulation with little or no chlorosis |
| 9 | Very susceptible | Abundant sporulation without chlorosis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zakaria, W.G.E.; Atia, M.M.; Ali, A.Z.; Abbas, E.E.A.; Salim, B.M.A.; Marey, S.A.; Hatamleh, A.A.; Elnahal, A.S.M. Assessing the Effectiveness of Eco-Friendly Management Approaches for Controlling Wheat Yellow Rust and Their Impact on Antioxidant Enzymes. Plants 2023, 12, 2954. https://doi.org/10.3390/plants12162954
Zakaria WGE, Atia MM, Ali AZ, Abbas EEA, Salim BMA, Marey SA, Hatamleh AA, Elnahal ASM. Assessing the Effectiveness of Eco-Friendly Management Approaches for Controlling Wheat Yellow Rust and Their Impact on Antioxidant Enzymes. Plants. 2023; 12(16):2954. https://doi.org/10.3390/plants12162954
Chicago/Turabian StyleZakaria, Waleed Gamal Eldein, Mahmoud Mohamed Atia, Ahmed Zaki Ali, Entsar E. A. Abbas, Bilkess M. A. Salim, Samy A. Marey, Ashraf Atef Hatamleh, and Ahmed Saeed Mohammed Elnahal. 2023. "Assessing the Effectiveness of Eco-Friendly Management Approaches for Controlling Wheat Yellow Rust and Their Impact on Antioxidant Enzymes" Plants 12, no. 16: 2954. https://doi.org/10.3390/plants12162954
APA StyleZakaria, W. G. E., Atia, M. M., Ali, A. Z., Abbas, E. E. A., Salim, B. M. A., Marey, S. A., Hatamleh, A. A., & Elnahal, A. S. M. (2023). Assessing the Effectiveness of Eco-Friendly Management Approaches for Controlling Wheat Yellow Rust and Their Impact on Antioxidant Enzymes. Plants, 12(16), 2954. https://doi.org/10.3390/plants12162954