
Resolving the Puzzle of Iris maackii (Iridaceae): A Morphological Insight into Its Taxonomy


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Morphological Study

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species | Origin | Voucher (*) |
|---|---|---|---|
| 1 | I. maackii | China, Ussuri River | R. Maack (LE01010783, holotype) |
| 2 | I. laevigata | Buryatia, Lake Baikal, near the Vydrinaya River estuary, 51°29′29.6″ N 104°50′39.3″ E | Yu.N. Pochinchik (VBGI) |
| 3 | Primorsky Krai, Bolshaya Ussurka River, Roshchino Village, 45°53′12.6″ N 134°50′48.3″ E | L.M. Pshennikova (VBGI) | |
| 4 | Primorsky Krai, Khasansky District, Cape L’va, 42°41′60.0″ N 131°14′12.3″ E | E.A. Chubar (VBGI) | |
| 5 | I. pseudacorus | Primorsky Krai, Vladivostok, BGI FEB RAS, 43°13′27″ N 131°59′38″ E | E.V. Boltenkov (VBGI, cult.) |
| 6 | Sakhalin Island, 3 km south of Shebunino Village | s. coll. (No. 2086, sub I. maackii) ** | |
| 7 | Sakhalin Island, Dolinsk | s. coll. (No. 2066, sub I. maackii) ** | |
| 8 | Rostov Oblast, Don River delta, 47°07′49.3″ N 39°28′07.7″ E | A.N. Shmaraeva (RWBG) |
2.2. Morphometric Analysis
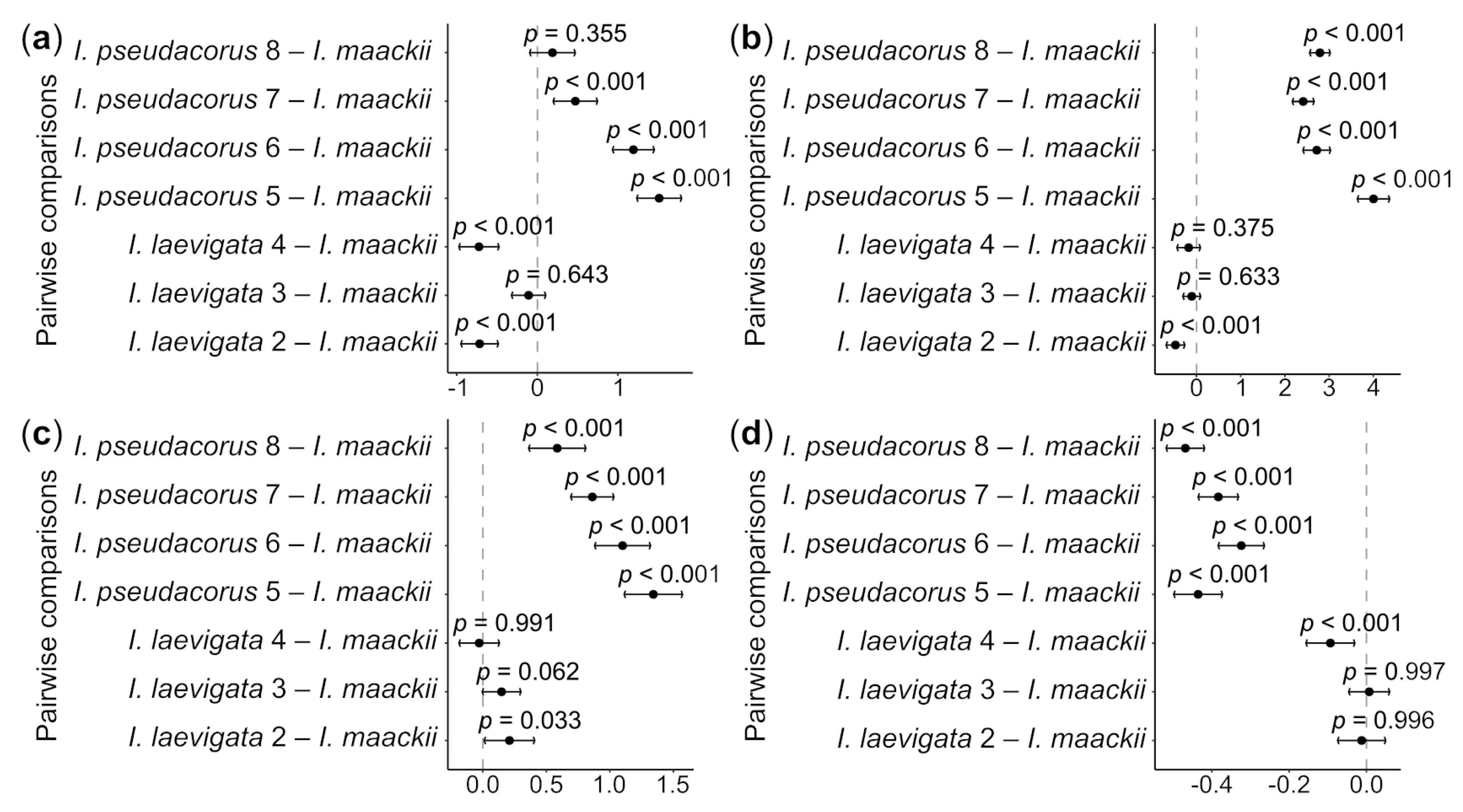
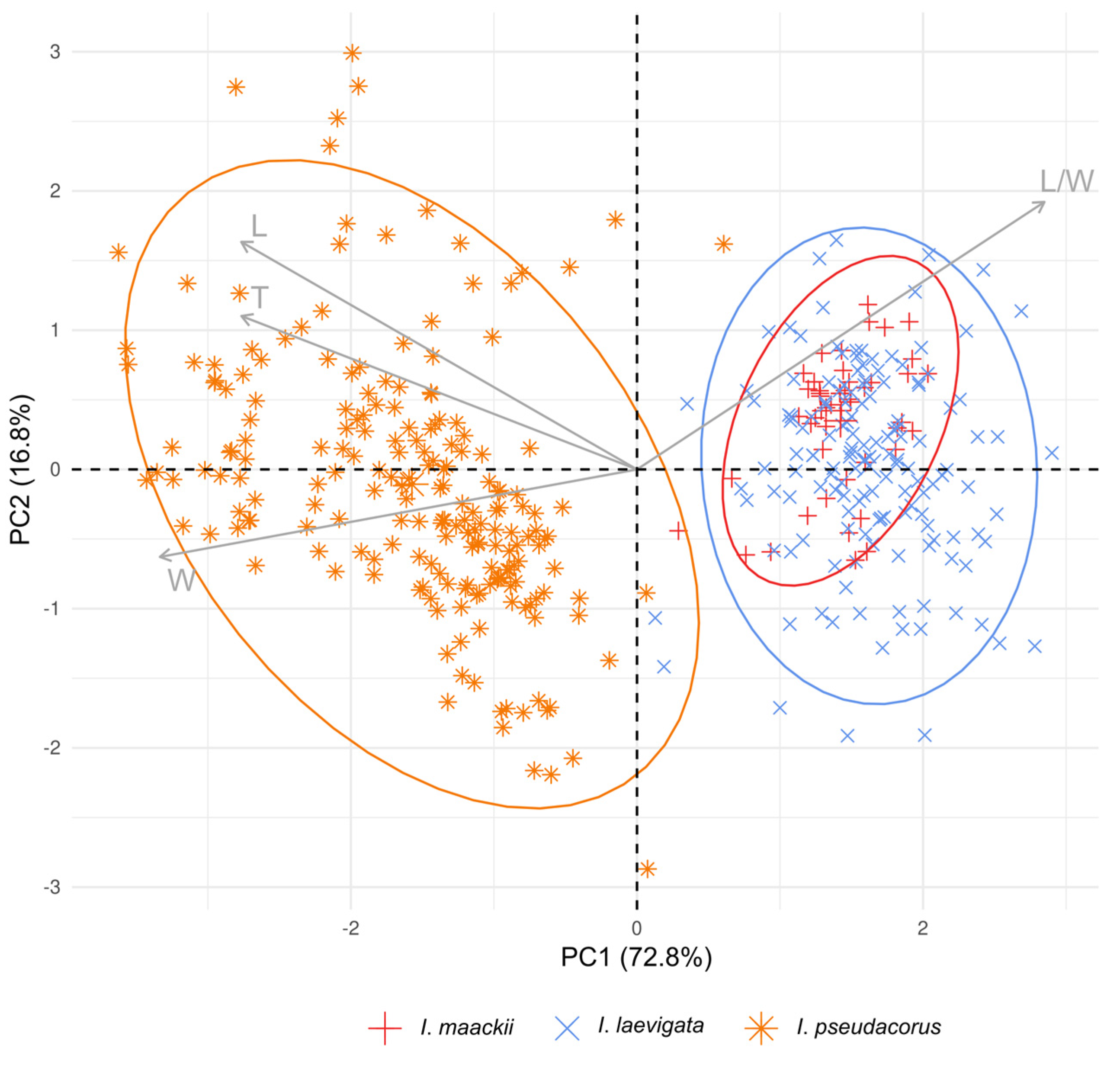
3. Results
4. Discussion
4.1. What Is Iris maackii from Northeast Asia According to the Literature?
4.2. What Is Iris maackii vs. I. laevigata and I. pseudacorus According to Morphology?
4.3. Taxonomic Treatment
5. Conclusions
Supplementary Materials
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maack, R. Journey Through the Valley of the Ussuri River; V. Bezobrazov & Co.: St. Petersburg, Russia, 1861; Volume 1. [Google Scholar]
- Regel, E. Tentamen Florae Ussuriensis Oder Versuch Einer Flora des Ussuri-Gebietes. Mémoires De L’académie Impériale Des Sci. De St. Pétersbourg Septième Série 1861, 4, 1–228. [Google Scholar]
- Maximowicz, C.J. Diagnoses plantarum novarum asiaticarum, III. Bull. Acad. Imp. Sci. Saint-Pétersbourg 1880, 26, 420–542. [Google Scholar]
- Fedtschenko, O. Irideen-Studien; Was ist Iris Maacki Maxim? Allg. Bot. Z. Syst. 1906, 12, 89–90. [Google Scholar]
- Komarov, V.L. Flora Manshuriae, III. Trudy Imp. S. Peterburgsk. Bot. Sada 1907, 25, 335–853. [Google Scholar]
- Koidzumi, G. Contributiones ad cognitionem florae Asiae orientalis. Bot. Mag. 1925, 39, 299–318. [Google Scholar] [CrossRef]
- Shishkin, I.K. Materialy k flore bassejna r. Imana (DVK, Khabarovskij okrug) [Materials to the flora of the Iman river basin (Khabarovsk Okrug)]. Zap. Vladivostoksk. Otd. Gosud. Russk. Geogr. Obsc. 1930, 5, 5–173. [Google Scholar]
- Fedtschenko, B.A. Iris. In Flora of the USSR; Komarov, V.L., Ed.; Izdatel’stvo Akademii nauk SSSR: Leningrad, Russia, 1935; Volume 4, pp. 511–557. [Google Scholar]
- Peckham, E.A.S. Alphabetical Iris Check List; Waverly Press: Baltimore, MD, USA, 1939. [Google Scholar]
- Kitagawa, M. Neo-Lineamenta Florae Manshuricae; J. Cramer: Vaduz, Liechtenstein, 1979. [Google Scholar]
- Pavlova, N.S. Iridaceae Juss. In Flora of the Russian Far East; Kozhevnikov, A.E., Probatova, N.S., Eds.; Dalnauka: Vladivostok, Russia, 2006; pp. 277–279. [Google Scholar]
- Linnegar, S. Series Laevigatae (Diels) Lawrence. In A Guide to Species Irises: Their Identification and Cultivation; The Species Group of the British Iris Society, Ed.; Cambridge University Press: Cambridge, UK, 2012; pp. 160–167. [Google Scholar]
- Hortipedia. Available online: https://en.hortipedia.com/Iris_laevigata (accessed on 13 August 2023).
- Lim, K.Y.; Matyasek, R.; Kovarik, A.; Leitch, A. Parental origin and genome evolution in the allopolyploid Iris versicolor. Ann. Bot. 2007, 100, 219–224. [Google Scholar] [CrossRef]
- Alekseeva, N.B. Genus Iris L. (Iridaceae) in Russia. Turczaninowia 2008, 11, 5–68. [Google Scholar]
- Alexeeva, N.B. Seed morphology in the genus Iris (Iridaceae) from Russia. Vavilovia 2020, 3, 5–28. [Google Scholar] [CrossRef]
- Catalogue of Life. Available online: https://www.catalogueoflife.org/data/taxon/3PZV6 (accessed on 13 August 2023).
- Catalogue of Life China 2023 Annual Checklist. Available online: http://www.sp2000.org.cn/species/show_species_details/T20171000043881 (accessed on 13 August 2023).
- National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=381594 (accessed on 13 August 2023).
- Plants of the World Online. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:438832-1 (accessed on 13 August 2023).
- The Global Biodiversity Information Facility. Available online: https://www.gbif.org/ (accessed on 13 August 2023).
- The Plant List. Available online: http://www.theplantlist.org/tpl1.1/record/kew-322090 (accessed on 13 August 2023).
- World Flora Online. Available online: http://www.worldfloraonline.org/taxon/wfo-0000783584 (accessed on 13 August 2023).
- World Plants. Available online: https://www.worldplants.de/world-plants-complete-list/complete-plant-list/#plantUid-74714 (accessed on 13 August 2023).
- Zhao, Y.-T.; Noltie, H.J.; Mathew, B. Iridaceae. In Flora of China; Wu, Z.-Y., Raven, P.H., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2000; Volume 24, pp. 297–313. [Google Scholar]
- Boltenkov, E.; Artyukova, E.; Kozyrenko, M.; Erst, A.; Trias-Blasi, A. Iris sanguinea is conspecific with I. sibirica (Iridaceae) according to morphology and plastid DNA sequence data. PeerJ 2020, 8, e10088. [Google Scholar] [CrossRef]
- Sutherland, W.J. Iris pseudacorus L. J. Ecol. 1990, 78, 833–848. [Google Scholar] [CrossRef]
- Rodionenko, G.I. On the taxonomic structure of Iris pseudacorus s.l. (Iridaceae). Bot. Zhurn. 2003, 88, 133–138. [Google Scholar]
- Yousefi, Z.; Mohseni-Bandpei, A. Nitrogen and phosphorus removal from wastewater by subsurface wetlands planted with Iris pseudacorus. Ecol. Eng. 2010, 36, 777–782. [Google Scholar] [CrossRef]
- Machado, A.I.; Fragoso, R.; Dordio, A.V.; Duarte, E. Performance of Iris pseudacorus and Typha domingensis for furosemide removal in a hydroponic system. Int. J. Phytoremediation 2020, 22, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, M.; Nawrot, N.; Wojciechowska, E.; Kuittinen, S.; Szczepańska, K.; Dembska, G.; Pappinen, A. Cadmium accumulation by Phragmites australis and Iris pseudacorus from stormwater in floating treatment wetlands microcosms: Insights into plant tolerance and utility for phytoremediation. J. Environ. Manag. 2023, 331, 117339. [Google Scholar] [CrossRef]
- Wang, Y.-N.; Cheng, L.-J.; Zhou, Q.-X. Phytoremediation of petroleum contaminated soils with Iris pseudacorus L. and the metabolic analysis in roots. J. Environ. Sci. 2016, 37, 1531–1538. [Google Scholar]
- Gervazoni, P.; Sosa, A.; Franceschini, C.; Coetzee, J.; Faltlhauser, A.; Fuentes-Rodriguez, D.; Martínez, A.; Hill, M. The alien invasive yellow flag (Iris pseudacorus L.) in Argentinian wetlands: Assessing geographical distribution through different data sources. Biol. Inv. 2020, 22, 3183–3193. [Google Scholar] [CrossRef]
- Asianflora. Available online: http://www.asianflora.com/Iridaceae/Iris-maackii.htm (accessed on 13 August 2023).
- Leipzig Catalogue of Vascular Plants. Available online: https://lifegate.idiv.de (accessed on 13 August 2023).
- Missouri Botanical Garden. Available online: http://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?taxonid=281340&isprofile=0& (accessed on 13 August 2023).
- SIGNA. Available online: http://www.signa.org/index.pl?Iris-maackii (accessed on 13 August 2023).
- The American Iris Society Iris Encyclopedia. Available online: https://wiki.irises.org/Spec/SpecMaackii (accessed on 13 August 2023).
- Wikipedia. Available online: https://en.wikipedia.org/wiki/Iris_maackii (accessed on 13 August 2023).
- Gallego-Tévar, B.; Grewell, B.J.; Whitcraft, C.R.; Futrell, J.C.; Bárcenas-Moreno, G.; Castillo, J.M. Contrasted impacts of yellow flag iris (Iris pseudacorus) on plant diversity in tidal wetlands within its native and invaded distribution ranges. Diversity 2022, 14, 326. [Google Scholar] [CrossRef]
- Minuti, G.; Stiers, I.; Coetzee, J.A. Climatic suitability and compatibility of the invasive Iris pseudacorus L. (Iridaceae) in the Southern Hemisphere: Considerations for biocontrol. Biol. Control 2022, 169, 104886. [Google Scholar] [CrossRef]
- Castillo, J.M.; Gallego-Tévar, B.; Grewell, B.J. Wrack burial limits germination and establishment of yellow flag iris (Iris pseudacorus L.). Plants 2023, 12, 1510. [Google Scholar] [CrossRef]
- Fischer, F.E.L.; Meyer, C.A. Index Quintus Seminum, Quae Hortus Botanicus Imperialis Petropolitanus Pro Mutua Commutatione Offert: Accedunt Animadversiones Botanicae Nonnullae; Academiae Caesareae Petropolitanae: Petropoli, Russia, 1839; Issue 5. [Google Scholar]
- Ponomarchuk, G.I.; Lyubitskaya, E.A. Floristic findings in south of Far East. Byull. Glavn. Bot. Sada 1983, 128, 51–52. [Google Scholar]
- Beentje, H. The Kew Plant Clossary; Kew Publishing: Kew, UK, 2012. [Google Scholar]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. (Eds.) International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017 [Regnum Vegetabile Volume 159]; Koeltz Botanical Books: Glashütten, Germany, 2018. [Google Scholar] [CrossRef]
- Index Herbariorum. Available online: https://sweetgum.nybg.org/ih/ (accessed on 13 August 2023).
- Ihaka, R.; Gentleman, R. R: A language for data analysis and graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.r-project.org/ (accessed on 13 August 2023).
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Navarro, D. Learning Statistics with R: A Tutorial for Psychology Students and Other Beginners. (Version 0.6.1). Available online: https://kpu.pressbooks.pub/learningstatistics/ (accessed on 13 August 2023).
- Dunnett, C.W. A multiple comparison procedure for comparing several treatments with a control. J. Am. Stat. Assoc. 1955, 50, 1096–1121. [Google Scholar] [CrossRef]
- Zeileis, A. Econometric computing with HC and HAC covariance matrix estimators. J. Stat. Soft. 2004, 11, 1–17. [Google Scholar] [CrossRef]
- Zeileis, A.; Köll, S.; Graham, N. Various versatile variances: An object-oriented implementation of clustered covariances in R. J. Stat. Softw. 2020, 95, 1–36. [Google Scholar] [CrossRef]
- Sneath, P.H.A.; Sokal, R.R. Numerical Taxonomy: The Principles and Practices of Numerical Classification; Freeman: San Francisco, CA, USA, 1973. [Google Scholar]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses, Version 1.0.7. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 13 August 2023).
- Linnaeus, C. Species Plantarum; Laurentii Salvii: Stockholm, Sweden, 1753; Volume 1. [Google Scholar] [CrossRef]
- Gmelin, J.G. Flora Sibirica sive Historia Plantarum Sibiriae; Typographia Academiae Scientiarum: St. Petersburg, Russia, 1747; Volume 1. [Google Scholar]
- Georgi, J.G. Bemerkungen einer Reise im Russischen Reich in den Jahren 1773 und 1774; Kaiserlichen Academie der Wissenschaften: St. Petersburg, Russia, 1775. [Google Scholar]
- Fedtschenko, B.A. Iridaceae. In Flora Transbaicalica; Fedtschenko, B.A., Ed.; Sumptibus Societatis Respublicanae Geographicae Troitzkosavsk–Kiachta: Leningrad, Russia, 1931; Volume 2, pp. 155–157. [Google Scholar]
- Komarov, V.L. Flora Manshuriae, I. Trudy Imp. S.-Peterburgsk. Bot. Sada 1901, 20, 1–559. [Google Scholar]
- Miyabe, K.; Kudô, Y. Flora of Hokkaido and Saghalien III: Monocotyledoneae Araceae to Orchidaceae. J. Fac. Agric. Hokkaido Imp. Univ. 1932, 26, 279–387. [Google Scholar]
- Voroschilov, V.N. Flora Sovetskogo Dal’nego Vostoka [Flora of the Soviet Far East]; Nauka: Moscow, Russia, 1966. [Google Scholar]
- Neczaeva, T.I. Iridaceae Juss. In Key for the Vascular Plants of Sakhalin and Kurile Islands; Tolmatchev, A.I., Ed.; Nauka: Leningrad, Russia, 1974; p. 121. [Google Scholar]
- Probatova, N.S.; Barkalov, V.Y.; Rudyka, E.G. Caryology of the Flora of Sakhalin and the Kurile Islands; Dalnauka: Vladivostok, Russia, 2007. [Google Scholar]
- Barkalov, V.Y. Flora of the Kuril Islands; Dalnauka: Vladivostok, Russia, 2009. [Google Scholar]
- Takahashi, H. Plants of the Kuril Islands; Hokkaido University Press: Sapporo, Japan, 2015. [Google Scholar]
- Ramtin, M.; Massiha, A.; Khoshkholgh-Pahlaviani, M.R.M.; Issazadeh, K.; Assmar, M.; Zarrabi, S. Evaluation of the antibacterial activities of essential oils of Iris pseudacorus and Urtica dioica. Zahedan J. Res. Med. Sci. 2014, 16, 35–39. [Google Scholar]
- Michalak, A.; Krauze-Baranowska, M.; Migas, P.; Kawiak, A.; Kokotkiewicz, A.; Królicka, A. Iris pseudacorus as an easily accessible source of antibacterial and cytotoxic compounds. J. Pharm. Biomed. Anal. 2021, 195, 113863. [Google Scholar] [CrossRef]
- Zhao, Y.-T. Some notes on the genus Iris of China. Acta Phytotax. Sin. 1980, 18, 53–62. Available online: https://www.jse.ac.cn/EN/Y1980/V18/I1/53 (accessed on 13 August 2023).
- Waddick, J.W.; Zhao, Y.-T. Iris of China; Timber Press: Portland, OR, USA, 1992. [Google Scholar]
- Fu, P.Y. Clavis Plantarum Chinae Boreali-Orientalis, 2nd ed.; Science Press: Beijing, China, 1995. [Google Scholar]
- Fu, L.; Hong, T. Higher Plants of China; Qingdao Publishing House: Qingdao, China, 2002; Volume 13, p. 286. [Google Scholar]
- Wu, Z.-Y.; Raven, P.H. Flora of China Illustrations; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2002; Volume 24, p. 344. [Google Scholar]
- Zhao, Y.-T. The wild irises of China. In Proceedings of the International Symposium on Iris, Celebrating the 50th Anniversary of the New Zealand Iris Society, Tauranga, New Zealand, 2–6 November 2000; pp. 39–43. [Google Scholar]
- Zheng, Y.; Meng, T.; Bi, X.; Lei, J. Investigation and evaluation of wild Iris resources in Liaoning Province, China. Genet. Resour. Crop Evol. 2017, 64, 967–978. [Google Scholar] [CrossRef]
- Jiménez-Mejías, P.; Luceño, M.; Martín-Bravo, S. Species boundaries within the southwest old world populations of the Carex flava group (Cyperaceae). Syst. Bot. 2014, 39, 117–131. [Google Scholar] [CrossRef]
- Dykes, W.R. Some garden irises. J. Roy. Hort. Soc. New Ser. 1914, 40, 226–233. [Google Scholar]
- Dykes, W.R. Iris laevigata, Fischer. Gard. Chron. Ser. 3 1920, 67, 274. [Google Scholar]
- Dykes, W.R. A Handbook of Garden Irises; M. Hopkinson & Co.: London, UK, 1924. [Google Scholar] [CrossRef]
- Alexeeva, N.B. Iridaceae Juss. In Catalogue of the Type Specimens of the Vascular Plants from Siberia and the Russian Far East kept in the Herbarium of the Komarov Botanical Institute (LE); Sokolova, I.V., Ed.; KMK Scientific Press: Moscow, Russia, 2012; pp. 415–419. [Google Scholar]
- Crespo, M.B. Nomenclatural types of Iberian irises (Iris and related genera, Iridaceae). Fl. Montiber. 2012, 53, 49–62. [Google Scholar]





| No. | Characters | Remarks |
|---|---|---|
| 1 | Rosette leaf width | Measured at the largest part of the widest rosette leaf |
| 2 | Rosette leaf texture | When dry, the surface of rosette leaves is finely ribbed, lacks a prominent midrib (smoothed), or has 1–2 large median veins generally running very close together, resembling in appearance a midrib (ribbed) |
| 3 | Stem branching | Flowering stem classified as simple, bearing only the terminal cluster (designated as 0), or branched, with lateral clusters (designated as 1, etc.) |
| 4 | Shoot length | Measured for the shoot of the upper lateral cluster |
| 5 | Cauline leaf length | Measured from the base to the apex of the upper cauline leaf |
| 6 | Bract length | Measured from the base to the apex of the outer bract of the terminal cluster |
| 7 | Bract texture | When fruiting, bracts dry or green |
| 8 | Pedicel length | Measured from the base of the terminal cluster to the ovary base of the first blooming flower |
| 9 | Fruit total | Number of fruit per stem |
| 10 | Fruit terminal | Number of fruit per terminal cluster |
| 11 | Fruit lateral | Number of fruit per upper lateral cluster |
| 12 | Fruit shape | Surface smoothed or angled, and characterized as sharply narrowed at the apex (obtuse) or conspicuously shortly beaked |
| 13 | Fruit length | Measured for the first fruit of the terminal cluster (using an Absolute Digimatic digital caliper, Mitutoyo, USA, to an accuracy of 0.1 mm) |
| 14 | Fruit width | |
| 15 | Seed shape | Inequilateral, tapering to hilum and chalaza, i.e., D-shaped or almost rounded |
| 16 | Seed color | Color was described in subjective terms |
| 17 | Seed length (L) | For seed characters, 50 samples from each collection site (see Table S2) were used (all were measured with the same сaliper) |
| 18 | Seed width (W) | |
| 19 | Seed thickness (T) | |
| 20 | Seed L/W ratio | The length-to-width ratio (L/W) provides additional data on the seed shape, i.e., degree of elongation along the hilum–chalaza axis (all calculations were manual) |
| No. | Characters | I. maackii | I. laevigata | I. pseudacorus |
|---|---|---|---|---|
| 1 | Rosette leaf width | 1.8 | 1.7 (0.85–2.9) | 3.0 (1.3–4.4) |
| 2 | Rosette leaf texture | Smoothed | Smoothed | Ribbed |
| 3 | Stem branching | 1 | 0–1 | 1–4 |
| 4 | Shoot length | 10.5 | 9 (4.2–13) | 4.7 (0.2–12.0) |
| 5 | Cauline leaf length | >16 | 18.7 (8.5–28.5) | 10.8 (4.7–26.2) |
| 6 | Bract length | ≥4.3 | 5.8 (3.8–10) | 5.7 (4–9.8) |
| 7 | Bract texture | Consisting of remnants | Dry | Green |
| 8 | Pedicel length | 2.0 | 2.1 (0.5–4.2) | 2.6 (1.4–4.8) |
| 9 | Fruit total | 7 | 4 (1–7) | 10 (5–17) |
| 10 | Fruit terminal | 4 | 4 (1–6) | 4 (2–5) |
| 11 | Fruit lateral | 3 | 1 (0–3) | 2 (1–3) |
| 12 | Fruit shape | Smoothed; obtuse | Smoothed; obtuse | 3-angled; beaked |
| 13 | Fruit length | 8.7 | 6.3 (4–8) | 7.6 (6–9.5) |
| 14 | Fruit width | 1.8 | 1.5 (1.2–1.8) | 1.9 (1.5–2.4) |
| 15 | Seed shape | Oblong, D-shaped | Mostly oblong, D-shaped, or almost round | Mostly almost round or D-shaped |
| 16 | Seed color | Brown | Brown | Brown |
| No. | Species | Seed | |||
|---|---|---|---|---|---|
| Length (L) | Width (W) | Thickness | L/W Ratio | ||
| 1 | I. maackii | 6.9 ± 0.5 (5.3–7.5) | 5.0 ± 0.4 (3.7–6.5) | 1.9 ± 0.3 (1.5–3.5) | 1.4 ± 0.1 (1.1–1.6) |
| 2 | I. laevigata | 6.2 ± 0.4 (4.5–6.9) | 4.6 ± 0.4 (3.8–5.4) | 2.1 ± 0.4 (1.5–3.6) | 1.4 ± 0.1 (1.1–1.7) |
| 3 | 6.8 ± 0.3 (6.1–7.5) | 4.9 ± 0.3 (4.3–5.8) | 2.1 ± 0.2 (1.6–2.5) | 1.4 ± 0.1 (1.1–1.6) | |
| 4 | 6.2 ± 0.5 (5.2–6.9) | 4.9 ± 0.6 (4.0–6.6) | 1.9 ± 0.3 (1.4–2.8) | 1.3 ± 0.1 (1.0–1.6) | |
| 5 | I. pseudacorus | 8.4 ± 0.6 (7.0–9.6) | 9.0 ± 0.9 (6.3–10.3) | 3.3 ± 0.5 (2.3–4.7) | 0.9 ± 0.1 (0.7–1.4) |
| 6 | 8.1 ± 0.5 (6.9–9.2) | 7.8 ± 0.7 (5.6–8.8) | 3.0 ± 0.5 (2.2–4.5) | 1.1 ± 0.1 (0.9–1.4) | |
| 7 | 7.4 ± 0.6 (6.4–8.9) | 7.5 ± 0.5 (5.2–8.4) | 2.8 ± 0.3 (2.2–3.5) | 1.0 ± 0.1 (0.9–1.5) | |
| 8 | 7.1 ± 0.6 (5.6–8.7) | 7.8 ± 0.5 (6.8–8.5) | 2.5 ± 0.5 (1.7–3.9) | 0.9 ± 0.1 (0.7–1.1) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boltenkov, E.V. Resolving the Puzzle of Iris maackii (Iridaceae): A Morphological Insight into Its Taxonomy. Plants 2023, 12, 3349. https://doi.org/10.3390/plants12193349
Boltenkov EV. Resolving the Puzzle of Iris maackii (Iridaceae): A Morphological Insight into Its Taxonomy. Plants. 2023; 12(19):3349. https://doi.org/10.3390/plants12193349
Chicago/Turabian StyleBoltenkov, Eugeny V. 2023. "Resolving the Puzzle of Iris maackii (Iridaceae): A Morphological Insight into Its Taxonomy" Plants 12, no. 19: 3349. https://doi.org/10.3390/plants12193349
APA StyleBoltenkov, E. V. (2023). Resolving the Puzzle of Iris maackii (Iridaceae): A Morphological Insight into Its Taxonomy. Plants, 12(19), 3349. https://doi.org/10.3390/plants12193349