
San Diego Thornmint (Acanthomintha ilicifolia) Populations Differ in Growth and Reproductive Responses to Differential Water Availability: Evidence from a Common Garden Experiment
 ,
,  and
and
Abstract
:1. Introduction
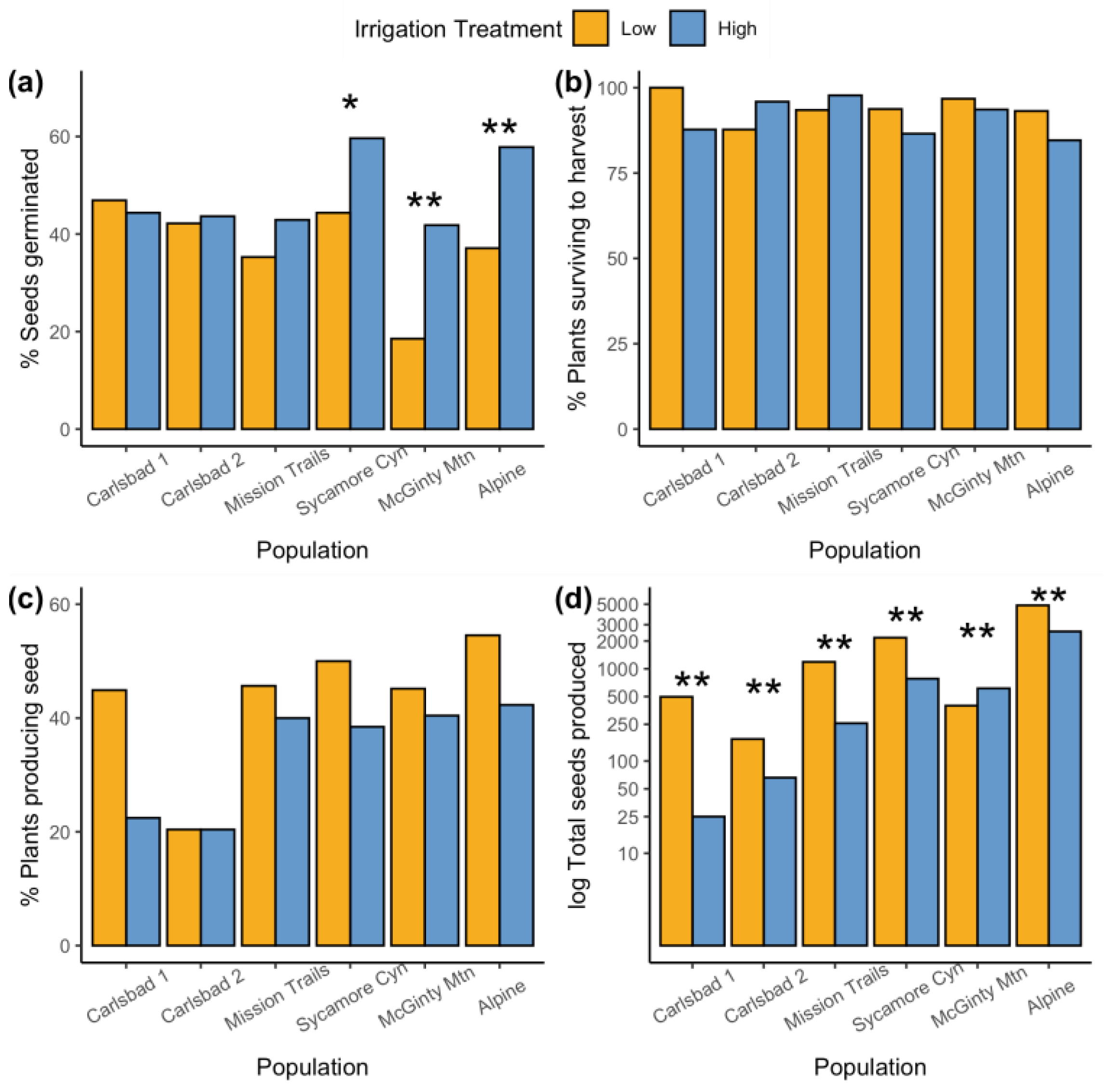
2. Results
2.1. Common Garden Experiment
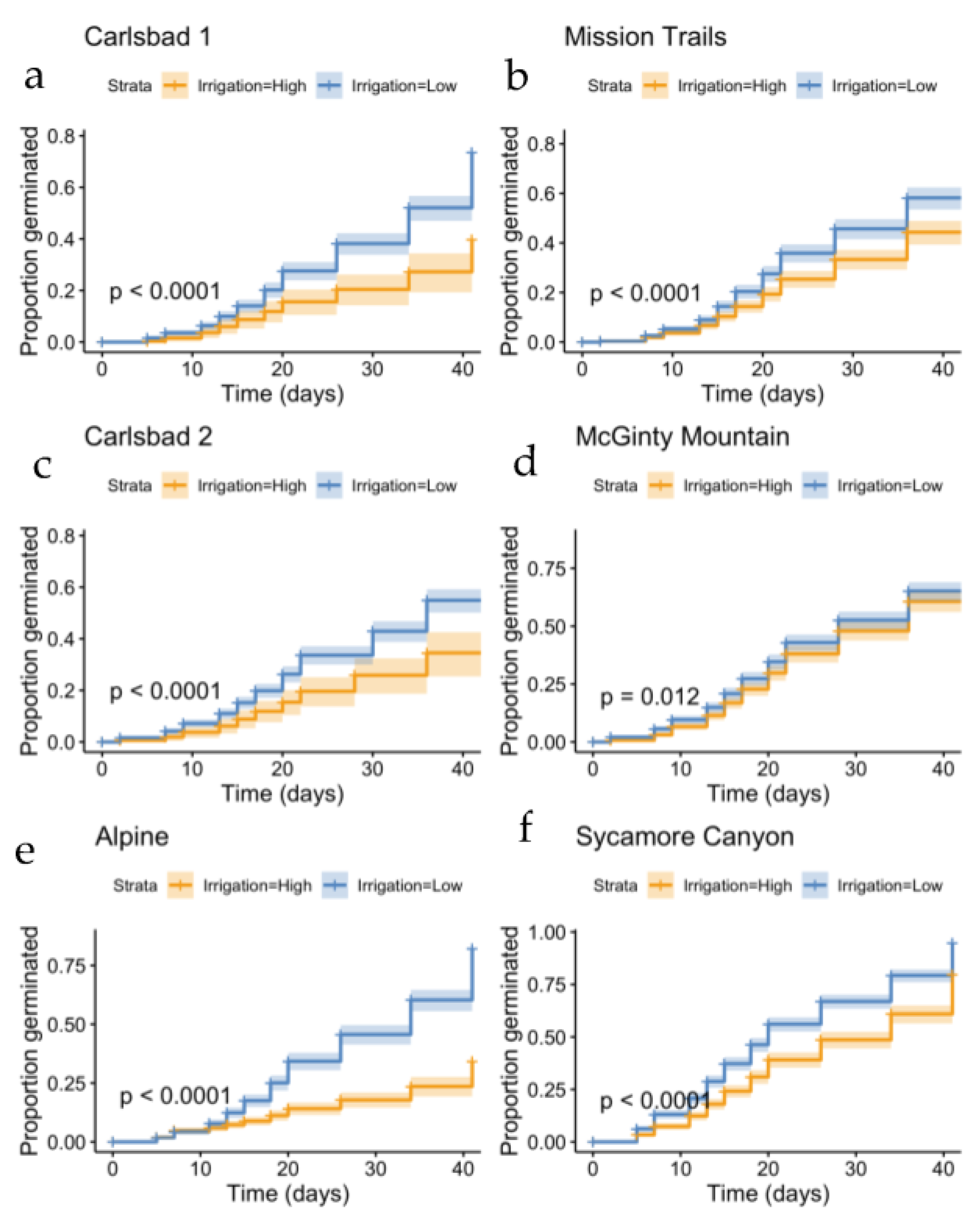
2.2. Ex Situ Germination Trials
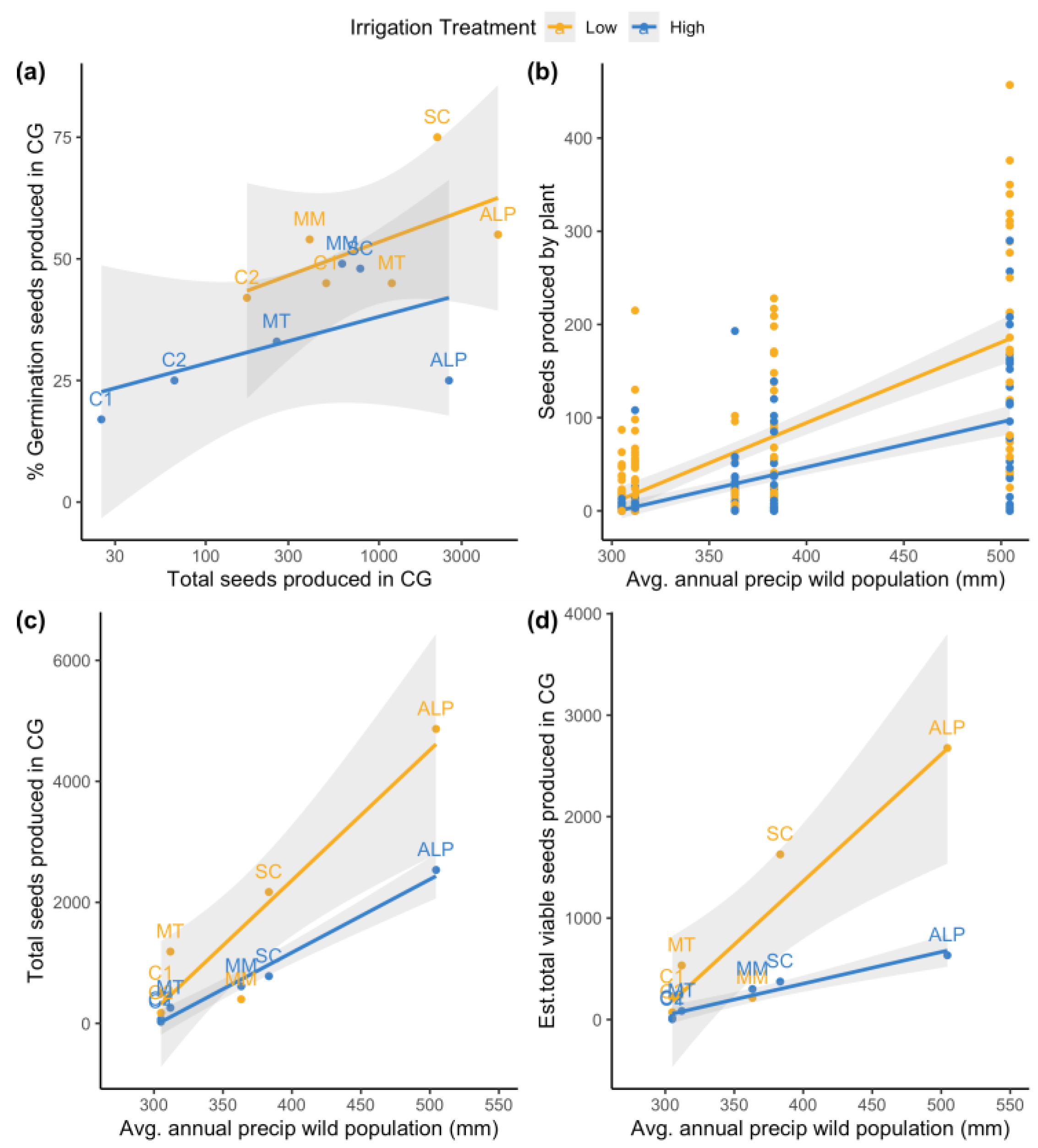
2.3. Relationship between Common Garden Reponses and Rainfall in Wild Source Populations
3. Discussion
3.1. Variable Growth Responses among Acanthomintha ilicifolia Populations in Common Garden

| Population Name | EO 3 (DeWoody et al. [22]) | Genetic Cluster (Milano et al. [23]) | Lat | Long | Elevation (m) | Wild Seed Collected | Mean Annual Rainfall (mm) 1 | CV Ann. Precip. 2 |
|---|---|---|---|---|---|---|---|---|
| Carlsbad1 (C1) | EO70A | Orange | 33.14 | −117.26 | 53 | 685 | 305 | 0.41 |
| Carlsbad2 (C2) | EO70B | Orange | 33.13 | −117.26 | 53 | 2757 | 305 | 0.41 |
| Sycamore Canyon (SC) | EO32-2 | Green/mixed | 32.93 | −116.98 | 341 | 975 | 383 | 0.32 |
| Mission Trails (MT) | EO33 | Green | 32.83 | −117.07 | 153 | 588 | 312 | 0.40 |
| McGinty Mountain (MM) | EO87l | Purple | 32.75 | −116.87 | 655 | 1369 | 363 | 0.34 |
| Alpine (ALP) | EO75 | Pink | 32.86 | −116.74 | 770 | 1705 | 504 | 0.25 |
3.2. Enhanced Reproductive Performance in Low-Irrigation Treatment
3.3. Conservation Strategy Implications of Results
4. Materials and Methods
4.1. Population Selection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
4.2. Common Garden Study

4.3. Germination Protocols
4.4. Statistical Methods
4.4.1. The Effect of Population and Watering Treatment on Plant Growth and Reproduction
4.4.2. The Effect of Watering Treatments on Ex Situ Seed Viability
4.4.3. The Effect of Source Population Climate and Geographic on ex Situ Reproductive Output
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Dependent Variable | Model Terms | X2 | df | p |
|---|---|---|---|---|
| Biomass at harvest | Treatment | 0.1 | 1 | 0.807 |
| Population | 12.2 | 5 | <0.001 | |
| Trt × pop | 19.1 | 5 | <0.001 | |
| Height at harvest | Treatment | 0.5 | 1 | 0.484 |
| Population | 175.0 | 5 | <0.001 | |
| Trt × pop | 19.8 | 5 | 0.001 | |
| Width at harvest | Treatment | 0.5 | 1 | 0.475 |
| Population | 74.0 | 5 | <0.001 | |
| Trt × pop | 43.1 | 5 | <0.001 | |
| Number of Floral nodes | Treatment | 9.2 | 1 | 0.002 |
| Population | 86.7 | 5 | <0.001 | |
| Trt × pop | 65.0 | 5 | <0.001 | |
| Days to flower | Treatment | 31.0 | 1 | <0.001 |
| Population | 41.2 | 5 | <0.001 | |
| Trt × pop | 3.8 | 5 | 0.002 | |
| Number of Seed produced | Treatment | 22.8 | 1 | <0.001 |
| Population | 165.5 | 5 | <0.001 | |
| Trt × pop | 13.0 | 5 | 0.024 | |
| Number of Seed/biomass | Treatment | 24.0 | 1 | <0.001 |
| Population | 199.8 | 5 | <0.001 | |
| Trt × pop | 14.5 | 5 | 0.013 | |
| % Germination | Treatment | 66.5 | 1 | <0.001 |
| Population | 41.4 | 5 | <0.001 | |
| Trt × pop | 35.2 | 5 | <0.001 | |
| % Survival | Treatment | 3.1 | 1 | 0.651 |
| Population | 3.3 | 5 | 0.08 | |
| Trt × pop | 15.3 | 5 | 0.009 | |
| % Plants producing seeds | Treatment | 6.1 | 5 | 0.014 |
| Population | 21.7 | 1 | <0.001 |
| Dependent Variable | Model Terms | X2 | df | p |
|---|---|---|---|---|
| Biomass at harvest | None retained | - | - | - |
| Height at harvest | None retained | - | - | - |
| Width at harvest | None retained | - | - | - |
| Number of floral nodes | Treatment | 6.5 | 1 | 0.011 |
| Days to flower | Treatment | 30.3 | 1 | <0.001 |
| Number of Seed produced | Treatment | 23.8 | 1 | <0.001 |
| Ann. rain wild pop | 74.4 | 1 | <0.001 | |
| Trt × ann. rain | 8.2 | 1 | 0.004 | |
| Number of Seed/biomass | Treatment | 24.5 | 1 | <0.001 |
| Ann. rain wild pop | 65.7 | 1 | <0.001 | |
| Trt × ann. rain | 13.4 | 1 | <0.001 | |
| % Germination | Treatment | 40.3 | 1 | <0.001 |
| Ann. rain wild pop | 0.5 | 1 | 0.49 | |
| Trt × ann. rain | 16.3 | 1 | <0.001 | |
| % Survival | None retained | - | - | - |
| % Plants producing seed | Treatment | 5.9 | 1 | 0.024 |
| Ann. rain wild pop | 5.1 | 1 | 0.016 |

| Population | Generation–Treatment | Ex Situ Germination (2019) |
|---|---|---|
| Carlsbad 1 | Wild | 100% |
| CG-Hi | 17% | |
| CG-Low | 45% | |
| Carlsbad 2 | Wild | 95% |
| CG-Hi | 25% | |
| CG-Low | 42% | |
| Alpine | Wild | N/A |
| CG-Hi | 25% | |
| CG-Low | 55% | |
| Mission Trails | Wild | 85% |
| CG-Hi | 33% | |
| CG-Low | 45% | |
| McGinty Mountain | Wild | 96% |
| CG-Hi | 49% | |
| CG-Low | 54% | |
| Sycamore | Wild | 95% |
| CG-Hi | 48% | |
| CG-Low | 75% |

| Dependent Variable | Model Terms | X2 | df | p |
|---|---|---|---|---|
| ANNUAL RAINFALL | ||||
| Total seeds | Ann. rain wild pop | 52.1 | 1 | <0.001 |
| Treatment | 10.1 | 1 | 0.002 | |
| Trt × ann. rain | 6.4 | 1 | 0.011 | |
| Ex situ germ | Treatment | 23.1 | 1 | <0.001 |
| Total viable seeds | Ann. rain wild pop | 36.2 | 1 | <0.001 |
| Treatment | 12.9 | 1 | <0.001 | |
| Trt × ann. rain | 13 | 1 | <0.001 | |
| ELEVATION | ||||
| Total seeds | Treatment | 4.7 | 1 | 0.027 |
| Ex situ germ | Elevation | 23.1 | 1 | <0.001 |
| Treatment | 23.6 | 1 | <0.001 | |
| Total viable seeds | Treatment | 3.8 | 1 | 0.052 |
| LATITUDE | ||||
| Total seeds | Treatment | 4.8 | 1 | 0.027 |
| Ex situ germ | Treatment | 23.1 | 1 | <0.001 |
| Total viable seeds | Treatment | 3.8 | 1 | 0.052 |
| LONGITUDE | ||||
| Total seeds | Treatment | 5.6 | 1 | 0.017 |
| Longitude | 6.6 | 1 | 0.01 | |
| Trt × longitude | 1.8 | 1 | 0.181 | |
| Ex situ germ | Treatment | 23.1 | 1 | <0.001 |
| Total viable seeds | Treatment | 5.2 | 1 | 0.022 |
| Longitude | 6.5 | 1 | 0.011 | |
| Trt × longitude | 2.9 | 1 | 0.091 |
References
- Nic Lughadha, E.; Bachman, S.P.; Leão, T.C.C.; Forest, F.; Halley, J.M.; Moat, J.; Acedo, C.; Bacon, K.L.; Brewer, R.F.A.; Gâteblé, G.; et al. Extinction Risk and Threats to Plants and Fungi. Plants People Planet 2020, 2, 389–408. [Google Scholar] [CrossRef]
- Vincent, H.; Bornand, C.N.; Kempel, A.; Fischer, M. Rare Species Perform Worse than Widespread Species under Changed Climate. Biol. Conserv. 2020, 246, 108586. [Google Scholar] [CrossRef]
- Thuiller, W.; Albert, C.; Araújo, M.B.; Berry, P.M.; Cabeza, M.; Guisan, A.; Hickler, T.; Midgley, G.F.; Paterson, J.; Schurr, F.M.; et al. Predicting Global Change Impacts on Plant Species’ Distributions: Future Challenges. Perspect. Plant Ecol. Evol. Syst. 2008, 9, 137–152. [Google Scholar] [CrossRef]
- Enquist, B.J.; Feng, X.; Boyle, B.; Maitner, B.; Newman, E.A.; Jørgensen, P.M.; Roehrdanz, P.R.; Thiers, B.M.; Burger, J.R.; Corlett, R.T. The Commonness of Rarity: Global and Future Distribution of Rarity across Land Plants. Sci. Adv. 2019, 5, eaaz0414. [Google Scholar] [CrossRef] [PubMed]
- Bartholomeus, R.P.; Witte, J.-P.M.; van Bodegom, P.M.; van Dam, J.C.; Aerts, R. Climate Change Threatens Endangered Plant Species by Stronger and Interacting Water-Related Stresses. J. Geophys. Res. Biogeosci. 2011, 116, G4. [Google Scholar] [CrossRef]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, Migration or Extirpation: Climate Change Outcomes for Tree Populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef]
- Christmas, M.J.; Breed, M.F.; Lowe, A.J. Constraints to and Conservation Implications for Climate Change Adaptation in Plants. Conserv. Genet. 2016, 17, 305–320. [Google Scholar] [CrossRef]
- Guerrant, E.O.; Havens, K.; Vitt, P. Sampling for Effective Ex Situ Plant Conservation. Int. J. Plant Sci. 2014, 175, 11–20. [Google Scholar] [CrossRef]
- Hoban, S.; Volk, G.; Routson, K.J.; Walters, C.; Richards, C. Sampling Wild Species to Conserve Genetic Diversity. In North American Crop Wild Relatives, Volume 1: Conservation Strategies; Greene, S.L., Williams, K.A., Khoury, C.K., Kantar, M.B., Marek, L.F., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 209–228. ISBN 978-3-319-95101-0. [Google Scholar]
- McGlaughlin, M.E.; Riley, L.; Brandsrud, M.; Arcibal, E.; Helenurm, M.K.; Helenurm, K. How Much Is Enough? Minimum Sampling Intensity Required to Capture Extant Genetic Diversity in Ex Situ Seed Collections: Examples from the Endangered Plant Sibara Filifolia (Brassicaceae). Conserv. Genet. 2015, 16, 253–266. [Google Scholar] [CrossRef]
- Havens, K.; Vitt, P.; Still, S.; Kramer, A.T.; Fant, J.B.; Schatz, K. Seed Sourcing for Restoration in an Era of Climate Change. Nat. Areas J. 2015, 35, 122–133. [Google Scholar] [CrossRef]
- Neale, J.R. Genetic Considerations in Rare Plant Reintroduction: Practical Applications (or How Are We Doing?). In Plant Reintroduction in a Changing Climate: Promises and Perils; Maschinski, J., Haskins, K.E., Raven, P.H., Eds.; The Science and Practice of Ecological Restoration; Island Press/Center for Resource Economics: Washington, DC, USA, 2012; pp. 71–88. ISBN 978-1-61091-183-2. [Google Scholar]
- Christie, K.; Strauss, S. Along the Speciation Continuum: Quantifying Intrinsic and Extrinsic Isolating Barriers across Five Million Years of Evolutionary Divergence in California Jewelflowers. Evolution 2018, 72, 1063–1079. [Google Scholar] [CrossRef] [PubMed]
- Berend, K.; Haynes, K.; MacKenzie, C.M. Common Garden Experiments as a Dynamic Tool for Ecological Studies of Alpine Plants and Communities in Northeastern North America. Rhodora 2019, 121, 174–212. [Google Scholar] [CrossRef]
- Hamann, E.; Kesselring, H.; Stöcklin, J. Plant Responses to Simulated Warming and Drought: A Comparative Study of Functional Plasticity between Congeneric Mid and High Elevation Species. J. Plant Ecol. 2018, 11, 364–374. [Google Scholar] [CrossRef]
- Huxman, T.E.; Winkler, D.E.; Mooney, K.A. A Common Garden Super-Experiment: An Impossible Dream to Inspire Possible Synthesis. J. Ecol. 2022, 110, 997–1004. [Google Scholar] [CrossRef]
- Aronson, J.; Kigel, J.; Shmida, A. Reproductive Allocation Strategies in Desert and Mediterranean Populations of Annual Plants Grown with and without Water Stress. Oecologia 1993, 93, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Schwinning, S.; Lortie, C.J.; Esque, T.C.; DeFalco, L.A. What Common-Garden Experiments Tell Us about Climate Responses in Plants. J. Ecol. 2022, 110, 986–996. [Google Scholar] [CrossRef]
- Corlett, R.T.; Tomlinson, K.W. Climate Change and Edaphic Specialists: Irresistible Force Meets Immovable Object? Trends Ecol. Evol. 2020, 35, 367–376. [Google Scholar] [CrossRef]
- Montrone, A.; Saito, L.; Weisberg, P.J.; Gosejohan, M.; Merriam, K.; Mejia, J.F. Climate Change Impacts on Vernal Pool Hydrology and Vegetation in Northern California. J. Hydrol. 2019, 574, 1003–1013. [Google Scholar] [CrossRef]
- US Fish and Wildlife Service Acanthomintha Ilicifolia (San Diego Thornmint). 5-Year Review: Summary and Evaluation. In Acanthomintha Ilicifolia (San Diego thornmint). 5-Year Review: Summary and Evaluation; U.S. Fish and Wildlife Service Carlsbad Fish and Wildlife Office: Carlsbad, CA, USA, 2009. [Google Scholar]
- DeWoody, J.; Rogers, D.L.; Hipkins, V.D.; Endress, B.A. Spatially Explicit and Multi-Sourced Genetic Information Is Critical for Conservation of an Endangered Plant Species, San Diego Thornmint (Acanthomintha Ilicifolia). Conserv. Genet. 2018, 19, 893–907. [Google Scholar] [CrossRef]
- Milano, E.R.; Vandergast, A.G. Population Genomic Surveys for Six Rare Plant Species in San Diego County, California; US Geological Survey: Reston, VA, USA, 2018; Volume 2018-1175, p. 72. [Google Scholar]
- Leiblein-Wild, M.C.; Tackenberg, O. Phenotypic Variation of 38 European Ambrosia Artemisiifolia Populations Measured in a Common Garden Experiment. Biol. Invasions 2014, 16, 2003–2015. [Google Scholar] [CrossRef]
- McKay, J.K.; Bishop, J.G.; Lin, J.-Z.; Richards, J.H.; Sala, A.; Mitchell-Olds, T. Local Adaptation across a Climatic Gradient despite Small Effective Population Size in the Rare Sapphire Rockcress. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2001, 268, 1715–1721. [Google Scholar] [CrossRef] [PubMed]
- Matesanz, S.; Ramos-Muñoz, M.; Moncalvillo, B.; Rubio Teso, M.L.; García de Dionisio, S.L.; Romero, J.; Iriondo, J.M. Plasticity to Drought and Ecotypic Differentiation in Populations of a Crop Wild Relative. AoB Plants 2020, 12, plaa006. [Google Scholar] [CrossRef] [PubMed]
- Valliere, J.M.; Allen, E.B. Interactive Effects of Nitrogen Deposition and Drought-Stress on Plant-Soil Feedbacks of Artemisia Californica Seedlings. Plant Soil 2016, 403, 277–290. [Google Scholar] [CrossRef]
- Roach, D.A.; Wulff, R.D. Maternal Effects in Plants. Annu. Rev. Ecol. Syst. 1987, 18, 209–235. [Google Scholar] [CrossRef]
- Li, N.; Li, Y. Maternal Control of Seed Size in Plants. J. Exp. Bot. 2015, 66, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Gremer, J.R.; Kimball, S.; Keck, K.R.; Huxman, T.E.; Angert, A.L.; Venable, D.L. Water-Use Efficiency and Relative Growth Rate Mediate Competitive Interactions in Sonoran Desert Winter Annual Plants. Am. J. Bot. 2013, 100, 2009–2015. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.J.; Sultan, S.E.; Horgan-Kobelski, T.; Riggs, C. Adaptive Transgenerational Plasticity in an Annual Plant: Grandparental and Parental Drought Stress Enhance Performance of Seedlings in Dry Soil. Integr. Comp. Biol. 2012, 52, 77–88. [Google Scholar] [CrossRef]
- Bauder, E.T.; Sakrison, J.A. Mechanisms of Persistence of San Diego Thornmint (Acanthomintha Ilicifolia); San Diego State Department of Biology: San Diego, CA, USA, 1999; p. 54. [Google Scholar]
- Memmott, J.; Craze, P.G.; Waser, N.M.; Price, M.V. Global Warming and the Disruption of Plant–Pollinator Interactions. Ecol. Lett. 2007, 10, 710–717. [Google Scholar] [CrossRef]
- Iler, A.M.; CaraDonna, P.J.; Forrest, J.R.K.; Post, E. Demographic Consequences of Phenological Shifts in Response to Climate Change. Annu. Rev. Ecol. Evol. Syst. 2021, 52, 221–245. [Google Scholar] [CrossRef]
- White, F.J.; Hay, F.R.; Abeli, T.; Mondoni, A. Two Decades of Climate Change Alters Seed Longevity in an Alpine Herb: Implications for Ex Situ Seed Conservation. Alp Bot. 2023, 133, 11–20. [Google Scholar] [CrossRef]
- Welt, R.S.; Litt, A.; Franks, S.J. Analysis of Population Genetic Structure and Gene Flow in an Annual Plant before and after a Rapid Evolutionary Response to Drought. AoB Plants 2015, 7, plv026. [Google Scholar] [CrossRef] [PubMed]
- Center for Plant Conservation CPC Best Plant Conservation Practices to Support Species Survival in the Wild; Center for Plant Conservation: Escondido, CA, USA, 2019.
- PRISM Climate Group. Oregon State University PRISM Climate Explorer. Available online: https://prism.oregonstate.edu/terms/ (accessed on 28 February 2023).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Lme4: Linear Mixed-Effects Models Using Eigen and S4. R Package Version 1.1-34. CRAN. 2014. Available online: https://cran.r-project.org/web/packages/lme4/lme4.pdf (accessed on 28 February 2023).
- Romano, A.; Stevanato, P. Germination Data Analysis by Time-to-Event Approaches. Plants 2020, 9, 617. [Google Scholar] [CrossRef] [PubMed]
- Therneau, T.M.; Elizabeth, A.; Cynthia, C.; Lumley, T. Survival: Survival Analysis, R Package Version 3.5-7. CRAN.; 2023. Available online: https://cran.r-project.org/web/packages/survival/survival.pdf/ (accessed on 28 February 2023).



| Population | Irrigation Treatment | Sample Size at Harvest | Biomass (g) | Height (cm) | Width (cm) | Floral Nodes | Seeds Produced per Plant | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | |||
| Carlsbad 1 | High | 43 | 0.74 | 0.04 | 20.2 | 1.04 | 13.7 | 0.7 | 26.0 | 1.7 | 2.3 | 0.5 |
| Low | 49 | 0.83 | 0.03 | 21.6 | 0.98 | 14.9 | 0.5 | 32.2 | 1.7 | 22.5 | 4.7 | |
| Carlsbad 2 | High | 49 | 0.42 | 0.05 | 12.6 | 0.76 | 9.8 | 0.7 | 14.4 | 1.6 | 6.6 | 1.1 |
| Low | 47 | 0.57 | 0.03 | 14.0 | 0.62 | 13.0 | 0.5 | 22.4 | 1.3 | 17.3 | 4.7 | |
| Alpine | High | 44 | 0.61 | 0.03 | 11.4 | 0.44 | 10.7 | 0.4 | 16.7 | 1.0 | 115.2 | 17.5 |
| Low | 41 | 0.63 | 0.03 | 11.1 | 0.42 | 9.61 | 0.3 | 19.1 | 0.8 | 202.8 | 25.4 | |
| Mission Trails | High | 44 | 0.59 | 0.05 | 14.0 | 0.60 | 9.8 | 0.5 | 19.6 | 1.7 | 14.3 | 5.7 |
| Low | 43 | 0.76 | 0.04 | 14.9 | 0.52 | 11.2 | 0.4 | 26.6 | 1.4 | 56.5 | 10.3 | |
| McGinty Mountain | High | 44 | 1.05 | 0.07 | 17.9 | 0.80 | 12.9 | 0.8 | 25.0 | 1.6 | 32.3 | 9.8 |
| Low | 30 | 0.42 | 0.03 | 13.3 | 0.78 | 7.7 | 0.4 | 10.6 | 0.7 | 28.4 | 8.9 | |
| Sycamore Canyon | High | 45 | 0.64 | 0.04 | 15.5 | 0.71 | 13.4 | 0.7 | 16.9 | 0.9 | 39.0 | 9.9 |
| Low | 45 | 0.76 | 0.03 | 13.9 | 0.50 | 14.4 | 0.4 | 20.8 | 1.1 | 90.5 | 15.6 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heineman, K.D.; Anderson, S.M.; Davitt, J.M.; Lippitt, L.; Endress, B.A.; Horn, C.M. San Diego Thornmint (Acanthomintha ilicifolia) Populations Differ in Growth and Reproductive Responses to Differential Water Availability: Evidence from a Common Garden Experiment. Plants 2023, 12, 3439. https://doi.org/10.3390/plants12193439
Heineman KD, Anderson SM, Davitt JM, Lippitt L, Endress BA, Horn CM. San Diego Thornmint (Acanthomintha ilicifolia) Populations Differ in Growth and Reproductive Responses to Differential Water Availability: Evidence from a Common Garden Experiment. Plants. 2023; 12(19):3439. https://doi.org/10.3390/plants12193439
Chicago/Turabian StyleHeineman, Katherine D., Stacy M. Anderson, Joseph M. Davitt, Laurie Lippitt, Bryan A. Endress, and Christa M. Horn. 2023. "San Diego Thornmint (Acanthomintha ilicifolia) Populations Differ in Growth and Reproductive Responses to Differential Water Availability: Evidence from a Common Garden Experiment" Plants 12, no. 19: 3439. https://doi.org/10.3390/plants12193439
APA StyleHeineman, K. D., Anderson, S. M., Davitt, J. M., Lippitt, L., Endress, B. A., & Horn, C. M. (2023). San Diego Thornmint (Acanthomintha ilicifolia) Populations Differ in Growth and Reproductive Responses to Differential Water Availability: Evidence from a Common Garden Experiment. Plants, 12(19), 3439. https://doi.org/10.3390/plants12193439