

Evaluation of the Carbohydrate Composition of Crabapple Fruit Tissues Native to Northern Asia


 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Morphological Traits
2.2. Content of Ascorbic Acid
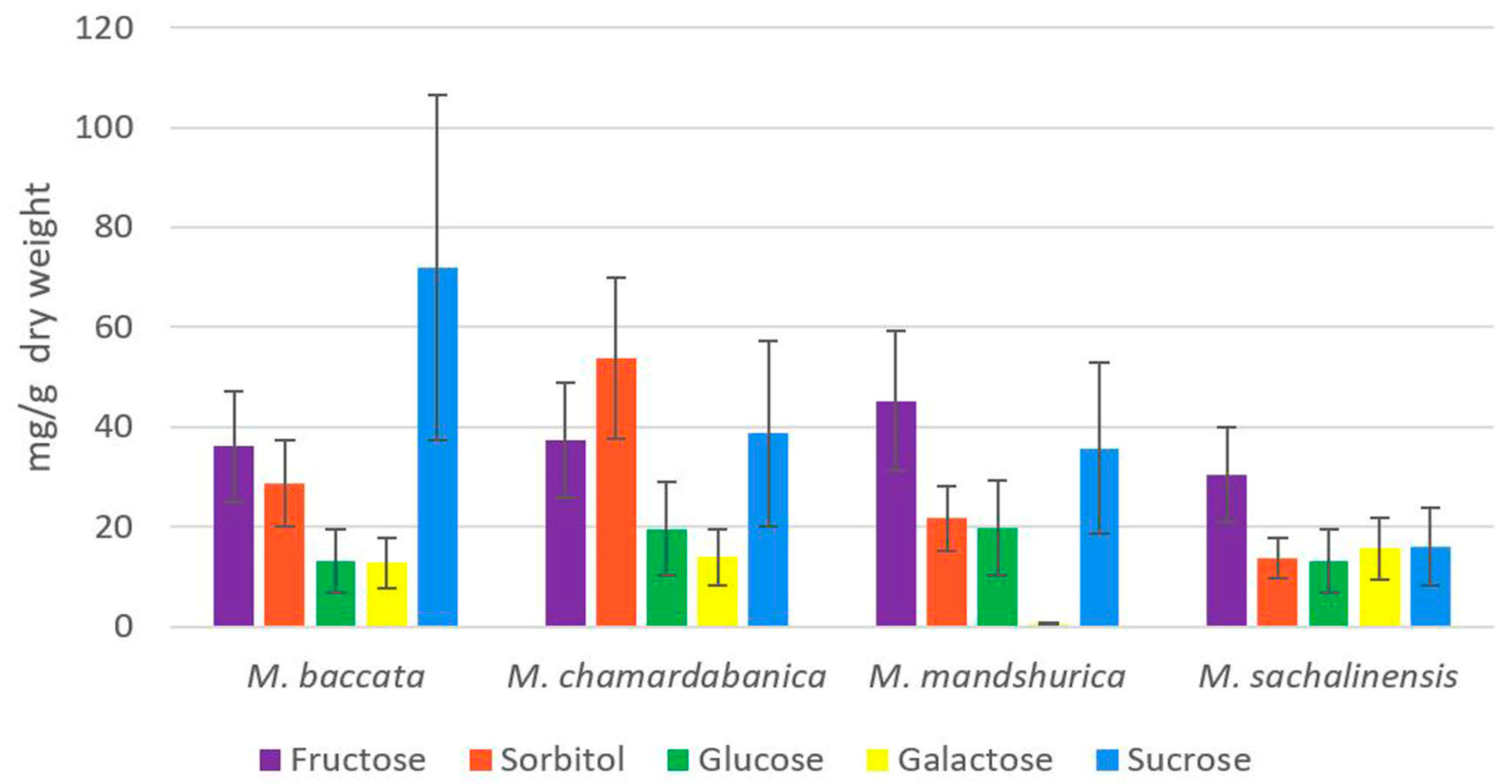
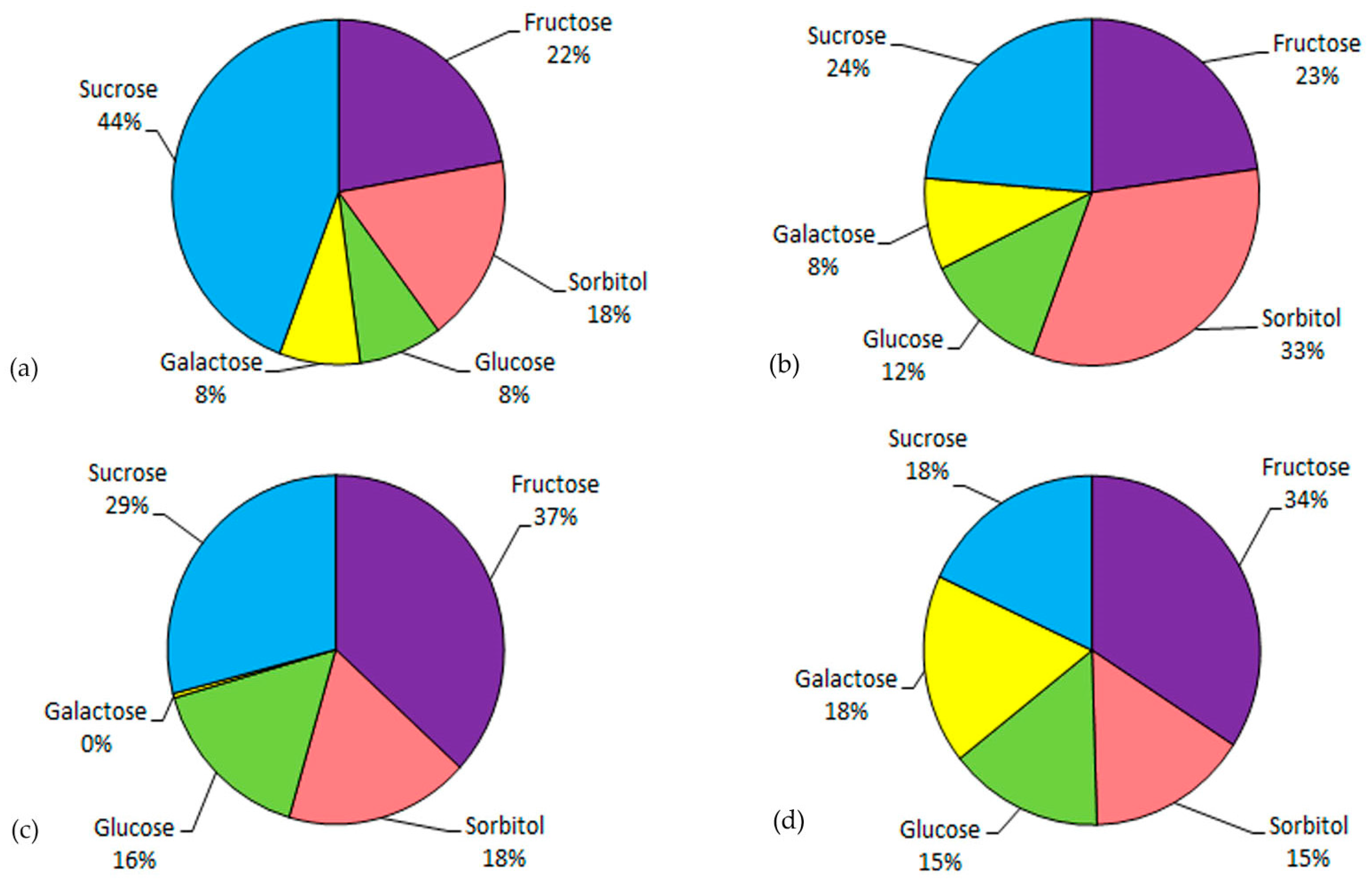
2.3. Content of the Main Soluble Sugars
2.4. Content of Pectin and Protopectin Fractions
2.5. Elemental Composition of Pectin and Protopectin Fractions
2.6. Principal Component Analysis
3. Discussion
3.1. Analysis of the Content of Main Soluble Sugars
3.2. Analysis of Biochemical Features of Pectin and Protopectin Fractions
4. Materials and Methods
4.1. Plant Material
4.2. Meteorological Data
4.3. Isolation and Analysis of Soluble Sugars
4.4. Isolation of Pectin Fractions
4.5. The Content of Pectic Substances
4.6. The Molecular Weight (MW)
4.7. Esterification of Pectin Chains
4.8. The Element Contents
4.9. Determination of Ascorbic Acid
4.10. Data Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reidel, E.J.; Rennie, E.; Amiard, V.; Cheng, L.; Turgeon, R. Phloem Loading Strategies in Three Plant Species that Transport Sugar Alcohols. Plant Physiol. 2009, 149, 1601–1608. [Google Scholar] [CrossRef] [PubMed]
- Shirataki, K.; Martinoia, E. Transporters in Fruit Vacuoles. Plant Biotech. 2007, 24, 127–133. [Google Scholar] [CrossRef]
- Reiter, W.-D.; Vanzin, G.F. Molecular Genetics of Nucleotide Sugar Interconversion Pathways in Plants. Plant Mol. Biol. 2001, 47, 95–113. [Google Scholar] [CrossRef]
- Roy, J.L.; Huss, B.; Creach, A.; Hawkins, S.; Neutelings, G. Glycosylation Is a Major Regulator of Phenylpropanoid Availability and Biological Activity in Plants. Front. Plant Sci. 2016, 7, 735. [Google Scholar] [CrossRef]
- Mellidou, I.; Kanellis, A.K. Genetic Control of Ascorbic Acid Biosynthesis and Recycling in Horticultural Crops. Front. Chem. 2017, 5, 50. [Google Scholar] [CrossRef] [PubMed]
- Ramon, M.; Rolland, F.; Sheen, J. Sugar Sensing and Signaling. TAB 2008, 6, e0117. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.-L. Signaling Role of Sucrose Metabolism in Development. Mol. Plant 2012, 5, 763–765. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Stewart, J.J.; Baker, C.R.; Adams, W.W. Optimization of Photosynthetic Productivity in Contrasting Environments by Regulons Controlling Plant Form and Function. Int. J. Mol. Sci. 2018, 19, 872. [Google Scholar] [CrossRef]
- Saddhe, A.A.; Manuka, R.; Penna, S. Plant Sugars: Homeostasis and Transport under Abiotic Stress in Plants. Physiol. Plant. 2021, 171, 739–755. [Google Scholar] [CrossRef]
- Eom, J.S.; Chen, L.Q.; Sosso, D.; Julius, B.T.; Lin, I.W.; Qu, X.Q.; Braun, D.M.; Frommer, W.B. SWEETs, Transporters for Intracellular and Intercellular Sugar Translocation. Curr. Opin. Plant Biol. 2015, 25, 53. [Google Scholar] [CrossRef]
- Yamaki, S. Metabolism and Accumulation of Sugars Translocated to Fruit and Their Regulation. Hortic. J. 2010, 79, 1–15. [Google Scholar] [CrossRef]
- Ma, B.; Chen, J.; Zheng, H.; Fang, T.; Ogutu, C.; Li, S.; Han, Y.; Wu, B. Comparative Assessment of Sugar and Malic Acid Composition in Cultivated and Wild Apples. Food Chem. 2015, 172, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, P.; Ma, F.; Dandekar, A.M.; Cheng, L. Sugar Metabolism and Accumulation in the Fruit of Transgenic Apple Trees with Decreased Sorbitol Synthesis. Hortic. Res. 2018, 5, 60. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Yaming, C.; Yang, Q.; Ogutu, C.O.; Liao, L.; Han, Y. Analysis of Sorbitol Content Variation in Wild and Cultivated Apples. J. Sci. Food Agric. 2019, 100, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Magwaza, L.; Opara, U. Analytical Methods for Determination of Sugars and Sweetness of Horticultural Products—A Review. Sci. Hortic. 2015, 184, 179–192. [Google Scholar] [CrossRef]
- Zhen, Q.; Fang, T.; Peng, Q.; Liao, L.; Zhao, L.; Owiti, A.; Han, Y. Developing Gene-Tagged Molecular Markers for Evaluation of Genetic Association of Apple SWEET Genes with Fruit Sugar Accumulation. Hortic. Res. 2018, 5, 14. [Google Scholar] [CrossRef]
- Rudikovskaya, E.G.; Dudareva, L.V.; Shishparenok, A.A.; Mitanova, N.B.; Petrova, I.G.; Rudikovskii, A.V. Phenolic Composition of Malus baccata Fruit. Chem. Nat. Compd. 2014, 4, 739–740. [Google Scholar] [CrossRef]
- Rudikovskaya, E.G.; Dudareva, L.V.; Shishparenok, A.A.; Rudikovskii, A.V. Peculiarities of Polyphenolic Profile of Fruits of Siberian Crab Apple and its Hybrids with Malus Domestica Borkh. Acta Physiol. Plant. 2015, 37, 238. [Google Scholar] [CrossRef]
- Darvill, A.G.; McNeil, M.; Albersheim, P. Structure of Plant Cell Walls. VIII. A New Pectic Polysaccharide. Plant Physiol. 1978, 62, 418–422. [Google Scholar] [CrossRef]
- Mohnen, D. Pectin Structure and Biosynthesis. Curr. Opin. Plant Biol. 2008, 11, 266–277. [Google Scholar] [CrossRef]
- Dranca, F.; Vargas, M.; Oroiana, M. Physicochemical Properties of Pectin from Malus domestica ‘Fălticeni’ Apple Pomace as Affected by non-Conventional Extraction Techniques. Food Hydrocoll. 2020, 100, 105383. [Google Scholar] [CrossRef]
- Kumari, A.; Dhaliwal, Y.S. A Study on Nutritional Composition and Value Addition of Crab Apple (Malus baccata). Am. J. Food Sci. Technol. 2017, 5, 19–22. [Google Scholar] [CrossRef]
- Petkova, N.; Ognyanov, M.; Inyutin, B.; Zhelev, P.; Denev, P. Phytochemical Composition and Antioxidant Activity of Malus baccata (L.) Borkh. Fruits. Food Sci. Appl. Biotechnol. 2020, 3, 47–55. [Google Scholar] [CrossRef]
- Vanina, L.S.; Vartapetyan, V.V. Wild Apple Trees (Collection of the Botanical Garden, Faculty of Biology. Lomonosov Moscow State University); KMK: Moscow, Russian, 2010. [Google Scholar]
- Zhang, L.Y.; Peng, Y.-B.; Pelleschi-Travier, S.; Fan, Y.; Lu, Y.-F.; Lu, Y.-M.; Gao, X.-P.; Shen, Y.-Y.; Delrot, S.; Zhang, D.-P. Evidence for Apoplasmic Phloem Unloading in Developing Apple Fruit. Plant Physiol. 2004, 135, 574–586. [Google Scholar] [CrossRef]
- Li, M.; Feng, F.; Cheng, L. Expression Patterns of Genes Involved in Sugar Metabolism and Accumulation During Apple Fruit Development. PLoS ONE 2012, 7, e33055. [Google Scholar] [CrossRef]
- Einhorn-Stoll, U.; Kastnera, H.; Urbischb, A.; Krohb, L.W.; Drusch, S. Thermal Degradation of Citrus Pectin in Low-Moisture Environment-Influence of Acidic and Alkaline Pre-treatment. Food Hydrocoll. 2019, 86, 104–115. [Google Scholar] [CrossRef]
- Willats, W.G.; Orfila, C.; Limberg, G.; Buchholt, H.C.; van Alebeek, G.J.; Voyage, A.G.; Marcus, S.E.; Christensen, T.M.; Mikkelsen, J.D.; Murray, B.S.; et al. Modulation of the Degree and Pattern of Methyl-Esterification of Pectic Homogalacturonan in Plant Cell Walls. Implications for Pectin Methyl Esterase Action, Matrix Properties and Cell Adhesion. J. Biol. Chem. 2001, 276, 19404–19413. [Google Scholar] [CrossRef]
- Wolf, S.; Mouille, G.; Pelloux, J. Homogalacturonan Methyl-Esterification and Plant Development. Mol. Plant 2009, 2, 85. [Google Scholar] [CrossRef]
- Schols, H.A.; Bakx, E.J.; Schipper, D.; Voragen, A.G.J. A Xylogalacturonan Subunit Present in the Modified Hairy Regions of Apple Pectin. Carbohydr. Res. 1995, 279, 265–279. [Google Scholar] [CrossRef]
- Schols, H.A.; Geraeds, C.C.J.M.; van Leeuwen, S.; Marjo, F.; Kormelink, F.J.M.; Voragen, A.G.J. Rhamnogalacturonase: Novel Enzyme that Degrades the Hairy Regions of Pectins. Carbohydr. Res. 1990, 206, 105–115. [Google Scholar] [CrossRef]
- Ridley, B.L.; O’Neill, M.A.; Mohnen, D. Pectins: Structure, Biosynthesis, and Oligogalacturonide-Related Signaling. Phytochemistry 2001, 57, 929–967. [Google Scholar] [PubMed]
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit Calcium: Transport and Physiology. Front. Plant Sci. 2016, 7, 569. [Google Scholar] [CrossRef]
- Manzoor, H.; Chiltz, A.; Madani, S.; Vatsa, P.; Schoefs, B.; Pugin, A.; Garcia-Brugger, A. Calcium Signatures and Signaling in Cytosol and Organelles of Tobacco Cells Induced by Plant Defense Elicitors. Cell Calcium 2012, 51, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Farmer, E.; Ryan, C. Interplant Communication: Airborne Methyl Jasmonate Induces Synthesis of Proteinase Inhibitors in Plant Leaves. Proc. Natl. Acad. Sci. USA 1990, 87, 7713–7716. [Google Scholar] [CrossRef]
- Moloshok, T.; Pearce, G.; Clarence, A.R. Oligouronide Signaling of Proteinase Inhibitor Genes in Plants: Structure-Activity Relationships of Di- and Trigalacturonic Acids and their Derivatives. Arch. Biochem. Biophys. 1992, 294, 731–734. [Google Scholar] [CrossRef] [PubMed]
- Legendre, L.; Rueter, S.; Heinstein, P.F.; Low, P.S. Characterization of the Oligogalacturonide-Induced Oxidative Burst in Cultured Soybean (Glycine max) Cells. Plant Physiol. 1993, 102, 233. [Google Scholar] [CrossRef]
- Binet, M.N.; Bourque, S.; Lebrun-Garcia, A.; Chiltz, A.; Pugin, A. Comparison of the Effects of Cryptogein and Oligogalacturonides on Tobacco Cells and Evidence of Different Forms of Desensitization Induced by these Elicitors. Plant Sci. 1998, 137, 33–41. [Google Scholar] [CrossRef]
- Low, P.S.; Merida, J.R. The Oxidative Burst in Plant Defense: Function and Signal Transduction. Physiol. Plant. 1996, 96, 533–542. [Google Scholar] [CrossRef]
- Lamb, C.J.; Dixon, R.A. The Oxidative Burst in Plant Disease Resistance. Annu. Rev. Plant Biol. 1997, 45, 251–275. [Google Scholar] [CrossRef]
- Bolwell, P.G. Role of Active Oxygen Species and NO in Plant Defense Responses. Curr. Opin. Plant Biol. 1999, 2, 287–294. [Google Scholar] [CrossRef]
- Côté, F.; Hahn, M.G. Oligosaccharins: Structure and Signal Transduction. Plant Mol. Biol. 1994, 12, 143–175. [Google Scholar]
- Hooda, P.S. (Ed.) Trace Elements in Soils; WileyBlackwell: Hoboken, NJ, USA, 2010. [Google Scholar] [CrossRef]
- Pinto, E.; Aguiar, A.A.R.M.; Ferreira, I.M.P.L.V.O. Influence of Soil Chemistry and Plant Physiology in the Phytoremediation of Cu, Mn, and Zn. Crit. Rev. Plant Sci. 2014, 33, 351–375. [Google Scholar] [CrossRef]
- Celus, M.; Kyomugasho, C.; Van Loey, A.M.; Grauwet, T.; Hendrickx, M.E. Influence of Pectin Structural Properties on Interactions with Divalent Cations and Its Associated Functionalities. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1576. [Google Scholar] [CrossRef] [PubMed]
- Minzanova, S.T.; Mironov, V.F.; Vyshtakalyuk, A.B.; Tsepaeva, O.V.; Mironova, L.G.; Mindubaev, A.Z.; Nizameev, I.R.; Kholin, K.V.; Milyukov, V.A. Complexation of Pectin with Macro- and Microelements. Antianemic Activity of Na, Fe and Na, Ca, Fe Complexes. Carbohydr. Polym. 2015, 134, 524–533. [Google Scholar] [CrossRef]
- Caffall, K.H.; Mohnen, D. The Structure, Function, and Biosynthesis of Plant Cell Wall Pectic Polysaccharides. Carbohydr. Res. 2009, 344, 1879. [Google Scholar] [CrossRef] [PubMed]
- Ezaki, N.; Kido, N.; Takahashi, K.; Katou, K. The Role of Wall Ca2+ in the Regulation of Wall Extensibility During the Acid-Induced Extension of Soybean Hypocotyl Cell Walls. Plant Cell Physiol. 2005, 46, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Chardonnet, C.O.; Sams, C.E.; Trigiano, R.N.; Conway, W.S. Variability of Three Isolates of Botrytis cinerea Affects the Inhibitory Effects of Calcium on this Fungus. Phytopathology. 2000, 90, 769–774. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, L.; Feng, Z.; Hao, W.; Mayo, K.H.; Zhou, Y.; Tai, G. Preparation of Individual Galactan Oligomers, their Prebiotic Effects, and Use in Estimating Galactan Chain Length in Pectin-Derived Polysaccharides. Carbohydr. Polym. 2018, 199, 526–533. [Google Scholar] [CrossRef]
- Glyad, V.M. Determination of Monosaccharides, Disaccharides, and Oligosaccharides in the Same Plant Sample by High-Performance Liquid Chromatography. Russ. J. Plant Physiol. 2002, 49, 277. [Google Scholar] [CrossRef]
- Ermakov, A.I.; Arasimovich, V.V.; Yarosh, N.P.; Peruanskii, U.V.; Lukovnikov, G.A.; Ikonnikova, M.I. Methods of Biochemical Research on Plants; Ermakov, A.I., Ed.; Agropromizdat: Leningrad, Russia, 1987; p. 429. (In Russian) [Google Scholar]
- Taylor, K.A.; Buchanan-Smith, J.G. A Calorimetric Method for the Quantitation of Uranic Acids and a Specific Assay for Galacturonic Acid. Anal. Biochem. 1992, 201, 190–196. [Google Scholar] [CrossRef]
- Babkin, V.A.; Neverova, N.A.; Medvedeva, E.N.; Fedorova, T.E.; Levchuk, A.A. Investigation of Physicochemical Properties of Arabinogalactan of Different Larch Species. Russ. J. Bioorg. Chem. 2016, 42, 707–711. [Google Scholar] [CrossRef]
- Rogošić, M.; Mencer, H.J.; Gomzi, Z. Polydispersity Index and Molecular Weight Distributions of Polymers. Eur. Polym. J. 1996, 32, 1337–1344. [Google Scholar] [CrossRef]
- Lara-Espinoza, C.; Carvajal-Millán, E.; Balandrán-Quintana, R.; López-Franco, Y.; Rascón-Chu, A. Pectin and Pectin-Based Composite Materials: Beyond Food Texture. Molecules 2018, 23, 942. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Diameter, cm | Length, cm | Weight, g | Ascorbic Acid, mg% | Water-Soluble Pectin, mg/g | Protopectin, mg/g |
|---|---|---|---|---|---|---|
| M. baccata | 1.35 ± 0.28 | 1.28 ± 0.36 | 1.22 ± 0.25 | 154 ± 26 | 4.81 ± 0.42 | 9.33 ± 1.11 |
| M. chamardabanica | 0.84 ± 0.04 | 1.10 ± 0.09 | 0.38 ± 0.04 | 129 ± 22 | 1.88 ± 0.31 | 5.07 ± 0.71 |
| M. mandshurica | 0.92 ± 0.10 | 1.42 ± 0.26 | 0.67 ± 0.08 | 117 ± 28 | 8.63 ± 0.86 | 9.18 ± 1.03 |
| M. sachalinensis | 1.21 ± 0.10 | 1.21 ± 0.08 | 0.93 ± 0.08 | 127 ± 28 | 6.02 ± 0.81 | 10.80 ± 1.06 |
| Water-Soluble Pectin | Protopectin | |||||||
|---|---|---|---|---|---|---|---|---|
| Species | Pectin Molecules, g/mol | D | LMWHPM #, g/mol | D | Pectin Molecules, g/mol | D | LMWHPM #, g/mol | D |
| M. baccata | 1.12 × 105 | 2.1 | 6.90 × 103 | 1.1 | 0.99 × 105 | 2.8 | 11.00 × 103 | 1.1 |
| M. chamardabanica | 4.81 × 105 | 1.5 | 7.00 × 103 | 1.1 | 4.32 × 105 | 8.2 | 23.00 × 103 | 1.2 |
| M. mandshurica | ND | ---- | 11.00 × 103 | 1.1 | 3.15 × 105 | 5.7 | 19.00 × 103 | 1.3 |
| M. sachalinensis | 0.11 × 105 | 6.5 | 6.80 × 103 | 1.1 | 1.03 × 105 | 4.0 | 7.30 × 103 | 1.1 |
| Species | Water-Soluble Pectin | Protopectin |
|---|---|---|
| M. baccata | 66 ± 2 | 70 ± 6 |
| M. chamardabanica | 65 ± 11 | 61 ± 11 |
| M. manshurica | 68 ± 6 | 72 ± 3 |
| M. sachalinensis | 60 ± 1 | 66 ± 6 |
| Vater-Soluble Pectin | Al | K | Mg | Ca | Cd | P | S | Cu | Fe | Zn |
|---|---|---|---|---|---|---|---|---|---|---|
| M. baccata | <0.15 | 79.52 | 8.17 | 21.01 | <0.02 | 63.31 | 50.03 | 0.12 | 0.23 | 0.15 |
| M. chamardabanica | <0.15 | 118.40 | 20.72 | 10.58 | <0.02 | 36.17 | 24.45 | 0.18 | 0.39 | 0.54 |
| M. manshurica | <0.15 | 24.88 | 4.21 | 10.34 | <0.02 | 14.91 | 22.54 | 0.04 | 0.10 | 0.11 |
| M.sachalinensis | <0.15 | 96.38 | 8.20 | 14.16 | <0.02 | 28.32 | 22.73 | ND | 0.19 | 0.12 |
| Protopectin | Al | K | Mg | Ca | Cd | P | S | Cu | Fe | Zn |
|---|---|---|---|---|---|---|---|---|---|---|
| M. baccata | <0.15 | 25.25 | 3.23 | 45.10 | <0.02 | 10.00 | 13.02 | 0.06 | 0.28 | 0.06 |
| M. chamardabanica | <0.15 | ND | 0.48 | 36.98 | <0.02 | 0.59 | 2.06 | 0.05 | 0.02 | 0.01 |
| M. manshurica | <0.15 | 24.54 | 5.77 | 42.70 | <0.02 | 6.20 | 614.92 | 0.04 | 0.33 | 0.13 |
| M.sachalinensis | <0.15 | 15.39 | 2.63 | 41.34 | <0.02 | 3.69 | 83.70 | 0.03 | 0.70 | 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stavitskaya, Z.; Dudareva, L.; Rudikovskii, A.; Garkava-Gustavsson, L.; Shabanova, E.; Levchuk, A.; Rudikovskaya, E. Evaluation of the Carbohydrate Composition of Crabapple Fruit Tissues Native to Northern Asia. Plants 2023, 12, 3472. https://doi.org/10.3390/plants12193472
Stavitskaya Z, Dudareva L, Rudikovskii A, Garkava-Gustavsson L, Shabanova E, Levchuk A, Rudikovskaya E. Evaluation of the Carbohydrate Composition of Crabapple Fruit Tissues Native to Northern Asia. Plants. 2023; 12(19):3472. https://doi.org/10.3390/plants12193472
Chicago/Turabian StyleStavitskaya, Zlata, Lyubov Dudareva, Alexander Rudikovskii, Larisa Garkava-Gustavsson, Elena Shabanova, Alexey Levchuk, and Elena Rudikovskaya. 2023. "Evaluation of the Carbohydrate Composition of Crabapple Fruit Tissues Native to Northern Asia" Plants 12, no. 19: 3472. https://doi.org/10.3390/plants12193472
APA StyleStavitskaya, Z., Dudareva, L., Rudikovskii, A., Garkava-Gustavsson, L., Shabanova, E., Levchuk, A., & Rudikovskaya, E. (2023). Evaluation of the Carbohydrate Composition of Crabapple Fruit Tissues Native to Northern Asia. Plants, 12(19), 3472. https://doi.org/10.3390/plants12193472