
Early Antioxidative Response to Desiccant-Stimulated Drought Stress in Field-Grown Traditional Wheat Varieties

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
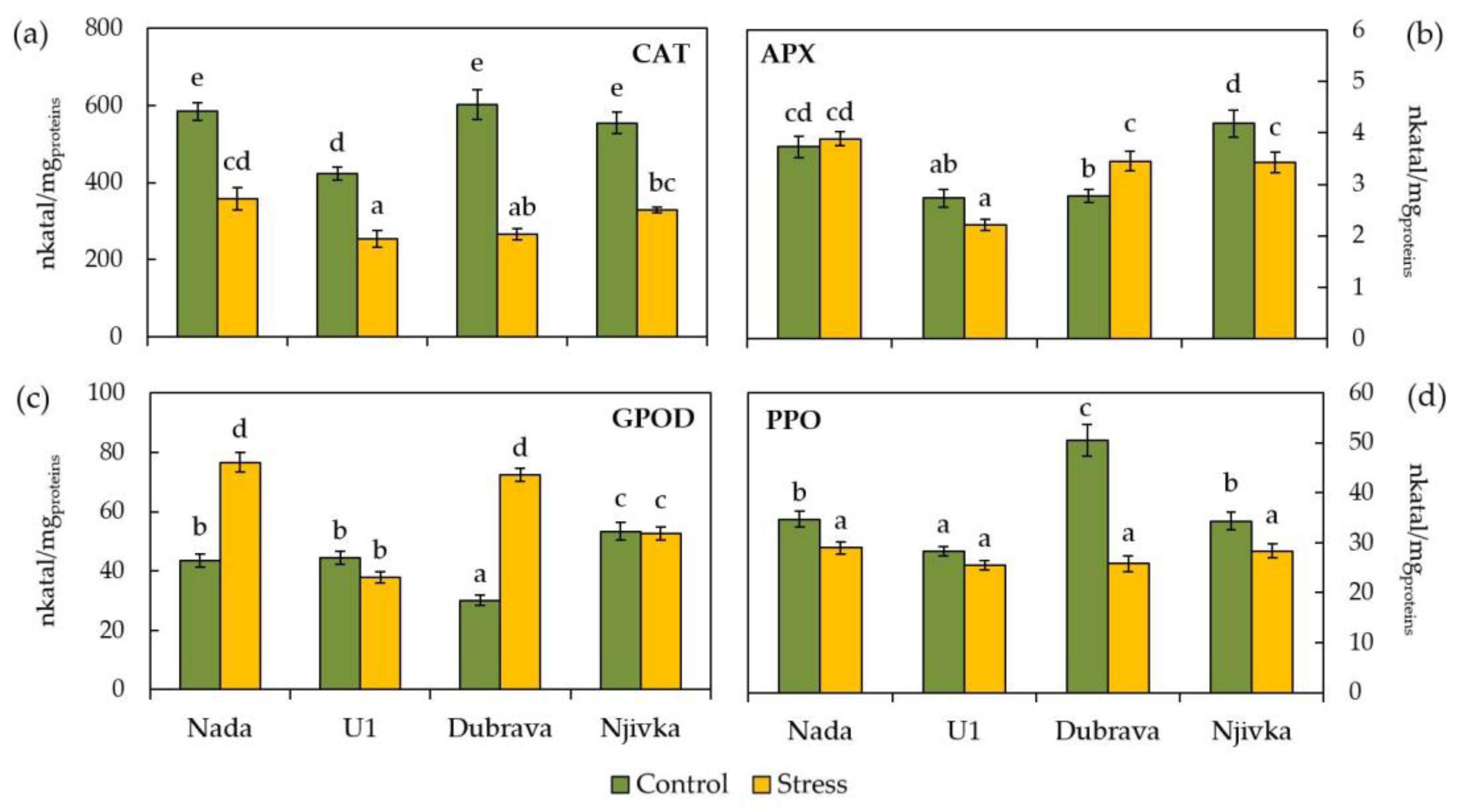
2. Results
2.1. Determination of Stress Occurrence and Severity
2.2. Hydrogen Peroxide, Lipid Peroxidation and Protein Content
2.3. Enzymatic Activity
2.4. Total Phenolics and Phenolic Acids Content
2.5. Photosynthetic Pigments
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Conditions
4.2. Water Content
4.3. Proline Content
4.4. Lipid Peroxidation and Hydrogen Peroxide
4.4.1. Lipid Peroxidation
4.4.2. Hydrogen Peroxide
4.5. Antioxidative Enzymes and Polyphenol Oxidase Activity
4.5.1. Soluble Protein Concentration
4.5.2. Catalase Activity
4.5.3. Ascorbate Peroxidase Activity
4.5.4. Guaiacol Peroxidase Activity
4.5.5. Polyphenol Oxidase Activity
4.6. Phenolic Acids Content
4.6.1. Extraction of Phenolic Acids for HPLC Analysis
4.6.2. HPLC Analysis of Phenolic Acids
4.7. Photosynthetic Pigments
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shewry, P.R.; Hey, S.J. The Contribution of Wheat to Human Diet and Health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef]
- Arora, N.K. Impact of Climate Change on Agriculture Production and Its Sustainable Solutions. Environ. Sustain. 2019, 2, 95–96. [Google Scholar] [CrossRef] [Green Version]
- Marinović, I.; Kalin, K.C.; Güttler, I.; Pasarić, Z. Dry Spells in Croatia: Observed Climate Change and Climate Projections. Atmosphere 2021, 12, 652. [Google Scholar] [CrossRef]
- Barić, M.; Kereša, S.; Jerčić, I.H.; Havrda, S.; Gelenčir, D. Evaluation and Characterization of Croatian Winter Wheat Genotypes (T. aestivum L.) for Drought Tolerance. Cereal Res. Commun. 2008, 36, 1031–1034. [Google Scholar]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The Role of the Plant Antioxidant System in Drought Tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Carvalho, M.H.C. Drought Stress and Reactive Oxygen Species: Production, Scavenging and Signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef] [Green Version]
- Blum, A. Drought Resistance and Its Improvement. In Plant Breeding for Water-Limited Environments; Springer: New York, NY, USA, 2011; pp. 53–152. ISBN 9781441974914. [Google Scholar]
- Mwadzingeni, L.; Figlan, S.; Shimelis, H.; Mondal, S.; Tsilo, T.J. Genetic Resources and Breeding Methodologies for Improving Drought Tolerance in Wheat. J. Crop Improv. 2017, 31, 648–672. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Van Breusegem, F. ROS Signaling: The New Wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Alché, J.D.D. A Concise Appraisal of Lipid Oxidation and Lipoxidation in Higher Plants. Redox Biol. 2019, 23, 101136. [Google Scholar] [CrossRef] [PubMed]
- Rawat, N.; Singla-Pareek, S.L.; Pareek, A. Membrane Dynamics during Individual and Combined Abiotic Stresses in Plants and Tools to Study the Same. Physiol. Plant. 2021, 171, 653–676. [Google Scholar] [CrossRef] [PubMed]
- Simova-Stoilova, L.; Vaseva, I.; Grigorova, B.; Demirevska, K.; Feller, U. Proteolytic Activity and Cysteine Protease Expression in Wheat Leaves under Severe Soil Drought and Recovery. Plant Physiol. Biochem. 2010, 48, 200–206. [Google Scholar] [CrossRef]
- Upadhyay, D.; Budhlakoti, N.; Singh, A.K.; Bansal, R.; Kumari, J.; Chaudhary, N.; Padaria, J.C.; Sareen, S.; Kumar, S. Drought Tolerance in Triticum aestivum L. Genotypes Associated with Enhanced Antioxidative Protection and Declined Lipid Peroxidation. 3 Biotech 2020, 10, 281. [Google Scholar] [CrossRef] [PubMed]
- Kiani, R.; Arzani, A.; Mirmohammady Maibody, S.A.M. Polyphenols, Flavonoids, and Antioxidant Activity Involved in Salt Tolerance in Wheat, Aegilops Cylindrica and Their Amphidiploids. Front. Plant Sci. 2021, 12, 646221. [Google Scholar] [CrossRef] [PubMed]
- Naderi, S.; Fakheri, B.A.; Maali-Amiri, R.; Mahdinezhad, N. Tolerance Responses in Wheat Landrace Bolani Are Related to Enhanced Metabolic Adjustments under Drought Stress. Plant Physiol. Biochem. 2020, 150, 244–253. [Google Scholar] [CrossRef]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef]
- Andrés, C.M.C.; de la Lastra, J.M.P.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Chemistry of Hydrogen Peroxide Formation and Elimination in Mammalian Cells, and Its Role in Various Pathologies. Stresses 2022, 2, 256–274. [Google Scholar] [CrossRef]
- Quan, L.J.; Zhang, B.; Shi, W.W.; Li, H.Y. Hydrogen Peroxide in Plants: A Versatile Molecule of the Reactive Oxygen Species Network. J. Integr. Plant Biol. 2008, 50, 2–18. [Google Scholar] [CrossRef]
- Herbette, S.; Lenne, C.; Leblanc, N.; Julien, J.L.; Drevet, J.R.; Roeckel-Drevet, P. Two GPX-like Proteins from Lycopersicon Esculentum and Helianthus Annuus Are Antioxidant Enzymes with Phospholipid Hydroperoxide Glutathione Peroxidase and Thioredoxin Peroxidase Activities. Eur. J. Biochem. 2002, 269, 2414–2420. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Asada, K. The Water-Water Cycle in Chloroplasts: Scavenging of Active Oxygens and Dissipation of Excess Photons. Annu. Rev. Plant Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef] [PubMed]
- Hernández, I.; Cela, J.; Alegre, L.; Munné-Bosch, S. Antioxidant Defenses against Drought Stress. In Plant Responses to Drought Stress; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 231–258. ISBN 9783642326530. [Google Scholar]
- Kumar, D.; Al Hassan, M.; Naranjo, M.A.; Agrawal, V.; Boscaiu, M.; Vicente, O. Effects of Salinity and Drought on Growth, Ionic Relations, Compatible Solutes and Activation of Antioxidant Systems in Oleander (Nerium oleander L.). PLoS ONE 2017, 12, e0185017. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, U.; Pradhan, B. Drought Stress-Induced Oxidative Stress and Antioxidative Responses in Four Wheat (Triticum aestivum L.) Varieties. Arch. Agron. Soil Sci. 2012, 58, 617–630. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Li, Y.; Guo, T. Expression of Flavonoid Biosynthesis Genes and Accumulation of Flavonoid in Wheat Leaves in Response to Drought Stress. Plant Physiol. Biochem. 2014, 80, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Boeckx, T.; Winters, A.L.; Webb, K.J.; Kingston-Smith, A.H. Polyphenol Oxidase in Leaves: Is There Any Significance to the Chloroplastic Localization? J. Exp. Bot. 2015, 66, 3571–3579. [Google Scholar] [CrossRef] [Green Version]
- Sofo, A.; Dichio, B.; Xiloyannis, C.; Masia, A. Antioxidant Defences in Olive Trees during Drought Stress: Changes in Activity of Some Antioxidant Enzymes. Funct. Plant Biol. 2005, 32, 45–53. [Google Scholar] [CrossRef]
- Thipyapong, P.; Melkonian, J.; Wolfe, D.W.; Steffens, J.C. Suppression of Polyphenol Oxidases Increases Stress Tolerance in Tomato. Plant Sci. 2004, 167, 693–703. [Google Scholar] [CrossRef]
- Lee, B.R.; Kim, K.Y.; Jung, W.J.; Avice, J.C.; Ourry, A.; Kim, T.H. Peroxidases and Lignification in Relation to the Intensity of Water-Deficit Stress in White Clover (Trifolium repens L.). J. Exp. Bot. 2007, 58, 1271–1279. [Google Scholar] [CrossRef] [Green Version]
- Blum, A. Osmotic Adjustment Is a Prime Drought Stress Adaptive Engine in Support of Plant Production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of Glycine Betaine and Proline in Improving Plant Abiotic Stress Resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Trovato, M.; Mattioli, R.; Costantino, P. Multiple Roles of Proline in Plant Stress Tolerance and Development. Rend. Lincei 2008, 19, 325–346. [Google Scholar] [CrossRef]
- Mwadzingeni, L.; Shimelis, H.; Tesfay, S.; Tsilo, T.J. Screening of Bread Wheat Genotypes for Drought Tolerance Using Phenotypic and Proline Analyses. Front. Plant Sci. 2016, 7, 1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, A.; Tuberosa, R. Dehydration Survival of Crop Plants and Its Measurement. J. Exp. Bot. 2018, 69, 975–981. [Google Scholar] [CrossRef]
- Dodig, D.; Zorić, M.; Jović, M.; Kandić, V.; Stanisavljević, R.; Šurlan-Momirović, G. Wheat Seedlings Growth Response to Water Deficiency and How It Correlates with Adult Plant Tolerance to Drought. J. Agric. Sci. 2015, 153, 466–480. [Google Scholar] [CrossRef]
- Breinl, K.; Di Baldassarre, G.; Mazzoleni, M.; Lun, D.; Vico, G. Extreme Dry and Wet Spells Face Changes in Their Duration and Timing. Environ. Res. Lett. 2020, 15, 074040. [Google Scholar] [CrossRef]
- Zhang, C.J.; Chen, G.X.; Gao, X.X.; Chu, C.J. Photosynthetic Decline in Flag Leaves of Two Field-Grown Spring Wheat Cultivars with Different Senescence Properties. S. Afr. J. Bot. 2006, 72, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R.A.; Maurer, R. Drought Resistance in Spring Wheat Cultivars. I. Grain Yield Responses. Aust. J. Agric. Res. 1978, 29, 897–912. [Google Scholar] [CrossRef]
- Ozturk, A.; Aydin, F. Effect of Water Stress at Various Growth Stages on Some Quality Characteristics of Winter Wheat. J. Agron. Crop Sci. 2004, 190, 93–99. [Google Scholar] [CrossRef]
- Allahverdiyev, T.I. Effect of Drought Stress on Some Physiological Traits of Durum (Triticum durum Desf.) and Bread (Triticum aestivum L.) Wheat Genotypes. J. Stress Physiol. Biochem. 2015, 11, 29–38. [Google Scholar]
- Schonfeld, M.A.; Johnson, R.C.; Carver, B.F.; Mornhinweg, D.W. Water Relations in Winter Wheat as Drought Resistance Indicators. Crop Sci. 1988, 28, 526–531. [Google Scholar] [CrossRef]
- Chowdhury, M.K.; Hasan, M.A.; Bahadur, M.M.; Islam, M.R.; Hakim, M.A.; Iqbal, M.A.; Javed, T.; Raza, A.; Shabbir, R.; Sorour, S.; et al. Evaluation of Drought Tolerance of Some Wheat (Triticum aestivum L.) Genotypes through Phenology, Growth, and Physiological Indices. Agronomy 2021, 11, 1792. [Google Scholar] [CrossRef]
- Baloch, M.J.; Dunwell, J.; Khan, N.U.; Jatoi, W.A.; Khakhwani, A.A.; Vessar, N.F.; Gul, S. Morpho-Physiological Characterization of Spring Wheat Genotypes under Drought Stress. Int. J. Agric. Biol. 2013, 15, 945–950. [Google Scholar]
- Larbi, A.; Mekliche, A.; Agronomique, I.N.; De Phytotechnie, D. Relative Water Content (RWC) and Leaf Senescence as Screening Tools for Drought Tolerance in Wheat. Growth 2004, 196, 193–196. [Google Scholar]
- Maghsoudi, K.; Emam, Y.; Niazi, A.; Pessarakli, M.; Arvin, M.J. P5CS Expression Level and Proline Accumulation in the Sensitive and Tolerant Wheat Cultivars under Control and Drought Stress Conditions in the Presence/Absence of Silicon and Salicylic Acid. J. Plant Interact. 2018, 13, 461–471. [Google Scholar] [CrossRef] [Green Version]
- Bowne, J.B.; Erwin, T.A.; Juttner, J.; Schnurbusch, T.; Langridge, P.; Bacic, A.; Roessner, U. Drought Responses of Leaf Tissues from Wheat Cultivars of Differing Drought Tolerance at the Metabolite Level. Mol. Plant 2012, 5, 418–429. [Google Scholar] [CrossRef] [Green Version]
- Sultan, M.A.R.F.; Hui, L.; Yang, L.J.; Xian, Z.H. Assessment of Drought Tolerance of Some Triticum L. Species through Physiological Indices. Czech J. Genet. Plant Breed. 2012, 48, 178–184. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, I.M.; Dai, H.; Zheng, W.; Cao, F.; Zhang, G.; Sun, D.; Wu, F. Genotypic Differences in Physiological Characteristics in the Tolerance to Drought and Salinity Combined Stress between Tibetan Wild and Cultivated Barley. Plant Physiol. Biochem. 2013, 63, 49–60. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic Stress and Reactive Oxygen Species: Generation, Signaling, and Defense Mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef]
- Abid, M.; Ali, S.; Qi, L.K.; Zahoor, R.; Tian, Z.; Jiang, D.; Snider, J.L.; Dai, T. Physiological and Biochemical Changes during Drought and Recovery Periods at Tillering and Jointing Stages in Wheat (Triticum aestivum L.). Sci. Rep. 2018, 8, 4615. [Google Scholar] [CrossRef] [Green Version]
- Hameed, A.; Goher, M.; Iqbal, N. Drought Induced Programmed Cell Death and Associated Changes in Antioxidants, Proteases, and Lipid Peroxidation in Wheat Leaves. Biol. Plant. 2013, 57, 370–374. [Google Scholar] [CrossRef]
- Peršić, V.; Ament, A.; Dunić, J.A.; Drezner, G.; Cesar, V. PEG-Induced Physiological Drought for Screening Winter Wheat Genotypes Sensitivity—Integrated Biochemical and Chlorophyll a Fl Uorescence Analysis. Front. Plant Sci. 2022, 13, 987702. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative Stress, Antioxidants and Stress Tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Arora, A.; Sairam, R.K.; Srivastava, G.C. Oxidative Stress and Antioxidative System in Plants. Curr. Sci. 2002, 82, 1227–1238. [Google Scholar]
- Vuković, R.; Čamagajevac, I.Š.; Vuković, A.; Šunić, K.; Begović, L.; Mlinarić, S.; Sekulić, R.; Sabo, N.; Španić, V. Physiological, Biochemical and Molecular Response of Different Winter Wheat Varieties under Drought Stress at Germination and Seedling Growth Stage. Antioxidants 2022, 11, 693. [Google Scholar] [CrossRef]
- Pour-Benab, S.M.; Fabriki-Ourang, S.; Mehrabi, A.A. Expression of Dehydrin and Antioxidant Genes and Enzymatic Antioxidant Defense under Drought Stress in Wild Relatives of Wheat. Biotechnol. Biotechnol. Equip. 2019, 33, 1063–1073. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, M.L. Beyond Brown: Polyphenol Oxidases as Enzymes of Plant Specialized Metabolism. Front. Plant Sci. 2015, 5, 783. [Google Scholar] [CrossRef] [Green Version]
- Gandía-Herrero, F.; García-Carmona, F. Biosynthesis of Betalains: Yellow and Violet Plant Pigments. Trends Plant Sci. 2013, 18, 334–343. [Google Scholar] [CrossRef]
- Boeckx, T.; Webster, R.; Winters, A.L.; Webb, K.J.; Gay, A.; Kingston-Smith, A.H. Polyphenol Oxidase-Mediated Protection against Oxidative Stress Is Not Associated with Enhanced Photosynthetic Efficiency. Ann. Bot. 2015, 116, 529–540. [Google Scholar] [CrossRef] [Green Version]
- Boeckx, T.; Winters, A.; Webb, K.J.; Kingston-Smith, A.H. Detection of Potential Chloroplastic Substrates for Polyphenol Oxidase Suggests a Role in Undamaged Leaves. Front. Plant Sci. 2017, 8, 237. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.; Ahmad, S.D.; Sabir, S.M.; Awan, S.I.; Hameed, A.; Abbas, S.R.; Shehzad, M.; Khan, M.F.; Shafique, S.; Ahmad, Z. Detection of Saline Tolerant Wheat Cultivars (Triticum aestivum L.) Using Lipid Peroxidation, Antioxidant Defense System, Glycine-Betaine and Proline Contents. J. Anim. Plant Sci. 2013, 23, 1742–1748. [Google Scholar]
- Moharramnejad, S.; Sofalian, O.; Valizadeh, M.; Asgari, A.; Shiri, M. Proline, Glycine Betaine, Total Phenolics and Pigment Contents in Response to Osmotic Stress in Maize Seedlings. J. Biosci. Biotechnol. 2015, 4, 313–319. [Google Scholar]
- Ashraf, M.A.; Ashraf, M.; Ali, Q. Response of Two Genetically Diverse Wheat Cultivars to Salt Stress at Different Growth Stages: Leaf Lipid Peroxidation and Phenolic Contents. Pak. J. Bot. 2010, 42, 559–565. [Google Scholar]
- Prado, N.B.D.; de Abreu, C.B.; Pinho, C.S.; Junior, M.M.d.N.; Silva, M.D.; Espino, M.; Silva, M.F.; Dias, F.D.S. Application of Multivariate Analysis to Assess Stress by Cd, Pb and Al in Basil (Ocimum basilicum L.) Using Caffeic Acid, Rosmarinic Acid, Total Phenolics, Total Flavonoids and Total Dry Mass in Response. Food Chem. 2022, 367, 130682. [Google Scholar] [CrossRef]
- Zafar-Ul-Hye, M.; Akbar, M.N.; Iftikhar, Y.; Abbas, M.; Zahid, A.; Fahad, S.; Datta, R.; Ali, M.; Elgorban, A.M.; Ansari, M.J.; et al. Rhizobacteria Inoculation and Caffeic Acid Alleviated Drought Stress in Lentil Plants. Sustainability 2021, 13, 9603. [Google Scholar] [CrossRef]
- Mehmood, H.; Abbasi, G.H.; Jamil, M.; Malik, Z.; Ali, M.; Iqbal, R. Assessing the Potential of Exogenous Caffeic Acid Application in Boosting Wheat (Triticum aestivum L.) Crop Productivity under Salt Stress. PLoS ONE 2021, 16, e0259222. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Xin, Z.; Yang, T.; Ma, X.; Zhang, Y.; Wang, Z.; Ren, Y.; Lin, T. Metabolomics Response for Drought Stress Tolerance. Plants 2020, 9, 520. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Singh Sidhu, G.P.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic Response of Plants Under Different Abiotic Stresses: A Review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Agathokleous, E.; Feng, Z.Z.; Peñuelas, J. Chlorophyll Hormesis: Are Chlorophylls Major Components of Stress Biology in Higher Plants? Sci. Total Environ. 2020, 726, 138637. [Google Scholar] [CrossRef]
- Vuletić, M.V.; Mihaljević, I.; Tomaš, V.; Horvat, D.; Zdunić, Z.; Vuković, D. Physiological Response to Short-Term Heat Stress in the Leaves of Traditional and Modern Plum (Prunus domestica L.) Cultivars. Horticulturae 2022, 8, 72. [Google Scholar] [CrossRef]
- Gür, A.; Demirel, U.; Özden, M.; Kahraman, A.; Çopur, O. Diurnal Gradual Heat Stress Affects Antioxidant Enzymes, Proline Accumulation and Some Physiological Components in Cotton (Gossypium hirsutum L.). Afr. J. Biotechnol. 2010, 9, 1008–1015. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under Stressful Environments: An Overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought Stress in Plants: A Review on Morphological Characteristics and Pigments Composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Jain, M.; Tiwary, S.; Gadre, R. Sorbitol-Induced Changes in Various Growth and Biochemical Parameters in Maize. Plant Soil Environ. 2010, 56, 263–267. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Gu, T.; Khan, I.; Zada, A. Research Progress in the Interconversion, Turnover and Degradation of Chlorophyll. Cells 2021, 10, 3134. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.Y.; Azmi, A.R.; Khan, A.H.; Ala, S. Effect of Water Stress on Total Phenol, Peroxidase Activity and Chlorophyll Contents in Wheat (Triticum aestivum L.). Acta Physiol. Plant. 1994, 16, 185–191. [Google Scholar]
- Krieger-Liszkay, A.; Fufezan, C.; Trebst, A. Singlet Oxygen Production in Photosystem II and Related Protection Mechanism. Photosynth. Res. 2008, 98, 551–564. [Google Scholar] [CrossRef]
- Woodrow, P.; Ciarmiello, L.F.; Annunziata, M.G.; Pacifico, S.; Iannuzzi, F.; Mirto, A.; D’Amelia, L.; Dell’Aversana, E.; Piccolella, S.; Fuggi, A.; et al. Durum Wheat Seedling Responses to Simultaneous High Light and Salinity Involve a Fine Reconfiguration of Amino Acids and Carbohydrate Metabolism. Physiol. Plant. 2017, 159, 290–312. [Google Scholar] [CrossRef]
- Verma, S.; Dubey, R.S. Lead Toxicity Induces Lipid Peroxidation and Alters the Activities of Antioxidant Enzymes in Growing Rice Plants. Plant Sci. 2003, 164, 645–655. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative Stress and Some Antioxidant Systems in Acid Rain-Treated Bean Plants: Protective Role of Exogenous Polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantitites of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase In Vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; pp. 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Siegel, B.Z.; Galston, W. The Isoperoxidases of Pisum sativum. Plant Physiol. 1967, 42, 221–226. [Google Scholar] [CrossRef] [Green Version]
- Raymond, J.; Rakariyatham, N.; Azanza, J.L. Purification and Some Properties of Polyphenoloxidase from Sunflower Seeds. Phytochemistry 1993, 34, 927–931. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Pedigree | Year of Release |
|---|---|---|
| Nada | OSJECKA-20/OSK-4.216-2-76 | 1984 |
| U1 | CARLOTTA STRAMPELLI/MARQUIS | 1936 |
| Dubrava | U1/PILOT//LIBERO | 1968 |
| Njivka | SLAVONKA/OSK-5-132-2-74 | 1987 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dvojković, K.; Plavšin, I.; Novoselović, D.; Šimić, G.; Lalić, A.; Čupić, T.; Horvat, D.; Viljevac Vuletić, M. Early Antioxidative Response to Desiccant-Stimulated Drought Stress in Field-Grown Traditional Wheat Varieties. Plants 2023, 12, 249. https://doi.org/10.3390/plants12020249
Dvojković K, Plavšin I, Novoselović D, Šimić G, Lalić A, Čupić T, Horvat D, Viljevac Vuletić M. Early Antioxidative Response to Desiccant-Stimulated Drought Stress in Field-Grown Traditional Wheat Varieties. Plants. 2023; 12(2):249. https://doi.org/10.3390/plants12020249
Chicago/Turabian StyleDvojković, Krešimir, Ivana Plavšin, Dario Novoselović, Gordana Šimić, Alojzije Lalić, Tihomir Čupić, Daniela Horvat, and Marija Viljevac Vuletić. 2023. "Early Antioxidative Response to Desiccant-Stimulated Drought Stress in Field-Grown Traditional Wheat Varieties" Plants 12, no. 2: 249. https://doi.org/10.3390/plants12020249
APA StyleDvojković, K., Plavšin, I., Novoselović, D., Šimić, G., Lalić, A., Čupić, T., Horvat, D., & Viljevac Vuletić, M. (2023). Early Antioxidative Response to Desiccant-Stimulated Drought Stress in Field-Grown Traditional Wheat Varieties. Plants, 12(2), 249. https://doi.org/10.3390/plants12020249