

Historical Assembly of Andean Tree Communities

 , , , , , , , , ,
, , , , , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
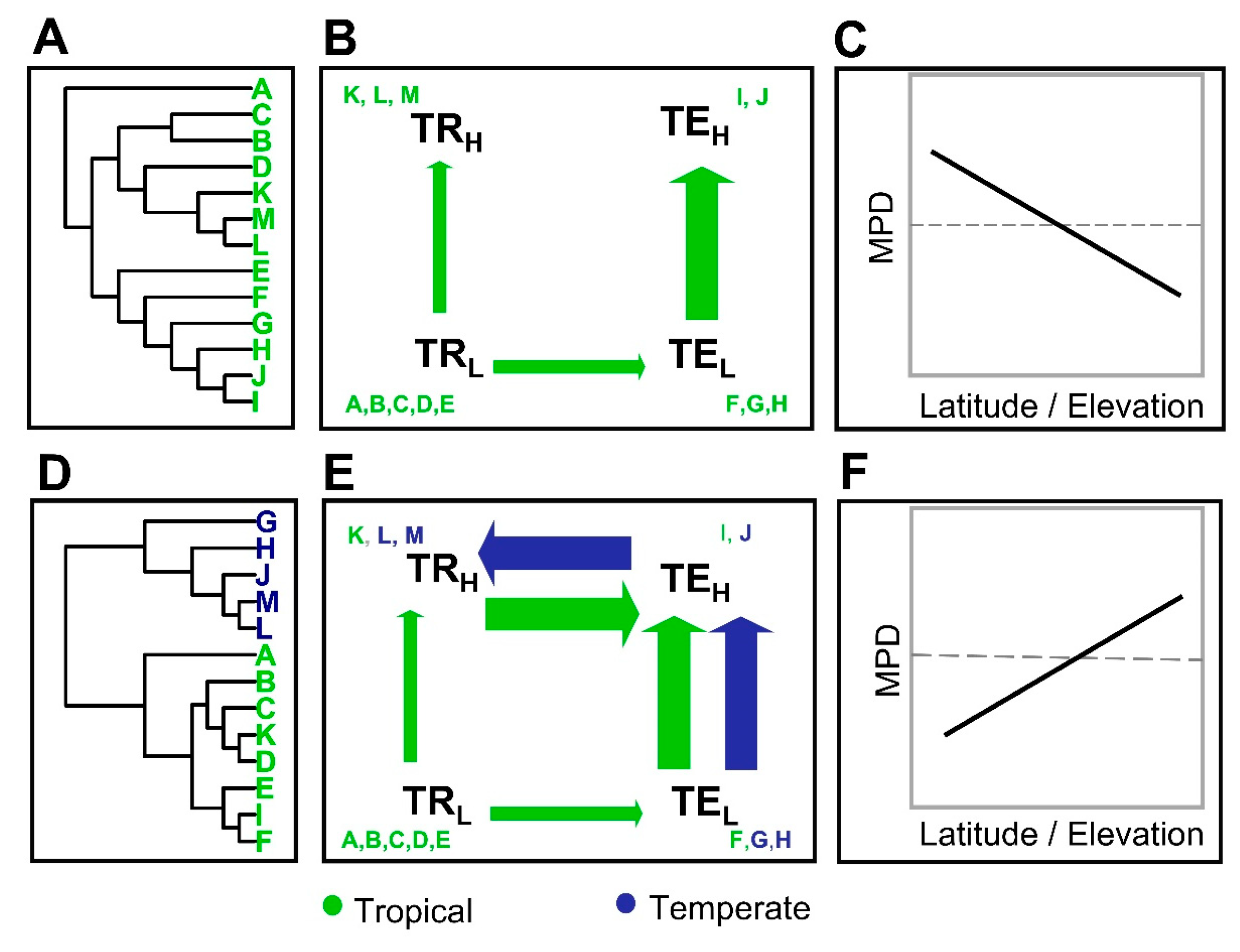
2.1. Validation of Predictions through Simulations of Biogeographic Diversification
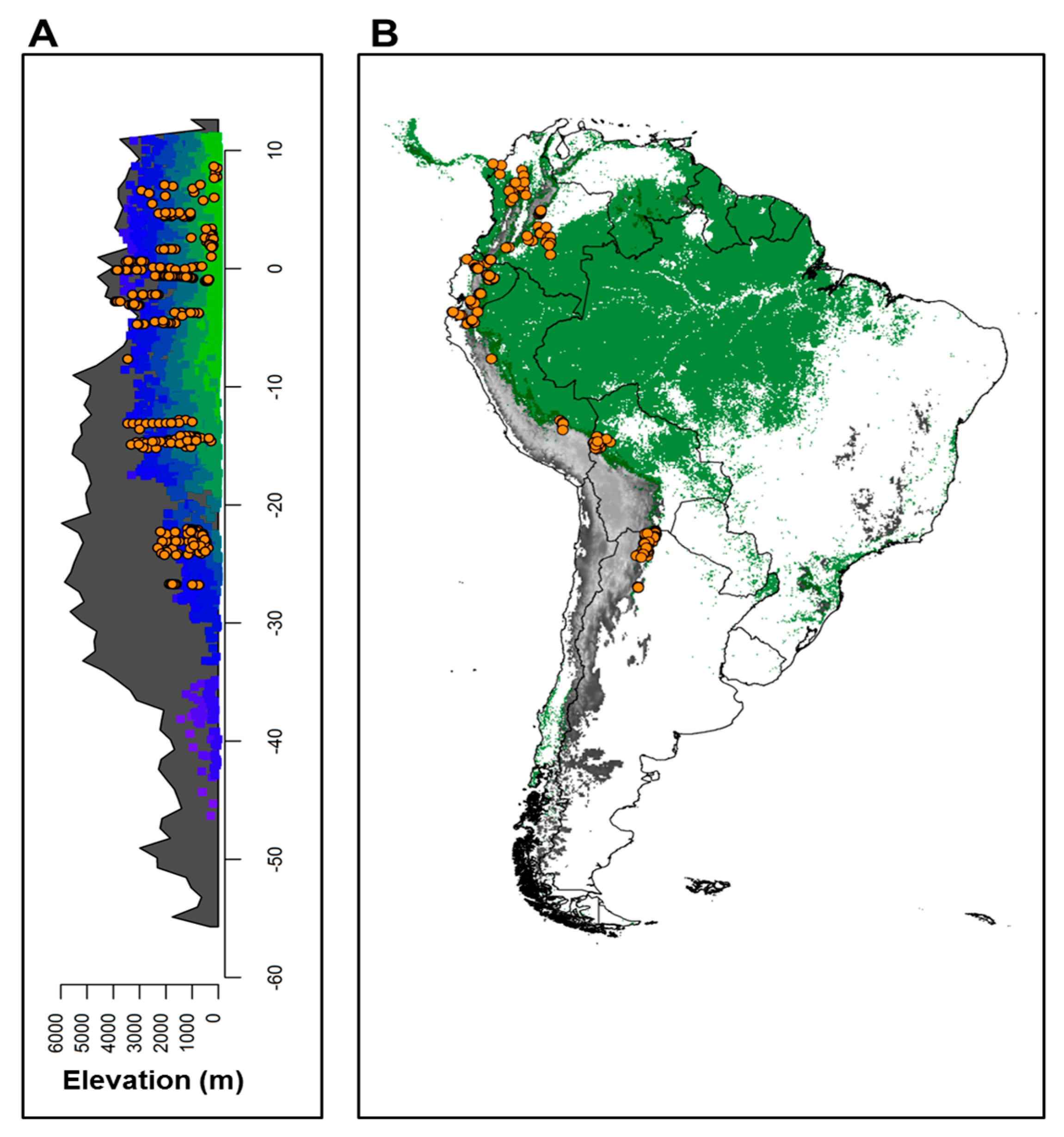
2.2. Forest Communities across the Andes
2.3. Measuring Local Phylogenetic Diversity
2.4. Testing Predictions with Empirical Data
3. Results
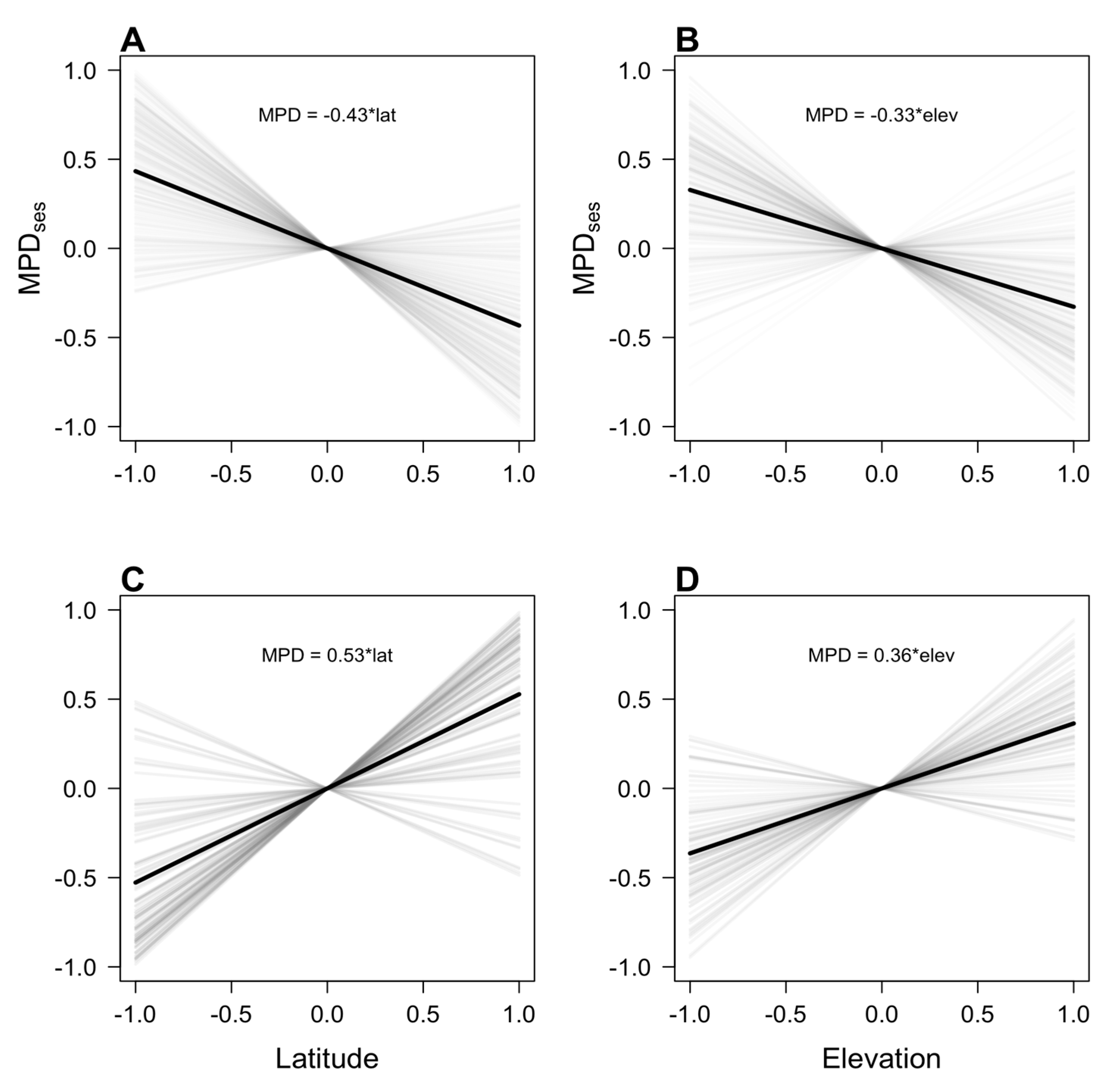
3.1. Simulations of the Tropical Niche Conservatism and Multiple Zones of Origin Hypotheses
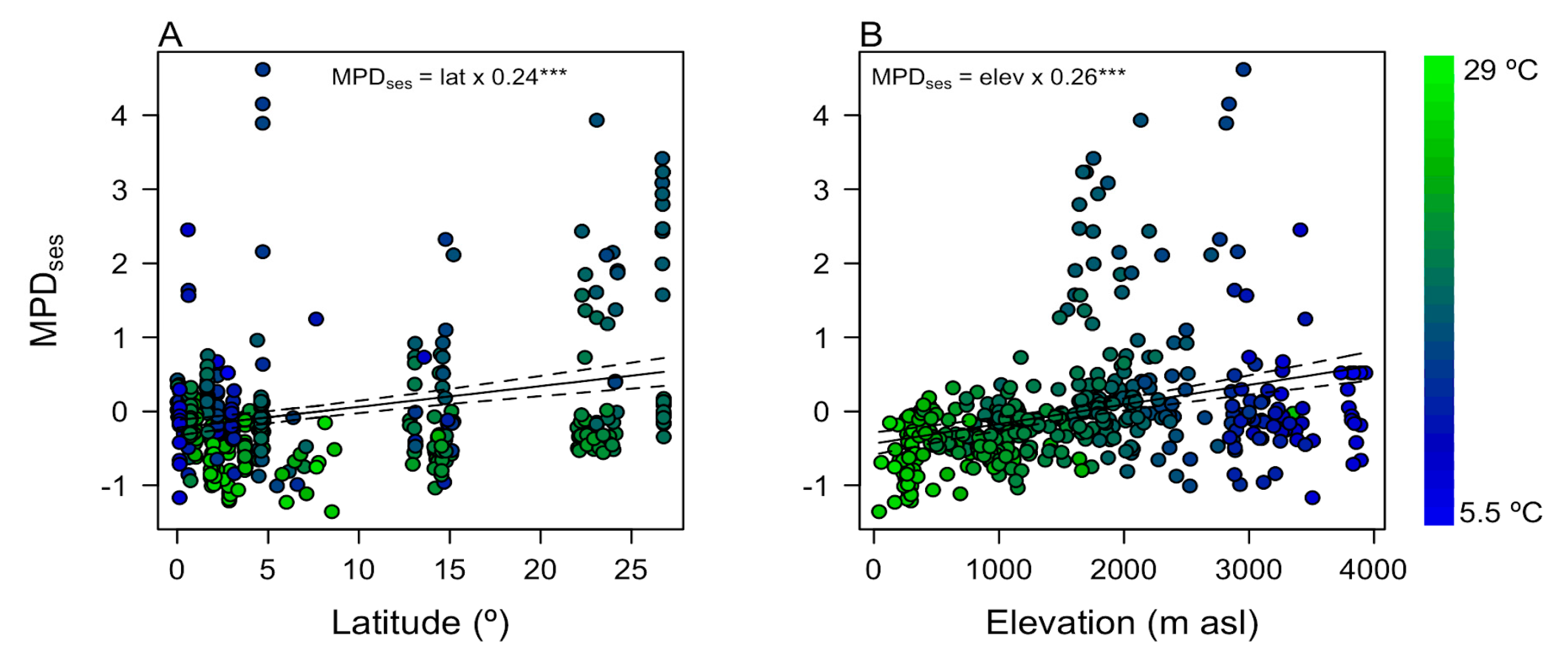
3.2. Local Phylogenetic Diversity and Phylogenetic Turnover along Latitude and Elevation in Andean Tree Communities
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Humboldt, A.; von Bonpland, A. Essai Sur La Géographie Des Plantes; Chez Levrault Schoell Campagnie Libr.: Paris, France, 1805. [Google Scholar]
- Mittelbach, G.G.; Schemske, D.W.; Cornell, H.V.; Allen, A.P.; Brown, J.M.; Bush, M.B.; Harrison, S.P.; Hurlbert, A.H.; Knowlton, N.; Lessios, H.A.; et al. Evolution and the Latitudinal Diversity Gradient: Speciation, Extinction and Biogeography. Ecol. Lett. 2007, 10, 315–331. [Google Scholar] [CrossRef]
- Laiolo, P.; Pato, J.; Obeso, J.R. Ecological and Evolutionary Drivers of the Elevational Gradient of Diversity. Ecol. Lett. 2018, 21, 1022–1032. [Google Scholar] [CrossRef] [PubMed]
- Etienne, R.S.; Cabral, J.S.; Hagen, O.; Hartig, F.; Hurlbert, A.H.; Pellissier, L.; Pontarp, M.; Storch, D. A Minimal Model for the Latitudinal Diversity Gradient Suggests a Dominant Role for Ecological Limits. Am. Nat. 2019, 194, E122–E133. [Google Scholar] [CrossRef] [PubMed]
- Kreft, H.; Jetz, W. Global Patterns and Determinants of Vascular Plant Diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 5925–5930. [Google Scholar] [CrossRef]
- Latham, R.E.; Ricklefs, R.E. Continental Comparisons of Temperate-Zone Tree Species Diversity. In Species Diversity in Ecological Communities; University of Chicago Press: Chicago, IL, USA, 1993; pp. 294–314. [Google Scholar]
- Ricklefs, R.E.; He, F. Region Effects Influence Local Tree Species Diversity. Proc. Natl. Acad. Sci. USA 2016, 113, 674–679. [Google Scholar] [CrossRef] [PubMed]
- Wiens, J.J.; Donoghue, M.J. Historical Biogeography, Ecology and Species Richness. Trends Ecol. Evol. 2004, 19, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Ricklefs, R.E. A Comprehensive Framework for Global Patterns in Biodiversity. Ecol. Lett. 2004, 7, 1–15. [Google Scholar] [CrossRef]
- Ricklefs, R.E. Evolutionary Diversification and the Origin of the Diversity-Environment Relationship. Ecology 2006, 87, 3–13. [Google Scholar] [CrossRef]
- Kerkhoff, A.J.; Moriarty, P.E.; Weiser, M.D. The Latitudinal Species Richness Gradient in New World Woody Angiosperms Is Consistent with the Tropical Conservatism Hypothesis. Proc. Natl. Acad. Sci. USA 2014, 111, 8125–8130. [Google Scholar] [CrossRef] [PubMed]
- Crisp, M.D.; Cook, L.G. Phylogenetic Niche Conservatism: What Are the Underlying Evolutionary and Ecological Causes? New Phytol. 2012, 196, 681–694. [Google Scholar] [CrossRef] [PubMed]
- González-Caro, S.; Umaña, M.N.; Álvarez, E.; Stevenson, P.R.; Swenson, N.G. Phylogenetic Alpha and Beta Diversity in Tropical Tree Assemblages along Regional-Scale Environmental Gradients in Northwest South America. J. Plant Ecol. 2014, 7, 145–153. [Google Scholar] [CrossRef]
- González-Caro, S.; Duque, Á.; Feeley, K.J.; Cabrera, E.; Phillips, J.; Ramirez, S.; Yepes, A. The Legacy of Biogeographic History on the Composition and Structure of Andean Forests. Ecology 2020, 101, e03131. [Google Scholar] [CrossRef] [PubMed]
- Segovia, R.A.; Pennington, R.T.; Baker, T.R.; de Souza, F.C.; Neves, D.M.; Davis, C.C.; Armesto, J.J.; Olivera-Filho, A.T.; Dexter, K.G. Freezing and Water Availability Structure the Evolutionary Diversity of Trees across the Americas. Sci. Adv. 2020, 6, eaaz5373. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, S.; González-Caro, S.; Phillips, J.; Cabrera, E.; Feeley, K.J.; Duque, Á. The Influence of Historical Dispersal on the Phylogenetic Structure of Tree Communities in the Tropical Andes. Biotropica 2019, 51, 500–508. [Google Scholar] [CrossRef]
- Hoorn, C.; Wesselingh, F.P.; Ter Steege, H.; Bermudez, M.A.; Mora, A.; Sevink, J.; Sanmartín, I.; Sanchez-Meseguer, A.; Anderson, C.L.; Figueiredo, J.P.; et al. Amazonia through Time: Andean Uplift, Climate Change, Landscape Evolution, and Biodiversity. Science 2010, 330, 927–931. [Google Scholar] [CrossRef]
- Qian, H.; Ricklefs, R.E. Out of the Tropical Lowlands: Latitude versus Elevation. Trends Ecol. Evol. 2016, 31, 738–741. [Google Scholar] [CrossRef]
- Scotese, C.R. A Continental Drift Flipbook. J. Geol. 2004, 112, 729–741. [Google Scholar] [CrossRef]
- Carvalho, M.R.; Jaramillo, C.; de la Parra, F.; Caballero-Rodríguez, D.; Herrera, F.; Wing, S.; Turner, B.L.; D’Apolito, C.; Romero-Báez, M.; Narváez, P.; et al. Extinction at the End-Cretaceous and the Origin of Modern Neotropical Rainforests. Science 2021, 372, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Fine, P.V.A.; Ree, R.H. Evidence for a Time-Integrated Species-Area Effect on the Latitudinal Gradient in Tree Diversity. Am. Nat. 2006, 168, 796–804. [Google Scholar] [CrossRef]
- Jaramillo, C.; Rueda, M.J.; Mora, G. Cenozoic Plant Diversity in the Neotropics. Science 2006, 311, 1893–1896. [Google Scholar] [CrossRef]
- Revell, L.J. Phytools: An R Package for Phylogenetic Comparative Biology (and Other Things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Ricklefs, R.E. Estimating Diversification Rates from Phylogenetic Information. Trends Ecol. Evol. 2007, 22, 601–610. [Google Scholar] [CrossRef]
- Pennell, M.W.; Eastman, J.M.; Slater, G.J.; Brown, J.W.; Uyeda, J.C.; Fitzjohn, R.G.; Alfaro, M.E.; Harmon, L.J. Geiger v2.0: An Expanded Suite of Methods for Fitting Macroevolutionary Models to Phylogenetic Trees. Bioinformatics 2014, 30, 2216–2218. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Jin, Y.; Qian, H.V. PhyloMaker: An R Package that Can Generate Very Large Phylogenies for Vascular Plants. Ecography 2019, 42, 1353–1359. [Google Scholar] [CrossRef]
- Smith, S.A.; Brown, J.W. Constructing a Broadly Inclusive Seed Plant Phylogeny. Am. J. Bot. 2018, 105, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Rangel, T.F.; Colwell, R.K.; Graves, G.R.; Fučíková, K.; Rahbek, C.; Diniz-Filho, J.A.F. Phylogenetic Uncertainty Revisited: Implications for Ecological Analyses. Evolution 2015, 69, 1301–1312. [Google Scholar] [CrossRef]
- Webb, C.O. Exploring the Phylogenetic Structure of Ecological Communities: An Example for Rain Forest Trees. Am. Nat. 2000, 156, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R Tools for Integrating Phylogenies and Ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [PubMed]
- Gentry, A.H. Neotropical Floristic Diversity: Phytogeographical Connections between Central and South America, Pleistocene Climatic Fluctuations, or an Accident of the Andean Orogeny? Ann. Mo. Bot. Gard. 1982, 69, 557–593. [Google Scholar] [CrossRef]
- Linan, A.G.; Myers, J.A.; Edwards, C.E.; Zanne, A.E.; Smith, S.A.; Arellano, G.; Cayola, L.; Farfan-Ríos, W.; Fuentes, A.F.; García-Cabrera, K.; et al. The Evolutionary Assembly of Forest Communities along Environmental Gradients: Recent Diversification or Sorting of Pre-Adapted Clades? New Phytol. 2021, 232, 2506–2519. [Google Scholar] [CrossRef] [PubMed]
- Malizia, A.; Blundo, C.; Carilla, J.; Acosta, O.O.; Cuesta, F.; Duque, A.; Aguirre, N.; Aguirre, Z.; Ataroff, M.; Baez, S.; et al. Elevation and Latitude Drives Structure and Tree Species Composition in Andean Forests: Results from a Large-Scale Plot Network. PLoS ONE 2020, 15, e0231553. [Google Scholar] [CrossRef] [PubMed]
- Madriñán, S.; Cortés, A.J.; Richardson, J.E. Páramo Is the World’s Fastest Evolving and Coolest Biodiversity Hotspot. Front. Genet. 2013, 4, 1–7. [Google Scholar] [CrossRef]
- Qian, H.; Ricklefs, R.E.; Thuiller, W. Evolutionary Assembly of Flowering Plants into Sky Islands. Nat. Ecol. Evol. 2021, 5, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Vargas, O.M.; Dick, C.W. Diversification History of Neotropical Lecythidaceae, an Ecologically Dominant Tree Family of Amazon Rain Forest. In Neotropical Diversification: Patterns and Processes. Fascinating Life Sciences; Springer: Cham, Switzerland, 2020; pp. 791–809. [Google Scholar] [CrossRef]
- Jaramillo, C.; Cárdenas, A. Global Warming and Neotropical Rainforests: A Historical Perspective. Annu. Rev. Earth Planet. Sci. 2013, 41, 741–766. [Google Scholar] [CrossRef]
- Montes, C.; Cardona, A.; Jaramillo, C.; Pardo, A.; Silva, J.C.; Valencia, V.; Ayala, C.; Pérez-Angel, L.C.; Rodriguez-Parra, L.A.; Ramirez, V.; et al. Middle Miocene Closure of the Central American Seaway. Science 2015, 348, 226–229. [Google Scholar] [CrossRef]
- Valiente-Banuet, A.; Verdú, M. Facilitation Can Increase the Phylogenetic Diversity of Plant Communities. Ecol. Lett. 2007, 10, 1029–1036. [Google Scholar] [CrossRef]
- Sargent, R.D.; Ackerly, D.D. Plant-Pollinator Interactions and the Assembly of Plant Communities. Trends Ecol. Evol. 2008, 23, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Culmsee, H.; Leuschner, C.; Moser, G.; Pitopang, R. Forest Aboveground Biomass along an Elevational Transect in Sulawesi, Indonesia, and the Role of Fagaceae in Tropical Montane Rain Forests. J. Biogeogr. 2010, 37, 960–974. [Google Scholar] [CrossRef]
- Olson, M.E.; Soriano, D.; Rosell, J.A.; Anfodillo, T.; Donoghue, M.J.; Edwards, E.J.; León-Gómez, C.; Dawson, T.; Julio Camarero Martínez, J.; Castorena, M.; et al. Plant Height and Hydraulic Vulnerability to Drought and Cold. Proc. Natl. Acad. Sci. USA 2018, 115, 7551–7556. [Google Scholar] [CrossRef]
- Duque, A.; Peña, M.A.; Cuesta, F.; González-Caro, S.; Kennedy, P.; Phillips, O.L.; Calderón-Loor, M.; Blundo, C.; Carilla, J.; Cayola, L.; et al. Mature Andean Forests as Globally Important Carbon Sinks and Future Carbon Refuges. Nat. Commun. 2021, 12, 2138. [Google Scholar] [CrossRef]
- Duque, A.; Stevenson, P.R.; Feeley, K.J. Thermophilization of Adult and Juvenile Tree Communities in the Northern Tropical Andes. Proc. Natl. Acad. Sci. USA 2015, 112, 10744–10749. [Google Scholar] [CrossRef]
- Fadrique, B.; Báez, S.; Duque, Á.; Malizia, A.; Blundo, C.; Carilla, J.; Osinaga-Acosta, O.; Malizia, L.; Silman, M.; Farfán-Ríos, W.; et al. Widespread but Heterogeneous Responses of Andean Forests to Climate Change. Nature 2018, 564, 207–212. [Google Scholar] [CrossRef]
- Vellend, M.; Cornwell, W.K.; Magnuson-Ford, K.; Mooers, A.Ø. Measuring phylogenetic biodiversity. Front. Meas. Biol. Divers. 2010, 194–207. [Google Scholar]
- Schluter, D.; Pennell, M.W. Speciation gradients and the distribution of biodiversity. Nature 2017, 546, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Rabosky, D.L.; Title, P.O.; Huang, H. Minimal effects of latitude on present-day speciation rates in New World birds. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142889. [Google Scholar] [CrossRef]
- Igea, J.; Tanentzap, A.J. Angiosperm speciation cools down in the tropics. Ecol. Lett. 2020, 23, 692–700. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Caro, S.; Tello, J.S.; Myers, J.A.; Feeley, K.; Blundo, C.; Calderón-Loor, M.; Carilla, J.; Cayola, L.; Cuesta, F.; Farfán, W.; et al. Historical Assembly of Andean Tree Communities. Plants 2023, 12, 3546. https://doi.org/10.3390/plants12203546
González-Caro S, Tello JS, Myers JA, Feeley K, Blundo C, Calderón-Loor M, Carilla J, Cayola L, Cuesta F, Farfán W, et al. Historical Assembly of Andean Tree Communities. Plants. 2023; 12(20):3546. https://doi.org/10.3390/plants12203546
Chicago/Turabian StyleGonzález-Caro, Sebastián, J. Sebastián Tello, Jonathan A. Myers, Kenneth Feeley, Cecilia Blundo, Marco Calderón-Loor, Julieta Carilla, Leslie Cayola, Francisco Cuesta, William Farfán, and et al. 2023. "Historical Assembly of Andean Tree Communities" Plants 12, no. 20: 3546. https://doi.org/10.3390/plants12203546
APA StyleGonzález-Caro, S., Tello, J. S., Myers, J. A., Feeley, K., Blundo, C., Calderón-Loor, M., Carilla, J., Cayola, L., Cuesta, F., Farfán, W., Fuentes, A. F., Garcia-Cabrera, K., Grau, R., Idarraga, Á., Loza, M. I., Malhi, Y., Malizia, A., Malizia, L., Osinaga-Acosta, O., ... Duque, Á. (2023). Historical Assembly of Andean Tree Communities. Plants, 12(20), 3546. https://doi.org/10.3390/plants12203546