


Effect of Ferulago lutea (Poir.) Grande Essential Oil on Molecular Hallmarks of Skin Aging

 , ,
, ,  , ,
, ,  and
and 
Abstract
:
1. Introduction
2. Results
2.1. Chemical Composition
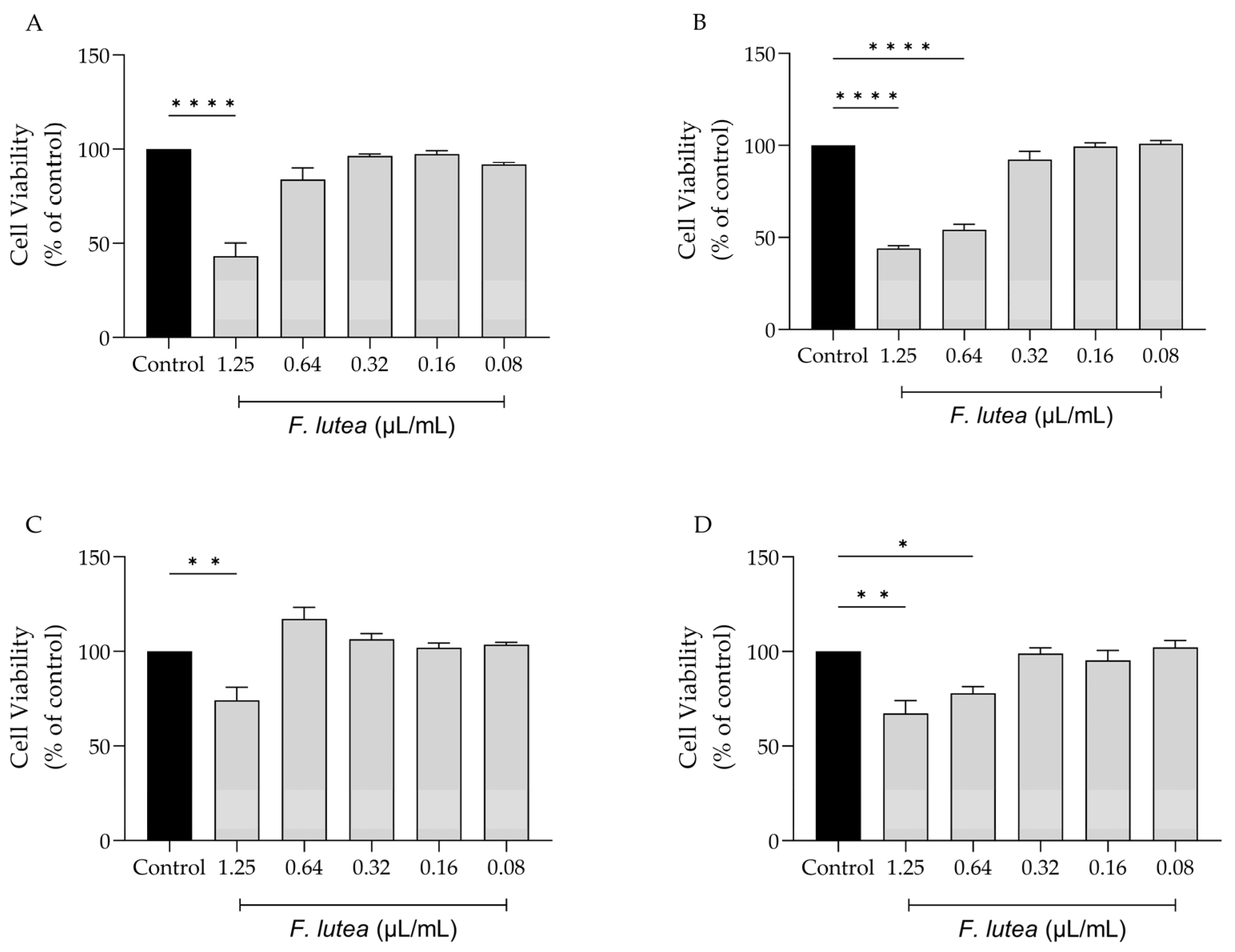
2.2. Effect of F. lutea on Cell Viability
2.3. The Essential Oil of F. lutea Exerts Anti-Inflammatory Effect via Inhibition of NF-κB Pathway
2.4. Ferulago lutea Essential Oil Promotes Cell Migration
2.5. Ferulago lutea Essential Oil Affects Lipogenesis Differentially
2.6. Ferulago lutea Essential Oil Has Depigmenting Properties
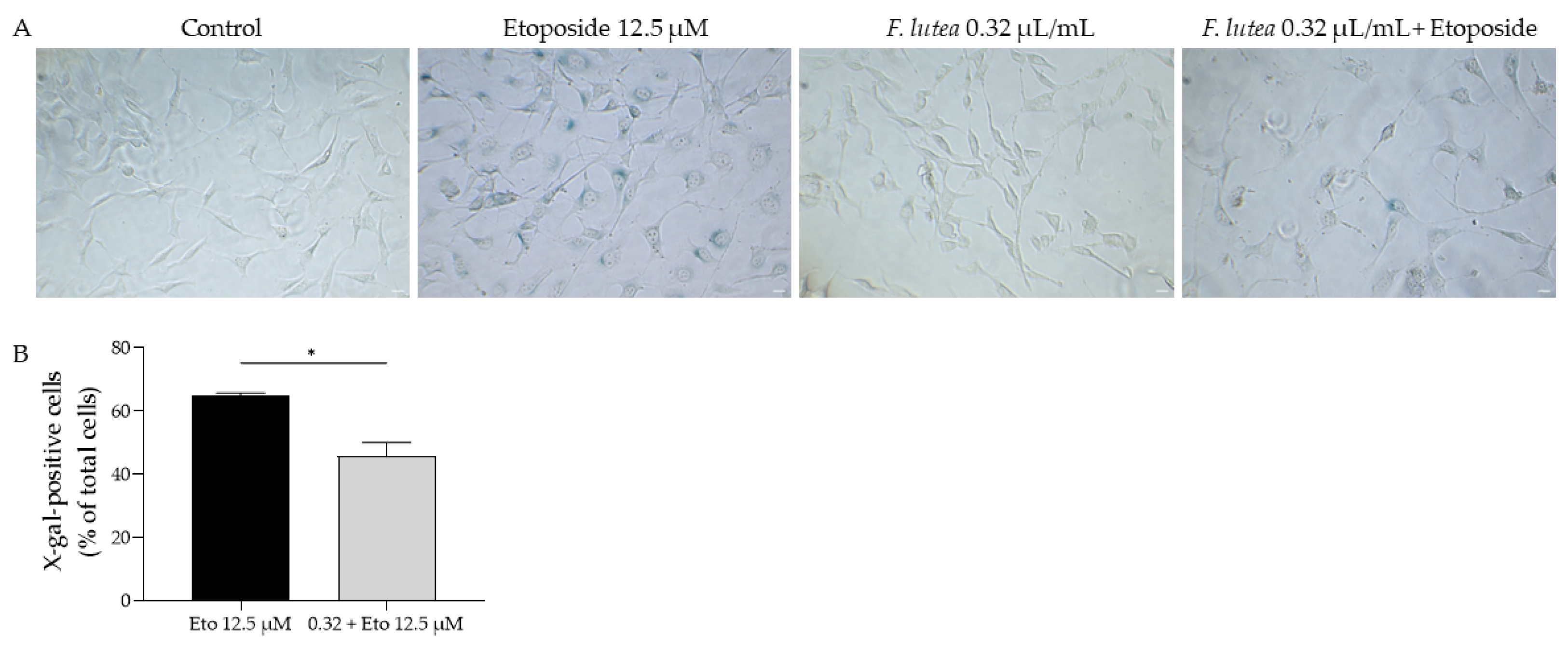
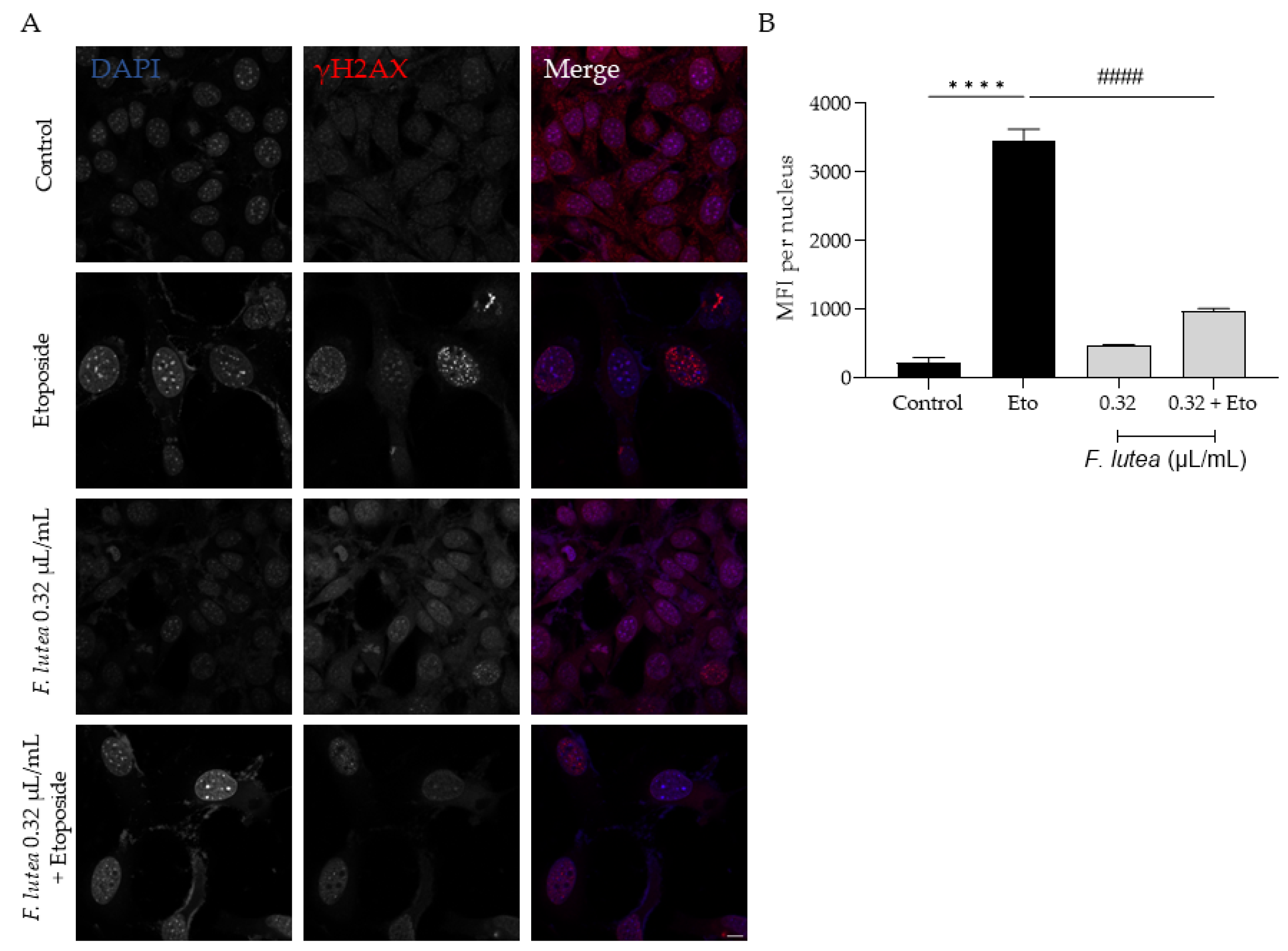
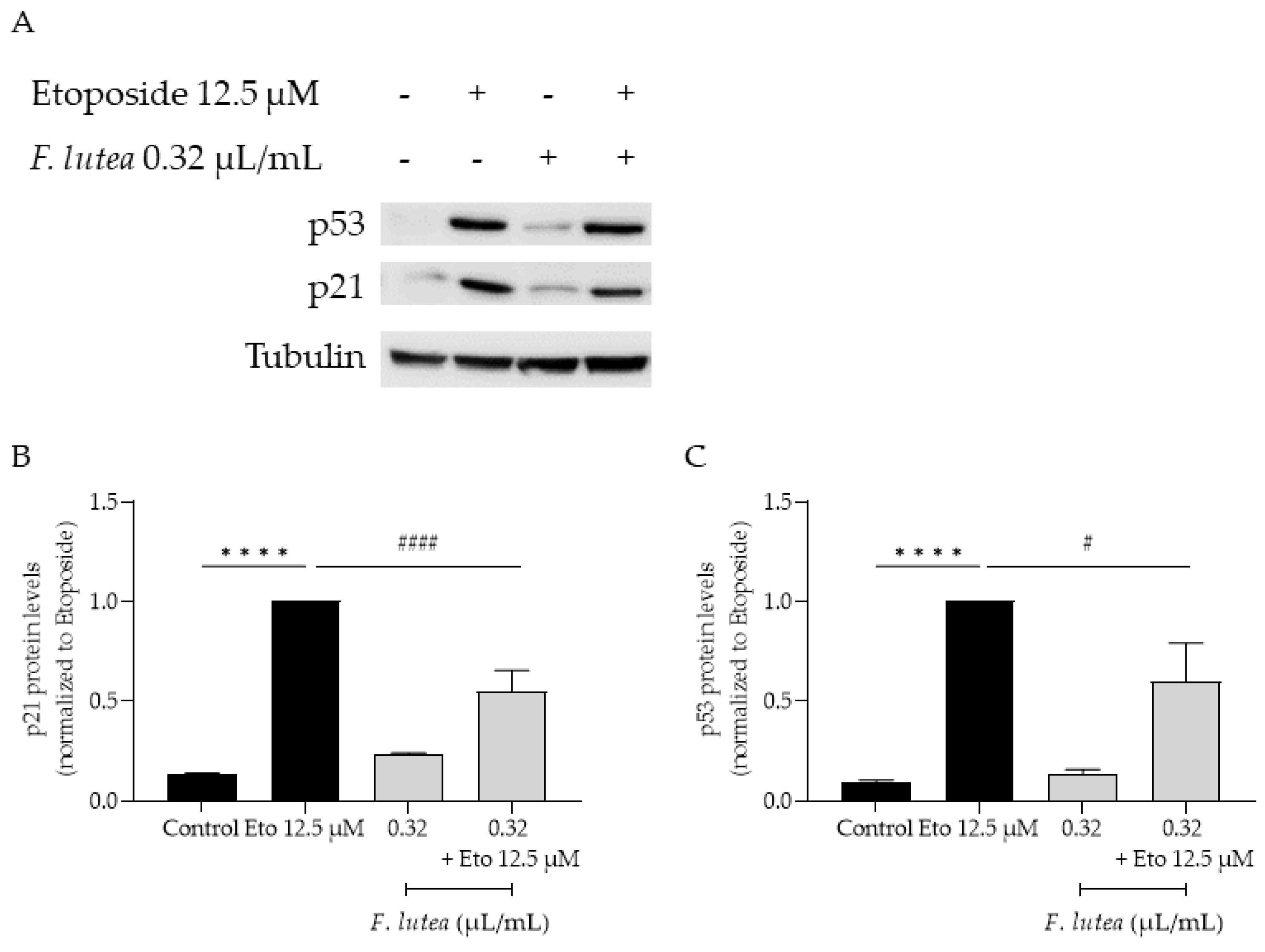
2.7. Ferulago lutea Essential Oil Has Anti-Senescence Properties
3. Discussion
4. Materials and Methods
4.1. Plant Material and Essential Oil Distillation
4.2. Chemical Characterization of the Essential Oil
4.3. Cell Culture
4.4. Effect on Cell Viability
4.5. Anti-Inflammatory Potential
4.5.1. Nitric Oxide Production
4.5.2. Western Blot Analysis of Pro-Inflammatory Mediators
4.6. Cell Migration
4.7. Inhibition of Lipogenesis
4.7.1. Lipogenesis Induction
4.7.2. Oil Red-O Fluorescent Staining
4.7.3. Lipid Accumulation Quantification by Oil Red O Staining
4.8. Depigmenting Effect
4.9. Anti-Senescence Potential
4.9.1. Senescence-Associated β-Galactosidase Activity
4.9.2. yH2AX Staining
4.9.3. p21 and p53 Protein Levels
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. World Report on Ageing and Health; World Health Organization: Geneva, Switzerland, 2015; ISBN 9789241565042. [Google Scholar]
- Pilkington, S.M.; Bulfone-Paus, S.; Griffiths, C.E.M.; Watson, R.E.B. Inflammaging and the Skin. J. Investig. Dermatol. 2021, 141, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.C.; Park, T.J.; Kang, H.Y. Skin-Aging Pigmentation: Who Is the Real Enemy? Cells 2022, 11, 2541. [Google Scholar] [CrossRef] [PubMed]
- Cavinato, M.; Waltenberger, B.; Baraldo, G.; Grade, C.V.C.; Stuppner, H.; Jansen-Dürr, P. Plant Extracts and Natural Compounds Used against UVB-Induced Photoaging. Biogerontology 2017, 18, 499–516. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.; Estanqueiro, M.; Oliveira, M.; Sousa Lobo, J. Main Benefits and Applicability of Plant Extracts in Skin Care Products. Cosmetics 2015, 2, 48–65. [Google Scholar] [CrossRef]
- WFO. Ferulago W.D.J.Koch. Available online: https://www.worldfloraonline.org/taxon/wfo-4000014699 (accessed on 19 September 2023).
- Badalamenti, N.; Ilardi, V.; Rosselli, S.; Bruno, M. The Ethnobotany, Phytochemistry and Biological Properties of Genus Ferulago—A Review. J. Ethnopharmacol. 2021, 274, 114050. [Google Scholar] [CrossRef]
- Pinto, E.; Hrimpeng, K.; Lopes, G.; Vaz, S.; Gonçalves, M.J.; Cavaleiro, C.; Salgueiro, L. Antifungal Activity of Ferulago capillaris Essential Oil against Candida, Cryptococcus, Aspergillus and Dermatophyte Species. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1311–1320. [Google Scholar] [CrossRef]
- Znati, M.; Jabrane, A.; Hajlaoui, H.; Harzallah-Skhiri, F.; Bouajila, J.; Casanova, J.; Jannet, H. Ben Chemical Composition and in Vitro Evaluation of Antimicrobial and Anti-Acetylcholinesterase Properties of the Flower Oil of Ferula lutea. Nat. Prod. Commun. 2012, 7, 1934578X1200700. [Google Scholar] [CrossRef]
- Ben Salem, S.; Znati, M.; Jabrane, A.; Casanova, J.; Ben Jannet, H. Chemical Composition, Antimicrobial, Anti-Acetylcholinesterase and Cytotoxic Activities of the Root Essential Oil from the Tunisian Ferula lutea (Poir.) Maire (Apiaceae). J. Essent. Oil Bear. Plants 2016, 19, 897–906. [Google Scholar] [CrossRef]
- Rahmouni, M.; Laouer, H.; Dahamna, S.; Gali, L.; Bensouici, C.; Flamini, G.; Akkal, S. Biological Activities and Phytochemical Content of Essential Oil and Methanol Extracts of Ferula lutea (Poir.) Maire Growing in Algeria. Biocatal. Agric. Biotechnol. 2021, 34, 102017. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Vu, R.; Jin, S.; Sun, P.; Haensel, D.; Nguyen, Q.H.; Dragan, M.; Kessenbrock, K.; Nie, Q.; Dai, X. Wound Healing in Aged Skin Exhibits Systems-Level Alterations in Cellular Composition and Cell-Cell Communication. Cell Rep. 2022, 40, 111155. [Google Scholar] [CrossRef] [PubMed]
- Makrantonaki, E.; Zouboulis, C.C. Skin Alterations and Diseases in Advanced Age. Drug Discov. Today Dis. Mech. 2008, 5, e153–e162. [Google Scholar] [CrossRef]
- Esler, W.P.; Tesz, G.J.; Hellerstein, M.K.; Beysen, C.; Sivamani, R.; Turner, S.M.; Watkins, S.M.; Amor, P.A.; Carvajal-Gonzalez, S.; Geoly, F.J.; et al. Human Sebum Requires de Novo Lipogenesis, Which Is Increased in Acne Vulgaris and Suppressed by Acetyl-CoA Carboxylase Inhibition. Sci. Transl. Med. 2019, 11, eaau8465. [Google Scholar] [CrossRef] [PubMed]
- Mitro, N.; Vargas, L.; Romeo, R.; Koder, A.; Saez, E. T0901317 Is a Potent PXR Ligand: Implications for the Biology Ascribed to LXR. FEBS Lett. 2007, 581, 1721–1726. [Google Scholar] [CrossRef]
- Kudlova, N.; De Sanctis, J.B.; Hajduch, M. Cellular Senescence: Molecular Targets, Biomarkers, and Senolytic Drugs. Int. J. Mol. Sci. 2022, 23, 4168. [Google Scholar] [CrossRef]
- de Sá, D.C.; Festa Neto, C. Inflammasomes and Dermatology. An. Bras. Dermatol. 2016, 91, 566–578. [Google Scholar] [CrossRef]
- Yu, S.; Li, Y.; Zhou, Y.; Follansbee, T.; Hwang, S.T. Immune Mediators and Therapies for Pruritus in Atopic Dermatitis and Psoriasis. J. Cutan. Immunol. Allergy 2019, 2, 4–14. [Google Scholar] [CrossRef]
- Schwingen, J.; Kaplan, M.; Kurschus, F.C. Review—Current Concepts in Inflammatory Skin Diseases Evolved by Transcriptome Analysis: In-Depth Analysis of Atopic Dermatitis and Psoriasis. Int. J. Mol. Sci. 2020, 21, 699. [Google Scholar] [CrossRef]
- Song, A.; Lee, S.E.; Kim, J.H. Immunopathology and Immunotherapy of Inflammatory Skin Diseases. Immune Netw. 2022, 22, e7. [Google Scholar] [CrossRef]
- Goh, B.H.; Mocan, A.; Xiao, J.; Mah, S.H.; Yap, W.H. Editorial: Targeting Human Inflammatory Skin Diseases with Natural Products: Exploring Potential Mechanisms and Regulatory Pathways. Front. Pharmacol. 2021, 12, 791151. [Google Scholar] [CrossRef]
- Li, W.; Papa, F.; Shi, J.; Maggi, F.; Chen, X. The Chemical Constituents and the Hepato-Protective Effect of the Essential Oil of Ferulago campestris (Besser) Grecescu (Apiaceae). J. Essent. Oil Bear. Plants 2016, 19, 1701–1708. [Google Scholar] [CrossRef]
- Bantal, V.; Biradar, S. Screening of Natural Antioxidants by Using L-Arginine Induced Acute Pancreatitis Model. Int. J. Drug Dev. Res. 2012, 4, 284–297. [Google Scholar]
- Yu, L.; Yan, J.; Sun, Z. D-Limonene Exhibits Anti-Inflammatory and Antioxidant Properties in an Ulcerative Colitis Rat Model via Regulation of INOS, COX-2, PGE2 and ERK Signaling Pathways. Mol. Med. Rep. 2017, 15, 2339–2346. [Google Scholar] [CrossRef] [PubMed]
- Santana, H.S.R.; de Carvalho, F.O.; Silva, E.R.; Santos, N.G.L.; Shanmugam, S.; Santos, D.N.; Wisniewski, J.O.; Junior, J.S.C.; Nunes, P.S.; Araujo, A.A.S.; et al. Anti-Inflammatory Activity of Limonene in the Prevention and Control of Injuries in the Respiratory System: A Systematic Review. Curr. Pharm. Des. 2020, 26, 2182–2191. [Google Scholar] [CrossRef]
- Rufino, A.T.; Ribeiro, M.; Judas, F.; Salgueiro, L.; Lopes, M.C.; Cavaleiro, C.; Mendes, A.F. Anti-Inflammatory and Chondroprotective Activity of (+)-α-Pinene: Structural and Enantiomeric Selectivity. J. Nat. Prod. 2014, 77, 264–269. [Google Scholar] [CrossRef]
- Li, X.-J.; Yang, Y.-J.; Li, Y.-S.; Zhang, W.K.; Tang, H.-B. α-Pinene, Linalool, and 1-Octanol Contribute to the Topical Anti-Inflammatory and Analgesic Activities of Frankincense by Inhibiting COX-2. J. Ethnopharmacol. 2016, 179, 22–26. [Google Scholar] [CrossRef]
- Rahbar, I.; Abbasnejad, M.; Haghani, J.; Raoof, M.; Kooshki, R.; Esmaeili-Mahani, S. The Effect of Central Administration of Alpha-pinene on Capsaicin-induced Dental Pulp Nociception. Int. Endod. J. 2019, 52, 307–317. [Google Scholar] [CrossRef]
- Khalid, K.A.; Nawi, A.F.M.; Zulkifli, N.; Barkat, M.A.; Hadi, H. Aging and Wound Healing of the Skin: A Review of Clinical and Pathophysiological Hallmarks. Life 2022, 12, 2142. [Google Scholar] [CrossRef]
- Salas-Oropeza, J.; Jimenez-Estrada, M.; Perez-Torres, A.; Castell-Rodriguez, A.E.; Becerril-Millan, R.; Rodriguez-Monroy, M.A.; Jarquin-Yañez, K.; Canales-Martinez, M.M. Wound Healing Activity of α-Pinene and α-Phellandrene. Molecules 2021, 26, 2488. [Google Scholar] [CrossRef]
- d’Alessio, P.; Mirshahi, M.; Bisson, J.-F.; Bene, M. Skin Repair Properties of D-Limonene and Perillyl Alcohol in Murine Models. Anti-Inflamm. Anti-Allergy Agents Med. Chem. 2014, 13, 29–35. [Google Scholar] [CrossRef]
- Ahmad, M.; Khan, T.H.; Ansari, M.N.; Ahmad, S.F. Enhanced Wound Healing by Topical Administration of D Limonene in Alloxan Induced Diabetic Mice through Reduction of pro-Inflammatory Markers and Chemokine Expression. BMC Genom. 2014, 15, P29. [Google Scholar] [CrossRef]
- Keskin, I.; Gunal, Y.; Ayla, S.; Kolbasi, B.; Sakul, A.; Kilic, U.; Gok, O.; Koroglu, K.; Ozbek, H. Effects of Foeniculum vulgare Essential Oil Compounds, Fenchone and Limonene, on Experimental Wound Healing. Biotech. Histochem. 2017, 92, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C.; Boschnakow, A. Chronological Ageing and Photoageing of the Human Sebaceous Gland. Clin. Exp. Dermatol. 2001, 26, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Soundharrajan, I.; Kim, D.H.; Srisesharam, S.; Kuppusamy, P.; Choi, K.C. R-Limonene Enhances Differentiation and 2-Deoxy-D-Glucose Uptake in 3T3-L1 Preadipocytes by Activating the Akt Signaling Pathway. Evid.-Based Complement. Altern. Med. 2018, 2018, 4573254. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.-T.; Huang, Y.-W.; Hou, C.-Y.; Wang, J.-J.; Wu, C.-C.; Hsieh, S.-L. D-Limonene Promotes Anti-Obesity in 3T3-L1 Adipocytes and High-Calorie Diet-Induced Obese Rats by Activating the AMPK Signaling Pathway. Nutrients 2023, 15, 267. [Google Scholar] [CrossRef] [PubMed]
- Hakozaki, T.; Swanson, C.L.; Bissett, D.L. Hyperpigmentation in Aging Skin. In Textbook of Aging Skin; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1–10. [Google Scholar]
- Zolghadri, S.; Beygi, M.; Mohammad, T.F.; Alijanianzadeh, M.; Pillaiyar, T.; Garcia-Molina, P.; Garcia-Canovas, F.; Munoz-Munoz, J.; Saboury, A.A. Targeting Tyrosinase in Hyperpigmentation: Current Status, Limitations and Future Promises. Biochem. Pharmacol. 2023, 212, 115574. [Google Scholar] [CrossRef]
- Kumar, K.J.S.; Vani, M.G.; Wang, S. Limonene Protects Human Skin Keratinocytes against UVB-induced Photodamage and Photoaging by Activating the Nrf2-dependent Antioxidant Defense System. Environ. Toxicol. 2022, 37, 2897–2909. [Google Scholar] [CrossRef]
- Liyanage, A.; Liyanage, G.; Sirimanna, G.; Schürer, N. Comparative Study on Depigmenting Agents in Skin of Color. J. Clin. Aesthet. Dermatol. 2022, 15, 12–17. [Google Scholar]
- Csekes, E.; Račková, L. Skin Aging, Cellular Senescence and Natural Polyphenols. Int. J. Mol. Sci. 2021, 22, 12641. [Google Scholar] [CrossRef]
- Franco, A.C.; Aveleira, C.; Cavadas, C. Skin Senescence: Mechanisms and Impact on Whole-Body Aging. Trends Mol. Med. 2022, 28, 97–109. [Google Scholar] [CrossRef]
- Passos, J.F.; Nelson, G.; Wang, C.; Richter, T.; Simillion, C.; Proctor, C.J.; Miwa, S.; Olijslagers, S.; Hallinan, J.; Wipat, A.; et al. Feedback between P21 and Reactive Oxygen Production Is Necessary for Cell Senescence. Mol. Syst. Biol. 2010, 6, 347. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.S.; François, M.; Fenech, M.F.; Leifert, W.R. Persistent ΓH2AX: A Promising Molecular Marker of DNA Damage and Aging. Mutat. Res. Rev. Mutat. Res. 2015, 766, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Secerli, J.; Erdem, O.; Bacanlı, M. Antiaging Effects of Limonene in Ageing-Induced HaCaT Cells. Genet. Appl. 2023, 7, 1–8. [Google Scholar] [CrossRef]
- Karthikeyan, R.; Kanimozhi, G.; Madahavan, N.R.; Agilan, B.; Ganesan, M.; Prasad, N.R.; Rathinaraj, P. Alpha-Pinene Attenuates UVA-Induced Photoaging through Inhibition of Matrix Metalloproteinases Expression in Mouse Skin. Life Sci. 2019, 217, 110–118. [Google Scholar] [CrossRef]
- Council of Europe. European Pharmacopoeia, 7th ed.; Directorate for the Quality of Medicines & HealthCare of the Council of Europe: Strasbourg, France, 2010; ISBN 978-92-871-6700-2. [Google Scholar]
- Alves-Silva, J.M.; Zuzarte, M.; Gonçalves, M.J.; Cruz, M.T.; Cavaleiro, C.; Salgueiro, L. Unveiling the Bioactive Potential of the Essential Oil of a Portuguese Endemism, Santolina impressa. J. Ethnopharmacol. 2019, 244, 112–120. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectroscopy, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Stein, S.E. “Retention Indices” by NIST Mass Spec Data Center. In NIST Chemistry WebBook; Linstrom, P.J., Mallard, W.J., Eds.; NIST Standard Reference Database Number 69; National Institute of Standards and Technology: Gaithersburg, MD, USA, 2017. [Google Scholar]
- Joulain, D.; König, W.A. The Atlas of Spectral Data of Sesquiterpene Hydrocarbons; E.B.-Verlag: Hamburg, Germany, 1998; ISBN 9783930826483. [Google Scholar]
- El-Sayed, A. The Pherobase: Database of Insect Pheromones and Semiochemicals. 2007. Available online: https://www.pherobase.com/database/floral-compounds/floral-taxa-compounds-index.php (accessed on 30 September 2022).
- McLafferty, F.W. Wiley Registry of Mass Spectral Data/NIST08, 9th ed.; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2009; ISBN 0470520361. [Google Scholar]
- Alves-Silva, J.M.; Guerra, I.; Gonçalves, M.J.; Cavaleiro, C.; Cruz, M.T.; Figueirinha, A.; Salgueiro, L. Chemical Composition of Crithmum maritimum L. Essential Oil and Hydrodistillation Residual Water by GC-MS and HPLC-DAD-MS/MS, and Their Biological Activities. Ind. Crops Prod. 2020, 149, 112329. [Google Scholar] [CrossRef]
- Moreira, P.; Sousa, F.J.; Matos, P.; Brites, G.S.; Gonçalves, M.J.; Cavaleiro, C.; Figueirinha, A.; Salgueiro, L.; Batista, M.T.; Branco, P.C.; et al. Chemical Composition and Effect against Skin Alterations of Bioactive Extracts Obtained by the Hydrodistillation of Eucalyptus globulus Leaves. Pharmaceutics 2022, 14, 561. [Google Scholar] [CrossRef] [PubMed]
- Piras, A.; Maccioni, A.; Falconieri, D.; Porcedda, S.; Gonçalves, M.J.; Alves-Silva, J.M.; Silva, A.; Cruz, M.T.; Salgueiro, L.; Maxia, A. Chemical Composition and Biological Activity of Essential Oil of Teucrium scordium L. subsp. Scordioides (Schreb.) Arcang. (Lamiaceae) from Sardinia Island (Italy). Nat. Prod. Res. 2022, 36, 5828–5835. [Google Scholar] [CrossRef]
- Cruz, M.T.; Duarte, C.B.; Gonçalo, M.; Figueiredo, A.; Carvalho, A.P.; Lopes, M.C. Granulocyte-Macrophage Coony-Stimulating Factor Activates the Transcription of Nuclear Factor Kappa B and Induces the Expression of Nitric Oxide Synthase in a Skin Dendritic Cell Line. Immunol. Cell Biol. 2001, 79, 590–596. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of Nitrate, Nitrite, and [15N]Nitrate in Biological Fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Zuzarte, M.; Alves-Silva, J.M.; Alves, M.; Cavaleiro, C.; Salgueiro, L.; Cruz, M.T. New Insights on the Anti-Inflammatory Potential and Safety Profile of Thymus carnosus and Thymus camphoratus Essential Oils and Their Main Compounds. J. Ethnopharmacol. 2018, 225, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Alves-Silva, J.M.; Pedreiro, S.; Cavaleiro, C.; Cruz, M.T.; Figueirinha, A.; Salgueiro, L. Effect of Thymbra capitata (L.) Cav. on Inflammation, Senescence and Cell Migration. Nutrients 2023, 15, 1930. [Google Scholar] [CrossRef] [PubMed]
- Martinotti, S.; Ranzato, E. Scratch Wound Healing Assay. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2020; Volume 2109, pp. 225–229. [Google Scholar]
- Suarez-Arnedo, A.; Torres Figueroa, F.; Clavijo, C.; Arbeláez, P.; Cruz, J.C.; Muñoz-Camargo, C. An Image J Plugin for the High Throughput Image Analysis of In Vitro Scratch Wound Healing Assays. PLoS ONE 2020, 15, e0232565. [Google Scholar] [CrossRef]
- Hong, I.; Rho, H.S.; Kim, D.H.; Lee, M.O. Activation of LXRα Induces Lipogenesis in HaCaT Cells. Arch. Pharm. Res. 2010, 33, 1443–1449. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds * | RI SPB-1 a | RI SW 10 b | % Peak Area |
|---|---|---|---|
| α-Pinene | 930 | 1030 | 36.5 |
| Camphene | 943 | 1077 | 1.5 |
| Sabinene | 964 | 1128 | 0.5 |
| β-Pinene | 970 | 1118 | 1.5 |
| Myrcene | 980 | 1161 | 5.0 |
| p-Cymene | 1009 | 1271 | 1.5 |
| Limonene | 1020 | 1206 | 31.2 |
| β-Phellandrene | 1020 | 1215 | 5.5 |
| (Z)-β-Ocimene | 1025 | 1235 | 3.5 |
| (E)-β-Ocimene | 1035 | 1253 | 1.0 |
| γ-Terpinene | 1046 | 1249 | 0.5 |
| (Z)-Linalool oxide | 1055 | 1439 | 1.0 |
| Terpinolene | 1076 | 1288 | 1.0 |
| p-Cymenene-8-ol | 1160 | 1621 | 0.5 |
| α-Copaene | 1364 | 1487 | 0.5 |
| (E)-Caryophyllene | 1408 | 1590 | 0.5 |
| Germacrene-D | 1466 | 1699 | 0.5 |
| δ-Cadinene | 1508 | 1751 | 0.6 |
| Total identified | 92.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves-Silva, J.M.; Moreira, P.; Cavaleiro, C.; Pereira, C.; Cruz, M.T.; Salgueiro, L. Effect of Ferulago lutea (Poir.) Grande Essential Oil on Molecular Hallmarks of Skin Aging. Plants 2023, 12, 3741. https://doi.org/10.3390/plants12213741
Alves-Silva JM, Moreira P, Cavaleiro C, Pereira C, Cruz MT, Salgueiro L. Effect of Ferulago lutea (Poir.) Grande Essential Oil on Molecular Hallmarks of Skin Aging. Plants. 2023; 12(21):3741. https://doi.org/10.3390/plants12213741
Chicago/Turabian StyleAlves-Silva, Jorge M., Patrícia Moreira, Carlos Cavaleiro, Cláudia Pereira, Maria Teresa Cruz, and Lígia Salgueiro. 2023. "Effect of Ferulago lutea (Poir.) Grande Essential Oil on Molecular Hallmarks of Skin Aging" Plants 12, no. 21: 3741. https://doi.org/10.3390/plants12213741
APA StyleAlves-Silva, J. M., Moreira, P., Cavaleiro, C., Pereira, C., Cruz, M. T., & Salgueiro, L. (2023). Effect of Ferulago lutea (Poir.) Grande Essential Oil on Molecular Hallmarks of Skin Aging. Plants, 12(21), 3741. https://doi.org/10.3390/plants12213741