
Further Molecular Diagnosis Determines Lack of Evidence for Real Seed Transmission of Tomato Leaf Curl New Delhi Virus in Cucurbits

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Obtaining Seeds from ToLCNDV-Infected Cucurbit Plants and Sampling
2.1.1. Seeds Obtained in the COMAV-UPV Greenhouse
2.1.2. Analysis of the Seeds from a Commercial Greenhouse
2.1.3. Analysis of the Seeds from a Commercial Nursery
2.2. Evaluation of ToLCNDV Transmission to Offspring through Seeds
2.3. DNA Extractions from the Different Plant Tissues, Seeds and Seedlings to Make a ToLCNDV Diagnosis
2.4. Detection of ToLCNDV by PCR
2.5. Relative Viral Titer Estimation by Quantitative PCR (qPCR)
2.6. Southern Blot Hybridization
2.7. Rolling-Circle Amplification (RCA)
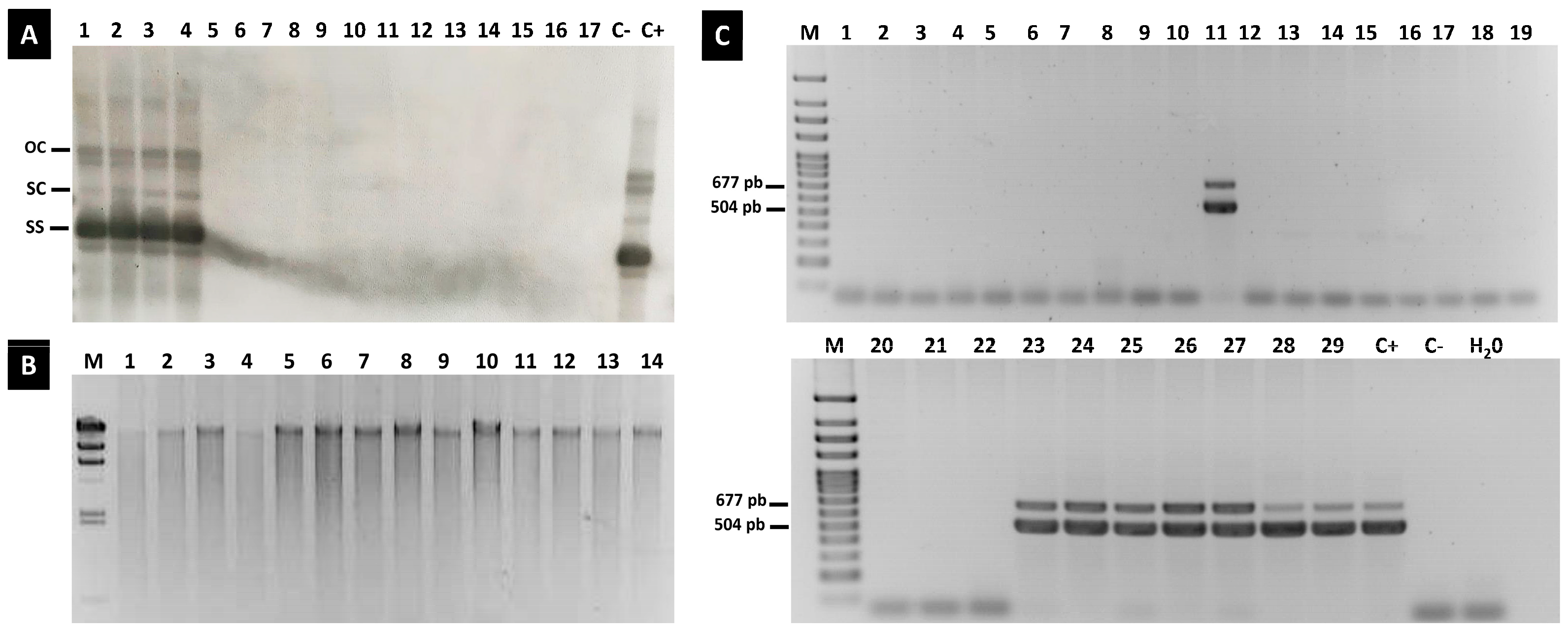
3. Results
3.1. Symptoms Development and ToLCNDV Detection in Vegetative and Reproductive Tissues of Infected Plants
3.2. ToLCNDV Detection by Quantitative PCR in Different Parts of Inoculated Plants, Seeds and Seedlings
3.3. Evaluation of ToLCNDV Presence in the Seedlings Obtained from the Inoculated Plants
3.4. Evaluation of ToLCNDV Presence in the Seeds from a Commercial Greenhouse and the Seedlings of Offspring
3.5. Evaluation of ToLCNDV Presence in the Commercial Seeds from a Nursery and the Seedlings of Their Progeny
3.6. Replicative Forms of ToLCNDV Were Not Identified in Any Seedling of Offspring
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. 2021. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 3 March 2023).
- De Moya-Ruiz, C.; Rabadán, P.; Juárez, M.; Gómez, P. Assessment of the Current Status of Potyviruses in Watermelon and Pumpkin Crops in Spain: Epidemiological Impact of Cultivated Plants and Mixed Infections. Plants 2021, 10, 138. [Google Scholar] [CrossRef]
- Pérez-De-Castro, A.; López-Martín, M.; Esteras, C.; Garcés-Claver, A.; Palomares-Ríus, F.J.; Picó, M.B.; Gómez-Guillamón, M.L. Melon Genome Regions Associated with TGR-1551-Derived Resistance to Cucurbit Yellow Stunting Disorder Virus. Int. J. Mol. Sci. 2020, 21, 5970. [Google Scholar] [CrossRef]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging Virus Diseases Transmitted by Whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef] [PubMed]
- Desbiez, C.; Wipf-Scheibel, C.; Millot, P.; Berthier, K.; Girardot, G.; Gognalons, P.; Hirsch, J.; Moury, B.; Nozeran, K.; Piry, S.; et al. Distribution and Evolution of the Major Viruses Infecting Cucurbitaceous and Solanaceous Crops in the French Mediterranean Area. Virus Res. 2020, 286, 198042. [Google Scholar] [CrossRef] [PubMed]
- Fortes, I.M.; Sánchez-Campos, S.; Fiallo-Olivé, E.; Díaz-Pendón, J.A.; Navas-Castillo, J.; Moriones, E. A Novel Strain of Tomato Leaf Curl New Delhi Virus Has Spread to the Mediterranean Basin. Viruses 2016, 8, 307. [Google Scholar] [CrossRef] [PubMed]
- Jyothsna, P.; Haq, Q.M.I.; Singh, P.; Sumiya, K.V.; Praveen, S.; Rawat, R.; Briddon, R.W.; Malathi, V.G. Infection of Tomato Leaf Curl New Delhi Virus (ToLCNDV), a Bipartite Begomovirus with Betasatellites, Results in Enhanced Level of Helper Virus Components and Antagonistic Interaction between DNA B and Betasatellites. Appl. Microbiol. Biotechnol. 2013, 97, 5457–5471. [Google Scholar] [CrossRef] [PubMed]
- Padidam, M.; Beachy, R.N.; Fauquet, C.M. Tomato Leaf Curl Geminivirus from India Has a Bipartite Genome and Coat Protein Is Not Essential for Infectivity. J. Gen. Virol. 1995, 76, 25–35. [Google Scholar] [CrossRef]
- Srivastava, K.M.; Hallan, V.; Raizada, R.K.; Chandra, G.; Singh, B.P.; Sane, P.V. Molecular Cloning of Indian Tomato Leaf Curl Virus Genolme Following a Simple Method of Concentrating the Supercoiled Replicative Form of Viral DNA. J. Virol. Methods 1995, 51, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Moriones, E.; Praveen, S.; Chakraborty, S. Tomato Leaf Curl New Delhi Virus: An Emerging Virus Complex Threatening Vegetable and Fiber Crops. Viruses 2017, 9, 264. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.S.-E.-A.; Martin, D.P.; Amin, I.; Farooq, M.; Mansoor, S. Tomato Leaf Curl New Delhi Virus: A Widespread Bipartite Begomovirus in the Territory of Monopartite Begomoviruses. Mol. Plant Pathol. 2017, 18, 901–911. [Google Scholar] [CrossRef]
- Juárez, M.; Rabadán, M.P.; Martínez, L.D.; Tayahi, M.; Grande-Pérez, A.; Gómez, P. Natural Hosts and Genetic Diversity of the Emerging Tomato Leaf Curl New Delhi Virus in Spain. Front. Microbiol. 2019, 10, 140. [Google Scholar] [CrossRef] [PubMed]
- Panno, S.; Caruso, A.G.; Troiano, E.; Luigi, M.; Manglli, A.; Vatrano, T.; Iacono, G.; Marchione, S.; Bertin, S.; Tomassoli, L.; et al. Emergence of Tomato Leaf Curl New Delhi Virus in Italy: Estimation of Incidence and Genetic Diversity. Plant Pathol. 2019, 68, 601–608. [Google Scholar] [CrossRef]
- Juárez, M.; Tovar, R.; Fiallo-Olivé, E.; Aranda, M.A.; Gosálvez, B.; Castillo, P.; Moriones, E.; Navas-Castillo, J. First Detection of Tomato Leaf Curl New Delhi Virus Infecting Zucchini in Spain. Plant Dis. 2014, 98, 857. [Google Scholar] [CrossRef]
- Kheireddine, A.; Sifres, A.; Sáez, C.; Picó, B.; López, C. First Report of Tomato Leaf Curl New Delhi Virus Infecting Cucurbit Plants in Algeria. Plant Dis. 2019, 103, 3291. [Google Scholar] [CrossRef]
- Mnari-Hattab, M.; Zammouri, S.; Belkadhi, M.S.; Bellon Doña, D.; Ben Nahia, E.; Hajlaoui, M.R. First Report of Tomato Leaf Curl New Delhi Virus Infecting Cucurbits in Tunisia. New Dis. Rep. 2015, 31, 21. [Google Scholar] [CrossRef]
- Panno, S.; Iacono, G.; Davino, M.; Marchione, S.; Zappardo, V.; Bella, P.; Tomassoli, L.; Accotto, G.P.; Davino, S. First Report of Tomato Leaf Curl New Delhi Virus Affecting Zucchini Squash in an Important Horticultural Area of Southern Italy. New Dis. Rep. 2016, 33, 6. [Google Scholar] [CrossRef]
- Sifres, A.; Sáez, C.; Ferriol, M.; Selmani, E.A.; Riado, J.; Picó, B.; López, C. First Report of Tomato Leaf Curl New Delhi Virus Infecting Zucchini in Morocco. Plant Dis. 2018, 102, 1045. [Google Scholar] [CrossRef]
- Simmons, H.E.; Holmes, E.C.; Gildow, F.E.; Bothe-Goralczyk, M.A.; Stephenson, A.G. Experimental Verification of Seed Transmission of Zucchini Yellow Mosaic Virus. Plant Dis. 2011, 95, 751–754. [Google Scholar] [CrossRef]
- Chang, H.-H.; Ku, H.-M.; Tsai, W.-S.; Chien, R.-C.; Jan, F.-J. Identification and Characterization of a Mechanical Transmissible Begomovirus Causing Leaf Curl on Oriental Melon. Eur. J. Plant Pathol. 2010, 127, 219–228. [Google Scholar] [CrossRef]
- Islam, S.; Munshi, A.D.; Mandal, B.; Kumar, R.; Behera, T.K. Genetics of Resistance in Luffa Cylindrica Roem. against Tomato Leaf Curl New Delhi Virus. Euphytica 2010, 174, 83–89. [Google Scholar] [CrossRef]
- Khan, M.S.; Ji, S.H.; Chun, C. Begomoviruses and Their Emerging Threats in South Korea: A Review. Plant Pathol. J. 2012, 28, 123–136. [Google Scholar] [CrossRef]
- López, C.; Ferriol, M.; Picó, M.B. Mechanical Transmission of Tomato Leaf Curl New Delhi Virus to Cucurbit Germplasm: Selection of Tolerance Sources in Cucumis Melo. Euphytica 2015, 204, 679–691. [Google Scholar] [CrossRef]
- Sohrab, S.S.; Karim, S.; Varma, A.; Azhar, E.I.; Mandal, B.; Abuzenadah, A.M.; Chaudhary, A.G. Factors Affecting Sap Transmission of Tomato Leaf Curl New Delhi Begomovirus Infecting Sponge Gourd in India. Phytoparasitica 2013, 41, 591–592. [Google Scholar] [CrossRef]
- Usharani, K.S.; Surendranath, B.; Paul-Khurana, S.M.; Garg, I.D.; Malathi, V.G. Potato Leaf Curl—A New Disease of Potato in Northern India Caused by a Strain of Tomato Leaf Curl New Delhi Virus. Plant Pathol. 2004, 53, 235. [Google Scholar] [CrossRef]
- Renukadevi, P.; Sangeetha, B.; Malathi, V.G.; Nakkeeran, S.; Satya, V.K. Enigmatic Emergence of Seed Transmission of Geminiviruses. In Geminivirus: Detection, Diagnosis and Management; Gaur, R.K., Pradeep, S., Czosnek, H., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 285–306. ISBN 978-0-323-90587-9. [Google Scholar]
- Kim, J.; Kil, E.-J.; Kim, S.; Seo, H.; Byun, H.-S.; Park, J.; Chung, M.-N.; Kwak, H.-R.; Kim, M.-K.; Kim, C.-S.; et al. Seed Transmission of Sweet Potato Leaf Curl Virus in Sweet Potato (Ipomoea Batatas). Plant Pathol. 2015, 64, 1284–1291. [Google Scholar] [CrossRef]
- Kothandaraman, S.V.; Devadason, A.; Ganesan, M.V. Seed-Borne Nature of a Begomovirus, Mung Bean Yellow Mosaic Virus in Black Gram. Appl. Microbiol. Biotechnol. 2016, 100, 1925–1933. [Google Scholar] [CrossRef] [PubMed]
- Anabestani, A.; Behjatnia, S.A.A.; Izadpanah, K.; Tabein, S.; Accotto, G.P. Seed Transmission of Beet Curly Top Virus and Beet Curly Top Iran Virus in a Local Cultivar of Petunia in Iran. Viruses 2017, 9, 299. [Google Scholar] [CrossRef] [PubMed]
- Kil, E.-J.; Kim, S.; Lee, Y.-J.; Byun, H.-S.; Park, J.; Seo, H.; Kim, C.-S.; Shim, J.-K.; Lee, J.-H.; Kim, J.-K.; et al. Tomato Yellow Leaf Curl Virus (TYLCV-IL): A Seed-Transmissible Geminivirus in Tomatoes. Sci. Rep. 2016, 6, 19013. [Google Scholar] [CrossRef] [PubMed]
- Kil, E.J.; Park, J.; Choi, H.S.; Kim, C.S.; Lee, S. Seed Transmission of Tomato Yellow Leaf Curl Virus in White Soybean (Glycine Max). Plant Pathol. J. 2017, 33, 424–428. [Google Scholar] [CrossRef]
- Kil, E.-J.; Park, J.; Choi, E.-Y.; Byun, H.-S.; Lee, K.-Y.; An, C.G.; Lee, J.-H.; Lee, G.-S.; Choi, H.-S.; Kim, C.-S.; et al. Seed Transmission of Tomato Yellow Leaf Curl Virus in Sweet Pepper (Capsicum Annuum). Eur. J. Plant Pathol. 2018, 150, 759–764. [Google Scholar] [CrossRef]
- Sangeetha, B.; Malathi, V.G.; Alice, D.; Suganthy, M.; Renukadevi, P. A Distinct Seed-Transmissible Strain of Tomato Leaf Curl New Delhi Virus Infecting Chayote in India. Virus Res. 2018, 258, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-H.; Gustian, D.; Chang, C.-J.; Jan, F.-J. Seed and Pollen Transmission of Tomato Leaf Curl New Delhi Virus, Tomato Leaf Curl Taiwan Virus, and Tomato Yellow Leaf Curl Thailand Virus in Cucumbers and Tomatoes. Plant Dis. 2023, 107, 2002–2008. [Google Scholar] [CrossRef] [PubMed]
- Gomathi Devi, R.; Jothika, C.; Sankari, A.; Lakshmi, S.; Malathi, V.G.; Renukadevi, P. Seed Transmission of Begomoviruses: A Potential Threat for Bitter Gourd Cultivation. Plants 2023, 12, 1396. [Google Scholar] [CrossRef]
- Krishnan, N.; Kumari, S.; Chaubey, T.; Kumar, R.V.; Chinnappa, M.; Dubey, V.; Pandey, K.K.; Singh, J.; Singh, A.K. Study on Mosaic Disease of Sponge Gourd (Luffa Cylindrica L.) Caused by Tomato Leaf Curl New Delhi Virus. J. Plant Pathol. 2023, 105, 573–580. [Google Scholar] [CrossRef]
- Kil, E.-J.; Vo, T.T.B.; Fadhila, C.; Ho, P.T.; Lal, A.; Troiano, E.; Parrella, G.; Lee, S. Seed Transmission of Tomato Leaf Curl New Delhi Virus from Zucchini Squash in Italy. Plants 2020, 9, 563. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Díaz, T.; Zhang, D.; Lozano-Durán, R. No Evidence of Seed Transmissibility of Tomato Yellow Leaf Curl Virus in Nicotiana Benthamiana. J. Zhejiang Univ.-Science B 2017, 18, 437–440. [Google Scholar] [CrossRef]
- Pérez-Padilla, V.; Fortes, I.M.; Romero-Rodríguez, B.; Arroyo-Mateos, M.; Castillo, A.G.; Moyano, C.; De León, L.; Moriones, E. Revisiting Seed Transmission of the Type Strain of Tomato Yellow Leaf Curl Virus in Tomato Plants. Phytopathology 2020, 110, 121–129. [Google Scholar] [CrossRef]
- Tabein, S.; Miozzi, L.; Matić, S.; Accotto, G.P.; Noris, E. No Evidence for Seed Transmission of Tomato Yellow Leaf Curl Sardinia Virus in Tomato. Cells 2021, 10, 1673. [Google Scholar] [CrossRef]
- Andreason, S.A.; Olaniyi, O.G.; Gilliard, A.C.; Wadl, P.A.; Williams, L.H.; Jackson, D.M.; Simmons, A.M.; Ling, K.-S. Large-Scale Seedling Grow-Out Experiments Do Not Support Seed Transmission of Sweet Potato Leaf Curl Virus in Sweet Potato. Plants 2021, 10, 139. [Google Scholar] [CrossRef] [PubMed]
- Fortes, I.M.; Pérez-Padilla, V.; Romero-Rodríguez, B.; Fernández-Muñoz, R.; Moyano, C.; Castillo, A.G.; De León, L.; Moriones, E. Begomovirus Tomato Leaf Curl New Delhi Virus Is Seedborne but Not Seed Transmitted in Melon. Plant Dis. 2023, 107, 473–479. [Google Scholar] [CrossRef]
- Bragard, C.; Dehnen-Schmutz, K.; Di Serio, F.; Gonthier, P.; Jacques, M.-A.; Miret, J.A.J.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; Milonas, P.; et al. Pest Categorisation of Tomato Leaf Curl New Delhi Virus. EFSA J. 2020, 18, 6179. [Google Scholar] [CrossRef]
- Martín-Hernández, A.M.; Picó, B. Natural Resistances to Viruses in Cucurbits. Agronomy 2021, 11, 23. [Google Scholar] [CrossRef]
- Sáez, C.; Martínez, C.; Ferriol, M.; Manzano, S.; Velasco, L.; Jamilena, M.; López, C.; Picó, B. Resistance to Tomato Leaf Curl New Delhi Virus in Cucurbita spp. Ann. Appl. Biol. 2016, 169, 91–105. [Google Scholar] [CrossRef]
- Dombrovsky, A.; Smith, E. Seed Transmission of Tobamoviruses: Aspects of Global Disease Distribution. In Advances in Seed Biology; Jimenez-Lopez, J.C., Ed.; InTechOpen: London, UK, 2017; pp. 233–260. ISBN 978-953-51-3622-4. [Google Scholar]
- Montes, N.; Pagán, I. Light Intensity Modulates the Efficiency of Virus Seed Transmission through Modifications of Plant Tolerance. Plants 2019, 8, 304. [Google Scholar] [CrossRef]
- Doyle, J. DNA Protocols for Plants. In Molecular Techniques in Taxonomy; Hewitt, G.M., Johnston, A.W.B., Young, J.P.W., Eds.; Springer: Berlin/Heidelberg, Germany, 1991; Volume 57, pp. 283–293. ISBN 978-3-642-83962-7. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative C T Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Sáez, C.; Ambrosio, L.G.M.; Miguel, S.M.; Valcárcel, J.V.; Díez, M.J.; Picó, B.; López, C. Resistant Sources and Genetic Control of Resistance to ToLCNDV in Cucumber. Microorganisms 2021, 9, 913. [Google Scholar] [CrossRef]
- Hohn, T. Plant Virus Transmission from the Insect Point of View. Proc. Natl. Acad. Sci. USA 2007, 104, 17905–17906. [Google Scholar] [CrossRef]
- Sastry, K.S. Seed-Borne Plant Virus Diseases; Springer Science & Business Media, Ed.; Springer: New Delhi, India, 2013; ISBN 978-81-322-0812-9. [Google Scholar]
- Kumar, P.L.; Cuervo, M.; Kreuze, J.F.; Muller, G.; Kulkarni, G.; Kumari, S.G.; Massart, S.; Mezzalama, M.; Alakonya, A.; Muchugi, A.; et al. Phytosanitary Interventions for Safe Global Germplasm Exchange and the Prevention of Transboundary Pest Spread: The Role of CGIAR Germplasm Health Units. Plants 2021, 10, 328. [Google Scholar] [CrossRef]
- Chalam, V.C.; Sharma, V.D.; Sharma, R.; Maurya, A.K. Modern Technologies for the Diagnosis and Assay of Plants Viruses. In Applied Plant Virology; Awasthi, L.P., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 69–77. ISBN 978-0-12-818654-1. [Google Scholar]
- Pagán, I. Movement Between Plants: Vertical Transmission. In Cucumber Mosaic Virus; Palukaitis, P., García-Arenal., F., Eds.; APS Publications: St. Paul, MN, USA, 2019; pp. 185–198. ISBN 978-0-89054-610-9. [Google Scholar]
- Pagán, I. Transmission through Seeds: The Unknown Life of Plant Viruses. PLoS Pathog. 2022, 18, e1010707. [Google Scholar] [CrossRef]
- Just, K.; Sattar, M.N.; Arif, U.; Luik, A.; Kvarnheden, A. Infectivity of Tomato Yellow Leaf Curl Virus Isolated from Imported Tomato Fruit in Estonia. Zemdirb.-Agric. 2017, 104, 47–52. [Google Scholar] [CrossRef]
- Suruthi, V.; Nakkeeran, S.; Renukadevi, P.; Malathi, V.G.; Rajasree, V. Evidence of Seed Transmission of Dolichos Yellow Mosaic Virus, a Begomovirus Infecting Lablab-Bean in India. Virus Dis. 2018, 29, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Shahmohammadi, N.; Mansourpour, M.; Golnaraghi, A. Current Challenges and Future Perspectives on Detection of Geminiviruses. In Geminivirus: Detection, Diagnosis and Management; Gaur, R.K., Pradeep, S., Czosnek, H., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 3–24. ISBN 978-0-323-90587-9. [Google Scholar]
- Liu, Y.; Li, X.; Zhao, J.; Tang, X.; Tian, S.; Chen, J.; Shi, C.; Wang, W.; Zhang, L.; Feng, X.; et al. Direct Evidence That Suspensor Cells Have Embryogenic Potential That Is Suppressed by the Embryo Proper during Normal Embryogenesis. Proc. Natl. Acad. Sci. USA 2015, 112, 12432–12437. [Google Scholar] [CrossRef]
- Shargil, D.; Zemach, H.; Belausov, E.; Lachman, O.; Luria, N.; Molad, O.; Smith, E.; Kamenetsky, R.; Dombrovsky, A. Insights into the Maternal Pathway for Cucumber Green Mottle Mosaic Virus Infection of Cucurbit Seeds. Protoplasma 2019, 256, 1109–1118. [Google Scholar] [CrossRef]
- Yamamoto, H.; Wakita, Y.; Kitaoka, T.; Fujishiro, K.; Kesumawati, E.; Koeda, S. Southeast Asian Isolate of the Tomato Leaf Curl New Delhi Virus Shows Higher Pathogenicity Against Tomato and Cucurbit Crops Compared to That of the Mediterranean Isolate. Hort. J. 2021, 90, 314–325. [Google Scholar] [CrossRef]
- Janssen, D.; Simón, A.; Boulares, M.; Ruiz, L. Host Species-Dependent Transmission of Tomato Leaf Curl New Delhi Virus-ES by Bemisia Tabaci. Plants 2022, 11, 390. [Google Scholar] [CrossRef] [PubMed]
- Farina, A.; Rapisarda, C.; Fiallo-Olivé, E.; Navas-Castillo, J. Tomato Leaf Curl New Delhi Virus Spain Strain Is Not Transmitted by Trialeurodes Vaporariorum and Is Inefficiently Transmitted by Bemisia Tabaci Mediterranean between Zucchini and the Wild Cucurbit Ecballium Elaterium. Insects 2023, 14, 384. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.D.; Logsdon, J.M.; Kelley, S.E. An Empirical Study of the Evolution of Virulence under Both Horizontal and Vertical Transmission. Evolution 2005, 59, 730–739. [Google Scholar] [CrossRef]
- Hamelin, F.M.; Allen, L.J.S.; Prendeville, H.R.; Hajimorad, M.R.; Jeger, M.J. The Evolution of Plant Virus Transmission Pathways. J. Theor. Biol. 2016, 396, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Behera, P.R.; Mishra, M.K.; Kumar, S.; Senapati, A.K. Mechanical and Seed Transmission Studies of Yellow Mosaic Virus Disease in Greengram. Pharma Innov. J. 2022, 11, 2153–2160. [Google Scholar]
- Lee, C.-H.; Zheng, Y.-X.; Chan, C.-H.; Ku, H.-M.; Chang, C.-J.; Jan, F.-J. A Single Amino Acid Substitution in the Movement Protein Enables the Mechanical Transmission of a Geminivirus. Mol. Plant Pathol. 2020, 21, 571–588. [Google Scholar] [CrossRef]
- Ward, C.W.; Shukla, D.D. Taxonomy of Potyviruses: Current Problems and Some Solutions. Intervirology 1991, 32, 269–296. [Google Scholar] [CrossRef] [PubMed]
- Domingo, E.; Escarmís, C.; Sevilla, N.; Moya, A.; Elena, S.F.; Quer, J.; Novella, I.S.; Holland, J.J. Basic Concepts in RNA Virus Evolution. FASEB J. 1996, 10, 859–864. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Species | Subspecies | Group | Number of Evaluated Genotypes | Number of Genotypes with Fruit |
|---|---|---|---|---|---|
| Cucumis | melo | melo | inodorus | 18 | 15 |
| cantalupensis | 3 | 1 | |||
| flexuosus | 3 | 1 | |||
| Cucurbita | moschata | 3 | 3 |
| C. melo subsp. melo | C. moschata | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Inodorus Group | Cant. | Flex. | |||||||||||||||||||
| Sample | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 1 | 1 | 1 | 2 | 3 | |
| Leaves | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | |
| Flowers | Petals ♀ | + | + | + | + | + | + | + | + | + | nd | + | + | + | + | + | + | + | + | nd | + |
| Ovary–pistils | + | + | + | + | + | + | + | + | + | nd | + | + | + | + | + | + | + | − | − | + | |
| Petals ♂ | + | + | + | − | + | + | + | + | + | nd | + | nd | + | + | nd | + | + | + | − | + | |
| Anthers | + | + | + | + | + | + | + | + | + | nd | + | nd | + | + | + | + | − | + | + | + | |
| Seeds | Coat | + | + | + | + | + | + | + | + | + | + | + | + | + | + | nd | nd | nd | + | + | + |
| Endosperm and embryo | + | − | − | + | − | + | − | + | + | + | + | − | + | + | + | − | + | − | − | − | |
| Species | Type | Varieties Analyzed | Varieties Positive for ToLCNDV | Positive Seeds/ Analyzed Seeds | Offspring Seedlings Positive for ToLCNDV |
|---|---|---|---|---|---|
| Watermelon (Citrullus lanatus) | Mini | 1 | - | ||
| Black | 10 | - | |||
| Striped | 19 | - | |||
| Cucumber (Cucumis sativus) | Short | 10 | - | ||
| Long | 33 | 1 | 1/5 | - | |
| Melon (Cucumis melo) | Blanco | 1 | - | ||
| Amarillo | 1 | - | |||
| Cantalupo | 2 | - | |||
| Galia | 8 | - | |||
| Piel de Sapo | 5 | 1 | 5/5 | 1 | |
| Zucchini (Cucurbita pepo) | Yellow | 1 | - | ||
| White | 1 | - | |||
| Round | 1 | 1 | 1/5 | - | |
| Dark green | 7 | - | |||
| Edium green | 9 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sáez, C.; Kheireddine, A.; García, A.; Sifres, A.; Moreno, A.; Font-San-Ambrosio, M.I.; Picó, B.; López, C. Further Molecular Diagnosis Determines Lack of Evidence for Real Seed Transmission of Tomato Leaf Curl New Delhi Virus in Cucurbits. Plants 2023, 12, 3773. https://doi.org/10.3390/plants12213773
Sáez C, Kheireddine A, García A, Sifres A, Moreno A, Font-San-Ambrosio MI, Picó B, López C. Further Molecular Diagnosis Determines Lack of Evidence for Real Seed Transmission of Tomato Leaf Curl New Delhi Virus in Cucurbits. Plants. 2023; 12(21):3773. https://doi.org/10.3390/plants12213773
Chicago/Turabian StyleSáez, Cristina, Amina Kheireddine, Arcadio García, Alicia Sifres, Alejandro Moreno, María Isabel Font-San-Ambrosio, Belén Picó, and Carmelo López. 2023. "Further Molecular Diagnosis Determines Lack of Evidence for Real Seed Transmission of Tomato Leaf Curl New Delhi Virus in Cucurbits" Plants 12, no. 21: 3773. https://doi.org/10.3390/plants12213773
APA StyleSáez, C., Kheireddine, A., García, A., Sifres, A., Moreno, A., Font-San-Ambrosio, M. I., Picó, B., & López, C. (2023). Further Molecular Diagnosis Determines Lack of Evidence for Real Seed Transmission of Tomato Leaf Curl New Delhi Virus in Cucurbits. Plants, 12(21), 3773. https://doi.org/10.3390/plants12213773