
Content and Yield of L-DOPA and Bioactive Compounds of Broad Bean Plants: Antioxidant and Anti-Inflammatory Activity In Vitro

 ,
,  , ,
, ,  and
and 
Abstract
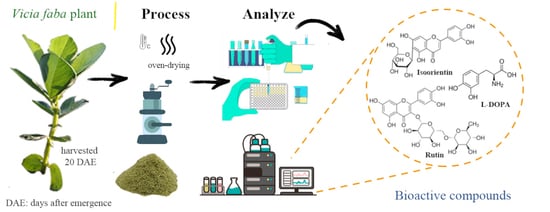
:
1. Introduction
2. Results and Discussion
2.1. Bioactive Compounds
2.1.1. Total Phenolic Compounds and Total Flavonoids
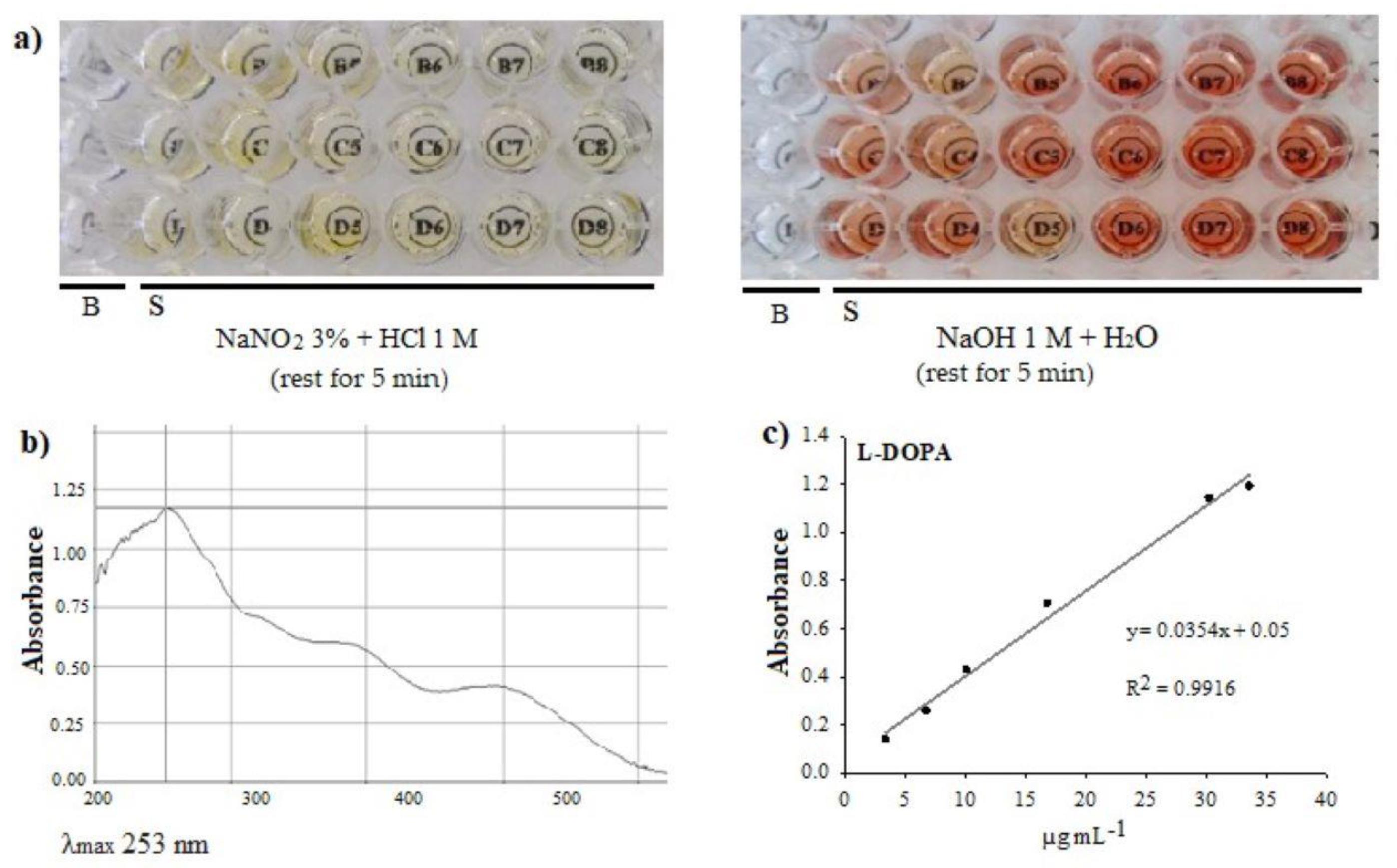
2.1.2. L-DOPA
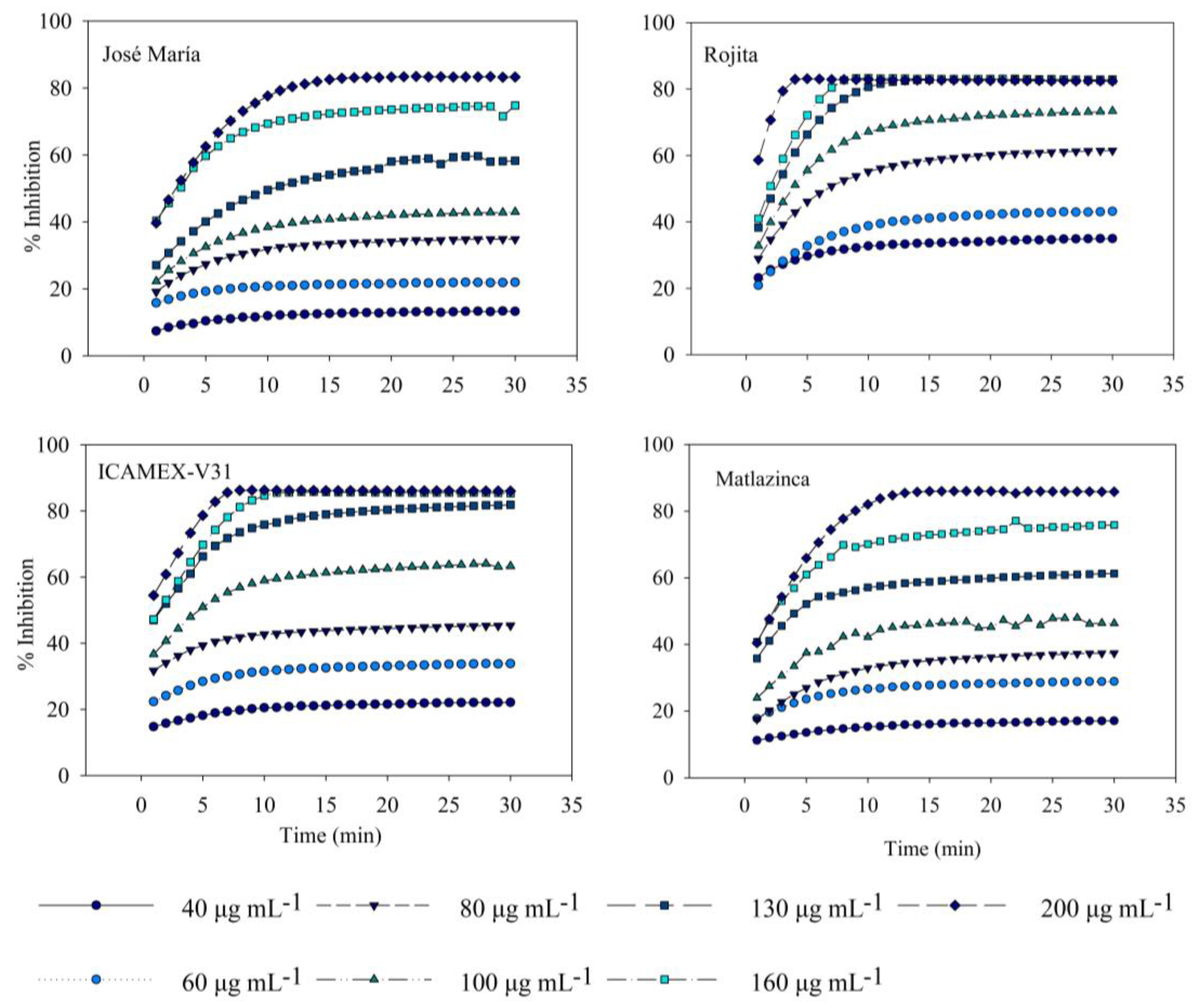
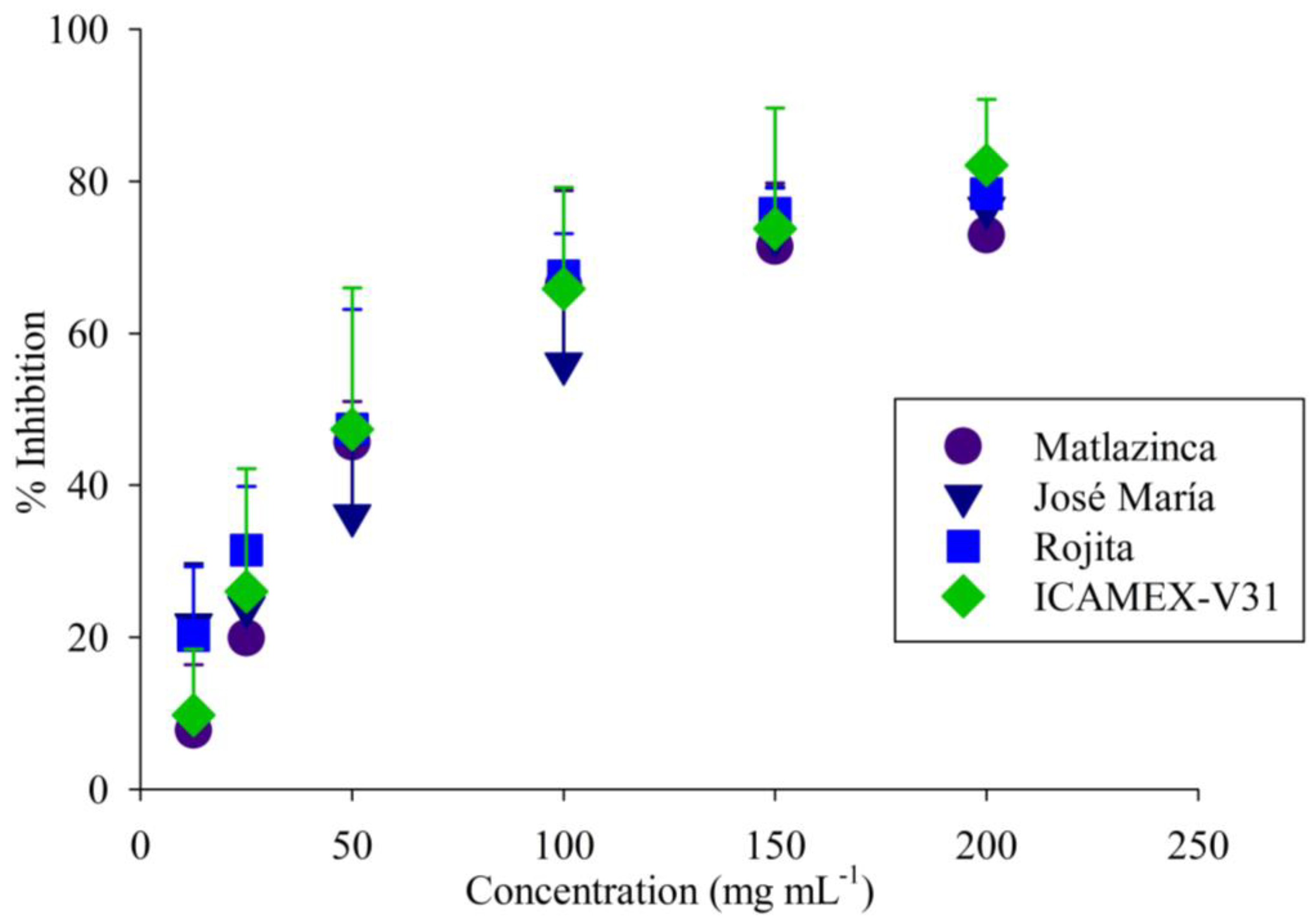
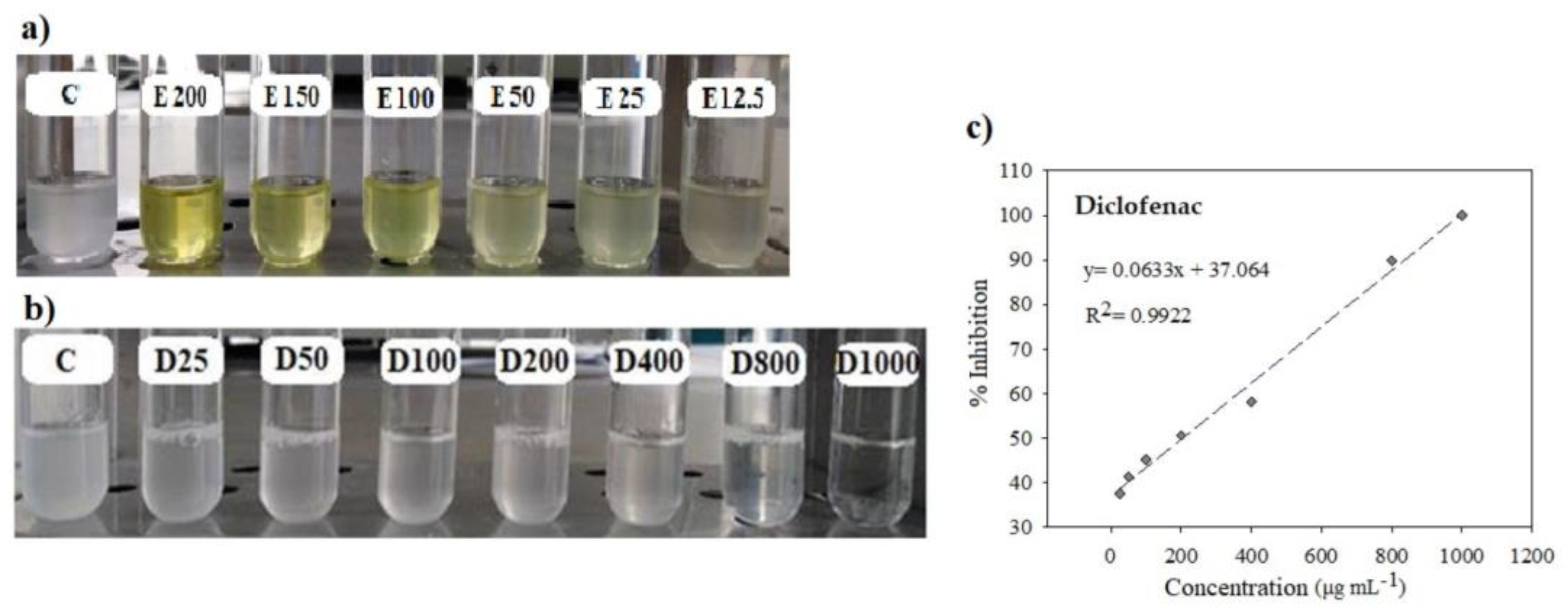
2.1.3. In Vitro Antioxidant and Anti-Inflammatory Activity
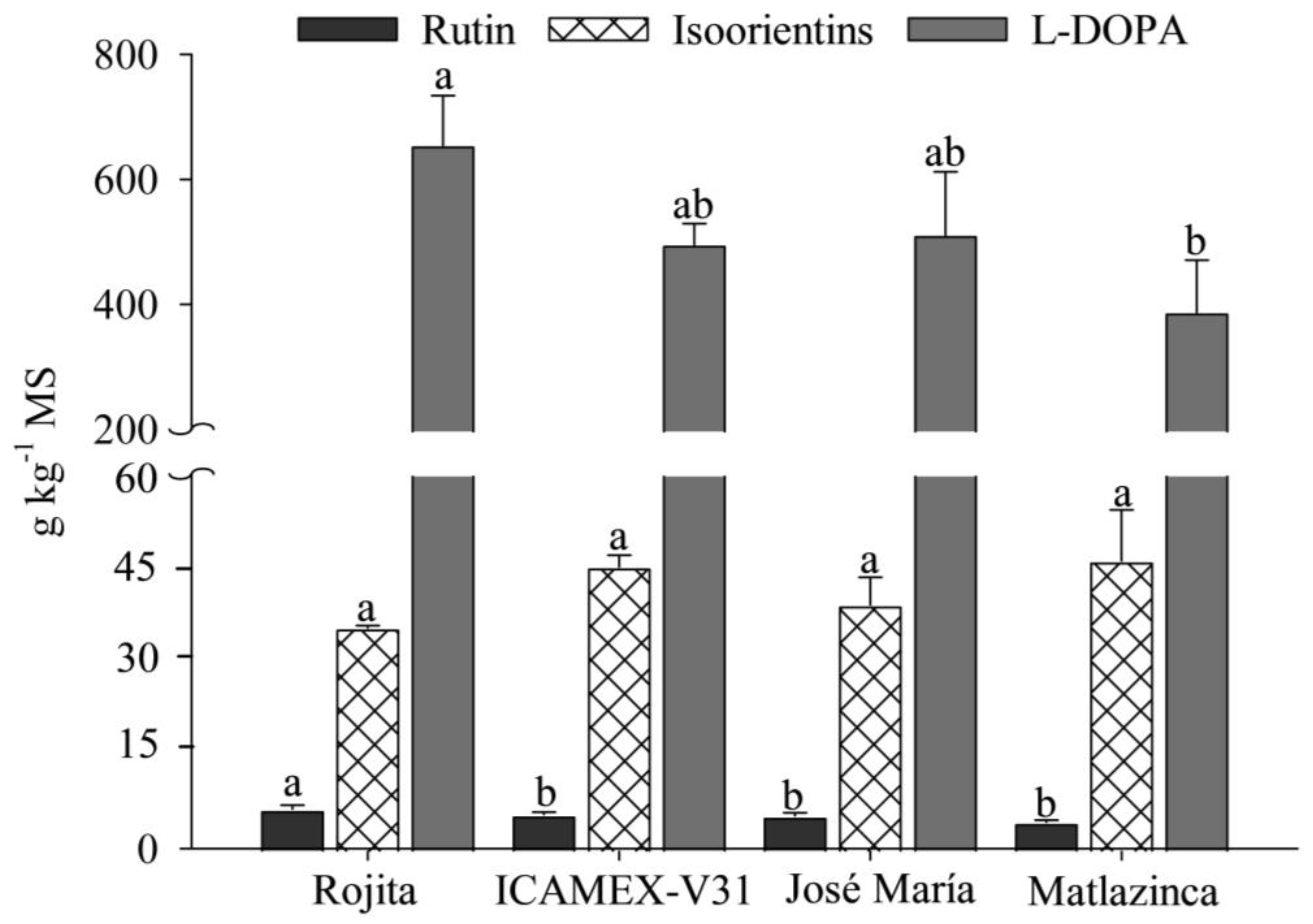
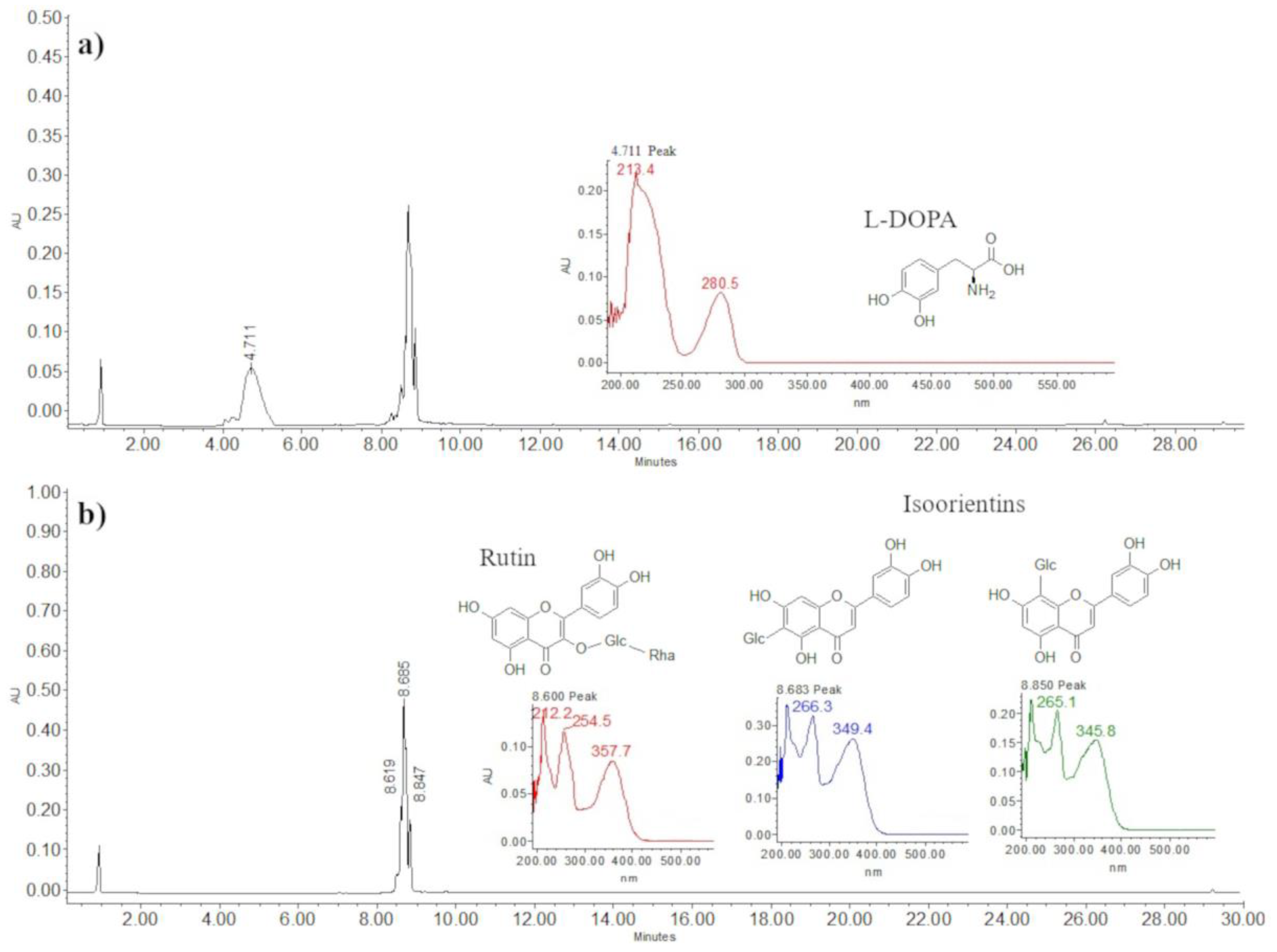
2.2. Bioactive Compounds Determined by HPLC: Concentration of L-DOPA, Rutin, and Isoorientins
2.3. Yield of Bioactive Compounds
2.4. The Broad Bean Plant as a Source of L-DOPA for Parkinson’s Disease Patients
3. Materials and Methods
3.1. Plant Material
3.2. Identification, Collection, and Processing of the Plant Material
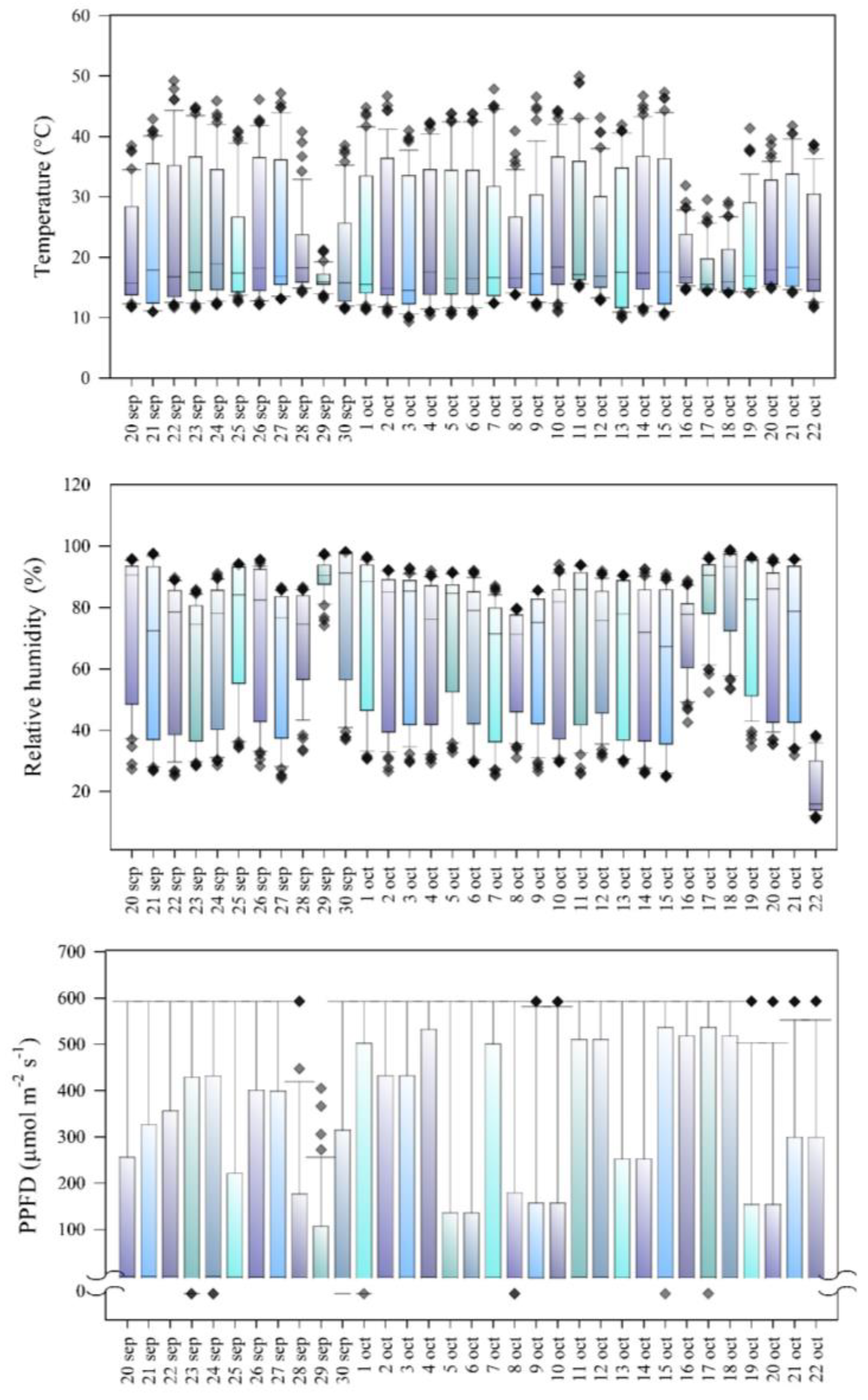
3.3. Agri-Environmental Variables
3.4. Analysis of Total Phenolic Compounds and Total Flavonoids
3.5. L-DOPA Analysis by Visible UV Spectrophotometry
3.6. Antioxidant Activity
- I%: Inhibition percentage;
- AB: Absorbance of the blank;
- AS: Absorbance of the sample.
3.7. In Vitro Assay of Anti-Inflammatory Activity
- I: Inhibition;
- ANC: Absorbance of the negative control;
- AS: Absorbance of the sample.
3.8. L-DOPA, Rutin, and Isoorientins by HPLC Analysis
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lim, T.K. Vicia faba. In Edible Medicinal and Non-Medicinal Plants; Springer: Dordrecht, The Netherlands, 2012; pp. 925–936. [Google Scholar]
- Prabhu, S.; Rajeswari, D. Nutritional and biological properties of Vicia faba L.: A perspective review. Int. Food Res. J. 2018, 25, 332–1340. [Google Scholar]
- Fuentes-Herrera, P.; Delgado-Alvarado, A.; Herrera-Cabrera, B.; Tornero-Campante, M.; Arévalo-Galarza, M.d.L.; Martínez-Ayala, A.; Barrera-Rodríguez, A. Effect of processing methods on the content of phenolic compounds in Vicia faba L. tissues grown in field and greenhouse. Bioagro 2022, 34, 221–232. [Google Scholar] [CrossRef]
- Etemadi, F.; Hashemi, M.; Autio, W.R.; Mangan, F.X.; Zandvakili, O. Yield and accumulation trend of biomass and L-DOPA in different parts of eight faba bean cultivars. Crop Sci. 2018, 58, 2020–2028. [Google Scholar] [CrossRef]
- Etemadi, F.; Hashemi, M.; Randhir, R.; ZandVakili, O.; Ebadi, A. Accumulation of L-DOPA in various organs of faba bean and influence of drought, nitrogen stress, and processing methods on L-DOPA yield. Crop J. 2018, 6, 426–434. [Google Scholar] [CrossRef]
- Vered, Y.; Rabey, J.M.; Palevitch, D.; Grosskopf, I.; Harsat, A.; Yanowski, A.; Shabtai, H.; Graff, E. Bioavailability of levodopa after consumption of Vicia faba seedlings by Parkinsonian patients and control subjects. Clin. Neuropharmacol. 2012, 17, 138–146. [Google Scholar] [CrossRef]
- Shetty, K.; Randhir, R.; Sarkar, D. Bioprocessing strategies to enhance L-DOPA and phenolic bioactives in the fava bean (Vicia faba). In Functional Foods and Biotechnology: Sources of Functional Foods and Ingredients, 1st ed.; Shetty, K., Sarkar, D., Eds.; Food biotechnology series; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2020; pp. 99–110. [Google Scholar]
- Patil, S.A.; Apine, O.A.; Surwase, S.N.; Jadhav, J.P. Biological sources of L-DOPA: An alternative approach. Adv. Park. Dis. 2013, 2, 81–87. [Google Scholar] [CrossRef]
- Rabey, J.M.; Vered, Y.; Shabtai, H.; Graff, E.; Korczyn, A.D. Improvement of Parkinsonian features correlate with high plasma levodopa values after broad bean (Vicia faba) consumption. J. Neurol. Neurosurg. Psychiatry 1992, 55, 725–727. [Google Scholar] [CrossRef]
- Rasool, H.B.A. Medicinal Plants (Importance and Uses). Pharm. Anal. Acta 2012, 3, e139. [Google Scholar] [CrossRef]
- Akinyemi, O.; Oyewole, S.O.; Jimoh, K.A. Medicinal plants and sustainable human health: A review. Horticult. Int. J. 2018, 2, 194–195. [Google Scholar]
- Organización Mundial de la Salud. Enfermedad de Parkinson. Available online: https://www.who.int/es/news-room/fact-sheets/detail/parkinson-disease (accessed on 2 August 2023).
- Etemadi, F.; Hashemi, M.; Shureshjani, R.A.; Autio, W.R. Application of data envelopment analysis to assess performance efficiency of eight faba bean varieties. Agron. J. 2017, 109, 1225–1231. [Google Scholar] [CrossRef]
- Ortiz, L.M.; Delgado-Alvarado, A.; Herrera-Cabrera, B.E.; Árevalo-Galarza, M.A.d.L.; Barrera-Rodríguez, A.I. Efecto de dos métodos de secado en los compuestos fenólicos totales, L-DOPA y la actividad antioxidante de Vicia faba L. Nova Sci. 2019, 11, 198–219. [Google Scholar] [CrossRef]
- Kirakosyan, A.; Kaufman, P.B.; Duke, J.A.; Warber, S.; Bolling, S. The production of L-dopa and isoflavones in seeds and seedlings of different cultivars of Vicia faba L. (fava bean). Evid.-Based Integr. Med. 2004, 1, 131–135. [Google Scholar] [CrossRef]
- Fuentes-Herrera, P.B.; Delgado-Alvarado, A.; Herrera, B.E.; Luna-Guevara, M.L.; Olvera-Hernández, J.I. Quantification of isoflavones in stems of faba bean (Vicia faba L.). Rev. Fac. Cienc. Agrar. 2020, 52, 43–51. [Google Scholar]
- Okumura, K.; Hosoya, T.; Kawarazaki, K.; Izawa, N.; Kumazawa, S. Antioxidant activity of phenolic compounds from fava bean sprouts. J. Food Sci. 2016, 81, C1394–C1398. [Google Scholar] [CrossRef]
- Boudjou, S.; Oomah, B.D.; Zaidi, F.; Hosseinian, F. Phenolics content and antioxidant and anti-inflammatory activities of legume fractions. Food Chem. 2013, 138, 1543–1550. [Google Scholar] [CrossRef]
- Valente, I.M.; Cabrita, A.R.J.; Malushi, N.; Oliveira, H.M.; Papa, L.; Rodrigues, J.A.; Fonseca, A.J.M.; Maia, M.R.G. Unravelling the phytonutrients and antioxidant properties of European Vicia faba L. Seeds. Food Res. Int. 2019, 116, 888–896. [Google Scholar] [CrossRef]
- Barros, L.; Dueñas, M.; Carvalho, A.M.; Ferreira, I.C.F.R.; Santos-Buelga, C. Characterization of phenolic compounds in flowers of wild medicinal plants from northeastern Portugal. Food Chem. Toxicol. 2012, 50, 1576–1582. [Google Scholar] [CrossRef]
- Sulaiman, C.; Balachandran, I. Total phenolics and total flavonoids in selected Indian medicinal Plants. Indian J. Pharm. Sci. 2012, 74, 258–260. [Google Scholar] [CrossRef]
- Duan, S.; Kwon, S.J.; Lim, Y.J.; Gil, C.S.; Jin, C.; Eom, S.H. L-3,4-Dihydroxyphenylalanine accumulation in faba bean (Vicia faba L.) tissues during different growth stages. Agronomy 2021, 11, 502. [Google Scholar] [CrossRef]
- Boukhanouf, S.; Louaileche, H.; Perrin, D. Phytochemical content and in vitro antioxidant activity of faba bean (Vicia faba L.) as affected by maturity stage and cooking practice. Int. Food Res. J. 2016, 23, 954–961. [Google Scholar]
- Clària, J. Los nuevos antiinflamatorios. Med. Integral 2001, 38, 175–183. [Google Scholar]
- Sakat, S.S.; Juvekar, A.R.; Gambhire, M.N. In vitro antioxidant and anti inflammatory activity of methanol extract of Oxalis corniculata Linn. Int. J. Pharm. Pharm. Sci. 2012, 2, 146–155. [Google Scholar]
- Anoop, M.V.; Bindu, A.R. In-vitro anti-inflammatory activity studies on Syzygium zeylanicum (L.) DC Leaves. Int. J. Pharm. Sci. Rev. Res. 2015, 4, 18–27. [Google Scholar]
- Ghorbani, A. Mechanisms of antidiabetic effects of flavonoid rutin. Biomed. Pharmacother. 2017, 96, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Ali, A.; Ali, J.; Sahni, J.K.; Baboota, S. Rutin: Therapeutic potential and recent advances in drug delivery. Expert Opin. Investig. Drugs 2013, 22, 1063–1079. [Google Scholar] [CrossRef]
- Gullón, B.; Lú-Chau, T.A.; Moreira, M.T.; Lema, J.M.; Eibes, G. Rutin: A review on extraction, identification and purification methods, biological activities and approaches to enhance its bioavailability. Trends Food Sci. Technol. 2017, 67, 220–235. [Google Scholar] [CrossRef]
- Hao, G.; Dong, Y.; Huo, R.; Wen, K.; Zhang, Y.; Liang, G. Rutin inhibits neuroinflammation and provides neuroprotection in an experimental rat model of subarachnoid hemorrhage, possibly through suppressing the RAGE–NF-κB inflammatory signaling pathway. Neurochem. Res. 2016, 41, 1496–1504. [Google Scholar] [CrossRef]
- Ziqubu, K.; Dludla, P.V.; Joubert, E.; Muller, C.J.F.; Louw, J.; Tiano, L.; Nkambule, B.B.; Kappo, A.P.; Mazibuko-Mbeje, S.E. Isoorientin: A dietary flavone with the potential to ameliorate diverse metabolic complications. Pharmacol. Res. 2020, 158, 104867. [Google Scholar] [CrossRef]
- Anilkumar, K.; Reddy, G.V.; Azad, R.; Yarla, N.S.; Dharmapuri, G.; Srivastava, A.; Kamal, M.A.; Pallu, R. Evaluation of anti-inflammatory properties of isoorientin isolated from tubers of Pueraria tuberosa. Oxid. Med. Cell. Longev. 2017, 2017, 5498054. [Google Scholar] [CrossRef]
- Siqueira-Soares, R.d.C.; Soares, A.R.; Parizotto, A.V.; Ferrarese, M.d.L.L.; Ferrarese-Filho, O. Root growth and enzymes related to the lignification of maize seedlings exposed to the allelochemical L-DOPA. Sci. World J. 2013, 2013, 134237. [Google Scholar] [CrossRef] [PubMed]
- Nagatsu, T.; Sawada, M. L-Dopa therapy for Parkinson’s disease: Past, present, and future. Park. Relat. Disord. 2009, 15, S3–S8. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.R.; Marchiosi, R.; Siqueira-Soares, R.d.C.; Barbosa de Lima, R.; Dantas dos Santos, W.; Ferrarese-Filho, O. The role of L-DOPA in plants. Plant Signal. Behav. 2014, 9, e28275. [Google Scholar] [CrossRef] [PubMed]
- Finefield, J.M.; Sherman, D.H.; Kreitman, M.; Williams, R.M. Enantiomeric Natural Products: Occurrence and Biogenesis. Angew. Chem. Int. Ed. Engl. 2012, 51, 4802–4836. [Google Scholar] [CrossRef]
- Hasegawa, T.; Takahashi, K.; Fukiwake, T.; Saijo, M.; Motoki, Y. Enantiomeric Determination of DOPA in Dietary Supplements Containing Mucuna pruriens by Liquid Chromatography/Mass Spectrometry. Food Hyg. Saf. Sci. 2013, 54, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Valadez-Barba, V.; Juárez-Navarro, K.; Padilla-Camberos, E.; Díaz, N.F.; Guerra-Mora, J.R.; Díaz-Martínez, N.E. Enfermedad de Parkinson: Actualización de estudios preclínicos con el uso de células troncales pluripotentes inducidas. Neurología 2021, 1571, 681–694. [Google Scholar] [CrossRef]
- Bumgarner, N.R.; Scheerens, J.C.; Kleinhenz, M.D. Nutritional yield: A proposed index for fresh food improvement illustrated with leafy vegetable data. Plant Foods Hum. Nutr. 2012, 67, 215–222. [Google Scholar] [CrossRef]
- Magalingam, K.B.; Radhakrishnan, A.K.; Haleagrahara, N. Protective mechanisms of flavonoids in Parkinson’s disease. Oxid. Med. Cell. Longev. 2015, 2015, 314560. [Google Scholar] [CrossRef]
- Instituto Nacional de las Personas Adultas Mayores. Secretaría de Bienestar. 2019. Available online: https://www.gob.mx/inapam/es/articulos/parkinson-segunda-enfermedad-neurodegenerativa-mas-frecuente-en-personas-mayores-de-50-anos?idiom=es (accessed on 12 May 2023).
- Apaydin, H.; Ertan, S.; Özekmekçi, S. Broad bean (Vicia faba)—A natural source of L-dopa—Prolongs “on” periods in patients with Parkinson’s disease who have “on–off” fluctuations. Mov. Disord. 2000, 15, 164–166. [Google Scholar] [CrossRef]
- Cassani, E.; Cilia, R.; Laguna, J.; Barichella, M.; Contin, M.; Cereda, E.; Isaias, I.U.; Sparvoli, F.; Akpalu, A.; Budu, K.O.; et al. Mucuna pruriens for Parkinson’s disease: Low-cost preparation method, laboratory measures and pharmacokinetics profile. J. Neurol. Sci. 2016, 365, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Pugalenthi, M.; Vadivel, V. L-Dopa (L-3,4-Dihydroxyphenylalanine): A non-protein toxic amino acid in Mucuna Pruriens seeds. Food 2007, 1, 322–343. [Google Scholar]
- Pool, N.L.; León, M.N.S.; González, S.C.; Figueroa, F.P. Frijol terciopelo, cultivo de cobertura en la agriculturaChol del valle del Tulija, Chiapas, México. Terra Latinoam. 1998, 16, 359–369. [Google Scholar]
- Kala, B.K.; Mohan, V.R. Nutritional and anti-nutritional potential of three accessions of itching bean (Mucuna Pruriens (L.) DC Var. Pruriens): An under-utilized tribal pulse. Int. J. Food Sci. Nutr. 2010, 61, 497–511. [Google Scholar] [PubMed]
- Martínez-Villaluenga, C.; Frias, J.; Vidal-Valverde, C. Alpha-galactosides: Antinutritional factors or functional ingredients? Crit. Rev. Food Sci. Nutr. 2008, 48, 301–316. [Google Scholar] [CrossRef]
- Tahir, M.; Lindeboom, N.; Båga, M.; Vandenberg, A.; Chibbar, R. Composition and correlation between major seed constituents in selected lentil (Lens Culinaris. Medik) genotypes. Can. J. Plant Sci. 2011, 91, 825–835. [Google Scholar] [CrossRef]
- Vadivel, V.; Pugalenthi, M. Effect of various processing methods on the levels of antinutritional constituents and protein digestibility of Mucuna pruriens (L.) DC. var. utilis (Wall. ex Wight) Baker ex Burck (velvet bean) seeds. J. Food Biochem. 2008, 32, 795–812. [Google Scholar]
- Organización de las Naciones Unidas para la Alimentación y la Agricultura. FAOSTAT. Cultivos y Productos de Ganadería. Visualizar Datos Estadísticos. Cantidades de Producción de Habas y Haba Caballar, Secas por País 1994—2021. Available online: https://www.fao.org/faostat/es/#data/QCL/visualize (accessed on 30 June 2023).
- Herald, T.J.; Gadgil, P.; Tilley, M. High-throughput micro plate assays for screening flavonoid content and DPPH-scavenging activity in sorghum bran and flour. J. Sci. Food Agric. 2012, 92, 2326–2331. [Google Scholar] [CrossRef]
- Polanowska, K.; Łukasik, R.; Kuligowski, M.; Nowak, J. Development of a sustainable, simple, and robust method for efficient L-DOPA extraction. Molecules 2019, 24, 2325. [Google Scholar] [CrossRef] [PubMed]
- Rahmani-Nezhad, S.; Dianat, S.; Saeedi, M.; Tehrani, M.; Ghadiri, A.; Hadjiakhoondi, A. Evaluating the accumulation trend of L-DOPA in dark-germinated seeds and suspension cultures of Phaseolus vulgaris L. by an efficient uv-spectrophotometric method. Quím. Nova 2018, 41, 386–393. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Rahman, M.; Islam, B.; Biswas, M.; Khurshid, A. In vitro antioxidant and free radical scavenging activity of different parts of Tabebuia pallida growing in Bangladesh. BMC Res. Notes 2015, 8, 621. [Google Scholar] [CrossRef]
- Mosquera, O.M.; Correa, Y.M.; Buitrago, D.C.; Niño, J. Antioxidant activity of twenty five plants from colombian biodiversity. Mem. Inst. Oswaldo Cruz 2007, 102, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, Y.; Kobayashi, M. Interaction of anti-inflammatory drugs with serum proteins, especially with some biologically active proteins. J. Pharm. Pharmacol. 2011, 20, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Statistical Analysis Software, version 9.1; SAS Institute: Cary, NC, USA, 2004.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | TPC | TF | L-DOPA | DPPH (IC50) | AANTI (IC50) |
|---|---|---|---|---|---|
| (mg GAE g−1 DM) | (mg QE g−1 DM) | (mg g−1 DM) | (µg mL−1) | (mg mL−1) | |
| Rojita | 131.65 a ± 15.44 | 111.17 a ± 9.97 | 88.04 a ± 10.02 | 87.68 a ± 2.49 | 74.40 a ± 13.27 |
| Matlazinca | 135.63 a ± 14.44 | 118.58 a ± 20.14 | 79.91 ab ± 5.46 | 97.42 c ± 2.49 | 87.87 a ± 10.55 |
| ICAMEX-V31 | 127.03 ab ± 12.86 | 91.98 b ± 11.94 | 79.98 ab ± 10.55 | 94.79 ab ± 6.57 | 99.14 a ± 19.32 |
| José María | 111.82 b ± 7.26 | 92.76 b ± 10.84 | 67.30 b ± 18.55 | 106.55 d ± 5.21 | 90.22 a ± 16.47 |
| Rojita | ICAMEX-V31 | José María | Matlazinca | |||||
|---|---|---|---|---|---|---|---|---|
| Biomass (g plant−1 FM) | 15.24 a | ±0.43 | 12.57 b | ±0.24 | 16.28 a | ±0.35 | 10.58 c | ±0.81 |
| Fresh plants (g m−2 FM) | 471.82 b | ±13.60 | 434.94 b | ±53.24 | 639.59 a | ±13.92 | 407.84 b | ±51.05 |
| Biomass (g plant−1 DM) | 1.97 a | ±0.03 | 1.73 b | ±0.01 | 2.11 a | ±0.03 | 1.46 c | ±0.12 |
| Dry plants (g m−2 DM) | 61.12 b | ±1.18 | 59.71 b | ±5.97 | 83.25 a | ±1.25 | 56.55 b | ±8.24 |
| TPC | ||||||||
| g plant−1 | 0.25 a | ±0.00 | 0.21 b | ±0.00 | 0.23 ab | ±0.00 | 0.19 c | ±0.01 |
| g kg−1 | 131.65 a | ±15.43 | 127.03 c | ±13.35 | 111.77 d | ±9.10 | 131.28 b | ±12.80 |
| g m−2 | 8.04 ab | ±0.15 | 7.58 ab | ±0.75 | 9.30 a | ±0.14 | 7.42 b | ±1.08 |
| TF | ||||||||
| g plant−1 | 0.21 a | ±0.00 | 0.16 c | ±0.00 | 0.19 b | ±0.00 | 0.16 c | ±0.01 |
| g kg−1 | 111.03 b | ±11.00 | 92.44 c | ±13.78 | 92.17 d | ±13.79 | 113.57 a | ±19.22 |
| g m−2 | 6.78 ab | ±0.13 | 5.52 b | ±0.55 | 8.08 a | ±0.12 | 6.42 b | ±0.93 |
| L-DOPA | ||||||||
| g plant−1 | 0.17 a | ±0.00 | 0.13 b | ±0.00 | 0.14 b | ±0.00 | 0.11 c | ±0.00 |
| g kg−1 | 87.53 a | ±11.11 | 80.48 b | ±10.99 | 67.81 d | ±18.94 | 79.68 c | ±10.05 |
| g m−2 | 5.34 ab | ±0.10 | 4.80 ab | ±0.48 | 5.64 a | ±0.08 | 4.50 b | ±0.65 |
| Patient | |||||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| Gender | M | M | M | F | M | M | M |
| Age (years) | 85 | 69 | 62 | 52 | 48 | 40 | 56 |
| Dose of L-DOPA (mg d−1) | 300 | 400 | 300 | 475 | 600 | 300 | 500 |
| Mobile Phase | ||
|---|---|---|
| Time (min) | A (%) | B (%) |
| 0 | 100 | 0 |
| 1 | 100 | 0 |
| 2 | 100 | 0 |
| 3 | 95 | 5 |
| 4 | 95 | 5 |
| 20 | 70 | 30 |
| 21 | 50 | 50 |
| 22 | 50 | 50 |
| 23 | 50 | 50 |
| 24 | 20 | 80 |
| 25 | 20 | 80 |
| 26 | 0 | 100 |
| 27 | 0 | 100 |
| 28 | 100 | 0 |
| 30 | 100 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuentes-Herrera, P.B.; Herrera-Cabrera, B.E.; Martínez-Ayala, A.L.; Zamilpa, A.; Delgado-Alvarado, A. Content and Yield of L-DOPA and Bioactive Compounds of Broad Bean Plants: Antioxidant and Anti-Inflammatory Activity In Vitro. Plants 2023, 12, 3918. https://doi.org/10.3390/plants12233918
Fuentes-Herrera PB, Herrera-Cabrera BE, Martínez-Ayala AL, Zamilpa A, Delgado-Alvarado A. Content and Yield of L-DOPA and Bioactive Compounds of Broad Bean Plants: Antioxidant and Anti-Inflammatory Activity In Vitro. Plants. 2023; 12(23):3918. https://doi.org/10.3390/plants12233918
Chicago/Turabian StyleFuentes-Herrera, Paula Beatriz, Braulio Edgar Herrera-Cabrera, Alma Leticia Martínez-Ayala, Alejandro Zamilpa, and Adriana Delgado-Alvarado. 2023. "Content and Yield of L-DOPA and Bioactive Compounds of Broad Bean Plants: Antioxidant and Anti-Inflammatory Activity In Vitro" Plants 12, no. 23: 3918. https://doi.org/10.3390/plants12233918
APA StyleFuentes-Herrera, P. B., Herrera-Cabrera, B. E., Martínez-Ayala, A. L., Zamilpa, A., & Delgado-Alvarado, A. (2023). Content and Yield of L-DOPA and Bioactive Compounds of Broad Bean Plants: Antioxidant and Anti-Inflammatory Activity In Vitro. Plants, 12(23), 3918. https://doi.org/10.3390/plants12233918