
Micropropagation as a Tool for the Conservation of Autochthonous Sorbus Species of Czechia


Abstract
:1. Introduction
2. Results
2.1. In Vitro Shoot Proliferation
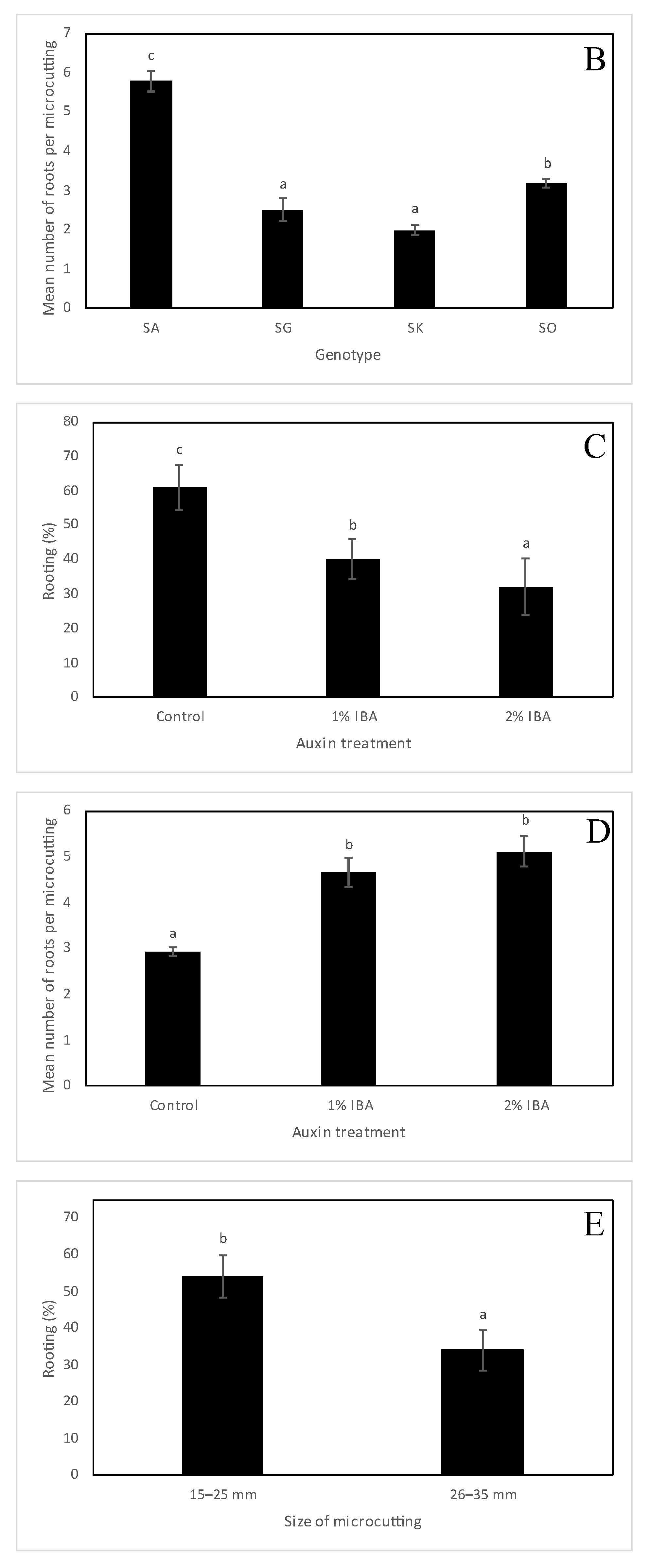
2.2. Ex Vitro Rooting and Acclimatization
3. Discussion
4. Materials and Methods
4.1. Plant Material and In Vitro Culture Initiation

4.2. In Vitro Shoot Proliferation
4.3. Ex Vitro Rooting and Acclimatization
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meyer, N.; Meierott, L.; Schuwerk, H.; Angerer, O. Beiträge zur Gattung Sorbus in Bayern. Ber. Bayer. Bot. Ges. Sonderband 2005, 5–216. [Google Scholar]
- Rich, T.C.G.; Houston, L.; Robertson, A.; Proctor, M.C.F. Whitebeams, Rowans and Service Trees of Britain and Ireland. A Monograph of British and Irish Sorbus L.; B.S.B.I. Handbook No. 14.; Botanical Society of the British Isles: London, UK, 2010; pp. 1–223. [Google Scholar]
- Grundt, H.H.; Salvesen, P.H. Kjenn din Sorbus. Rogn og asal i Norge. Rapp. Fra Skog Og Landsk. 2011, 23, 1–104. [Google Scholar]
- Lepší, M.; Lepší, P.; Koutecký, P.; Bílá, J.; Vít, P. Taxonomic revision of Sorbus subgenus Aria occurring in the Czech Republic. Preslia 2015, 87, 109–162. [Google Scholar]
- Velebil, J.; Lepší, M.; Nosková, J.; Lepší, P. Taxonomic assessment of Sorbus subgenus Aria in the Malé Karpaty Mountains. Preslia 2022, 94, 305–334. [Google Scholar] [CrossRef]
- Sennikov, A.N.; Kurtto, A. A phylogenetic checklist of Sorbus s.l. (Rosaceae) in Europe. Memo. Soc. Fauna Fl. Fenn. 2017, 93, 1–78. [Google Scholar]
- Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W. (Eds.) The Global Flora. A Practical Flora to Vascular Plant Species of the World; Special Edition, GLOVAP Nomenclature Part 1, Volume 4; Plant Gateway Ltd.: Bradford, UK, 2018; pp. 1–155. [Google Scholar]
- Kurtto, A.; Sennikov, A.N.; Lampinen, R. (Eds.) Atlas Florae Europaeae. Distribution of Vascular Plants in Europe. 17; Rosaceae (Sorbus s. lato); The Committee for Mapping the Flora of Europe & Societas Biologica Fennica Vanamo: Helsinki, Finland, 2018; pp. 1–132. [Google Scholar]
- Levin, J.; Fay, M.F.; Pellicer, J.; Hedrén, M. Multiple independent origins of intermediate species between Sorbus aucuparia and S. hybrida (Rosaceae) in the Baltic region. Nord. J. Bot. 2018, 36, 1–19. [Google Scholar] [CrossRef]
- Raimondo, F.M.; Gabrieljan, E.; Greuter, W. The genus Aria (Sorbus s. l., Rosaceae) in the Sicilian flora: Taxonomic updating, re-evaluation, description of a new species and two new combinations for one Sicilian and one SW Asian species. Bot. Chron. 2019, 22, 15–37. [Google Scholar]
- Meyer, N.; Feulner, M.; Voss, R.; Rich, T.; Gregor, T.; Paule, J. Sorbus dubronensis, eine neue endemische Art aus der Untergattung Aria (Sorbus s. l., Rosaceae) für Süddeutschland, und ihre Abgrenzung zu verwandten Arten. Ber. Bayer. Bot. Ges. 2020, 90, 83–106. [Google Scholar]
- Meyer, N.; Meierott, L. Ergänzende Beiträge zur Sorbus-Flora von Bayern. Ber. Bayer. Bot. Ges. 2021, 91, 21–48. [Google Scholar]
- Rivers, M.C.; Beech, E.; Bazos, I.; Bogunić, F.; Buira, A.; Caković, D.; Carapeto, A.; Carta, A.; Cornier, B.; Fenu, G.; et al. European Red List of Trees; IUCN: Cambridge, UK; Brussels, Belgium, 2019; pp. 1–59.
- Kaplan, Z.; Danihelka, J.; Lepší, M.; Lepší, P.; Ekrt, L.; Chrtek, J., Jr.; Kocián, J.; Prančl, J.; Kobrlová, L.; Hroneš, M.; et al. Distributions of vascular plants in the Czech Republic. Part 3. Preslia 2016, 88, 459–544. [Google Scholar]
- Kaplan, Z.; Danihelka, J.; Chrtek, J., Jr.; Kirschner, J.; Kubát, K.; Štech, M.; Štěpánek, J. (Eds.) Klíč ke Květeně České Republiky, 2nd ed.; Academia: Praha, Czech Republic, 2019; pp. 1–1168. (In Czech) [Google Scholar]
- Grulich, V. Červený seznam cévnatých rostlin ČR. Příroda 2017, 35, 75–132. (In Czech) [Google Scholar]
- Kociánová, M.; Štursová, H. Revize rozšíření a ekologie jeřábu krkonošského (Sorbus sudetica). Opera Corcon. 1986, 23, 77–110. (In Czech) [Google Scholar]
- Kovanda, M. Jeřáby (Sorbus) České republiky a jejich ochrana. Příroda 1999, 15, 31–47. (In Czech) [Google Scholar]
- Kociánová, M.; Štursová, H.; Zahradníková, J. Klonální růst endemického jeřábu Sorbus sudetica v Krkonoších. Opera Corcon. 2005, 42, 99–105. (In Czech) [Google Scholar]
- Pías, B.; Guitián, P. Breeding system and pollen limitation in the masting tree Sorbus aucuparia L. (Rosaceae) in the NW Iberian Peninsula. Acta Oecol. 2006, 29, 97–103. [Google Scholar] [CrossRef]
- Hoebee, S.E.; Arnold, U.; Düggelin, C.; Gugerli, F.; Brodbeck, S.; Rotach, P.; Holderegger, R. Mating patterns and contemporary gene flow by pollen in a large continuous and a small isolated population of the scattered forest tree Sorbus torminalis. Heredity 2007, 99, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Liljefors, A. Studies on propagation, embryology, and pollination in Sorbus. Acta Horti Bergiani 1953, 16, 277–329. [Google Scholar]
- Jankun, A.; Kovanda, M. Apomixis in Sorbus sudetica (Embryological studies in Sorbus 1). Preslia 1986, 58, 7–19. [Google Scholar]
- Jankun, A.; Kovanda, M. Apomixis and origin of Sorbus bohemica (Embryological studies in Sorbus 2). Preslia 1987, 59, 97–116. [Google Scholar]
- Dickinson, T.A.; Campbell, C.S. Population Structure and Reproductive Ecology in the Maloideae (Rosaceae). Syst. Bot. 1991, 16, 350–362. [Google Scholar] [CrossRef]
- Bärtels, A. Rozmnožování Dřevin; Státní Pedagogické Nakladatelství: Praha, Czechoslovakia, 1988; pp. 1–452. (In Czech) [Google Scholar]
- Hartmann, H.T.; Kester, D.E.; Davies, F.T.; Geneve, R.L. Hartmann and Kester’s Plant Propagation: Principles and Practices, 8th ed.; Prentice Hall: Hoboken, NJ, USA, 2011; pp. 1–915. [Google Scholar]
- Dujíčková, M.; Malá, J.; Chalupa, V. Vegetativní množení Sorbus torminalis (L.) Crantz a Sorbus domestica L. in vitro. Práce Vúlhm 1991, 77, 27–48. (In Czech) [Google Scholar]
- Malá, J.; Máchová, P.; Cvrčková, H.; Čížková, L. Využití mikropropagace pro reprodukci genových zdrojů vybraných ušlechtilých listnatých dřevin (Malus sylvestris, Pyrus pyraster, Sorbus torminalis, S. aucuparia a Prunus avium). Zprávy Lesn. Výzkumu 2005, 4, 219–224. (In Czech) [Google Scholar]
- Malá, J.; Máchová, P.; Cvrčková, H.; Karady, M.; Novák, O.; Mikulík, J.; Hauserová, E.; Greplová, J.; Strnad, M.; Doležal, K. Micropropagation of wild service tree (Sorbus torminalis (L.) Crantz): The regulative role of different aromatic cytokinins during organogenesis. J. Plant Growth Regul. 2009, 28, 341–348. [Google Scholar] [CrossRef]
- Chalupa, V. In vitro propagation of willows (Salix spp.), European mountain-ash (Sorbus aucuparia L.) and black locust (Robinia pseudoacacia L.). Biol. Plant. 1983, 25, 305–307. [Google Scholar] [CrossRef]
- Chalupa, V. Vegetativní rozmnožování listnatých dřevin řízky a metodou in vitro. Lesnictví 1987, 33, 501–510. (In Czech) [Google Scholar]
- Lall, S.; Mandegaran, Z.; Roberts, A.V. Shoot multiplication and adventitious regeneration in Sorbus aucuparia. Plant Cell Tissue Organ Cult. 2006, 85, 23–29. [Google Scholar] [CrossRef]
- Arrillaga, I.; Marzo, T.; Segura, J. Micropropagation of juvenile and adult Sorbus domestica L. Plant Cell Tissue Organ Cult. 1991, 27, 341–348. [Google Scholar] [CrossRef]
- Meier-Dinkel, A. In vitro Vermehrung von Speierling (Sorbus domestica L.). Corminaria 1998, 9, 9–13. [Google Scholar]
- Miko, M.; Gažo, J.; Biroščíková, M. In vitro klonové množenie genetických zdrojov jarabiny oskorušovej (Sorbus domestica L.) z územia Slovenska. Acta Fytotechn. Zootechn. 2004, 7, 85–89. (In Slovak) [Google Scholar]
- Nikolaou, P.; Zagas, D.; Scaltsoyiannes, V.; Balas, E.; Xilogianni, V.; Tsoulpha, P.; Tsaktsira, M.; Voulgaridou, E.; Iliev, I.; Triantafyllou, K.; et al. Advances in the micropropagation of service tree (Sorbus domestica L.). Propag. Ornam. Plants 2008, 8, 154–157. [Google Scholar]
- Ďurkovič, J.; Mišalová, A. Wood formation during ex vitro acclimatisation in micropropagated true service tree (Sorbus domestica L.). Plant Cell Tissue Organ Cult. 2009, 96, 343–348. [Google Scholar] [CrossRef]
- Malá, J.; Cvrčková, H.; Máchová, P.; Dostál, J. Mikropropagace Jeřábu Oskeruše (Sorbus domestica L.); Certified Methodology 4/2011; Forestry and Game Management Research Institute: Strnady, Czech Republic, 2011; pp. 1–17. (In Czech) [Google Scholar]
- Piagnani, M.C.; Zaccheo, P.; Crippa, L. Micropropagation of service tree (Sorbus domestica L.): Role of some factors on in vitro proliferation and rooting, and extra vitro acclimatization. Agrochimica 2012, 56, 219–233. [Google Scholar]
- Martini, A.N.; Papafotiou, M. Season and explant origin affect phenolic content, browning of explants, and micropropagation of ×Malosorbus florentina (Zucc.) Browicz. HortScience 2013, 48, 102–107. [Google Scholar] [CrossRef] [Green Version]
- Máchová, P.; Malá, J.; Cvrčková, H.; Dostál, J.; Buriánek, V. In vitro reproduction of rare and endemic species of rowan tree. J. For. Sci. 2013, 59, 386–390. [Google Scholar] [CrossRef] [Green Version]
- Chalupa, V. In vitro propagation of mature trees of Sorbus aucuparia L. and field performance of micropropagated tree. J. For. Sci. 2002, 48, 529–535. [Google Scholar] [CrossRef] [Green Version]
- Bhojwani, S.S.; Razdan, M.K. Plant Tissue Culture: Theory and Practice; Elsevier: Amsterdam, The Netherlands, 1983; pp. 1–502. [Google Scholar]
- Hackett, W.P. Juvenility, maturation and rejuvenation in woody plants. Hort. Rev. 1985, 7, 109–155. [Google Scholar]
- Basheer-Salimia, R. Juvenility, maturity and rejuvenation in woody plants. Hebron Univ. Res. J. 2007, 3, 17–43. [Google Scholar]
- Ďurkovič, J.; Mišalová, A. Micropropagation of temperate noble hardwoods: An overview. Funct. Plant Sci. Biotechnol. 2008, 2, 1–19. [Google Scholar]
- Werbrouck, S.P.O.; van der Jeugt, B.; Dewitte, W.; Prinsen, E.; Van Onckelen, H.A.; Debergh, P.C. The metabolism of benzyladenine in Spathiphyllum floribundum ‘Schott Petite’ in relation to acclimatisation problems. Plant Cell Tissue Organ Cult. 1995, 14, 662–665. [Google Scholar] [CrossRef]
- Bairu, M.W.; Stirk, W.A.; Doležal, K.; Van Staden, J. Optimizing the micropropagation protocol for the endangered Aloe polyphylla: Can meta-topolin and its derivatives serve as replacement for benzyladenine and zeatin? Plant Cell Tissue Organ Cult. 2007, 90, 15–23. [Google Scholar] [CrossRef]
- Tarkowská, D.; Doležal, K.; Tarkowski, P.; Astot, C.; Holub, J.; Fuksová, K.; Schmülling, T.; Sandberg, G.; Strnad, M. Identification of new aromatic cytokinins in Arabidopsis thaliana and Populus × canadensis leaves by LC-(+)ESI-MS and capillary liquid chromatography/frit-fast atom bombardment mass spektrometry. Physiol. Plant. 2003, 117, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Strnad, M.; Hanuš, J.; Vaněk, T.; Kamínek, M.; Ballantine, J.A.; Fussell, B.; Hanke, D.E. Meta-topolin, a highly active aromatic cytokinin from poplar leaves (Populus × canadensis Moench., cv. Robusta. Phytochemistry 1997, 45, 213–218. [Google Scholar] [CrossRef]
- Werbrouck, S.P.O.; Strnad, M.; Van Onckelen, H.A.; Debergh, P.C. Meta-topolin, an alternative to benzyladenine in tissue culture? Physiol. Pl. 2008, 98, 291–297. [Google Scholar] [CrossRef]
- Amoo, S.O.; Van Staden, J. Influence of plant growth regulators on shoot proliferation and secondary metabolite production in micpropagated Huernia hystrix. Plant Cell Tissue Organ Cult. 2013, 112, 249–256. [Google Scholar] [CrossRef]
- Moyo, M.; Finnie, J.F.; Van Staden, J. Recalcitrant effects associated with the development of basal callus-like tissue on caulogenesis and rhizogenesis in Sclerocarya birrea. Plant Growth Regul. 2011, 63, 187–195. [Google Scholar] [CrossRef]
- Šedivá, J.; Vlašínová, H.; Klemš, M.; Vejsadová, H.; Švajdlenka, E.; Mertelík, J.; Kloudová, K.; Havel, L. Behaviour of resistant and non-resistant clones of Aesculus hippocastanum in vitro. Acta Hort. 2013, 988, 123–128. [Google Scholar] [CrossRef]
- van der Westhuizen, A. The use of meta-topolin as an alternative cytokinin in the tissue culture of Eucalyptus species. Acta Hort. 2014, 1055, 25–28. [Google Scholar] [CrossRef]
- Ördögh, M.; Jambor-Benczúr, E.; Tilly-Mándy, A.; Lelik, L. Effects of different cytokinins on proliferation of Sorbus borbasii ‘Herkulesfürdö’. Propag. Ornam. Plants 2009, 9, 43–46. [Google Scholar]
- Jeong, B.R.; Sivanesan, I. Micropropagation of Sorbus commixta Hedl. Propag. Ornam. Plants 2015, 15, 142–146. [Google Scholar]
- Ördögh, M.; Jambor-Benczúr, E.; Tilly-Mándy, A.; Lelik, L. The effects of growth regulators in proliferation of Sorbus redliana “Burokvölgy”. Int. J. Hortic. Sci. 2006, 12, 77–83. [Google Scholar] [CrossRef]
- Meyer, E.M.; Touchell, D.H.; Ranney, T.G. In vitro shoot regeneration and polyploidy induction from leaves of Hypericum species. Hortscience 2009, 44, 1957–1961. [Google Scholar] [CrossRef] [Green Version]
- Kucharska, D.; Orlikowska, T.; Maciorowski, R.; Kunka, M.; Wójcik, D.; Pluta, S. Application of meta-Topolin for improving micropropagation of gooseberry (Ribes grossularia). Sci. Hort. 2020, 272, 109529. [Google Scholar] [CrossRef]
- Hlophe, N.P.; Aremu, A.O.; Doležal, K.; Van Staden, J.; Finnie, J.F. Cytokinin-facilitated plant regeneration of three Brachystelma species with different conservation status. Plants 2020, 9, 1657. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Ito, J.; Sugiura, A. Comparison of growth and rooting characteristics of micropropagated adult plants and juvenile seedlings of persimmon (Diospyros kaki L.). J. Jpn. Soc. Hortic. Sci. 1994, 63, 537–541. [Google Scholar] [CrossRef]
- Mendonça, E.G.; Batista, T.R.; Stein, V.C.; Balieiro, F.P.; Abreu, J.R.; Pires, M.F.; de Souza, P.A.; Paiva, L.V. In vitro serial subculture to improve rooting of Eucalyptus urophylla. New For. 2020, 51, 801–816. [Google Scholar] [CrossRef]
- Grant, N.J.; Hammatt, N. Increased root and shoot production during micropropagation of cherry and apple rootstocks: Effect of subculture frequency. Tree Physiol. 1999, 19, 899–903. [Google Scholar] [CrossRef] [Green Version]
- Economou, A.S. From microcutting rooting to microplant establishment: Key points to consider for maximum success in woody plants. Acta Horicult. 2013, 988, 43–56. [Google Scholar] [CrossRef]
- Májovský, J.; Bernátová, D. Nové hybridogénne podrody rodu Sorbus L. emend. Crantz. Acta Hortic. Regiotect. 2001, 1, 20–21. (In Slovak) [Google Scholar]
- Aroonpong, P.; Chang, J.C. Micropropagation of a difficult-to-root weeping mulberry (Morus alba var. Shidareguwa): A popular variety for ornamental purposes. Sci. Hort. 2015, 194, 320–326. [Google Scholar] [CrossRef]
- Iapichino, G.; Airo, M. Multiplication of Crataegus monogyna by in vitro culture of nodal segments. Acta Hort. 2009, 812, 135–140. [Google Scholar] [CrossRef]
- Noshad, D.; Miresmaili, S.; Riseman, A.; Ekramoddoullah, A. In vitro propagation of seven Daphne L. species. Plant Cell Tissue Organ Cult. 2009, 96, 201–209. [Google Scholar] [CrossRef]
- Debergh, P.C.; Topoonyanont, N.; Van Huylenbroeck, J.; Moreira da Silva, H.; Oyaert, E. Preparation of microplants for ex vitro establishment. Acta Hort. 2000, 530, 269–276. [Google Scholar] [CrossRef]
- Herman, E.B. Recent Advances in Plant Tissue Culture XVIII. Plant Micropropagation: 2010–2013; Agritech Consultants, Inc.: Mohegan Lake, NY, USA, 2013; pp. 1–138. [Google Scholar]
- Apter, R.C.; McWilliams, E.L.; Davies, F.T., Jr. In vitro and ex vitro adventitious root formation in Asian jasmine (Trachelospermum asiaticum). I. Comparative morphology. J. Am. Soc. Hort. Sci. 1993, 118, 902–905. [Google Scholar] [CrossRef] [Green Version]
- Soukup, A.; Malá, J.; Hrubcová, M.; Kálal, J.; Votrubová, O.; Cvikrová, M. Differences in anatomical structure and lignin content of roots of pedunculate oak and wild cherry-tree plantlets during acclimation. Biol. Pl. 2004, 48, 481–489. [Google Scholar] [CrossRef]
- Hatzilazarou, S.P.; Syros, T.D.; Yupsanis, T.A.; Bosabalidis, A.M.; Economou, A.S. Peroxidases, lignin and anatomy during in vitro and ex vitro rooting of gardenia (Gardenia jasminoides Ellis) microshoots. J. Pl. Physiol. 2006, 163, 827–836. [Google Scholar] [CrossRef]
- Pruski, K.W.; Tina, L.; Astatkie, T.; Nowak, J. Micropropagation of chokecherry and pincherry cultivars. Plant Cell Tissue Organ Cult. 2000, 63, 93–100. [Google Scholar] [CrossRef]
- Marques, S.L.; Canhoto, J.; Gonçalves, J.C.; Diogo, M.G. Micropropagation of Sorbus torminalis (L.) Crantz: Hormonal effects during multiplication and rooting phases. Acta Hort. 2013, 990, 397–404. [Google Scholar] [CrossRef]
- Šedivá, J.; Businský, R.; Pospíšková, M.; Velebil, J.; Drahošová, H.; Zýka, V. Conservation methods of Czech gene pool of whitebeams. Acta Hort. 2021, 1324, 41–46. [Google Scholar] [CrossRef]
- Pierik, R.L.M. In Vitro Culture of Higher Plants; Kluwer Academic Publishers Group: Dordrecht, The Netherlands, 1989; pp. 1–344. [Google Scholar]
- De Klerk, G.J.; Van der Krieken, W.; De Jong, J.C. Review the formation of adventitious roots: New concepts, new possibilities. Vitr. Cell Dev. Biol.-Plant 1999, 35, 189–199. [Google Scholar] [CrossRef]
- De Klerk, G.J.M. Rooting in vitro. Unexpected effects of deficient gas exchange. Prophyta Annu. 2013, 2013, 46–47. [Google Scholar]
- De Klerk, G.J.M.; Paffen, A.K. MnO4-coated grains prevent a massive, detrimental rise of ethylene during in vitro rooting of rose microcuttings. Propag. Ornam. Plants 2020, 20, 96–102. [Google Scholar]
- Šedivá, J.; Zahumenická, P.; Fernández Cusimamani, E. An efficient in vitro propagation protocol for snowdrop anemone (Anemone sylvestris L.). Hort. Sci. 2017, 44, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Sreekissoon, A.; Plačková, L.; Doležal, K.; Finnie, J.F.; Van Staden, J. In vitro and ex vitro vegetative propagation and cytokinin profiles of Sceletium tortuosum (L.) N. E. Br.: A South African medicinal plant. Plant Cell Tissue Organ Cult. 2021, 145, 191–202. [Google Scholar] [CrossRef]
- Haq, I.U.; Ahmad, T.; Hafiz, I.A.; Abbasi, N.A. Influence of microcutting sizes and IBA concentrations on in vitro rooting of olive cv. ‘Dolce Agogia’. Pak. J. Bot. 2009, 41, 1213–1222. [Google Scholar]
- OuYang, F.; Wang, J.; Li, Y. Effects of cutting size and exogenous hormone treatment on rooting of shoot cuttings in Norway spruce [Picea abies (L.) Karst.]. New For. 2015, 46, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Lepší, M.; Koutecký, P.; Nosková, J.; Lepší, P.; Urfus, T.; Rich, T.C.G. Versatility of reproductive modes and ploidy level interactions in Sorbus s.l. (Malinae, Rosaceae). Bot. J. Linn. Soc. 2019, 191, 502–522. [Google Scholar] [CrossRef]
- Velebil, J. Sorbus omissa, a new endemic hybridogenous species from the lower Vltava river valley. Preslia 2012, 84, 375–390. [Google Scholar]
- Sdmtoolbox. Available online: http://www.sdmtoolbox.org (accessed on 7 July 2022).
- Pladias. Database of the Czech Flora and Vegetation. Available online: http://www.pladias.cz/en/ (accessed on 7 July 2022).
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Montgomery, D.C. Design and Analysis of Experiments; Willey: New York, NY, USA, 2012; pp. 1–730. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PGR | Genotype | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. × abscondita | S. gemella | S. × kitaibeliana | S. omissa | |||||||||
| n | SE | Group | n | SE | Group | n | SE | Group | n | SE | Group | |
| Control | 2.00 | 0.28 | abcd | 1.11 | 0.09 | abcd | 1.28 | 0.09 | ab | 1.00 | 0.00 | a |
| BA | 2.97 | 0.41 | de | 2.36 | 0.39 | bcde | 3.42 | 0.59 | de | 2.42 | 0.36 | abcde |
| BA+IBA | 2.42 | 0.33 | abcde | 1.69 | 0.22 | abcd | 2.42 | 0.33 | abcde | 2.11 | 0.27 | abcd |
| mT | 3.72 | 0.36 | e | 2.39 | 0.42 | abcde | 2.83 | 0.36 | cde | 1.89 | 0.14 | abcd |
| mT+IBA | 3.47 | 0.31 | e | 2.69 | 0.43 | bcde | 2.81 | 0.34 | cde | 2.13 | 0.37 | abcd |
| IBA | 1.69 | 0.33 | abcd | 1.03 | 0.03 | a | 1.36 | 0.12 | abc | 1.03 | 0.03 | a |
| PGR | Genotype | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. × abscondita | S. gemella | S. × kitaibeliana | S. omissa | |||||||||
| l | SE | Group | l | SE | Group | l | SE | Group | l | SE | Group | |
| Control | 56.69 | 8.9 | bcdef | 22.97 | 1.0 | a | 31.69 | 3.6 | abc | 20.67 | 0.6 | a |
| BA | 59.92 | 8.3 | cdef | 40.56 | 5.3 | abcde | 71.00 | 12.9 | ef | 42.36 | 5.7 | abcde |
| BA+IBA | 54.94 | 7.8 | abcde | 30.50 | 3.2 | abc | 56.03 | 7.6 | bcde | 42.03 | 5.2 | abcde |
| mT | 68.13 | 6.5 | def | 50.53 | 5.5 | abcde | 61.81 | 5.9 | cdef | 36.08 | 1.9 | abcd |
| mT+IBA | 89.22 | 8.1 | f | 44.86 | 4.1 | abcde | 65.50 | 6.3 | def | 50.49 | 8.8 | abcde |
| IBA | 39.99 | 7.3 | abcde | 27.00 | 1.2 | ab | 36.31 | 3.9 | abcd | 24.25 | 0.8 | ab |
| Rooting | Mean Number of Roots per Microcutting | |||||||
|---|---|---|---|---|---|---|---|---|
| df | Mean Squares | F | df | Mean Squares | F | |||
| Genotype | 3 | 2.42 | 53.98 | *** | 3 | 23.06 | 72.66 | *** |
| Auxin treatment | 2 | 1.13 | 25.31 | *** | 2 | 5.83 | 18.37 | *** |
| Microcutting size | 1 | 1.15 | 25.71 | *** | 1 | 1.52 | 4.80 | * |
| Genotype × auxin treatment | 6 | 0.35 | 7.97 | *** | 6 | 3.98 | 12.56 | *** |
| Genotype × microcutting size | 3 | 0.25 | 5.65 | ** | 3 | 0.48 | 15.14 | ns |
| Auxin treatment × microcutting size | 2 | 0.02 | 0.59 | ns | 2 | 0.10 | 0.32 | ns |
| Error | 54 | 0.04 | 568 | 0.31 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šedivá, J.; Velebil, J.; Zahradník, D. Micropropagation as a Tool for the Conservation of Autochthonous Sorbus Species of Czechia. Plants 2023, 12, 488. https://doi.org/10.3390/plants12030488
Šedivá J, Velebil J, Zahradník D. Micropropagation as a Tool for the Conservation of Autochthonous Sorbus Species of Czechia. Plants. 2023; 12(3):488. https://doi.org/10.3390/plants12030488
Chicago/Turabian StyleŠedivá, Jana, Jiří Velebil, and Daniel Zahradník. 2023. "Micropropagation as a Tool for the Conservation of Autochthonous Sorbus Species of Czechia" Plants 12, no. 3: 488. https://doi.org/10.3390/plants12030488
APA StyleŠedivá, J., Velebil, J., & Zahradník, D. (2023). Micropropagation as a Tool for the Conservation of Autochthonous Sorbus Species of Czechia. Plants, 12(3), 488. https://doi.org/10.3390/plants12030488