


The Ovule Number Variation Provides New Insights into Taxa Delimitation in Willows (Salix subgen. Salix; Salicaceae)

Abstract
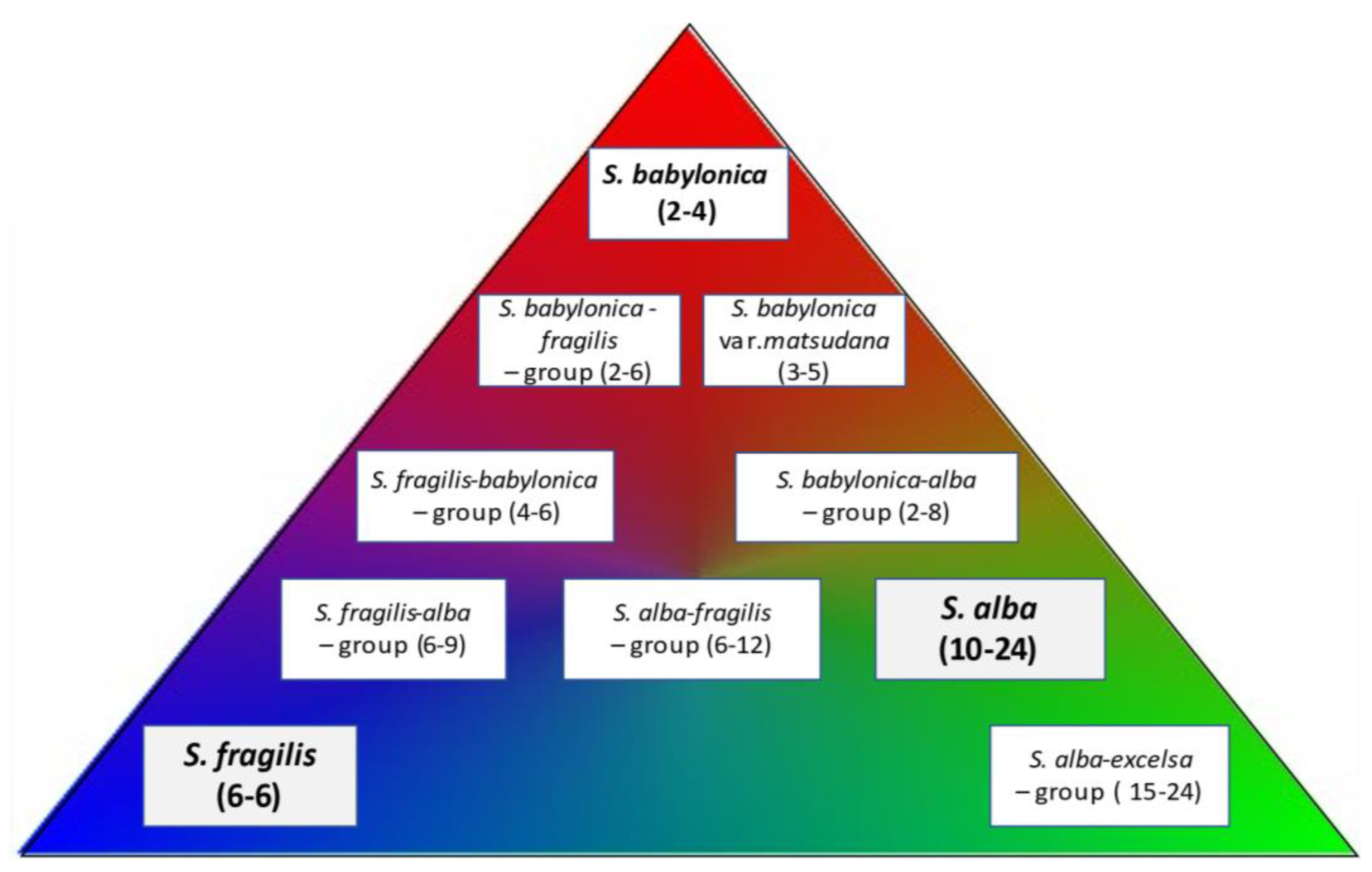
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Ovule Count
3. Results
3.1. Salix babylonica
3.1.1. S. babylonica var. sacramenta Marchenko et Kuzovkina, ined.
3.1.2. S. babylonica ‘Babylon’
3.1.3. S. babylonica var. matsudana
3.1.4. S. babylonica-fragilis—Group (The Groups Were Originally Designated as Varieties by A.M.; such Designations Were not in Accord with the ICBN and Were Replaced with the Non-Nomenclatural Designations Such as Groups
3.1.5. S. babylonica-alba—Group
3.1.6. Salix babylonica dolorosa
3.2. S. fragilis
3.2.1. S. fragilis-babylonica—Group
3.2.2. The Pure S. fragilis
3.2.3. S. fragilis-alba—Group
3.3. S. alba
3.3.1. S. alba-fragilis—Group
3.3.2. The Pure S. alba
3.3.3. S. alba-excelsa—Group
3.4. Specimen Identifications
3.5. Salix tetrasperma and S. mucronata
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Species | Region, Collector, Herbarium |
|---|---|---|
| 1 | S. babylonica var. sacramenta ‘Soveny Americano’ | Argentina, Delta del Parana, Las Carabelas Farm, 29/10/2018; Y.A.Kuzovkina. CONN |
| 2 | S. babylonica | USA, Boston, Arnold Arboretum, cultivated, 05/05/1996. A.K.Skvortsov A.R.Brach. MHA |
| 3 | Salix—specimen No. 1 in the folder “S. babylonica L.” | China, Yunnan Province, Kunming surroundings, foothills, 2000–2500 m. Clonal plantings along the river, 25–28/03/1998. A.K.Skvortsov. MHA |
| 4 | Salix—specimen No. 2 in the folder “S. babylonica L.’ | China, Yunnan province, Kunming surroundings, foothills, 2000–2500 m. Clonal plantings along the river. 25–28/03/1998. MHA |
| 5 | Salix—specimen No. 3 in the folder “S. babylonica L.” | China, Yunnan province, Kunming surroundings, foothills, 2000–2500 m. Clonal plantings along the river, 25–28/03/1998. A.K.Skvortsov. MHA |
| 6 | S. babylonica | NA 46541; USA, Washington, National Arboretum. Collection of willows. Origin unknown. Planted as Salix matsudana ‘Pendula’, collected by F. G. Meyer, 07/05/1987. (Skvortsov’s note: “very simi-lar to my samples from China in 1998”). MHA |
| 7 | S. babylonica | Spain, province of Castellón, Morella; grown in the Botanical Garden of Madrid from cuttings brought from the bank of the Morella River. Collected during the first flowering in the garden. 18/03/1996. P.Blanco, S.Cirujano, R.Morales. MHA |
| 8 * | S. babylonica | Argentina, Patagonia, Neuquen, planted Villa El Chocon; 4/11//2018. Y.A.Kuzovkina. CONN |
| 9 | S. babylonica | Russia, Crimea, Nikitsky Botanical Garden, Collect. I.Artamonova; 03/05/1956. A.K.Skvortsov. MHA |
| 10 | S. babylonica ‘Babylon’ | NA 44011, USA, Washington, National Arboretum. NA |
| 11 | S. babylonica var. annularis | Edson (S. babylonica var. crispa; Loudon) 0–777 Highland Park, Rochester, NY. 6/8/1918. A |
| 12 | S. matsudana f. tortuosa Rehder | No. 360-60-B, USA, Boston, Arnold Arboretum, plot 5-c. Cuttings obtained in 1960 from AA9046 (Western Nursery, Massachusetts, in 1946); D. Michener, E. Bennett, J. Dricker B. Porter; 782; 05/06/1984. MHA |
| 13 * | S. babylonica ‘Tortuosa’ | No. NA 44014, USA, Washington, National Arboretum |
| 14 * | S. matsudana ‘Tortuosa’ | USA, Chicago, Morton Arboretum |
| 15 | S. matsudana var. tortuosa | Czechoslovakia, Moravia, Brno, Dornic. 19/04/1960. Ing. Chmelař. MHA |
| 16 * | S. matsudana | No. 366-79, USA, Chicago, Morton Arboretum |
| 17 | S. matsudana | No. 17177, USA, Boston, Arnold Arboretum, Single-trunk tree 15–16 m high; 160 cm diameter; received from John Vietch Son nursery (England) in 1913 by D. Michener, E. Bennett S. Erwin; 94; 01/05/1984. MHA |
| 18 | S. matsudana f. pendula Schneider | No. 175-61-A, Boston, Arnold Arboretum; map 4-b. Cuttings from Morton’s Arboretum in 1961. S.Elsik, I.Hay, J.Carey, E. Fisch P. Megowen. 771. 04/06/1984. MHA |
| 19 | S. babylonica | Japan, Iwate Prefecture, Usugin, Kawasaki, 02/05/1949. Masao Kikuchi. MHA |
| 20 | S. matsudana | China, Beijing, 1856. Dr. Tatarinow. MHA |
| 21 | S. matsudana var. tortuosa | Spain, Catalonia, Girona: Banyoles, Lake Banyoles. 459813. 04/05/1998. Blanco, Paloma. MHA |
| 22 | S. babylonica ‘Tortuosa’ | USA, Montgomery, Wheaton, Maryland. Japanese Garden, Brookside Gardens, 24/04/1986. F.G.Meyer, P.M. Mazzeo. MHA |
| 23 * | S. matsudana f. umbraculifera | No. 166-82, USA, Chicago, Morton Arboretum |
| 24 | S. matsudana f. umbraculifera | No. 634-60-A, USA, Boston, Arnold Arboretum. Plot 6-b. Origin: from Morton Arboretum in 1960 S.Elsik, I.Hay, J.Carey, E. Fisch P. Megowen. 772. 04/06/1984. MHA |
| 25 | S. babylonica | Transbaikalia, now the microdistrict of Ulan-Ude, Selenga River. 15/07/1913. S.J Enander/A.K. Skvortsov? MHA |
| 26 | S. babylonica | Japan, Honshu, Nagano Prefecture, Matsumoto, Shinshu University Campus 04/04/1976. T. Shimizu. MHA |
| 27 | S. babylonica | China, Alashan, Dyn-yuan-ying oasis; col. S.S.Chetyrkin. 1909. A.K.Skvortsov. MHA |
| 28 | S. babylonica | China, Prov. Liaoning, st. Xunyaochen, 22/05/1950. Lion Tchen-ngo/A.K. Skvortsov? MHA |
| 29 | S. fragilis | Russia, Smolensk region, Kardymovsky district, near Kardymovo village, the bank of the Khmost River. 11/05/1975. A.K.Skvortsov. MHA |
| 30 | S. fragilis | Russia, Smolensk region, okr. Smolensk. Talashkino, along ditch at border of the homestead, probably not planted. 07/05/1975. A.K.Skvortsov. MHA |
Appendix B
| No | Species | Region, Collector, Herbarium |
|---|---|---|
| 1 | S. fragilis f. latifolia Anderss. | Russia, Mordovia, Saransk. E.T.Malyutina. MHA |
| 2 | S. alba | Portugal, Mertola. P. da Silva, M. Bravo Lima, B. Rainha. MHA |
| 3 | S. alba × S. fragilis | Portugal, Province of Baxio Alentejo, Mertola. P. da Silva, M. Bravo Lima, B. Rainha. MHA |
| 4 | S. alba | Austria, Vienna. A. Neumann. MHA |
| 5 | S. aegyptiaca | Iran, Mazandaran Province: Galanrud, №37175 E. 26/04/1956. Sabeti/S. babylonica L.—A.K.Skvortsov). MHA |
| 6 | S. euxina | 001191972Bulgaria, River Kalnitza, near Botevgrad. 24/04/2015. BM |
| 7 | S. alba | Kazakhstan, Taldy-Kurgan region, r. Karatal, 36–38 km from Ushtobe, the outskirts of the Saryesik-Atyrau desert. A.K.Skvortsov. MHA |
| 8 | S. fragilis | Russia, Mordovia, Saransk. E.T.Malyutina. MHA |
| 9 | S. fragilis | Russia, Moscow region, Serpukhov district, near the village Luzhki. A.K.Skvortsov. MHA |
| 10 | S. fragilis | USSR, Minsk region, Logai district, r. Gaina. A.K.Skvortsov. MHA |
| 11 | S. fragilis var. decipiens | Portugal, Province of Baxio Alentejo. Bento V. Rainha. MHA |
| 12 * | S. alba | Russia, Yekaterinburg, Botanical Garden, salicetum, row 1. I.V.Belyaeva |
| 13 | S. alba | Russia, Moscow region, near Zvenigorod, bank of the river Storozki. A.K.Skvortsov. MHA |
| 14 | S. fragilis | Bohemia, (now Czech Republic), Litomysl. A.K.Skvortsov. MHA |
| 15 | S. alba | Austria, Murau. Gratg/I.V.Belyaeva. K |
| 16 | S. alba | Russia, Stalingrad (now Volgograd) region, near the village Pereshepnoe. A.K.Skvortsov. MHA |
| 17 | S. fragilis | Russia, Ulyanovsk region, Barysh district, s. Rumyantsevo. E.T.Malyutina. MHA |
| 18 | S. fragilis | Russia, Moscow region, Serpukhov district, s. Luzhki. A.K.Skvortsov. MHA |
| 19 | S. alba | Russia, Ulyanovsk region, Barysh district, s. Surskie Vershini. E.T.Malyutina. MHA |
| 20 | S. fragilis L. × S. alba | Russia, Moscow region, the left bank of the Oka below Serpukhov, near s. Luzhki. A.K.Skvortsov. MHA |
| 21 | S. fragilis (×S. alba?) | Russia, Moscow region, right bank of the Oka below Kashira, near the village of Kropotovo. A.K.Skvortsov. MHA |
| 22 | S. alba | Southern Albania. A.H.G.Alston, N.Y. Sandwith/ I.V.Belyaeva. K |
| 23 | S. alba | Russia, Moscow region, Zvenigorod district, Molodenovo village. A.K. Skvortsov. MHA |
| 24 | S. alba | Russia, Moscow region, near Zvenigorod, the bank of the Moscow River. A.K. Skvortsov. MHA |
| 25 | S. alba | Kazakhstan, Taldy-Kurgan region, Ushtobe, the outskirts of the Saryesik-Atyrau desert. A.K. Skvortsov. MHA |
| 26 | S. alba | England, East Yorkshire, Hull, on the road Beverly Road to the village of Newland. C.Waterfall/I.V.Belyaeva. K |
| 27 | S. fragilis × S. alba | Russia, Moscow region, left bank of the Oka below Serpukhov, near the village of Luzhki. A.K.Skvortsov. MHA |
| 28 | S. alba | England, Devon. Drummond/I.V.Belyaeva. K |
| 29 | S. fragilis | Russia, Ulyanovsk region, Gremyachka village. E.T.Malyutina. MHA |
| 30 | S. fragilis | Bulgaria, near the village of Kasitsene, Sofia district. N.Vihodcevsky. MHA |
| 31 | S. alba | Russia, Stalingrad (now Volgograd) region, Kotovsky district, near the village of Popki. A.K.Skvortsov. MHA |
| 32 | S. alba | Russia, Penza region, lake New Sura. E.T.Malyutina. MHA |
| 33 | S. fragilis | Lithuania, outskirts of Kaunas. A.K.Skvortsov. MHA |
| 34 | S. alba var. caerulea | Middlesex, (England), near the water near Longmore. A.B.Jackson/S. alba L.—I.V.Belyaeva. K |
| 35 | S. fragilis | Russia, Orenburg region, Buguruslan district, Kiryushkino village. E.T.Malyutina. MHA |
| 36 | S. alba | Russia, Zap. Altai, Leninogorsk. A.K.Skvortsov. MHA |
| 37 | S. alba | Russia, Saratov region, valley of the river Khoper near Balashov. MHA |
| 38 | S. alba | Russia, Moscow region, Serpukhov district, near the village of Luzhki, floodplain of the Oka. A.K.Skvortsov. MHA |
| 39 | S. fragilis | Russia, Penza region, Kuznetsk district, Tyukhmenevo village. E.T.Malyutina. MHA |
| 40 | S. fragilis | Russia, Penza region, Kuznetsk district, Starye Chasi village. E.T.Malyutina. MHA |
| 41 | S. fragilis | Slovakia, Liptovský Mikuláš district, Parizovice. A.K.Skvortsov? MHA |
| 42 | S. fragilis × S. alba | Russia, Penza region, Kuznetsk district, between the village of Dvoriki and the village of Mordovsky Kachim. E.T.Malyutina. MHA |
| 43 | S. fragilis | USA, Massachusetts, Hampshire, Amherst. H.E.Ahles. MHA |
| 44 | S. alba × S. fragilis | Kazakhstan, Kokchetav region, Shchuchinsk, arboretum KazNIILKH. A.K.Skvortsov. MHA |
| 45 | S. fragilis | Russia, Ulyanovsk region, Inza, on the bank of the Syuksum River. E.T.Malyutina. MHA |
| 46 | S. alba | Russia, Kaluga region, near Tarusa, in the floodplain of the Tarusa River. A.K.Skvortsov. MHA |
| 47 | S. fragilis | Russia, Moscow region, Serpukhov district, near the village of Luzhki. A.K.Skvortsov. MHA |
| 48 | S. fragilis | Russia, Penza region, lake New Sura. E.T.Malyutina. MHA |
| 49 | S. euxina | 001191975 Poland, River Panna, at Tylawa 28/04/2015 BM |
| 50 | S. fragilis | Belgium, Pleinvaux (community of Neupre), (Belgium, province Liège, [bord de l’Qurthe] left bank, not far from Rosiere farm, IFBL/G7.12.22. No. 12931. R. Renard, Vidit J. Lambinon. 11/05/ 1987. MHA |
| 51 | Salix | Greece, North-West Macedonia. A.H.G.Alston, N.Y. Sandwith/S. alba L.—I.V.Belyaeva. K |
| 52 | S. alba | France, Cher, (Centre-Val-de-Loire). I.V.Belyaeva. № 4657. K |
| 53 | Salix | Greece, Macedonia, Lake Kerkini S. alba L.- I.V.Belyaeva. K |
| 54 | Salix | Greece, North-West Macedonia. A.H.G. Alston, N.Y. Sandwith/S. alba L.—I.V.Belyaeva. K |
| 55 | S. alba | Russia, Moscow region, (b. Venevsky Uyezd of Tula Gub.) near the village of Serebryanye Prudy. A.K.Skvortsov. MHA |
| 56 | S. fragilis | 01065618. Turkey, Kars province, near Promezhutochnaya station, bank of the river Kyaklik], 245 ♂ and ♀”. S. Turkevich/S. euxina—I.V.Belyaeva. LE |
| 57 | S. alba | England. Dorling Mills/I.V.Belyaeva. K |
| 58 | S. alba × S. fragilis | Slovakia, Liptovský Mikuláš district. A.K. Skvortsov? MHA |
| 59 | S. alba | Kazakhstan, Taldy-Kurgan region, Karatal river, 36–38 km from Ushtobe, the outskirts of the Saryesik-Atyrau desert. A.K.Skvortsov. MHA |
| 60 | S. alba | Russia, Moscow region, Serpukhov district, Luzhki village. A.K.Skvortsov. MHA |
| 61 | S. alba | Northeast Turkey, A8, Gümüşhane, the valley of the Chorokh River between Maden and Bayburt. A.K.Skvortsov. MHA |
| 62 | S. alba | Greek Macedonia, Struma River near the Kopriva Bridge. M.B. Jurrill/I.V.Belyaeva. K |
| 63 | S. alba var. splendens | Serbia, near Niš (C.K. Schneider/S.×fragilis L. (S. alba × S.euxina I.V.Belyaeva)- I.V.Belyaeva. K |
| 64 | S. alba | England, Hemingford Gray. I.V.Belyaeva. K |
| 65 | S. alba | France, Cher (Centre-Val-de-Loire). I.V.Belyaeva. № 4655. K |
| 66 | S. alba | 0556278 Germany, Hesse nursery, Baumschulen, 7/05/1987. F.G.Meyer. (NA 44016) NA |
| 67 | S. alba | Turkey, B-1, Izmir—Manisa. A.P.Khokhryakov, M.T.Mazurenko/A.K.Skvortsov? MHA |
| 68 | S. alba | Armenia, Vedinsky district, Saray-bulak gorge. A.K.Skvortsov. MHA |
| 69 | S. alba | North Turkey, B8, Erzurum, Palandeken Mountains. A.K.Skvortsov. MHA |
| 70 | S. alba | Iran, Isfahan Province. A.K.Skvortsov. MHA |
| 71 | S. excelsa | Azerbaijan, Kuba district, Cuba-Derbent. A.K.Skvortsov. MHA |
| 72 | S. excelsa | Afghanistan, Takhar Province, about 3 km south of Taloqan. A.K.Skvortsov/I.V.Belyaeva. MHA |
| 73 | S. alba | Western Georgia, Arsian Ridge, road to Goderdza Pass. E.E. Gogina. MHA |
| 74 | S. alba | Azerbaijan, Zuvand basin. A.K.Skvortsov. MHA |
| 75 | S. fragilis | Armenia, Meghri district, okr. Tashtun village. E.E. Gogina, Proskuryakova. MHA |
| 76 | S. alba | Iran, Gilan province, Pahlavi city (now Bandar Anzali). A.K.Skvortsov. MHA |
References
- Skvortsov, A.K. Willows of Russia and Adjacent Countries: Taxonomical and Geographical Revision; University of Joensuu Faculty of Mathematics and Natural Sciences Report Series; University of Joensuu: Joensuu, Finland, 1999; Volume 39. [Google Scholar]
- Argus, G.W. Salix. In Flora of North America north of Mexico; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA, 2010; Volume 7, pp. 23–162. [Google Scholar]
- Gramlich, S.G.; Wagner, N.D.; Horandl, E. RAD-seq reveals genetic structure of the F2-generation of natural willow hybrids (Salix L.) and a great potential for interspecific introgression. BMC Plant Biol. 2018, 18, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, N.D.; He, L.; Horandl, E. Phylogenomic relationships and evolution of polyploid Salix species revealed by RAD sequencing data. Front. Plant Sci. 2020, 11, 1077. [Google Scholar] [CrossRef] [PubMed]
- Marchenko, A.M.; Kuzovkina, Y.A. Calculation of the ovule number in the genus Salix: A method for taxa differentiation. Appl. Plant Sci. 2021, 9, e11450. [Google Scholar] [CrossRef] [PubMed]
- Skvortsov, A.K.; Golisheva, M.D. Issledovanie anatomii lista uv v svyazi s sistematikoy roda i semeistva. [A study of the leaf anatomy of Salix for the systematics of the genus and the family]. Acta Bot. 1966, 12, 125–174. (In Russian) [Google Scholar]
- Fang, Z.-F.; Zhao, S.-D.; Skvortsov, A.K. Salicaceae. In Flora of China; Wu, Z.-Y., Raven, P.H., Eds.; Science Press: Beijing China; Missouri Botanical Garden Press: St. Louis, MO, USA, 1999; Volume 4, pp. 162–274. [Google Scholar]
- Santamour, F.S.; McArdle, A.J. Cultivars of Salix babylonica and other weeping willows. J. Arboric. 1988, 14, 180–184. [Google Scholar] [CrossRef]
- Meikle, R.D. Willows and Poplars of Great Britain and Ireland; Botanical Society of the British Isles: London, UK, 1984. [Google Scholar]
- Triest, L.; De Greef, B.; Vermeersch, S.; Van Slycken, J.; Coart, E. Genetic variation and putative hybridisation in Salix alba and Salix fragilis: Evidence from allozyme data. Plant Syst. Evol. 2000, 215, 169–187. [Google Scholar] [CrossRef]
- Triest, L. Hybridization in staminate and pistillate Salix alba and S. fragilis (Salicaceae): Morphology versus RAPDs. Plant Syst. Evol. 2001, 226, 143–154. [Google Scholar] [CrossRef]
- De Cock, K.; Lybeer, B.; Vander Mijnsbrugge, K.; Zwaenepoel, A.; Van Peteghem, P.; Quataert, P.; Breyne, P.; Goetghebeur, P.; Slycken, J.V. Diversity of the willow complex Salix alba–S. x rubens-S-fragilis. Silvae Genet. 2003, 52, 148–153. [Google Scholar]
- King, R.A.; Harris, S.L.; Karp, A. Characterization and inheritance of nuclear microsatellite loci for use in population studies of the allotetraploid Salix alba–Salix fragilis complex. Tree Genet. Genomes 2010, 6, 247–258. [Google Scholar] [CrossRef]
- Skvortsov, A.K. Sovremennoye rasprostraneniye i veroyatnyĭ pervichnyĭ areal lomkoĭ ivy (Salix fragilis L.) [Present distribution and probable primary range of brittle willow (Salix fragilis L.)]. In Problemy Biogeotsenologii, Geobotaniki i Botanicheskoĭ Geografii [Problems in Biogeocoenology, Geobotany and Botanical Geography]; Tikhomirov, B.A., Ed.; Nauka: Leningrad, Russia, 1973; pp. 236–278. Available online: http://www.salicicola.com/translations/Skv1973SF.html (accessed on 20 January 2023)(In Russian, English Translation).
- Marchenko, A.M.; Kuzovkina, Y.A. Notes on the nomenclature and taxonomy of Salix fragilis (Salicaceae). Taxon 2022, 71, 721–732. [Google Scholar] [CrossRef]
- Meneghetti, S.; Barcaccia, G.; Paiero, P.; Lucchin, M. Genetic characterization of Salix alba L. and Salix fragilis L. by means of different PCR-derived marker systems. Plant Biosyst. 2007, 141, 283–291. [Google Scholar] [CrossRef]
- Kehl, A.; Aas, G.; Rambold, G. Genotypical and multiple phenotypical traits discriminate Salix × rubens Schrank clearly from its parent species. Plant Syst. Evol. 2008, 275, 169–179. [Google Scholar] [CrossRef]
- Oberprieler, C.; Dietz, L.; Harlander, C.; Heilmann, J. Molecular and phytochemical evidence for the taxonomic integrity of Salix alba, S. fragilis, and their hybrid S. ×rubens (Salicaceae) in mixed stands in SE Germany. Plant Syst. Evol. 2013, 299, 1107–1118. [Google Scholar] [CrossRef]
- Barcaccia, G.; Meneghetti, S.; Lucchin, M.; De Jong, H. Genetic segregation and genomic hybridization patterns support an allotetraploid structure and disomic inheritance for Salix species. Diversity 2014, 6, 633–651. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Nyman, T.; Wang, D.-C.; Argus, G.W.; Yang, Y.-P.; Chen, J.-H. Phylogeny of Salix subgenus Salix s.l. (Salicaceae): Delimitation, biogeography, and reticulate evolution. BMC Evol. Biol. 2015, 15, 31. [Google Scholar] [CrossRef] [Green Version]
- Sitzia, T.; Barcaccia, G.; Lucchin, M. Genetic diversity and stand structure of neighboring white willow (Salix alba L.) populations along fragmented riparian corridors: A case study. Silvae Genet. 2018, 67, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Barcaccia, G.; Meneghetti, S.; Albertini, E.; Triest, L.; Lucchin, M. Linkage mapping in tetraploid willows: Segregation of molecular markers and estimation of linkage phases support an allotetraploid structure for Salix alba × Salix fragilis interspecific hybrids. Heredity 2003, 90, 169–180. [Google Scholar] [CrossRef]
- Primack, R.B. Relationships among flowers, fruits, and seeds. Annu. Rev. Ecol. Syst. 1987, 18, 409–430. [Google Scholar] [CrossRef]
- Endress, P.K. Angiosperm ovules: Diversity, development, evolution. Ann. Bot. 2011, 107, 1465–1489. [Google Scholar] [CrossRef] [Green Version]
- Strelin, M.M.; Aizen, M.A. The interplay between ovule number, pollination and resources as determinants of seed set in a modular plant. PeerJ 2018, 6, e5384. [Google Scholar] [CrossRef]
- Chmelař, J. Taxonomický význam pocˇtu zakladu semen kapslí u rodu Salix L. [Taxonomic importance of capsule seed number in the genus Salix L.]. Acta Mus. Sil. Ser. C Dendrol. 1977, 26, 1–7. (In Czech) [Google Scholar]
- Argus, G.W. The genus Salix (Salicaceae) in the southeastern United States. Syst. Bot. Monogr. 1986, 9, 1–170. [Google Scholar] [CrossRef]
- Valyagina-Malutina, E.T. Willows of the European Part of Russia; KMK: Moscow, Russia, 2004. (In Russian) [Google Scholar]
- Marchenko, A.M. Semyazachatki i Identifikaciya iv (Salix) [Ovules and Identification of Willows (Salix)]; Publishing House: Moscow, Russia, 2019. (In Russian) [Google Scholar]
- Marchenko, A.M.; Kuzovkina, Y.A. Identification of hybrid formulae of a few willows (Salix) using ovule numbers. Silvae Genet. 2021, 70, 75–83. [Google Scholar] [CrossRef]
- INTA. 2021. Available online: https://inta.gob.ar/variedades/soveny-americano (accessed on 30 June 2022).
- Ragonese, A.E.; Alberti, F.R. Sauces hibridas originados naturalmente en la Republica Argentina. Rev. Investig. Agric. 2015, 12, 111–153. (In Spanish) [Google Scholar]
- Krüssmann, G. Manual of Cultivated Broad-Leaved Trees and Shrubs; Timber Press: Beaverton, OR, USA, 1984. [Google Scholar]
- Dirr, M.A. Manual of Woody Landscape Plants: Their Identification, Ornamental Characteristics Culture, Propagation and Uses, 6th ed.; Stipes Publishing: Champaign, IL, USA, 2009. [Google Scholar]
- Vermont Willow Nursery. Salix matsudana. 2021. Available online: https://www.willowsvermont.com/home.html (accessed on 15 July 2021).
- Ohashi, H.; Yonekura, K. Additions and corrections for Salicaceae of Japan 3. J. Jpn. Bot. 2015, 90, 1–14. [Google Scholar]
- Bailey, L.H. Cyclopedia of American Horticulture; Virtue: Toronto, ON, Canada, 1902; Volume 4, p. 1600. [Google Scholar]
- Bailey, L.H. The Standard Cyclopedia of Horticulture; Macmillan, 1917; Volume 6, p. 3053. [Google Scholar]
- Rehder, A. Bibliography of Cultivated Trees and Shrubs; Arnold Arboretum of Harvard University: Jamaica Plain, MA, USA, 1949. [Google Scholar]
- Cronk, Q.; Ruzzier, E.; Belyaeva, I.; Percy, D. Salix transect of Europe: Latitudinal patterns in willow diversity from Greece to arctic Norway. Biodivers. Data J. 2015, 3, e6258. [Google Scholar] [CrossRef] [Green Version]
- Christensen, K.; Jonsell, B. Proposal to conserve the name Salix fragilis with a conserved type (Salicaceae). Taxon 2005, 54, 555–556. [Google Scholar] [CrossRef]







| No | Specimen | Percentage of Valves with Each Ovule Number | No. of Ovules per Ovary | Ovule Index | Trichomes | Nectaries | Ovary | No of Ovaries | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | Min/ Often | Max/ Often | |||||||
| 1 | S. babylonica var. sacramenta ‘Soveny Americano’ | 61 | 39 | 2/2 | 3/3 | 2–3 | A | 1 | G | 50 | ||
| 2 | S. babylonica | 28 | 72 | 2/3 | 4/4 | 2–4 | A | 1 | G | 68 | ||
| 3 | Salix—specimen No. 1 in the folder “S. babylonica” | 17 | 83 | 2/3 | 4/4 | 2–4 | A | 1 | G | 58 | ||
| 4 | Salix—specimen No. 2 in the folder “S. babylonica.” | 16 | 84 | 2/3 | 4/4 | 2–4 | A | 2 | G | 50 | ||
| 5 | Salix—specimen No. 3 in the folder “S. babylonica” | 9 | 91 | 2/3 | 4/4 | 2–4 | A | 2 | G | 92 | ||
| 6 | S. babylonica | 8 | 92 | 2/3 | 4/4 | 2–4 | A | 1 | G | 56 | ||
| 7 | S. babylonica | 5 | 95 | 3/4 | 4/4 | 3–4 | A | 2 | G, P | 130 | ||
| 8 | S. babylonica | 4 | 96 | 3/4 | 4/4 | 3–4 | A | 1 | G | 60 | ||
| 9 | S. babylonica | 1 | 99 | 3/4 | 4/4 | 3–4 | A | 1 | P | 58 | ||
| 10 | S. babylonica ‘Babylon’ | 1 | 99 | 3/4 | 4/4 | 3–4 | A | 1 | G | 119 | ||
| 11 | S. babylonica var. annularis | 8 | 92 | 3/3 | 4/4 | 3–4 | A | 1 | P at base | 109 | ||
| 12 | S. matsudana f. tortuosa | 4 | 96 | 3/3 | 4/4 | 3–4 | A | 1 | G | 56 | ||
| 13 | S. babylonica ‘Tortuosa’ | 1 | 99 | 3/4 | 4/4 | 3–4 | A | 1 | G | 46 | ||
| 14 | S. matsudana ‘Tortuosa’ | 0 | 100 | 4/4 | 4/4 | 4–4 | A | 1 | G | 52 | ||
| 15 | S. matsudana var. tortuosa | 0 | 100 | 4/4 | 4/4 | 4–4 | A | 1 | G | 55 | ||
| 16 | S. matsudana | 0 | 100 | 4/4 | 4/4 | 4–4 | A | 1 | G | 32 | ||
| 17 | S. matsudana | 0 | 100 | 4/4 | 4/4 | 4–4 | A | 1 | G | 65 | ||
| 18 | S. matsudanaf. pendula | 0 | 100 | 4/4 | 4/4 | 4–4 | A | 1 | P | 56 | ||
| 19 | S. babylonica | 3 | 96 | 1 | 3/4 | 5/4 | 3–5 | A | 1, 2 | G, P | 91 | |
| 20 | S. matsudana | 1 | 98 | 1 | 3/4 | 5/4 | 3–5 | A | 1 | G | 105 | |
| 21 | S. matsudana var. tortuosa | 1 | 93 | 6 | 3/4 | 5/5 | 3–5 | A | 1 | G | 97 | |
| 22 | S. babylonica ‘Tortuosa | 0 | 99 | 1 | 4/4 | 5/4 | 4–5 | A | 1 | G | 56 | |
| 23 | S. matsudana ‘Umbraculifera’ | 0 | 85 | 15 | 4/4 | 6/5 | 4–6 | A | 1, 2 | G | 35 | |
| 24 | S. matsudana ‘Umbraculifera’ | 0 | 78 | 22 | 4/4 | 6/5 | 4–6 | A | 1 | G | 45 | |
| 25 | S. babylonica | 0 | 80 | 20 | 4/4 | 6/5 | 4–6 | A | 1 | G | 20 | |
| 26 | S. babylonica | 0 | 99 | 1 | 4/4 | 5/4 | 4–5 | A | 1, 2 | G, P | 132 | |
| 27 | S. babylonica | 0 | 98 | 2 | 4/4 | 5/4 | 4–5 | A | 1 | G | 191 | |
| 28 | S. babylonica | 0 | 97 | 3 | 4/4 | 5/4 | 4–5 | A | 1 | G | 57 | |
| 29 | S. fragilis | 10 | 61 | 29 | 2/4 | 6/5 | 2–6 | L | 1 | G | 38 | |
| 30 | S. fragilis | 3 | 24 | 56 | 17 | 2/4 | 8/6 | 2–8 | L | 1 | G | 38 |
| No | Specimen | Percentage of Valves with Each Ovule Number | No. of Ovules per Ovary | Ovule Index | Trichomes | No of Ovaries | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | Min/ Often | Max/ Often | |||||
| 1 | S. fragilis f. latifolia | 33 | 67 | 0 | 0 | 0 | 4/4 | 6/6 | 4–6 | L | 70 |
| 2 | S. alba | 27 | 73 | 0 | 0 | 0 | 4/5 | 6/6 | 4–6 | L | 44 |
| 3 | S. alba × S. fragilis | 12 | 88 | 0 | 0 | 0 | 5/5 | 6/6 | 5–6 | L | 82 |
| 4 | S. alba | 2 | 98 | 0 | 0 | 0 | 5/6 | 6/6 | 5–6 | N | 18 |
| 5 | S. aegyptiaca | 1 | 99 | 0 | 0 | 0 | 5/6 | 6/6 | 5/6 | L | 72 |
| 6 | S. euxina | 0 | 100 | 0 | 0 | 0 | 6/6 | 6/6 | 6/6 | L | 45 |
| 7 | S. alba | 3 | 94 | 3 | 0 | 0 | 4/6 | 7/7 | 4–7 | S | 75 |
| 8 | S. fragilis | 3 | 80 | 17 | 0 | 0 | 5/6 | 8/7 | 5–8 | N | 75 |
| 9 | S. fragilis | 1 | 76 | 23 | 0 | 0 | 6/6 | 8/7 | 6–8 | L | 51 |
| 10 | S. fragilis | 1 | 53 | 44 | 2 | 0 | 6/6 | 9/8 | 6–9 | L | 48 |
| 11 | S. fragilis var. decipiens | 3 | 50 | 33 | 14 | 0 | 4/6 | 9/8 | 4–9 | N | 30 |
| 12 | S. alba | 1 | 50 | 47 | 2 | 0 | 6/6 | 9/7 | 6–9 | S | 146 |
| 13 | S. alba | 0 | 98 | 2 | 0 | 0 | 6/6 | 7/6 | 6–7 | S | 70 |
| 14 | S. fragilis | 0 | 93 | 7 | 0 | 0 | 6/6 | 7/6 | 6–7 | L | 65 |
| 15 | S. alba | 0 | 92 | 8 | 0 | 0 | 6/6 | 7/6 | 6–7 | A | 18 |
| 16 | S. alba | 0 | 90 | 10 | 0 | 0 | 6/6 | 7/6 | 6–7 | A | 35 |
| 17 | S. fragilis | 0 | 90 | 10 | 0 | 0 | 6/6 | 7/6 | 6–7 | L | 92 |
| 18 | S. fragilis | 0 | 89 | 11 | 0 | 0 | 6/6 | 8/7 | 6–8 | A | 66 |
| 19 | S. alba | 0 | 89 | 11 | 0 | 0 | 6/6 | 8/7 | 6–8 | S | 106 |
| 20 | S. fragilis × S. alba | 0 | 64 | 36 | 0 | 0 | 6/6 | 8/7 | 6–8 | S | 40 |
| 21 | S. fragilis (×S. alba?) | 0 | 56 | 44 | 0 | 0 | 6/6 | 8/7 | 6–8 | L | 39 |
| 22 | S. alba | 0 | 50 | 50 | 0 | 0 | 6/6 | 8/8 | 6–8 | N | 20 |
| 23 | S. alba | 0 | 95 | 3 | 2 | 0 | 6/6 | 8/6 | 6–8 | S | 60 |
| 24 | S. alba | 0 | 92 | 6 | 2 | 0 | 6/6 | 8/6 | 6–8 | S | 85 |
| 25 | S. alba | 0 | 90 | 7 | 3 | 0 | 6/6 | 9/7 | 6–9 | S | 73 |
| 26 | S. alba | 0 | 79 | 18 | 3 | 0 | 6/6 | 8/7 | 6–8 | A | 14 |
| 27 | S. fragilis × S. alba | 0 | 76 | 23 | 1 | 0 | 6/6 | 8/7 | 6–8 | S | 40 |
| 28 | S. alba | 0 | 74 | 19 | 7 | 0 | 6/6 | 9/7 | 6–9 | S | 31 |
| 29 | S. fragilis | 0 | 70 | 26 | 4 | 0 | 6/6 | 8/8 | 6–8 | S | 73 |
| 30 | S. fragilis | 0 | 56 | 42 | 2 | 0 | 6/6 | 8/7 | 6–8 | L | 44 |
| 31 | S. alba | 0 | 56 | 39 | 5 | 0 | 6/6 | 9/7 | 6–9 | N | 97 |
| 32 | S. alba | 0 | 60 | 35 | 5 | 0 | 6/6 | 9/7 | 6–9 | S | 71 |
| 33 | S. fragilis | 0 | 38 | 52 | 10 | 0 | 6/7 | 10/8 | 6–10 | L | 54 |
| 34 | S. alba var. caerulea | 0 | 36 | 27 | 37 | 0 | 6/6 | 10/9 | 6–10 | A | 28 |
| 35 | S. fragilis | 0 | 34 | 41 | 25 | 0 | 6/6 | 10/9 | 6–10 | N | 71 |
| 36 | S. alba | 0 | 27 | 46 | 27 | 0 | 6/6 | 10/9 | 6–10 | N | 33 |
| 37 | S. alba | 0 | 25 | 59 | 16 | 0 | 6/7 | 10/9 | 6–10 | N | 65 |
| 38 | S. alba | 0 | 24 | 33 | 43 | 0 | 6/7 | 10/8 | 6–10 | N | 63 |
| 39 | S. fragilis | 0 | 23 | 67 | 10 | 0 | 6/7 | 10/9 | 6–10 | N | 73 |
| 40 | S. fragilis | 0 | 19 | 63 | 18 | 0 | 6/7 | 10/9 | 6–10 | N | 63 |
| 41 | S. fragilis | 0 | 15 | 62 | 23 | 0 | 6/8 | 10/9 | 6–10 | L | 41 |
| 42 | S. fragilis × S. alba | 0 | 13 | 84 | 3 | 0 | 6/7 | 10/8 | 6–10 | A | 91 |
| 43 | S. fragilis | 0 | 13 | 81 | 6 | 0 | 7/8 | 9/8 | 7–9 | L | 34 |
| 44 | S. alba × S. fragilis | 0 | 10 | 53 | 27 | 0 | 6/8 | 10/9 | 6–10 | N | 31 |
| 45 | S. fragilis | 0 | 4 | 65 | 31 | 0 | 7/8 | 10/9 | 7–10 | S | 70 |
| 46 | S. alba | 0 | 4 | 30 | 66 | 0 | 8/9 | 10/9 | 8–10 | S | 69 |
| 47 | S. fragilis | 0 | 3 | 41 | 56 | 0 | 8/9 | 10/9 | 8–10 | A | 43 |
| 48 | S. fragilis | 0 | 3 | 56 | 39 | 2 | 7/8 | 11/10 | 7–11 | L | 94 |
| 49 | S. euxina | 0 | 26 | 46 | 26 | 2 | 7/8 | ? | 7–10 | L | 51 |
| 50 | S. fragilis | 0 | 19 | 54 | 23 | 4 | 7/8 | 10/9 | 7–10 | L | 39 |
| 51 | Salix | 0 | 0 | 30 | 70 | 0 | 8/9 | 10 | 8–10 | S | 15 |
| 52 | S. alba | 0 | 0 | 21 | 79 | 0 | 9 | 10 | 9–10 | N | 14 |
| 53 | Salix | 0 | 0 | 4 | 96 | 0 | 9 | 10 | 9–10 | S | 12 |
| 54 | Salix | 0 | 0 | 3 | 97 | 0 | 9 | 10 | 9–10 | N | 16 |
| 55 | S. alba | 0 | 0 | 25 | 74 | 1 | 8/9 | 11/10 | 8–11 | S | 55 |
| 56 | S. fragilis | 0 | 0 | 44 | 51 | 5 | 8/9 | 12/10 | 8–12 | S | 38 |
| 57 | S. alba | 0 | 0 | 14 | 79 | 7 | 9/9 | 11/10 | 9–11 | N | 22 |
| 58 | S. alba × S. fragilis | 0 | 0 | 10 | 88 | 2 | 8/9 | 11/10 | 8–11 | S | 26 |
| 59 | S. alba | 0 | 0 | 4 | 95 | 1 | 9/9 | 11/10 | 9–11 | N | 65 |
| 60 | S. alba | 0 | 0 | 2 | 84 | 14 | 9/10 | 12 | 9–12 | A | 58 |
| 61 | S. alba | 0 | 0 | 1 | 78 | 21 | 10/10 | 12/11 | 10–12 | N | 36 |
| 62 | S. alba | 0 | 0 | 4 | 46 | 50 | 10 | 12/11 | 10–12 | N | 12 |
| 63 | S. alba var. splendens | 0 | 0 | 0 | 93 | 7 | 10 | 12/11 | 10–12 | N | 14 |
| 64 | S. alba | 0 | 0 | 0 | 80 | 20 | 10 | 12 | 10–12 | A | 20 |
| 65 | S. alba | 0 | 0 | 0 | 71 | 29 | 10 | 12 | 10–12 | N | 12 |
| 66 | S. alba | 0 | 0 | 0 | 63 | 37 | 10 | 12 | 10–12 | N | 38 |
| No | Specimen | Percentage of Valves with Each Ovule Number | No. of Ovules per Ovary | Ovule Index | Trichomes | No of Ovaries | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 6 | 7 | 8 | 9 | 10 | 11 | 12 | Min/ Often | Max/ Often | |||||
| 67 | S. alba | 91 | 7 | 2 | 0 | 0 | 0 | 0 | 11/12 | 14 | 11–14 | N | 42 |
| 68 | S. alba | 2 | 5 | 78 | 15 | 0 | 0 | 0 | 13/16 | 17 | 13–17 | A | 48 |
| 69 | S. alba | 0 | 5 | 84 | 11 | 0 | 0 | 0 | 14/16 | 18 | 14–18 | S | 55 |
| 70 | S. alba | 0 | 24 | 71 | 5 | 0 | 0 | 0 | 15 | 17 | 15–17 | N | 19 |
| 71 | S. excelsa | 0 | 5 | 67 | 28 | 0 | 0 | 0 | 15 | 18 | 15–18 | N | 9 |
| 72 | S. excelsa | 0 | 0 | 14 | 73 | 9 | 3 | 1 | 16/17 | 21/18 | 16–21 | N | 54 |
| 73 | S. alba | 0 | 0 | 0 | 88 | 8 | 4 | 0 | 18 | 20 | 18–20 | N | 36 |
| 74 | S. alba | 0 | 0 | 0 | 62 | 20 | 9 | 9 | 18 | 21/19 | 18–21 | A | 17 |
| 75 | S. fragilis | 0 | 0 | 0 | 55 | 27 | 9 | 9 | 18/19 | 22/19 | 18–22 | A | 11 |
| 76 | S. alba | 0 | 0 | 0 | 0 | 26 | 48 | 26 | 20/21 | 24/23 | 20–24 | A | 27 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchenko, A.M.; Kuzovkina, Y.A. The Ovule Number Variation Provides New Insights into Taxa Delimitation in Willows (Salix subgen. Salix; Salicaceae). Plants 2023, 12, 497. https://doi.org/10.3390/plants12030497
Marchenko AM, Kuzovkina YA. The Ovule Number Variation Provides New Insights into Taxa Delimitation in Willows (Salix subgen. Salix; Salicaceae). Plants. 2023; 12(3):497. https://doi.org/10.3390/plants12030497
Chicago/Turabian StyleMarchenko, Alexander M., and Yulia A. Kuzovkina. 2023. "The Ovule Number Variation Provides New Insights into Taxa Delimitation in Willows (Salix subgen. Salix; Salicaceae)" Plants 12, no. 3: 497. https://doi.org/10.3390/plants12030497
APA StyleMarchenko, A. M., & Kuzovkina, Y. A. (2023). The Ovule Number Variation Provides New Insights into Taxa Delimitation in Willows (Salix subgen. Salix; Salicaceae). Plants, 12(3), 497. https://doi.org/10.3390/plants12030497