

Effects of Date Palm Waste Compost Application on Root Proteome Changes of Barley (Hordeum vulgare L.)

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Effect of Date Palm Waste Compost on Growth Parameters and Yield of Barley Crop
2.2. Identification of DAPs Using LC-MS/MS
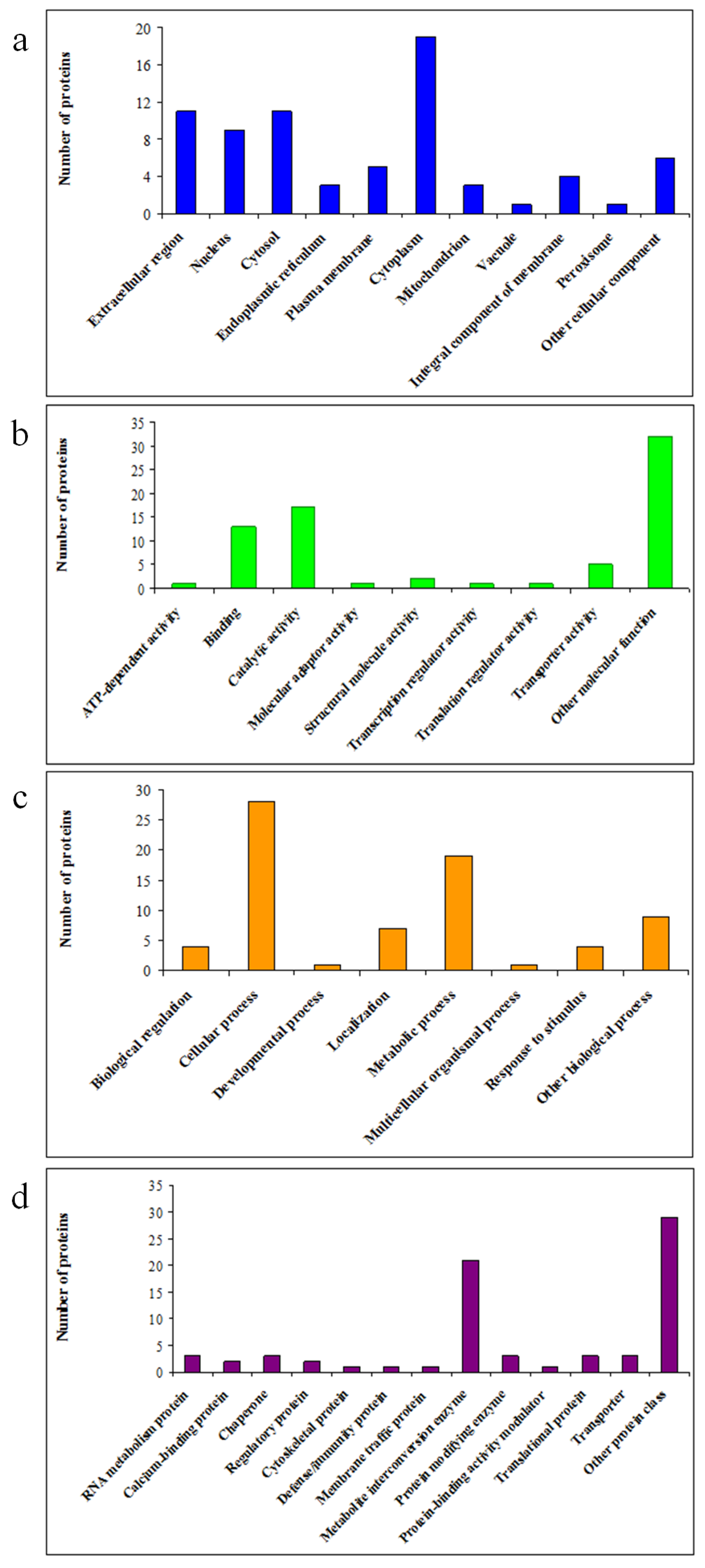
2.3. Functional Categories and Subcellular Localisation of DAPs
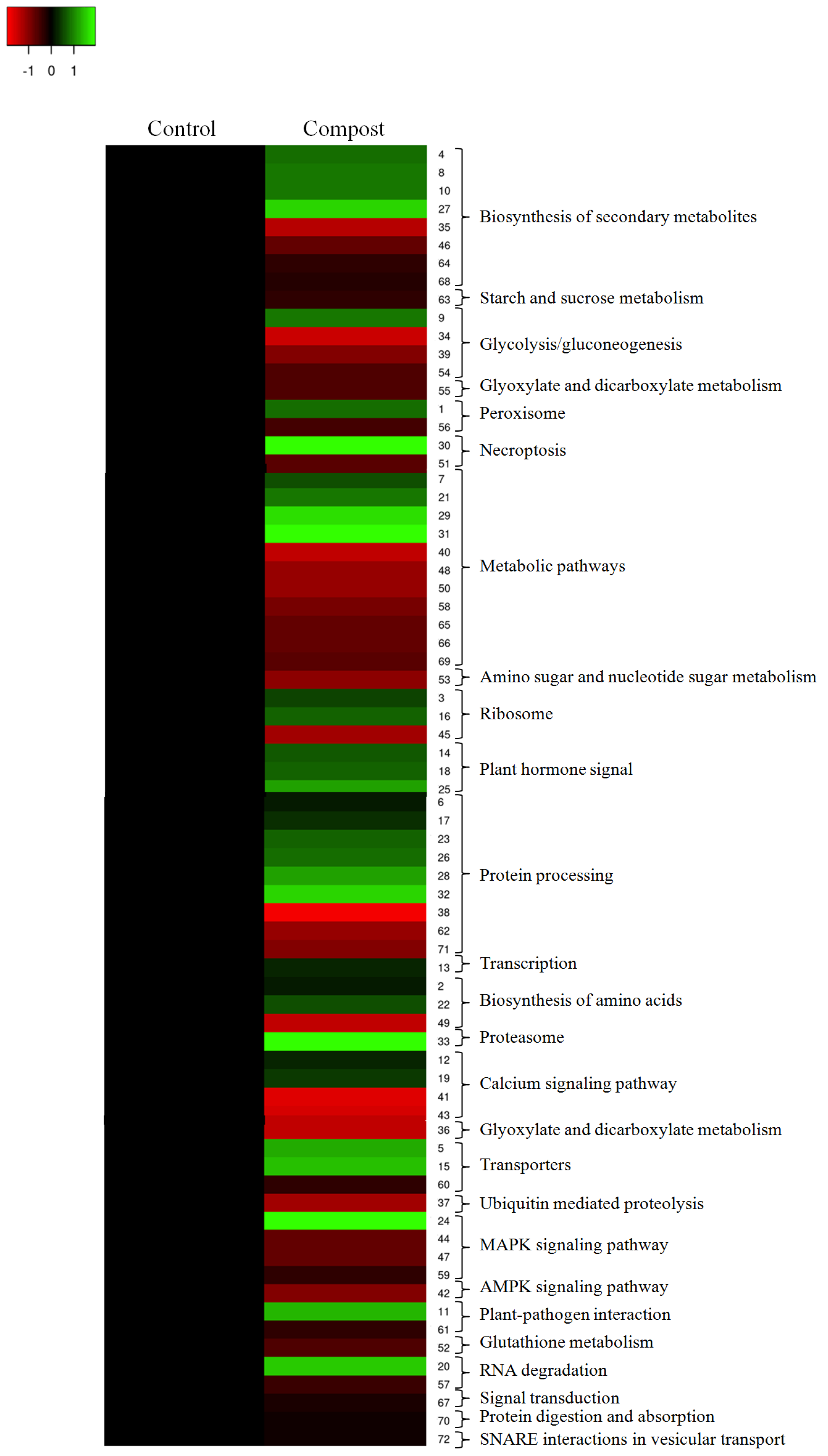
2.4. KEGG Pathway Enrichment Analysis of DAPs Induced by Date Palm Waste Compost Application in Barley Roots
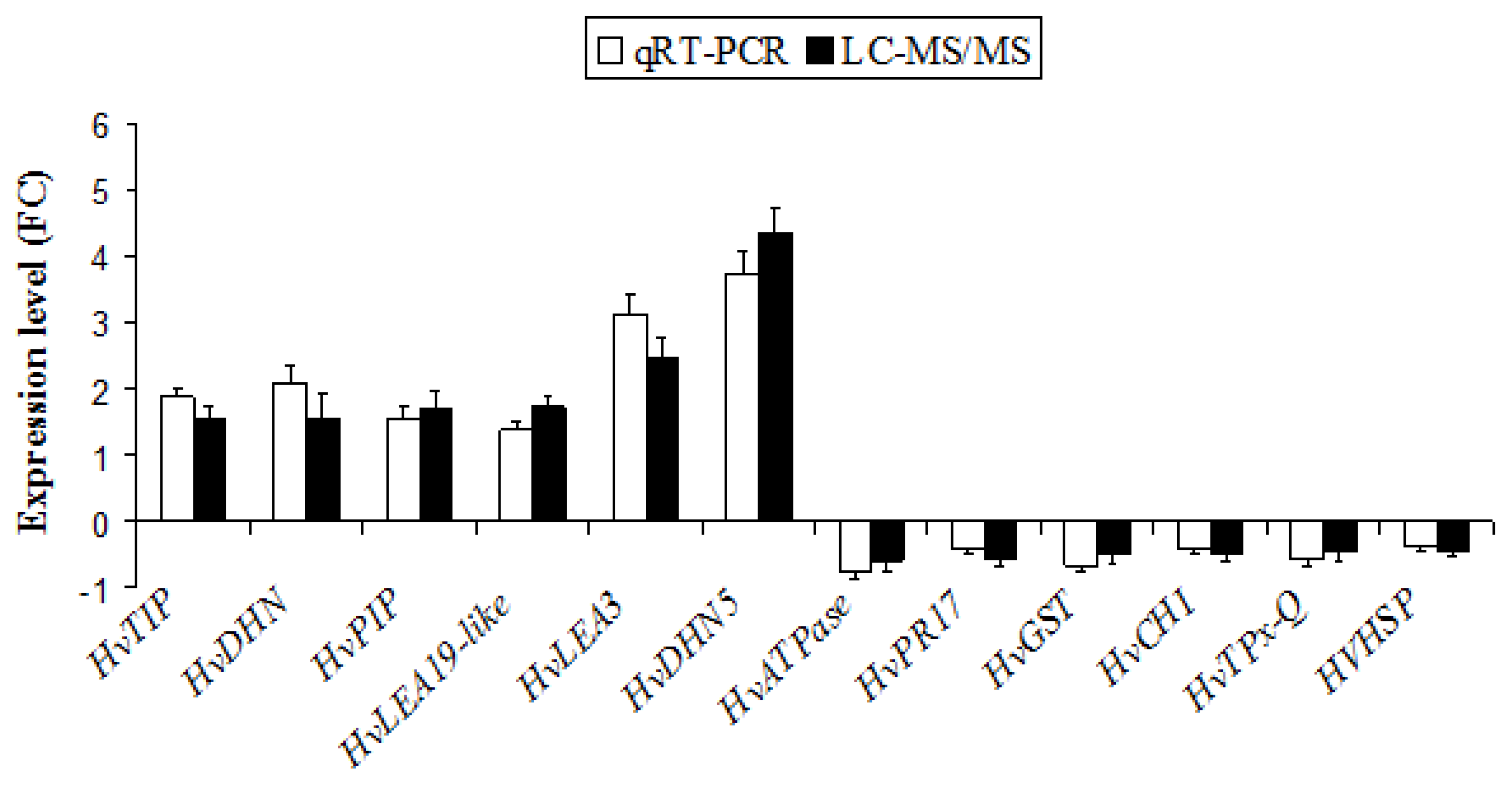
2.5. Validation of the Expression of DAPs in Barley Roots by Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
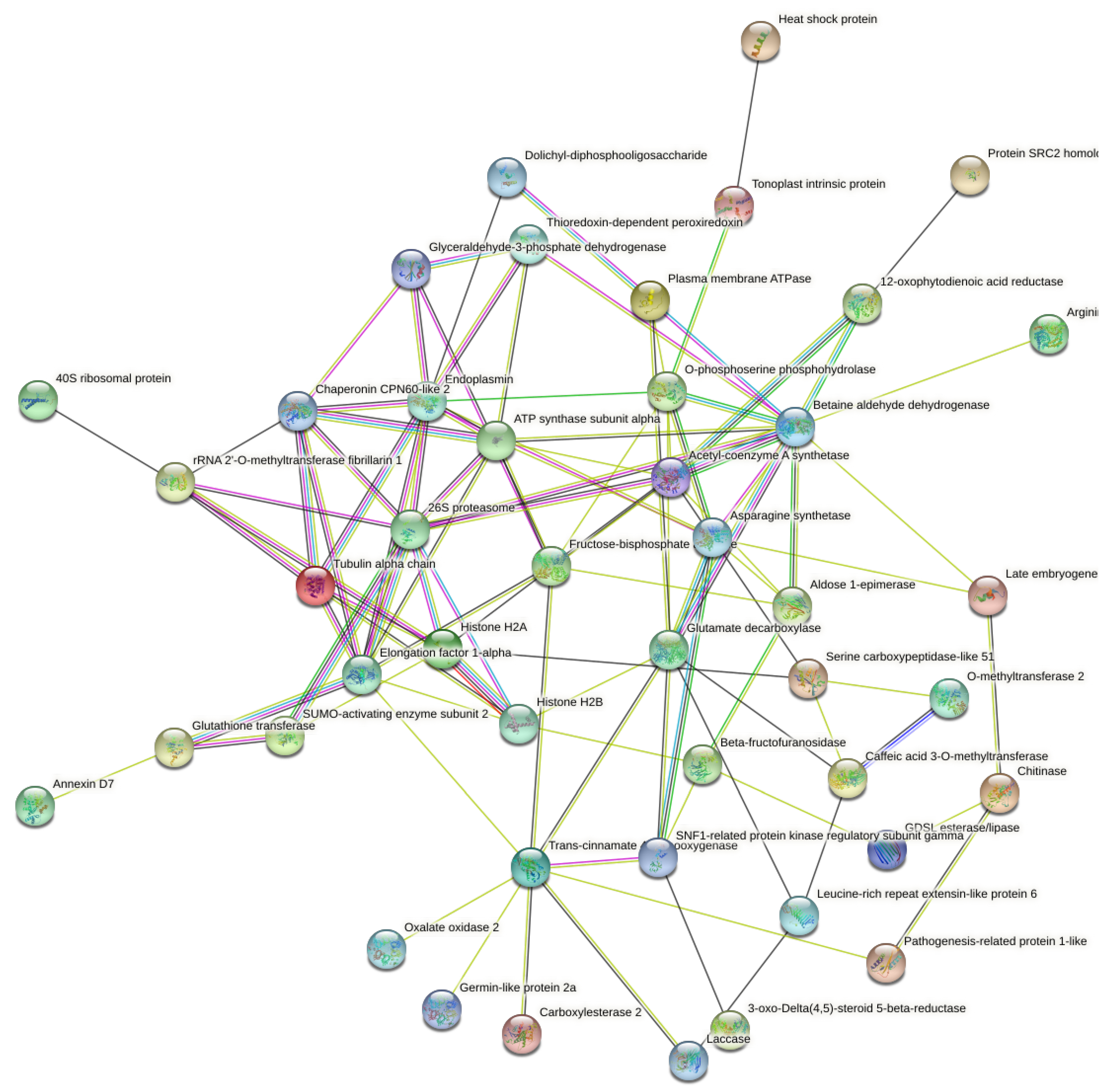
2.6. Putative Protein-Protein Interaction (PPI) Networks of DAPs to Date Palm Compost Waste Application
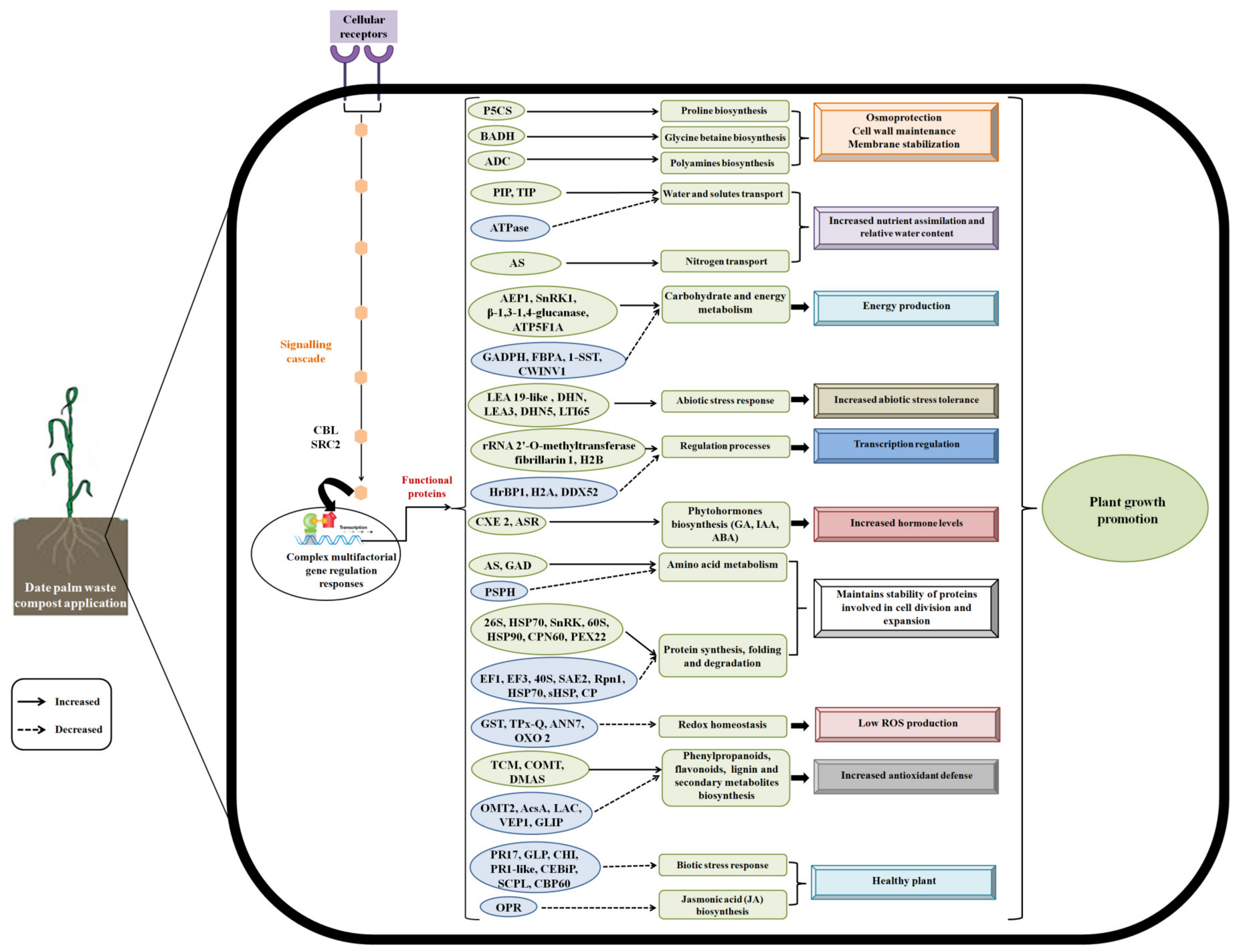
3. Discussion
3.1. Date Palm Waste Compost Promote the Growth and Development of Barley Plants
3.2. Date Palm Waste Compost Induced Changes in the Barley Root Proteomes
3.3. Detoxification Proteins
3.4. Plant Stress and Defense Proteins
3.5. Transport Proteins
3.6. Carbohydrate and Energy Metabolism Proteins
3.7. Amino Acids Metabolism Proteins
3.8. Folding, Synthesis and Degradation Proteins
3.9. Regulation Processes Proteins
3.10. Cytoskeleton Dynamics Proteins
3.11. Plant Hormone Metabolism Proteins
3.12. Secondary Metabolism Proteins
4. Materials and Methods
4.1. Plant Growth and Treatment with Date Palm Waste Compost
4.2. Extraction, Digestion and LC-MS/MS Analyses
4.3. Identification and Quantification of Proteins
4.4. Bioinformatics Analysis
4.5. Quantitative Real-Time PCR (qRT-PCR)
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alkharabsheh, H.M.; Seleiman, M.F.; Hewedy, O.A.; Battaglia, M.L.; Jalal, R.S.; Alhammad, B.A.; Schillaci, C.; Ali, N.; Al-Doss, A. Field crop responses and management strategies to mitigate soil salinity in modern agriculture: A review. Agronomy 2021, 11, 2299. [Google Scholar] [CrossRef]
- FAOSTAT. 2020. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 1 August 2022).
- Ho, T.T.K.; Tra, V.T.; Le, T.H.; Nguyen, N.K.Q.; Tran, C.S.; Nguyen, P.T.; Vo, T.D.H.; Thai, V.N.; Bui, X.T. Compost to improve sustainable soil cultivation and crop productivity. Case Stud. Chem. Environ. Eng. 2022, 6, 100211. [Google Scholar] [CrossRef]
- Heisey, S.; Ryals, R.; Maaz, T.M.; Nguyen, N.H. A single application of compost can leave lasting impacts on soil microbial community structure and alter cross-domain interaction networks. Front. Soil Sci. 2022, 2, 749212. [Google Scholar] [CrossRef]
- Darimani, H.S.; Adams, A.; Abudul Rahaman, A.B.; Nwang, J.; Yunus, R. Bio-compost as a soil supplement to improve growth and yield of tomato (Lycopersicum esculentum). J. Agric. Chem. Environ. 2022, 11, 67–82. [Google Scholar] [CrossRef]
- Liu, D.; Ding, Z.; Ali, E.F.; Kheir, A.M.S.; Eissa, M.A.; Ibrahim, O.H.M. Biochar and compost enhance soil quality and growth of roselle (Hibiscus sabdariffa L.) under saline conditions. Sci. Rep. 2021, 11, 8739. [Google Scholar] [CrossRef]
- Gaiotti, F.; Marcuzzo, P.; Belfiore, N.; Lovat, L.; Fornasier, F.; Tomasi, D. Influence of compost addition on soil properties, root growth and vine performances of Vitis vinifera cv. Cabernet sauvignon. Sci. Hortic. 2017, 225, 88–95. [Google Scholar] [CrossRef]
- Soobhany, N.; Mohee, R.; Garg, V.K. A comparative analysis of composts and vermicomposts derived from municipal solid waste for the growth and yield of green bean (Phaseolus vulgaris). Environ. Sci. Pollut. Res. 2017, 24, 11228–11239. [Google Scholar] [CrossRef]
- Rashtbari, M.; Ali, A.H.; Ghorchiani, M. Effect of vermicompost and municipal solid waste compost on growth and yield of canola under drought stress conditions. Commun. Soil Sci. Plant Anal. 2020, 51, 2215–2222. [Google Scholar] [CrossRef]
- Omara, A.E.D.; Hafez, E.M.; Osman, H.S.; Rashwan, E.; El-Said, M.A.A.; Alharbi, K.; Abd El-Moneim, D.; Gowayed, S.M. Collaborative Impact of Compost and Beneficial Rhizobacteria on Soil Properties, Physiological Attributes, and Productivity of Wheat Subjected to Deficit Irrigation in Salt Affected Soil. Plants 2022, 11, 877. [Google Scholar] [CrossRef]
- Ghouili, E.; Hidri, Y.; Cheikh M’Hamed, H.; Somenahally, A.; Xue, Q.; El Akram Znaïdi, I.; Jebara, M.; Nefissi Ouertani, R.; Muhovski, Y.; Riahi, J.; et al. Date palm waste compost promotes plant growth and nutrient transporter genes expression in barley (Hordeum vulgare L.). S. Afr. J. Bot. 2022, 149, 247–257. [Google Scholar] [CrossRef]
- Ait Rahou, Y.; Douira, A.; Tahiri, A.I.; Cherkaoui, E.M.; Benkirane, R.; Meddich, A. Application of plant growth-promoting rhizobacteria combined with compost as a management strategy against Verticillium dahliae in tomato. Can. J. Plant Pathol. 2022, 44, 806–827. [Google Scholar] [CrossRef]
- Franzoni, G.; Cocetta, G.; Prinsi, B.; Ferrante, A.; Espen, L. Biostimulants on crops: Their impact under abiotic stress conditions. Horticulturae 2022, 8, 189. [Google Scholar] [CrossRef]
- Tammam, A.; El-Aggan, W.; Helaly, A.; Badr, G.; El-Dakak, R. Proteomics and photosynthetic apparatus response to vermicompost attenuation of salinity stress Vicia faba leaves. Acta Physiol. Plant 2023, 45, 17. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, S.; Liu, K.; Wang, S.; Huang, L.; Guo, L. Proteomics: A powerful tool to study plant responses to biotic stress. Plant Methods 2019, 15, 135. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, R. Analysis of the barley malt rootlet proteome. Int. J. Mol. Sci. 2020, 21, 179. [Google Scholar] [CrossRef] [Green Version]
- Qin, Q.; Liu, J.; Hu, S.; Dong, J.; Yu, J.; Fang, L.; Huang, S.; Wang, L. Comparative proteomic analysis of different barley cultivars during seed germination. J. Cereal Sci. 2021, 102, 103357. [Google Scholar] [CrossRef]
- Rekaby, S.A.; Awad, M.Y.M.; Hegab, S.A.; Eissa, M.A. Effect of some organic amendments on barley plants under saline condition. J. Plant Nutr. 2020, 43, 1840–1851. [Google Scholar] [CrossRef]
- Miezah, K.; Ofosu-Anim, J.; Budu, G.K.O.; Enu-Kwesi, L.; Cofie, O. Isolation and identification of some plant growth promoting substances in compost and co-compost. Int. J. Virol. 2008, 4, 30–40. Available online: https://scialert.net/abstract/?doi=ijv.2008.30.40 (accessed on 12 November 2022). [CrossRef] [Green Version]
- Mutlu, A. The effect of organic fertilizers on grain yield and some yield components of barley (Hordeum vulgare L.). Fresenius Environ. Bull. 2020, 29, 10840–10846. [Google Scholar]
- Mohamed, M.F.; Thalooth, A.T.; Elewa, T.A.; Ahmed, A.G. Yield and nutrient status of wheat plants (Triticum aestivum) as affected by sludge, compost, and biofertilizers under newly reclaimed soil. Bull. Natl. Res. Cent. 2019, 43, 31. [Google Scholar] [CrossRef]
- Agegnehu, G.; Tsigie, A.; Tesfaye, A. Evaluation of crop residue retention, compost and inorganic fertilizer application on barley productivity and soil chemical properties in the central Ethiopian highlands. Ethiop. J. Agric. Sci. 2012, 22, 45–61. [Google Scholar]
- Roomi, S.; Masi, A.; Conselvan, G.B.; Trevisan, S.; Quaggiotti, S.; Pivato, M.; Arrigoni, G.; Yasmin, T.; Carletti, P. Protein profiling of Arabidopsis roots treated with humic substances: Insights into the metabolic and interactome networks. Front. Plant Sci. 2018, 9, 1812. [Google Scholar] [CrossRef] [PubMed]
- Zulfikar Ali, S.; Sandhya, V.; Grover, M.; Rao Linga, V.; Bandi, V. Effect of inoculation with a thermotolerant plant growth promoting Pseudomonas putida strain AKMP7 on growth of wheat (Triticum spp.) under heat stress. J. Plant Interact. 2011, 6, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Suleiman, R.K.; Iali, W.; El Ali, B.; Umoren, S.A. New constituents from the leaves of date palm (Phoenix dactylifera L.) of Saudi origin. Molecules 2021, 26, 4192. [Google Scholar] [CrossRef]
- Ebinezer, L.B.; Franchin, C.; Trentin, A.R.; Carletti, P.; Trevisan, S.; Agrawal, G.K.; Rakwal, R.; Quaggiotti, S.; Arrigoni, G.; Masi, A. Quantitative proteomics of maize roots treated with a protein hydrolysate: A comparative study with transcriptomics highlights the molecular mechanisms responsive to biostimulants. J. Agric. Food Chem. 2020, 68, 7541–7553. [Google Scholar] [CrossRef]
- Copley, T.; Bayen, S.; Jabaji, S. Biochar amendment modifies expression of soybean and rhizoctonia solani genes leading to increased severity of rhizoctonia foliar blight. Front. Plant Sci. 2017, 8, 221. [Google Scholar] [CrossRef] [Green Version]
- Campobenedetto, C.; Mannino, G.; Beekwilder, J.; Contartese, V.; Karlova, R.; Bertea, C.M. The application of a biostimulant based on tannins affects root architecture and improves tolerance to salinity in tomato plants. Sci. Rep. 2021, 11, 354. [Google Scholar] [CrossRef]
- Ait-El-Mokhtar, M.; Baslam, M.; Ben-Laouane, R.; Anli, M.; Boutasknit, A.; Mitsui, T.; Wahbi, S.; Meddich, A. Alleviation of Detrimental Effects of Salt Stress on Date Palm (Phoenix dactylifera L.) by the Application of Arbuscular Mycorrhizal Fungi and/or Compost. Front. Sustain. Food Syst. 2020, 4, 131. [Google Scholar] [CrossRef]
- Yacoubi, I.; Gadaleta, A.; Mathlouthi, N.; Hamdi, K.; Giancaspro, A. Abscisic acid-stress-ripening genes involved in plant response to high salinity and water deficit in durum and common wheat. Front. Plant Sci. 2022, 13, 789701. [Google Scholar] [CrossRef]
- Gupta, K.; Wani, S.H.; Razzaq, A.; Skalicky, M.; Samantara, K.; Gupta, S.; Pandita, D.; Goel, S.; Grewal, S.; Hejnak, V.; et al. Abscisic acid: Role in fruit development and ripening. Front. Plant Sci. 2022, 13, 817500. [Google Scholar] [CrossRef]
- de Azevedo, I.G.; Olivares, F.L.; Ramos, A.C.; Bertolazi, A.A.; Canellas, L.P. Humic acids and herbaspirillum seropedicae change the extracellular H+ fux and gene expression in maize roots seedlings. Chem. Biol. Technol. Agric. 2019, 6, 8. [Google Scholar] [CrossRef]
- Abdul Aziz, M.; Sabeem, M.; Mullath, S.K.; Brini, F.; Masmoudi, K. Plant group II LEA proteins: Intrinsically disordered structure for multiple functions in response to environmental stresses. Biomolecules 2021, 11, 1662. [Google Scholar] [CrossRef] [PubMed]
- Wetzler, D.E.; Fuchs Wightman, F.; Bucci, H.A.; Rinaldi, J.; Caramelo, J.J.; Iusem, N.D.; Ricardi, M.M. Conformational plasticity of the intrinsically disordered protein ASR1 modulates its function as a drought stress-responsive gene. PLoS ONE 2018, 13, e0202808. [Google Scholar] [CrossRef] [PubMed]
- Bellini, A.; Ferrocino, I.; Cucu, M.A.; Pugliese, M.; Garibaldi, A.; Gullino, M.L. A compost treatment acts as a suppressive agent in Phytophthora capsici-Cucurbita pepo pathosystem by modifying the rhizosphere microbiota. Front. Plant Sci. 2020, 11, 885. [Google Scholar] [CrossRef] [PubMed]
- Maynard, D.; Kumar, V.; Sproï, J.; Dietz, K.J. 12-oxophytodienoic acid reductase 3 (OPR3) functions as NADPH-dependent α,β-ketoalkene reductase in detoxification and monodehydroascorbate reductase in redox homeostasis. Plant Cell Physiol. 2020, 61, 584–595. [Google Scholar] [CrossRef]
- Liu, L.; Sonbol, F.M.; Huot, B.; Gu, Y.; Withers, J.; Mwimba, M.; Yao, J.; He, S.Y.; Dong, X. Salicylic acid receptors activate jasmonic acid signalling through a non-canonical pathway to promote effector-triggered immunity. Nat. Commun. 2016, 7, 13099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, Y.H.; Zamora, O.R.; Troncoso-Rojas, R.; Tiznado-Hernández, M.E.; Báez-Flores, M.E.; Carvajal-Millan, E.; Rascón-Chu, A. Toward understanding the molecular recognition of fungal chitin and activation of the plant defense mechanism in horticultural crops. Molecules 2021, 26, 6513. [Google Scholar] [CrossRef]
- Kulcsár, L.; Flipphi, M.; Jónás, A.; Sándor, E.; Fekete, E.; Karaffa, L. Identification of a mutarotase gene involved in D-galactose utilization in Aspergillus nidulans. FEMS Microbiol. Lett. 2017, 364, fnx202. [Google Scholar] [CrossRef]
- Perrot, T.; Pauly, M.; Ramírez, V. Emerging roles of β-Glucanases in plant development and adaptative responses. Plants 2022, 11, 1119. [Google Scholar] [CrossRef]
- Han, M.; Xu, M.; Su, T.; Wang, S.; Wu, L.; Feng, J.; Ding, C. Transcriptome analysis reveals critical genes and pathways in carbon metabolism and ribosome biogenesis in poplar fertilized with glutamine. Int. J. Mol. Sci. 2022, 23, 9998. [Google Scholar] [CrossRef]
- Crozet, P.; Margalha, L.; Confraria, A.; Rodrigues, A.; Martinho, C.; Adamo, M.; Elias, C.A.; Baena-González, E. Mechanisms of regulation of SNF1/AMPK/SnRK1 protein kinases. Front. Plant Sci. 2014, 5, 190. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, T.B.; Chapman, L.M. The importance of thiamine (vitamin B1) in plant health: From crop yield to biofortification. J. Biol. Chem. 2020, 295, 12002–12013. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, U.K.; Islam, M.N.; Siddiqui, M.N.; Rahman Khan, M.A. Understanding the roles of osmolytes for acclimatizing plants to changing environment: A review of potential mechanism. Plant Signal. Behav. 2021, 16, e1913306. [Google Scholar] [CrossRef]
- Othibeng, K.; Nephali, L.; Myoli, A.; Buthelezi, N.; Jonker, W.; Huyser, J.; Tugizimana, F. Metabolic circuits in sap extracts reflect the effects of a microbial biostimulant on maize metabolism under drought conditions. Plants 2022, 11, 510. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Xi, Y.; Zhang, Z.; Mohammadi, M.A.; Joshi, J.; Borza, T.; Wang-Pruski, G. Effects of phosphite as a plant biostimulant on metabolism and stress response for better plant performance in Solanum tuberosum. Ecotoxicol. Environ. Saf. 2021, 210, 111873. [Google Scholar] [CrossRef] [PubMed]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.T.; Chen, H.H.W.; Feun, L.G.; Savaraj, N. Targeting the proline-glutamine-asparagines-arginine metabolic axis in amino acid starvation cancer therapy. Pharmaceuticals 2021, 14, 72. [Google Scholar] [CrossRef]
- Kavi Kishor, P.B.; Hima Kumari, P.; Sunita, M.S.L.; Sreenivasulu, N. Role of proline in cell wall synthesis and plant development and its implications in plant ontogeny. Front. Plant Sci. 2015, 6, 544. [Google Scholar] [CrossRef] [Green Version]
- Oddy, J.; Rafan, S.; Wilkinson, M.D.; Elmore, J.S.; Halford, N.G. Stress, nutrients and genotype: Understanding and managing asparagine accumulation in wheat grain. CABI Agric. Biosci. 2020, 1, 10. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, L.; Zhao, W.; Fu, L.; Han, Y.; Wang, K.; Yan, L.; Li, Y.; Zhang, X.H.; Min, D.H. Genome-wide analysis of the serine carboxypeptidase-like protein family in Triticum aestivum reveals TaSCPL184-6D is involved in abiotic stress response. BMC Genom. 2021, 22, 350. [Google Scholar] [CrossRef]
- Byrne, M.E. A role for the ribosome in development. Trends Plant Sci. 2009, 14, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Ayoola, M.B.; Nakamya, M.F.; Shack, L.A.; Park, S.; Lim, J.; Lee, J.H.; Ross, M.K.; Eoh, H.; Nanduri, B. SP_0916 is an arginine decarboxylase that catalyzes the synthesis of agmatine, which is critical for capsule biosynthesis in Streptococcus pneumoniae. Front. Microbiol. 2020, 11, 578533. [Google Scholar] [CrossRef] [PubMed]
- Marchesani, F.; Zangelmi, E.; Bruno, S.; Bettati, S.; Peracchi, A.; Campanini, B. A novel assay for phosphoserine phosphatase exploiting serine acetyltransferase as the coupling enzyme. Life 2021, 11, 485. [Google Scholar] [CrossRef] [PubMed]
- Freitas, I.S.; Trennepohl, B.I.; Acioly, T.M.S.; Conceição, V.J.; Mello, S.C.; Dourado Neto, D.; Kluge, R.A.; Azevedo, R.A. Exogenous application of L-arginine improves protein content and increases yield of Pereskia aculeata Mill. grown in soilless media container. Horticulturae 2022, 8, 142. [Google Scholar] [CrossRef]
- González-Hernández, A.I.; Scalschi, L.; Vicedo, B.; Marcos-Barbero, E.L.; Morcuende, R.; Camañes, G. Putrescine: A key metabolite involved in plant development, tolerance and resistance responses to stress. Int. J. Mol. Sci. 2022, 23, 2971. [Google Scholar] [CrossRef]
- Xia, J.; Yamaji, N.; Che, J.; Shen, R.F.; Ma, J.F. Normal root elongation requires arginine produced by argininosuccinate lyase in rice. Plant J. 2014, 78, 215–226. [Google Scholar] [CrossRef]
- Salih, K.J.; Duncan, O.; Li, L.; O’Leary, B.; Fenske, R.; Trösch, J.; Millar, A.H. Impact of oxidative stress on the function, abundance, and turnover of the arabidopsis 80S cytosolic ribosome. Plant J. 2020, 103, 128–139. [Google Scholar] [CrossRef]
- Xu, B.; Liu, L.; Song, G. Functions and regulation of translation elongation factors. Front. Mol. Biosci. 2022, 8, 816398. [Google Scholar] [CrossRef]
- Hummel, M.; Cordewener, J.H.G.; de Groot, J.C.M.; Smeekens, S.; America, A.H.P.; Hanson, J. Dynamic protein composition of Arabidopsis thaliana cytosolic ribosomes in response to sucrose feeding as revealed by label free MSE proteomics. Proteomics 2012, 12, 1024–1038. [Google Scholar] [CrossRef]
- Nunes, R.O.; Domiciano, G.A.; Alves, W.S.; Amaral Melo, A.C.; Sousa Nogueira, F.C.; Canellas, L.P.; Olivares, F.L.; Zingali, R.B.; Soares, M.R. Evaluation of the effects of humic acids on maize root architecture by label-free proteomics analysis. Sci. Rep. 2019, 9, 12019. [Google Scholar] [CrossRef] [Green Version]
- Kurepa, J.; Smalle, J.A. Structure, function and regulation of plant proteasomes. Biochimie 2008, 90, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Gao, T.; Hu, J.; Zhao, L.; Yu, C.; Ma, F. Research advances in function and regulation mechanisms of plant small heat shock proteins (sHSPs) under environmental stresses. Sci. Total Environ. 2022, 825, 154054. [Google Scholar] [CrossRef] [PubMed]
- Cocetta, G.; Landoni, M.; Pilu, R.; Repiso, C.; Nolasco, J.; Alajarin, M.; Ugena, L.; Levy, C.C.B.; Scatolino, G.; Villa, D.; et al. Priming treatments with biostimulants to cope the short-term heat stress response: A transcriptomic profile evaluation. Plants 2022, 11, 1130. [Google Scholar] [CrossRef] [PubMed]
- Azevedo-Favory, J.; Gaspin, C.; Ayadi, L.; Montacié, C.; Marchand, V.; Jobet, E.; Rompais, M.; Carapito, C.; Motorin, Y.; Sáez-Vásquez, J. Mapping rRNA 2′-O-methylations and identification of C/D snoRNAs in Arabidopsis thaliana plants. RNA Biol. 2021, 18, 1760–1777. [Google Scholar] [CrossRef]
- Asensi-Fabado, M.A.; Amtmann, A.; Perrella, G. Plant responses to abiotic stress: The chromatin context of transcriptional regulation. Biochim. Biophys. Acta (BBA) 2017, 1860, 106–122. [Google Scholar] [CrossRef] [Green Version]
- Pichereaux, C.; Laurent, E.A.; Gargaros, A.; Viudes, S.; Durieu, C.; Lamaze, T.; Grieu, P.; Burlet-Schiltz, O. Analysis of durum wheat proteome changes under marine and fungal biostimulant treatments using large-scale quantitative proteomics: A useful dataset of durum wheat proteins. J. Proteom. 2019, 200, 28–39. [Google Scholar] [CrossRef]
- Lloyd, C.; Chan, J. The parallel lives of microtubules and cellulose microfibrils. Curr. Opin. Plant Biol. 2008, 11, 641–646. [Google Scholar] [CrossRef]
- Rui, C.; Peng, F.; Fan, Y.; Zhang, Y.; Zhang, Z.; Xu, N.; Zhang, H.; Wang, J.; Li, S.; Yang, T.; et al. Genome-wide expression analysis of carboxylesterase (CXE) gene family implies GBCXE49 functional responding to alkaline stress in cotton. BMC Plant Biol. 2022, 22, 194. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Omary, M.; Hu, Y.; Doron, O.; Hoermayer, L.; Chen, Q.; Megides, O.; Chekli, O.; Ding, Z.; Friml, J.; et al. Cell kinetics of auxin transport and activity in Arabidopsis root growth and skewing. Nat. Commun. 2021, 12, 1657. [Google Scholar] [CrossRef]
- Bjorklund, S.; Antti, H.; Uddestrand, I.; Moritz, T.; Sundberg, B. Cross-talk between gibberellin and auxin in development of Populus wood: Gibberellin stimulates polar auxin transport and has a common transcriptome with auxin. Plant J. 2007, 52, 499–511. [Google Scholar] [CrossRef]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lephatsi, M.; Nephali, L.; Meyer, V.; Piater, L.A.; Buthelezi, N.; Dubery, I.A.; Opperman, H.; Brand, M.; Huyser, J.; Tugizimana, F. Molecular mechanisms associated with microbial biostimulant-mediated growth enhancement, priming and drought stress tolerance in maize plants. Sci. Rep. 2022, 12, 10450. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, B.; Wang, L.; Ali, S.; Guo, Y.; Liu, J.; Wang, J.; Xie, L.; Zhang, Q. Genome-wide identification and characterization of caffeic acid o-methyltransferase gene family in soybean. Plants 2021, 10, 2816. [Google Scholar] [CrossRef] [PubMed]
- Faurobert, M.; Pelpoir, E.; Chaïb, J. Phenol extraction of proteins for proteomic studies of recalcitrant plant tissues. Methods Mol. Biol. 2007, 355, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Belouah, I.; Benard, C.; Denton, A.; Blein-Nicolas, M.; Balliau, T.; Teyssier, E.; Gallusci, P.; Bouchez, O.; Usadel, B.; Zivy, M.; et al. Transcriptomic and proteomic data in developing tomato fruit. Data Brief 2020, 28, 105015. [Google Scholar] [CrossRef]
- Balliau, T.; Duruflé, H.; Blanchet, N.; Blein-Nicolas, M.; Langlade, N.B.; Zivy, M. Proteomic data from leaves of twenty-four sunflower genotypes under water deficit. Oilseeds Fats Crops Lipids (OCL) 2021, 28, 12. [Google Scholar] [CrossRef]
- Langella, O.; Valot, B.; Balliau, T.; Blein-Nicolas, M.; Bonhomme, L.; Zivy, M. X!TandemPipeline: A tool to manage sequence redundancy for protein inference and phosphosite identification. J. Proteome Res. 2017, 16, 494–503. [Google Scholar] [CrossRef]
- Valot, B.; Langella, O.; Nano, E.; Zivy, M. MassChroQ: A versatile tool for mass spectrometry quantification. Proteomics 2011, 11, 3572–3577. [Google Scholar] [CrossRef] [Green Version]
- Belouah, I.; Blein-Nicolas, M.; Balliau, T.; Gibon, Y.; Zivy, M.; Colombié, S. Peptide filtering differently affects the performances of XIC-based quantification methods. J. Proteom. 2019, 193, 131–141. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Jensen, L.J. STRING v11: Protein_protein association networks with increased coverage supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Report. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H.J. Primer3 on the WWW for general users and for biologist programmers. In Bioinformatics Methods and Protocols: Methods in Molecular Biology; Krawetz, S., Misener, S., Eds.; Humana Press: Totowa, NJ, USA, 2000; pp. 365–386. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Shoot Length (cm) | Root Length (cm) | Shoot Dry Weight (g) | Root Dry Weight (g) | Grain Yield (t ha−1) |
|---|---|---|---|---|---|
| Control | 32.00 ± 2.00 b | 14.83 ± 1.75 a | 2.78 ± 0.13 b | 1.18 ± 0.09 b | 2.89 ± 0.54 b |
| Compost | 49.66 ± 1.52 a | 16.90 ± 2.12 a | 5.99 ± 0.25 a | 1.44 ± 0.07 a | 4.33 ± 0.67 a |
| No | Protein Name | Function | Regulated Type | Fold Change | Protein Accession | KEGG Pathway |
|---|---|---|---|---|---|---|
| 1 | Peroxisome biogenesis protein 22-like (PEX22) | Protein translocation | Up | 1.50 | F2DXG1 | Peroxisome |
| 2 | Delta-1-pyrroline-5-carboxylate synthase (P5CS) | Amino acid metabolism | Up | 1.51 | A0A224MLP5 | Biosynthesis of amino acids |
| 3 | 60S ribosomal protein L7a (60S) | Protein biosynthesis | Up | 1.51 | F2DE13 | Ribosome |
| 4 | Trans-cinnamate 4-monooxygenase (TCM) | Flavonoid biosynthesis | Up | 1.54 | F2E6M8 | Biosynthesis of secondary metabolites |
| 5 | Tonoplast intrinsic protein (TIP) | Solute transport | Up | 1.55 | D2KZ38 | Transporters |
| 6 | Dehydrin | Stress response | Up | 1.56 | M0UW32 | Protein processing |
| 7 | Betaine aldehyde dehydrogenase (BADH) | Glycine betaine biosynthesis | Up | 1.59 | Q94IC0 | Metabolic pathways |
| 8 | Caffeic acid 3-O-methyltransferase (COMT) | Lignin biosynthesis | Up | 1.60 | F2D692 | Biosynthesis of secondary metabolites |
| 9 | Aldose 1-epimerase (AEP 1) | Galactose metabolism | Up | 1.61 | M0X1Y4 | Glycolysis/gluconeogenesis |
| 10 | Deoxymugineic acid synthase 1 (DMAS) | Phytosiderophore biosynthesis | Up | 1.61 | F2DHN0 | Biosynthesis of secondary metabolites |
| 11 | Calcium-binding protein CBP-like (CBL) | Multi-process regulation | Up | 1.64 | A0A287NLZ0 | Plant-pathogen interaction |
| 12 | Annexin D7 (ANN7) | Stress response | Up | 1.64 | A0A287UPG1 | Calcium signaling pathway |
| 13 | La protein 1-like | RNA biosynthesis transcription | Up | 1.66 | F2DIB4 | Transcription |
| 14 | Low-temperature-induced 65 kDa protein-like isoform X1 (LTI65) | Stress response | Up | 1.67 | F2D3S4 | Plant hormone signal |
| 15 | PIP aquaporin isoform (PIP) | Solute transport | Up | 1.70 | A0A287IVR7 | Transporters |
| 16 | rRNA 2′-O-methyltransferase fibrillarin 1 | Protein biosynthesis | Up | 1.71 | F2E7G3 | Ribosome |
| 17 | Late embryogenesis abundant protein 19-like (LEA 19-like) | Stress response | Up | 1.72 | F2CUJ9 | Protein processing |
| 18 | Carboxylesterase 2 (CXE 2) | Hydrolysis | Up | 1.79 | A0A287TVH9 | Plant hormone signal |
| 19 | Protein SRC2 homolog (SRC2) | Signal transduction | Up | 1.80 | F2EAD0 | Calcium signaling pathway |
| 20 | Chaperonin CPN60-like 2 (CPN60) | Protein homeostasis | Up | 1.80 | F2CZD9 | RNA degradation |
| 21 | Arginine decarboxylase (ADC) | Putrescine biosynthesis | Up | 1.92 | M0XCI1 | Metabolic pathways |
| 22 | Asparagine synthetase (AS) | Amino acid metabolism | Up | 2.06 | A0A287P6Q4 | Biosynthesis of amino acids |
| 23 | Endoplasmin (HSP90) | Protein homeostasis | Up | 2.27 | P36183 | Protein processing |
| 24 | SNF1-related protein kinase regulatory subunit gamma-1 (SnRK) | Protein modification | Up | 2.30 | M0XDA4 | MAPK signaling pathway |
| 25 | Abcissic acid stress and ripening (ASR) | Stress response | Up | 2.41 | A0A1Z3GD05 | Plant hormone signal |
| 26 | Late embryogenesis abundant protein (LEA3) | Stress response | Up | 2.44 | B5TWC9 | Protein processing |
| 27 | Glutamate decarboxylase (GAD) | Amino acid metabolism | Up | 2.89 | F2E7E5 | Biosynthesis of secondary metabolites |
| 28 | Embryonic protein DC-8-like isoform X1 | Embryogenesis | Up | 3.26 | A0A287PP23 | Protein processing |
| 29 | (1-3,1-4)-beta-D-glucanase | Hydrolysis | Up | 3.33 | Q02345 | Metabolic pathways |
| 30 | Histone H2B | Chromatin organisation | Up | 3.72 | A0A287T8I9 | Necroptosis |
| 31 | Tubulin alpha chain | Cytoskeleton organisation | Up | 4.09 | F2E847 | Metabolic pathways |
| 32 | Dehydrin (DHN5) | Stress response | Up | 4.33 | Q40042 | Protein processing |
| 33 | 26S proteasome non-ATPase regulatory subunit 6 | Protein homeostasis | Up | 5.59 | F2DJW8 | Proteasome |
| 34 | Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | Carbohydrate degradation | Down | 0.25 | F2D714 | Glycolysis/gluconeogenesis |
| 35 | GDSL esterase/lipase (GLIP) | Hydrolysis | Down | 0.28 | A0A287TDE4 | Biosynthesis of secondary metabolites |
| 36 | Oxalate oxidase 2 (OXO 2) | H2O2 generation | Down | 0.30 | P45851 | Glyoxylate and dicarboxylate metabolism |
| 37 | SUMO-activating enzyme subunit 2 (SAE2) | Protein homeostasis | Down | 0.34 | A0A287HPI7 | Ubiquitin mediated proteolysis |
| 38 | Harpin binding protein 1 (HrBP1) | RNA-binding protein | Down | 0.35 | Q5QJB5 | Protein processing |
| 39 | Fructose-bisphosphate aldolase (FBPA) | Gluconeogenesis | Down | 0.37 | F2ELD1 | Glycolysis/gluconeogenesis |
| 40 | Dolichyl-diphosphooligosaccharide--protein glycosyltransferase subunit 1 (Rpn1) | Protein modification | Down | 0.38 | M0Z968 | Metabolic pathways |
| 41 | Plasma membrane ATPase | Solute transport | Down | 0.39 | F2DW68 | Calcium signaling pathway |
| 42 | Elongation factor 1-alpha (EF1) | Protein biosynthesis | Down | 0.40 | F2D525 | AMPK signaling pathway |
| 43 | Calmodulin-binding protein 60 B-like isoform X2 (CBP60) | Stress response | Down | 0.41 | F2DIS1 | Calcium signaling pathway |
| 44 | Pathogenesis related protein (PR17) | Plant defense responses | Down | 0.44 | A7YA66 | MAPK signaling pathway |
| 45 | 40S ribosomal protein SA (40S) | Protein biosynthesis | Down | 0.45 | F2D7E4 | Ribosome |
| 46 | Serine carboxypeptidase-like 51 (SCPL) | Hydrolysis | Down | 0.45 | A0A287WND1 | Biosynthesis of secondary metabolites |
| 47 | Pathogenesis-related protein 1-like (PR1-like) | Plant defense responses | Down | 0.46 | A0A287VZQ8 | MAPK signaling pathway |
| 48 | O-methyltransferase 2 (OMT2) | Aromatic compound biosynthetic process | Down | 0.47 | A0A287PZJ8 | Metabolic pathways |
| 49 | O-phosphoserine phosphohydrolase (PSPH) | Amino acid metabolism | Down | 0.47 | F2D8E1 | Biosynthesis of amino acids |
| 50 | Lipase | Lipid hydrolysis | Down | 0.47 | UCW116 | Metabolic pathways |
| 51 | Histone H2A | Chromatin organisation | Down | 0.49 | F2DGG6 | Necroptosis |
| 52 | Glutathione S-transferase (GST) | Redox homeostasis | Down | 0.49 | F2D4L0 | Glutathione metabolism |
| 53 | Chitinase (CHI) | Plant defense responses | Down | 0.50 | F2CQW5 | Amino sugar and nucleotide sugar metabolism |
| 54 | Acetyl-coenzyme A synthetase (AcsA) | Fatty acid production | Down | 0.51 | F2DVB4 | Glycolysis/gluconeogenesis |
| 55 | Germin-like protein 2a (GLP) | Plant defense responses | Down | 0.51 | Q0GR10 | Glyoxylate and dicarboxylate metabolism |
| 56 | Thioredoxin-dependent peroxiredoxin (TPx-Q) | Redox homeostasis | Down | 0.53 | F2DTT4 | Peroxisome |
| 57 | DEAD-box ATP-dependent RNA helicase 52C (DDX52) | Vesicle trafficking | Down | 0.54 | A0A287NPY4 | RNA degradation |
| 58 | Leucine-rich repeat extensin-like protein 6 | Cell wall organisation | Down | 0.56 | A0A287TFV1 | Metabolic pathways |
| 59 | Heat shock protein (HSP 70) | Protein homeostasis | Down | 0.56 | F2DT51 | MAPK signaling pathway |
| 60 | Thiamine pyrophosphate carrier 1 | Solute transport | Down | 0.56 | F2DJG4 | Transporters |
| 61 | Chitin elicitor-binding protein (CEBiP) | Stress response | Down | 0.57 | F2D6S9 | Plant-pathogen interaction |
| 62 | Elongation factor 3 (EF3) | Protein biosynthesis | Down | 0.58 | F2DTZ6 | Protein processing |
| 63 | Sucrose 1-fructosyltransferase (1-SST) | Fructan biosynthesis | Down | 0.61 | J7GM45 | Starch and sucrose metabolism |
| 64 | 12-oxophytodienoic acid reductase (OPR) | Oxylipins biosynthesis | Down | 0.62 | M1EUV0 | Biosynthesis of secondary metabolites |
| 65 | 3-oxo-Delta(4,5)-steroid 5-beta-reductase (VEP1) | Enzyme oxidoreductase | Down | 0.62 | M0Y0N6 | Metabolic pathways |
| 66 | ATP synthase subunit alpha (ATP5F1A) | Cellular respiration | Down | 0.62 | F2DBC9 | Metabolic pathways |
| 67 | Multiprotein-bridging factor 1a-like | Transcription regulation | Down | 0.63 | F2CW93 | Signal transduction |
| 68 | Beta-fructofuranosidase, insoluble isoenzyme 4-like isoform X1 (CWINV1) | Hydrolysis | Down | 0.63 | A0A287T167 | Biosynthesis of secondary metabolites |
| 69 | Laccase (LAC) | Enzyme oxidoreductase | Down | 0.64 | F2DUK8 | Metabolic pathways |
| 70 | Carboxypeptidase (CP) | Hydrolysis | Down | 0.65 | F2DJ52 | Protein digestion and absorption |
| 71 | Small heat shock protein (sHSP) | Protein homeostasis | Down | 0.65 | A0A287RG57 | Protein processing |
| 72 | Plant SNARE 13 (PSN13) | Vesicle trafficking | Down | 0.66 | M0X338 | SNARE interactions in vesicular transport |
| Parameters | Value |
|---|---|
| Total organic carbon (%) | 18.58 |
| Total N (%) | 1.21 |
| C/N | 15.36 |
| P (%) | 0.54 |
| K (%) | 0.95 |
| Ca (%) | 8.18 |
| Mg (%) | 1.05 |
| Na (%) | 0.42 |
| Alkalinity (% CaCO3) | 11.50 |
| Zn (mg kg−1 DW compost) | 70.10 |
| Fe (g kg−1 DW compost) | 70 |
| Mn (mg kg−1 DW compost) | 130 |
| Cu (mg kg−1 DW compost) | 11.60 |
| Cd (mg kg−1 DW compost) | 0.20 |
| Pb (mg kg−1 DW compost) | 4.15 |
| Cr (mg kg−1 DW compost) | 11.50 |
| Ni (mg kg−1 DW compost) | 5.88 |
| Total coliforms (MPN g DW−1 compost) | 143.33 ± 5.77 |
| Faecal coliforms (MPN g DW−1 compost) | 120 ± 17.32 |
| Escherichia coli (MPN g DW−1 compost) | 114 ± 23.79 |
| Faecal Streptococci (MPN g DW−1 compost) | 114.33 ± 23.8 |
| Salmonella spp. (MPN g DW−1 compost) | <0.3 |
| Shigella spp. (MPN g DW−1 compost) | <0.3 |
| Gene | Sequence (5′–3′) | Product Size (bp) | Tm (°C) |
|---|---|---|---|
| Plasma membrane ATPase (HvATPase) | F: 5′-CGTTGGTGTCTCCATTGTTG-3′ R: 5′-TTGCAACCGGTGGTGTAGTA-3′ | 105 | 60 |
| Pathogenesis related protein PR17 (HvPR17) | F: 5′-CAGAGCTGATGGTCGACGTA-3′ R: 5′-CGCTCCAGTCAATACAGCAA-3′ | 122 | 60 |
| Glutathione S-transferase (HvGST) | F: 5′-AAGCTGTACGGGATGATGCT-3′ R: 5′-GGTTGAGCTTGAGGAAGTCG-3′ | 139 | 60 |
| Chitinase (HvCHI) | F: 5′-CTACACGTACGACGCCTTCA-3′ R: 5′-TAGTCTCGTGGGAGGTCTGG-3′ | 122 | 61 |
| Thioredoxin-dependent peroxiredoxin (HvTPx-Q) | F: 5′-GTCCAAGAAAACCCAGACGA-3′ R: 5′-AAACGGCCATGACAAAACTC-3′ | 149 | 60 |
| Heat shock protein (HvHSP) | F: 5′-CTCAAATCGAGATCCCCGTA-3′ R: 5′-TCGATCTTGGCTTGTCCTCT-3′ | 118 | 60 |
| Tonoplast intrinsic protein (HvTIP) | F: 5′- GCTTCCTCCTCCGCTTCT-3′ R: 5′- ATGACGATCTCCAGGACCAC-3′ | 97 | 59 |
| Dehydrin (HvDHN) | F: 5′-CGTGTCAAGATGGAGGGATT-3′ R: 5′-CTGAAGCCCGTATACCCAAA-3′ | 98 | 60 |
| Aquaporin (HvPIP) | F: 5′-CTGGCCACTATCCCAATCAC-3′ R: 5′-CACCCAGAAGATCCAGTGGT-3′ | 111 | 60 |
| Late embryogenesis abundant protein 19-like (HvLEA19-like) | F: 5′-ACGAAGGAGAGGGACAGGAT-3′ R: 5′-CGCGTACAGATTTCCAGACA-3′ | 106 | 59 |
| Late embryogenesis abundant protein LEA3 (HvLEA3) | F: 5′-CATGGGAGGGGACAACAC-3′ R: 5′-GATTCCTGGTGGTGGTGGT-3′ | 86 | 61 |
| Dehydrin DHN5 (HvDHN5) | F: 5′-TTACATGCCGACACTTCCAA-3′ R: 5′-CGAAAACATCCGATCCTTGT-3′ | 85 | 59 |
| Actin (HvActin) | F: 5′-CGACAATGGAACCGGAATG-3′ R: 5′-CCCTTGGCGCATCATCTC-3′ | 56 | 61 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghouili, E.; Sassi, K.; Hidri, Y.; M’Hamed, H.C.; Somenahally, A.; Xue, Q.; Jebara, M.; Nefissi Ouertani, R.; Riahi, J.; de Oliveira, A.C.; et al. Effects of Date Palm Waste Compost Application on Root Proteome Changes of Barley (Hordeum vulgare L.). Plants 2023, 12, 526. https://doi.org/10.3390/plants12030526
Ghouili E, Sassi K, Hidri Y, M’Hamed HC, Somenahally A, Xue Q, Jebara M, Nefissi Ouertani R, Riahi J, de Oliveira AC, et al. Effects of Date Palm Waste Compost Application on Root Proteome Changes of Barley (Hordeum vulgare L.). Plants. 2023; 12(3):526. https://doi.org/10.3390/plants12030526
Chicago/Turabian StyleGhouili, Emna, Khaled Sassi, Yassine Hidri, Hatem Cheikh M’Hamed, Anil Somenahally, Qingwu Xue, Moez Jebara, Rim Nefissi Ouertani, Jouhaina Riahi, Ana Caroline de Oliveira, and et al. 2023. "Effects of Date Palm Waste Compost Application on Root Proteome Changes of Barley (Hordeum vulgare L.)" Plants 12, no. 3: 526. https://doi.org/10.3390/plants12030526
APA StyleGhouili, E., Sassi, K., Hidri, Y., M’Hamed, H. C., Somenahally, A., Xue, Q., Jebara, M., Nefissi Ouertani, R., Riahi, J., de Oliveira, A. C., Abid, G., & Muhovski, Y. (2023). Effects of Date Palm Waste Compost Application on Root Proteome Changes of Barley (Hordeum vulgare L.). Plants, 12(3), 526. https://doi.org/10.3390/plants12030526