

Effects of Isoflavone-Rich NADES Extract of Pueraria lobata Roots and Astaxanthin-Rich Phaffia rhodozyma Extract on Prostate Carcinogenesis in Rats
 ,
,
Abstract
:1. Introduction
2. Results
2.1. FTICR-MS Molecular Characterization of Isoflavone-Rich NADES Extract of Pueraria Lobata Roots and Astaxanthin-Rich Phaffia Rhodozyma Extract
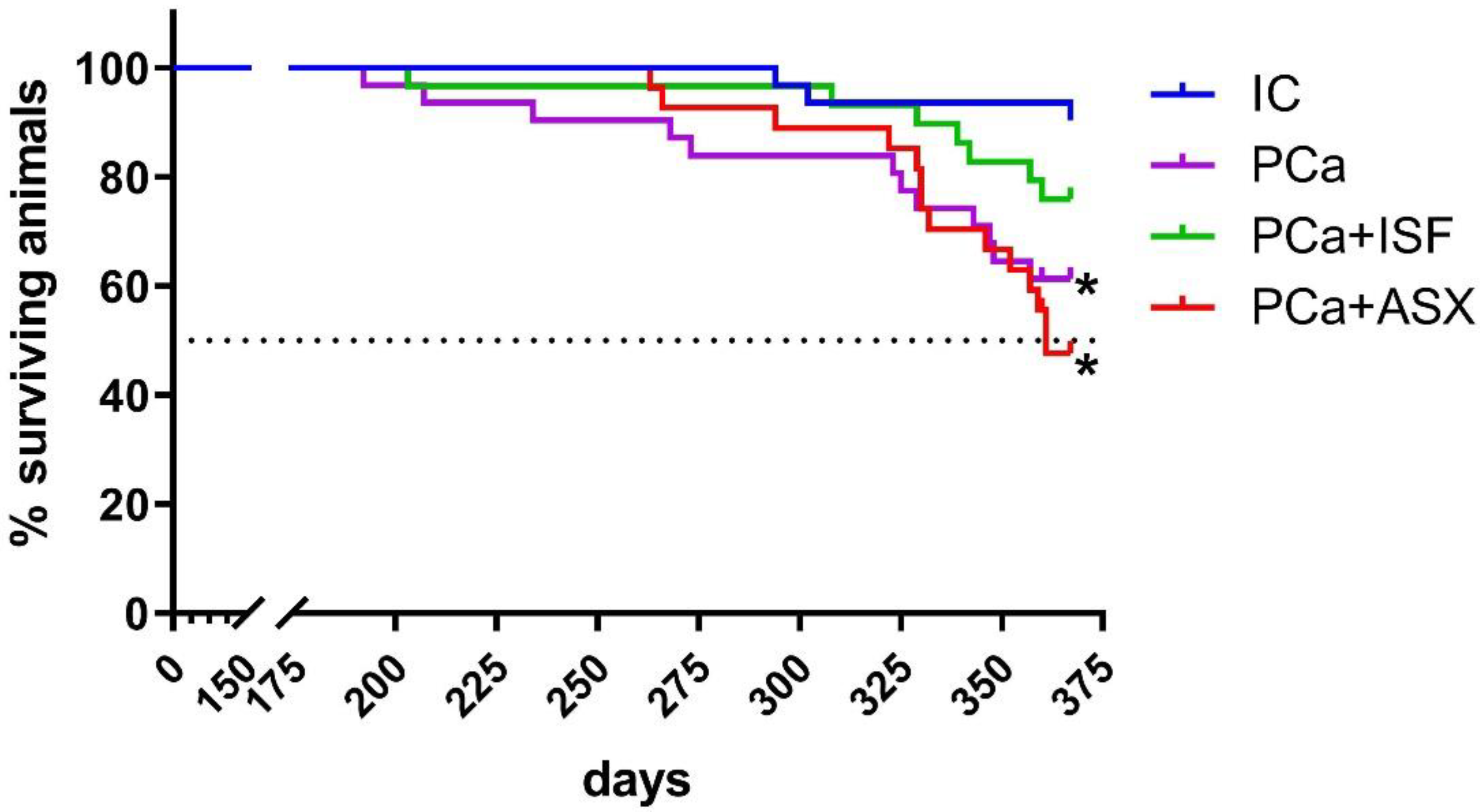
2.2. Survival Data and Long-Term Toxicity of Isoflavone-Rich NADES Extract of Pueraria Lobata Roots and Astaxanthin-Rich Phaffia Rhodozyma Extract
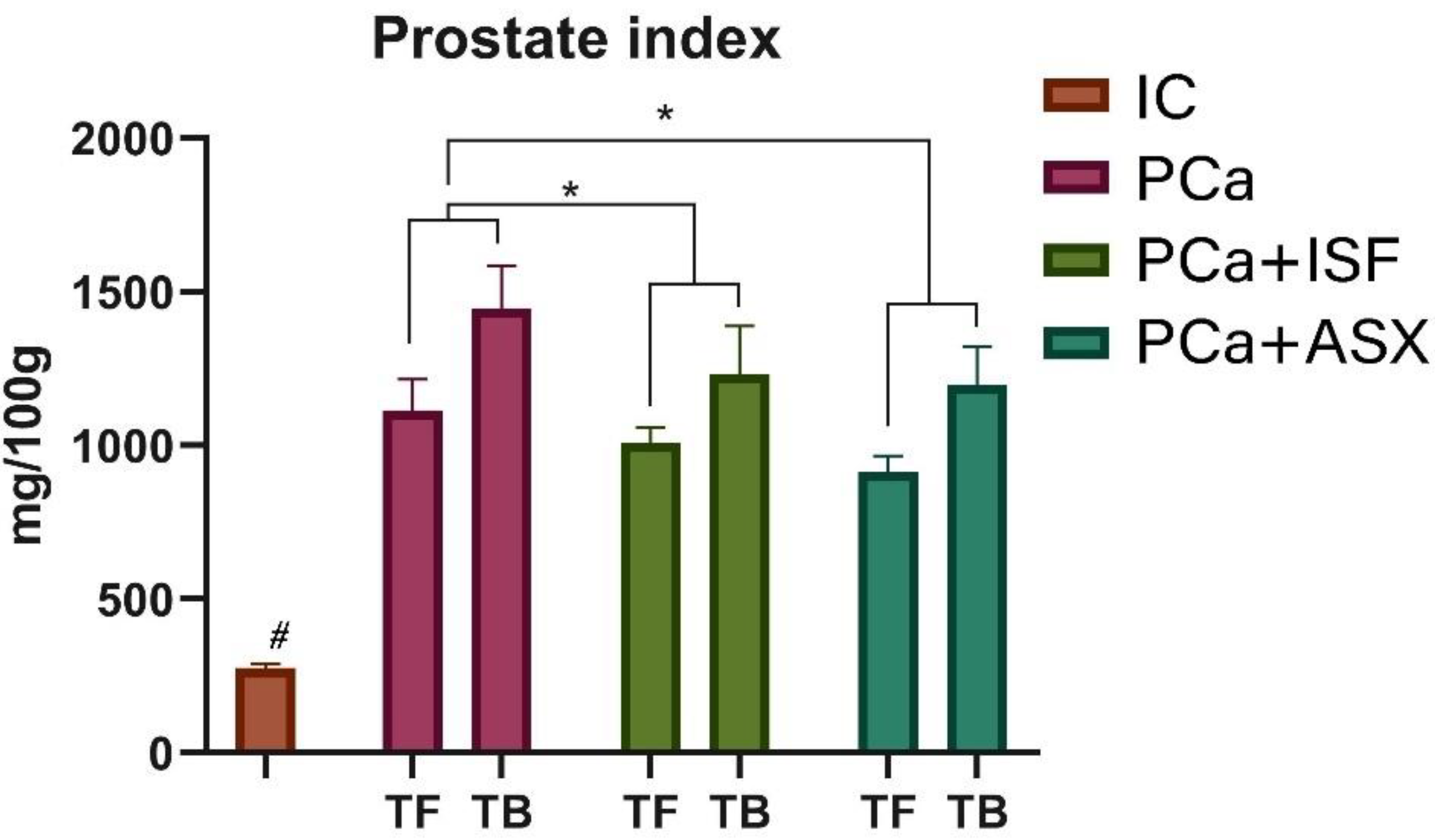
2.3. Prostate Cancer Incidence, Prostate Index and Urinary Retention
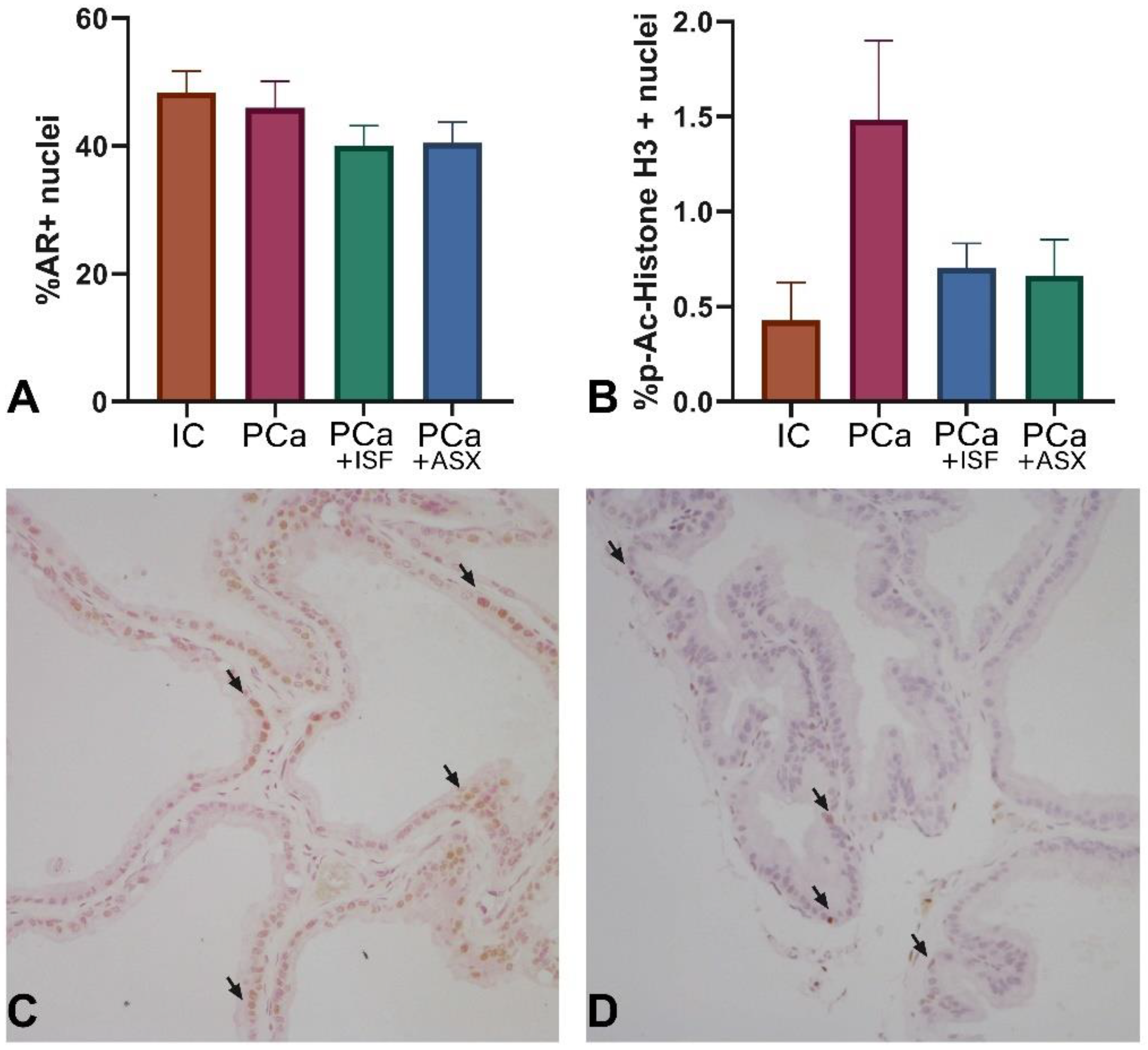
2.4. Assessment of Androgen Receptor Expression and Proliferation
2.5. Prostate Tissue Biochemical Analysis
2.6. Blood Biochemical Analysis
3. Discussion
4. Materials and Methods
4.1. Study Materials
4.2. High-Resolution Mass Spectrometry Characterization of Study Materials
4.3. Animals
4.4. Experimental Design
4.5. Pathomorphological Examination
4.6. Biochemical Analysis, Antioxidant Activity Evaluation, Blood Count and ELISA Assay
4.7. Ethical Standards
4.8. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Culp, M.B.; Soerjomataram, I.; Efstathiou, J.A.; Bray, F.; Jemal, A. Recent Global Patterns in Prostate Cancer Incidence and Mortality Rates. Eur. Urol. 2020, 77, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Rawla, P. Epidemiology of Prostate Cancer. World J. Oncol. 2019, 10, 63–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Abajobir, A.A.; Abate, K.H.; Abbafati, C.; Abbas, K.M.; Abd-Allah, F.; Abera, S.F.; Abraha, H.N.; Abu-Raddad, L.J.; Abu-Rmeileh, N.M.E.; et al. Global, Regional, and National under-5 Mortality, Adult Mortality, Age-Specific Mortality, and Life Expectancy, 1970–2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1084–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.; Hlatky, L.; Jeong, Y.-S.; Kim, D. Therapeutic Effectiveness of Anticancer Phytochemicals on Cancer Stem Cells. Toxins 2016, 8, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenov, A.; Gubareva, E.; Ermakova, E.; Dorofeeva, A.; Tumanyan, I.; Radetskaya, E.; Yurova, M.; Aboushanab, S.; Kanwugu, O.; Fedoros, E.; et al. Astaxantin and Isoflavones Inhibit Benign Prostatic Hyperplasia in Rats by Reducing Oxidative Stress and Normalizing Ca/Mg Balance. Plants 2021, 10, 2735. [Google Scholar] [CrossRef]
- Sarkar, F.H.; Li, Y. Soy Isoflavones and Cancer Prevention. Cancer Investig. 2003, 21, 744–757. [Google Scholar] [CrossRef]
- Sawada, N.; Iwasaki, M.; Yamaji, T.; Shimazu, T.; Inoue, M.; Tsugane, S. Soy and Isoflavone Consumption and Subsequent Risk of Prostate Cancer Mortality: The Japan Public Health Center-Based Prospective Study. Int. J. Epidemiol. 2020, 49, 1553–1561. [Google Scholar] [CrossRef]
- Applegate, C.; Rowles, J.; Ranard, K.; Jeon, S.; Erdman, J. Soy Consumption and the Risk of Prostate Cancer: An Updated Systematic Review and Meta-Analysis. Nutrients 2018, 10, 40. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.L. A Preliminary Investigation of the Enzymatic Inhibition of 5alpha-Reduction and Growth of Prostatic Carcinoma Cell Line LNCap-FGC by Natural Astaxanthin and Saw Palmetto Lipid Extract in Vitro. J. Herb. Pharmacother. 2005, 5, 17–26. [Google Scholar] [CrossRef]
- Faraone, I.; Sinisgalli, C.; Ostuni, A.; Armentano, M.F.; Carmosino, M.; Milella, L.; Russo, D.; Labanca, F.; Khan, H. Astaxanthin Anticancer Effects Are Mediated through Multiple Molecular Mechanisms: A Systematic Review. Pharmacol. Res. 2020, 155, 104689. [Google Scholar] [CrossRef]
- Zherebker, A.Y.; Rukhovich, G.D.; Kharybin, O.N.; Fedoros, E.I.; Perminova, I.V.; Nikolaev, E.N. Fourier Transform Ion Cyclotron Resonance Mass Spectrometry for the Analysis of Molecular Composition and Batch-to-batch Consistency of Plant-derived Polyphenolic Ligands Developed for Biomedical Application. Rapid Commun. Mass Spectrom. 2020, 34, e8850. [Google Scholar] [CrossRef] [PubMed]
- Zherebker, A.; Rukhovich, G.D.; Sarycheva, A.; Lechtenfeld, O.J.; Nikolaev, E.N. Aromaticity Index with Improved Estimation of Carboxyl Group Contribution for Biogeochemical Studies. Environ. Sci. Technol. 2022, 56, 2729–2737. [Google Scholar] [CrossRef] [PubMed]
- Maia, M.; Figueiredo, A.; Cordeiro, C.; Sousa Silva, M. FT-ICR-MS-based Metabolomics: A Deep Dive into Plant Metabolism. Mass Spectrom. Rev. 2021, Volume, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Chaleckis, R.; Meister, I.; Zhang, P.; Wheelock, C.E. Challenges, Progress and Promises of Metabolite Annotation for LC–MS-Based Metabolomics. Curr. Opin. Biotechnol. 2019, 55, 44–50. [Google Scholar] [CrossRef]
- McCormick, D.L.; Rao, K.V.; Dooley, L.; Steele, V.E.; Lubet, R.A.; Kelloff, G.J.; Bosland, M.C. Influence of N-Methyl-N-Nitrosourea, Testosterone, and N-(4-Hydroxyphenyl)-All-Trans-Retinamide on Prostate Cancer Induction in Wistar-Unilever Rats. Cancer Res. 1998, 58, 3282–3288. [Google Scholar]
- Rojas-Armas, J.P.; Arroyo-Acevedo, J.L.; Ortiz-Sánchez, J.M.; Palomino-Pacheco, M.; Herrera-Calderón, O.; Calva, J.; Rojas-Armas, A.; Justil-Guerrero, H.J.; Castro-Luna, A.; Hilario-Vargas, J.; et al. Flowers: A Promising Medicinal Plant as Chemopreventive in Induced Prostate Carcinogenesis in Rats. Evid. Based Complement. Altern. Med. 2020, 2020, 1–8. [Google Scholar] [CrossRef]
- Sivoňová, M.; Kaplán, P.; Tatarková, Z.; Lichardusová, L.; Dušenka, R.; Jurečeková, J. Androgen Receptor and Soy Isoflavones in Prostate Cancer (Review). Mol. Clin. Oncol. 2018, 10, 191–204. [Google Scholar] [CrossRef] [Green Version]
- Nogueira-De-Almeida, C.A.; Ferraz, I.S.; Ued, F.D.V.; Almeida, A.C.F.; Del Ciampo, L.A. Impact of Soy Consumption on Human Health: Integrative Review. Braz. J. Food Technol. 2020, 23, e2019129. [Google Scholar] [CrossRef]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, A.M.; Yang, W.; Bosland, M.C. Soy Isoflavones and Prostate Cancer: A Review of Molecular Mechanisms. J. Steroid Biochem. Mol. Biol. 2014, 140, 116–132. [Google Scholar] [CrossRef] [Green Version]
- Ong, S.; Shanmugam, M.; Fan, L.; Fraser, S.; Arfuso, F.; Ahn, K.; Sethi, G.; Bishayee, A. Focus on Formononetin: Anticancer Potential and Molecular Targets. Cancers 2019, 11, 611. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.F.; Ahmad, A.; Zubair, H.; Khan, H.Y.; Wang, Z.; Sarkar, F.H.; Hadi, S.M. Soy Isoflavone Genistein Induces Cell Death in Breast Cancer Cells through Mobilization of Endogenous Copper Ions and Generation of Reactive Oxygen Species. Mol. Nutr. Food Res. 2011, 55, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Eren-Guzelgun, B.; Ince, E.; Gurer-Orhan, H. In Vitro Antioxidant/Prooxidant Effects of Combined Use of Flavonoids. Nat. Prod. Res. 2018, 32, 1446–1450. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Sofic, E.; Prior, R.L. Antioxidant and Prooxidant Behavior of Flavonoids: Structure-Activity Relationships. Free. Radic. Biol. Med. 1997, 22, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.H.; Wang, Y.; Xin, M. Low Concentration of Formononetin Stimulates the Proliferation of Nasopharyngeal Carcinoma Cell Line CNE2 by Upregulating Bcl-2 and p-ERK1/2 Expression. Pharm. Biol. 2016, 54, 1841–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Xiong, C.; Xu, P.; Luo, Q.; Zhang, R. Puerarin Induces Apoptosis in Prostate Cancer Cells via Inactivation of the Keap1/Nrf2/ARE Signaling Pathway. Bioengineered 2021, 12, 402–413. [Google Scholar] [CrossRef]
- Fakhri, S.; Abbaszadeh, F.; Dargahi, L.; Jorjani, M. Astaxanthin: A Mechanistic Review on Its Biological Activities and Health Benefits. Pharmacol. Res. 2018, 136, 1–20. [Google Scholar] [CrossRef]
- Sun, S.-Q.; Zhao, Y.-X.; Li, S.-Y.; Qiang, J.-W.; Ji, Y.-Z. Anti-Tumor Effects of Astaxanthin by Inhibition of the Expression of STAT3 in Prostate Cancer. Mar. Drugs 2020, 18, 415. [Google Scholar] [CrossRef]
- Montazeri-Najafabady, N.; Chatrabnous, N.; Arabnezhad, M.; Azarpira, N. Anti-androgenic Effect of Astaxanthin in LNCaP Cells Is Mediated through the Aryl Hydrocarbon-androgen Receptors Cross Talk. J. Food Biochem. 2021, 45, e13702. [Google Scholar] [CrossRef]
- Jin, B.-R.; Kim, H.-J.; Park, S.-K.; Kim, M.-S.; Lee, K.-H.; Yoon, I.-J.; An, H.-J. Anti-Proliferative Effects of HBX-5 on Progression of Benign Prostatic Hyperplasia. Molecules 2018, 23, 2638. [Google Scholar] [CrossRef] [Green Version]
- Dose, J.; Matsugo, S.; Yokokawa, H.; Koshida, Y.; Okazaki, S.; Seidel, U.; Eggersdorfer, M.; Rimbach, G.; Esatbeyoglu, T. Free Radical Scavenging and Cellular Antioxidant Properties of Astaxanthin. Int. J. Mol. Sci. 2016, 17, 103. [Google Scholar] [CrossRef] [PubMed]
- Body, J.-J.; Niepel, D.; Tonini, G. Hypercalcaemia and Hypocalcaemia: Finding the Balance. Support Care Cancer 2017, 25, 1639–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zherebker, A.; Shirshin, E.; Rubekina, A.; Kharybin, O.; Kononikhin, A.; Kulikova, N.A.; Zaitsev, K.V.; Roznyatovsky, V.A.; Grishin, Y.K.; Perminova, I.V. Optical Properties of Soil Dissolved Organic Matter Are Related to Acidic Functions of Its Components as Revealed by Fractionation, Selective Deuteromethylation, and Ultrahigh Resolution Mass Spectrometry. Environ. Sci. Technol. 2020, 54, 2667–2677. [Google Scholar] [CrossRef] [PubMed]
- Leefmann, T.; Frickenhaus, S.; Koch, B.P. UltraMassExplorer: A Browser-based Application for the Evaluation of High-resolution Mass Spectrometric Data. Rapid Commun. Mass Spectrom. 2019, 33, 193–202. [Google Scholar] [CrossRef]
- Kellerman, A.M.; Dittmar, T.; Kothawala, D.N.; Tranvik, L.J. Chemodiversity of Dissolved Organic Matter in Lakes Driven by Climate and Hydrology. Nat. Commun. 2014, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Bespalov, V.G.; Panchenko, A.V.; Murazov, I.G.; Chepik, O.F. Influence of sodium selenite on carcinogenesis of the prostate and other organs induced by methylnitrosourea and testosterone in rats. Vopr. Onkol. 2011, 57, 486–492. (In Russian) [Google Scholar]
- Herrera, G.M.; Meredith, A.L. Diurnal Variation in Urodynamics of Rat. PLoS ONE 2010, 5, e12298. [Google Scholar] [CrossRef] [Green Version]
- Panchenko, A.V.; Tyndyk, M.L.; Fedoros, E.I.; Maydin, M.A.; Semenov, A.L.; Gubareva, E.A.; Golubev, A.G.; Anisimov, V.N. Comparative Analysis of Experimental Data on the Effects of Different Polyphenols on Lifespan and Aging. Adv. Gerontol. 2019, 9, 396–401. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Group | Mean Animal Weight (g) | Relative Organ Weight, mg/100 g | ||||
|---|---|---|---|---|---|---|
| At the Start of PCa Induction | At the End of Experiment | Liver | Kidneys | Heart | Spleen | |
| IC | 328.9 ± 5.2 | 537.9 ± 15.2 | 3207 ± 47 | 533 ± 12 | 281 ± 8 | 218 ± 7 |
| PCa | 321.9 ± 5.3 | 401.4 ± 9.2 a | 3603 ± 49 a | 830 ± 13 a | 344 ± 9 a | 302 ± 16 a |
| PCa + ISF | 320.2 ± 5.1 | 394.9 ± 8.7 a | 3622 ± 50 a | 810 ± 17 a | 336 ± 9 a | 326 ± 17 a |
| PCa + ASX | 318.6 ± 4.1 | 402.3 ± 9.2 a | 3652 ± 96 a | 832 ± 20 a | 343 ± 9 a | 288 ± 14 a |
| Group | Effective Number of Animals | Number of Animals with Cancer (%) | |||
|---|---|---|---|---|---|
| Whole Prostate | Dorsolateral Lobe | Ventral Lobes | Anterior Lobes | ||
| IC | 31 | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| PCa | 30 | 11 (37%) a | 9 (30%) a | 5 (17%) a | 0 (0%) |
| PCa + ISF | 27 | 10 (37%) a | 9 (33%) a | 4 (15%) a | 0 (0%) |
| PCa + ASX | 29 | 12 (41%) a | 9 (31%) a | 8 (28%) a | 0 (0%) |
| Group | SOD, Relative Activity | Ca, µmol/g Protein | Mg, µmol/g Protein | Ca/Mg |
|---|---|---|---|---|
| IC | 100.0 ± 8.1 | 4.64 ± 0.74 | 18.4 ± 4.4 | 0.30 ± 0.07 |
| PCa | 71.3 ± 9.1 a | 12.92 ± 0.78 a | 18.0 ± 2.1 | 0.76 ± 0.09 a |
| PCa + ISF | 76.3 ± 6.4 a | 12.85 ± 0.95 a | 16.2 ± 2.8 | 0.82 ± 0.08 a |
| PCa + ASX | 65.0 ± 6.1 a | 8.32 ± 1.62 a, b | 17.3 ± 4.7 | 0.64 ± 0.11 a |
| Group | Cholesterol, mmol/L | Calcium, mmol/L | Magnesium, mmol/L | Ca/Mg | Phosphorus, mmol/L | SOD, Relative Activity | MDA, µmol/L12 RBC | PSA, ng/mL | Testosterone, ng/mL | DHT, pg/mL |
|---|---|---|---|---|---|---|---|---|---|---|
| IC | 1.07 ± 0.06 | 2.28 ± 0.04 | 0.73 ± 0.02 | 3.14 ± 0.08 | 1.61 ± 0.04 | 100.0 ± 1.4 | 22.0 ± 1.5 | 1.87±0.15 | 0.55±0.21 | 43.6±18.7 |
| PCa | 0.93 ± 0.05 a | 2.00 ± 0.03 a | 0.78 ± 0.01 | 2.61 ± 0.05 a | 1.38 ± 0.05 a | 94.6 ± 1.4 a | 29.9 ± 2.1 | 1.65±0.16 | 11.03±1.08 a | 421.5±40.9 a |
| PCa + ISF | 0.76 ±0.04 a,b | 2.16 ±0.04 a,b | 0.78 ± 0.03 | 2.84 ±0.08 a,b | 1.62 ± 0.06 b | 89.0 ±0.9 a,b | 38.1 ± 3.6 a | 1.28 ±0.11 a,b | 16.37±1.05 a,b | 547.3±53.9 a |
| PCa + ASX | 0.91 ± 0.03 a | 2.11 ±0.06 a,b | 0.75 ± 0.03 | 2.85 ±0.10 a,b | 1.49 ± 0.09 a | 94.7 ± 2.0 a | 31.3 ± 1.5 | 0.79 ±0.13 a,b | 24.96±3.95 a,b | 588.6±103.0 a,b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semenov, A.L.; Tyndyk, M.L.; Von, J.D.; Ermakova, E.D.; Dorofeeva, A.A.; Tumanyan, I.A.; Radetskaya, E.A.; Yurova, M.N.; Zherebker, A.; Gorbunov, A.Y.; et al. Effects of Isoflavone-Rich NADES Extract of Pueraria lobata Roots and Astaxanthin-Rich Phaffia rhodozyma Extract on Prostate Carcinogenesis in Rats. Plants 2023, 12, 564. https://doi.org/10.3390/plants12030564
Semenov AL, Tyndyk ML, Von JD, Ermakova ED, Dorofeeva AA, Tumanyan IA, Radetskaya EA, Yurova MN, Zherebker A, Gorbunov AY, et al. Effects of Isoflavone-Rich NADES Extract of Pueraria lobata Roots and Astaxanthin-Rich Phaffia rhodozyma Extract on Prostate Carcinogenesis in Rats. Plants. 2023; 12(3):564. https://doi.org/10.3390/plants12030564
Chicago/Turabian StyleSemenov, Alexander L., Margarita L. Tyndyk, Julia D. Von, Elena D. Ermakova, Anastasia A. Dorofeeva, Irina A. Tumanyan, Ekaterina A. Radetskaya, Maria N. Yurova, Alexander Zherebker, Alexander Yu. Gorbunov, and et al. 2023. "Effects of Isoflavone-Rich NADES Extract of Pueraria lobata Roots and Astaxanthin-Rich Phaffia rhodozyma Extract on Prostate Carcinogenesis in Rats" Plants 12, no. 3: 564. https://doi.org/10.3390/plants12030564
APA StyleSemenov, A. L., Tyndyk, M. L., Von, J. D., Ermakova, E. D., Dorofeeva, A. A., Tumanyan, I. A., Radetskaya, E. A., Yurova, M. N., Zherebker, A., Gorbunov, A. Y., Fedoros, E. I., Panchenko, A. V., & Anisimov, V. N. (2023). Effects of Isoflavone-Rich NADES Extract of Pueraria lobata Roots and Astaxanthin-Rich Phaffia rhodozyma Extract on Prostate Carcinogenesis in Rats. Plants, 12(3), 564. https://doi.org/10.3390/plants12030564