
Abutilon theophrasti’s Resilience against Allelochemical-Based Weed Management in Sustainable Agriculture – Due to Collection of Highly Advantageous Microorganisms?




Abstract
:
1. Introduction
2. Biological Control of Abutilon theophrasti by Microorganisms
Natural Compounds—Monoterpenoids, Benzyl-Isothiocyanate, Phenolics, and Sorgoleone in the Management of Abutilon theophrasti
3. Resilience of Abutilon theophrasti against Benzoxazinoid Allelochemicals from Rye (Secale cereale L.) Mulches
3.1. Microorganisms Associated with Abutilon theophrasti and Microbial Role in Benzoxazinoid Tolerance
3.2. Defense Strategies against Benzoxazolinone (BOA)—Uptake, Detoxification, Membrane Repair, and Polymerizations at the Root Surface
4. Secondary Metabolites of Velvetleaf
5. Considerations and Further Perspective
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cardina, J. Lives of Weeds: Opportunism, Resistance, Folly; Cornell University Press: Ithaca, NY, USA, 2021; Chapter 3; pp. 73–100. ISBN 9781501758980. [Google Scholar]
- Warwick, S.I.; Black, L.D. The biology of Canadian Weeds. 90. Abutilon theophrasti. Can. J. Plant Sci. 1988, 68, 1069–1085. [Google Scholar] [CrossRef]
- Stegink, S.J.; Spencer, N.R. Using Protein Electrophoresis to Investigate the Phylogeny of Velvetleaf (Abutilon theophrasti). Weed Sci. 1988, 36, 172–175. Available online: https://www.jstor.org/stable/4044866 (accessed on 31 January 2023). [CrossRef]
- Rojas-Sandoval, J. Abutilon theophrasti (Velvet Leaf). Available online: https://cabidigitallibrary.org/terms-and-conditions (accessed on 3 March 2022).
- Cortés, J.A.; Mendiola, M.A.; Castejón, M. Competition of velvetleaf (Abutilon theophrasti) weed with cotton (Gossypium hirsutum L.). Economic damage threshold. Span. J. Agric. Res. 2010, 8, 391–399. [Google Scholar] [CrossRef]
- Rain, A.S.; Cârciu, G. Abutilon theophrasti L. (velvetleaf): A problem-weed for grain maize. J. Hortic. Sci. Biotechnol. 2018, 22, 124–127. Available online: https://journal-hfb.usab-tm.ro/2018/Volum%2022(3)%20PDF/21Rain%20A.%202.pdf (accessed on 31 January 2023).
- Follak, S.; Aldrian, U.; Schwarz, M. Spread dynamics of Abutilon theophrasti in Central Europe. Plant Protect. Sci. 2014, 50, 157–163. [Google Scholar] [CrossRef]
- Tanji, A.; Taleb, A. New weed species recently introduced into Morocco. Weed Res. 1997, 37, 27–31. [Google Scholar] [CrossRef]
- Ghanizadeh, H.; James, T.K. Behaviour of Abutilon theophrasti in Different Climatic Niches: A New Zealand Case Study. Front. Plant Sci. 2022, 13, 885779. [Google Scholar] [CrossRef]
- Mahklouf, M.H.; El-Ahamir, S.M. The First Record of Abutilon theophrasti (Malvaceae) in the Flora of Libya. Phytotaxa 2019, 402, 259–264. [Google Scholar] [CrossRef]
- Scheliga, M.; Brand, U.; Türk, O.; Gruber, S.; Petersen, J. Field Trials Indicating the Potential of Abutilon theophrasti (Medic.) as a New Fibre Crop in Southwest Germany. J. Adv. Agric. 2019, 10, 1726–1739. [Google Scholar] [CrossRef]
- Bensky, D.; Gamble, A.; Kaptchuk, T.J. Chinese Herbal Medicine: Materia Medica; Eastland Press: Seattle, WA, USA, 1993; pp. 142–143. ISBN 0939616157. [Google Scholar]
- Spencer, N.R. Velvetleaf, Abutilon theophrasti (Malvaceae). History and Economic Impact in the United States. Econ. Bot. 1984, 38, 407–416. Available online: https://www.jstor.org/stable/4254678 (accessed on 31 January 2023). [CrossRef]
- Medovic, A.; Horvath, F. Content of a Storage Jar from the Late Neolithic Site of Hódmezővásárhely-Gorzsa, South Hungary: A Thousand Carbonized Seeds of Abutilon theophrasti Medic. Veg. Hist. Archaeobot. 2012, 21, 215–220. [Google Scholar] [CrossRef]
- Vaughn, L.G.; Bernards, M.L.; Arkebauer, T.J.; Lindquist, J.L. Corn and Velvetleaf (Abutilon theophrasti) Growth and Transpiration Efficiency under Varying Water Supply. Weed Sci. 2016, 64, 596–604. Available online: https://www.jstor.org/stable/26420728 (accessed on 31 January 2023). [CrossRef]
- Lindquist, J.L.; Mortensen, D.A. Tolerance and Velvetleaf (Abutilon theophrasti) Suppressive Ability of Two Old and Two Modern Corn (Zea mays) Hybrids. Weed Sci. 1998, 46, 569–574. [Google Scholar] [CrossRef]
- Ma, X.; Yang, J.; Wu, H.; Jiang, W.; Ma, Y.; Ma, Y. Growth Analysis of Cotton in Competition with Velvetleaf (Abutilon theophrasti). Weed Technol. 2016, 30, 123–136. [Google Scholar] [CrossRef]
- Sterling, T.M.; Putnam, A.R. Possible role of glandular trichome exudates in interference by velvetleaf (Abutilon theophrasti). Weed Sci. 1987, 35, 308–314. Available online: https://www.jstor.onarg/stable/4044589 (accessed on 31 January 2023). [CrossRef]
- De Sanctis, J.; Jhala, A. Interaction of dicamba, fluthiacet-methyl, and glyphosate for control of velvetleaf (Abutilon theophrasti) in dicamba/glyphosate–resistant soybean. Weed Technol. 2021, 35, 761–767. [Google Scholar] [CrossRef]
- Striegel, A.; Eskridge, K.M.; Lawrence, N.C.; Knezevic, S.Z.; Kruger, G.R.; Proctor, C.A.; Hein, G.L.; Jhala, A.J. Economics of herbicide programs for weed control in conventional, glufosinate, and dicamba/glyphosate-resistant soybean across Nebraska. Agron. J. 2020, 112, 5158–5179. [Google Scholar] [CrossRef]
- Travlos, I.; Kanatas, P.; Economou, G.; Kotoulas, V.; Chachalis, D.; Tsioros, S. Evaluation of velvetleaf interference with maize hybrids as influenced by relative time of emergence. Exp. Agric. 2012, 48, 127–137. [Google Scholar] [CrossRef]
- Tataridas, A.; Kanatas, P.; Chatzigeorgiou, A.; Zannopoulos, S.; Travlos, I. Sustainable Crop and Weed Management in the Era of the EU Green Deal: A Survival Guide. Agronomy 2022, 12, 589. [Google Scholar] [CrossRef]
- Barnes, E.R.; Lawrence, N.C.; Knezevic, S.Z.; Rodriguez, O.; Irmak, S.; Jhala, A.J. Weed control and response of yellow and white popcorn hybrids to herbicides. Agron. J. 2020, 112, 458–469. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide Resistant Weeds. Available online: www.weedscience.org (accessed on 12 December 2022).
- Cardina, J.; Norquay, H.M. Seed Production and Seedbank Dynamics in Subthreshold Velvetleaf (Abutilon theophrasti) Populations. Weed Sci. 1997, 45, 85–90. Available online: https://www.jstor.org/stable/4045717 (accessed on 31 January 2023). [CrossRef]
- Vatovec, C.; Jordan, N.; Huerd, S. Responsiveness of certain agronomic weed species to arbuscular mycorrhizal fungi. Renew. Agric. Food Syst. 2005, 20, 181–189. [Google Scholar] [CrossRef]
- O’Neil, C.M.; Nash, J.; Tiemann, L.K.; Miesel, J.R. Mycorrhizal Symbioses Enhance Competitive Weed Growth in Biochar and Nutrient-Amended Soils. Front. Agron. 2021, 3, 731184. [Google Scholar] [CrossRef]
- Horvath, D.P.; Llewellyn, D.; Clay, S.A. Heterologous Hybridization of Cotton Microarrays with Velvetleaf (Abutilon theophrasti) Reveals Physiological Responses due to Corn Competition. Weed Sci. 2007, 55, 546–557. Available online: https://www.jstor.org/stable/4539616 (accessed on 31 January 2023). [CrossRef]
- Horvath, D.; Gulden, R.; Clay, S. Microarray analysis of late-season velvetleaf (Abutilon theophrasti) effect on corn. Weed Sci. 2006, 54, 983–994. [Google Scholar] [CrossRef]
- Kremer, R.J.; Hughes, L.B., Jr.; Aldrich, R.J. Examination of Microorganisms and Deterioration Resistance Mechanisms Associated with Velvetleaf Seed. Agron. J. 1984, 76, 745–749. [Google Scholar] [CrossRef]
- Kremer, R.J. Antimicrobial Activity of Velvetleaf (Abutilon theophrasti) Seeds. Weed Sci. 1986, 34, 617–622. [Google Scholar] [CrossRef]
- Kremer, R.J.; Kennedy, A.C. Rhizobacteria as biocontrol agents of weeds. Weed Technol. 1996, 10, 601–609. [Google Scholar] [CrossRef]
- Boyetchko, S. Impact of soil microorganisms on weed biology and ecology. Phytoprotection 1996, 77, 41–56. [Google Scholar] [CrossRef]
- Kremer, R.J.; Spencer, N.R. Impact of a Seed-Feeding Insect and Microorganisms of Velvetleaf (Abutilon theophrasti) Seed Viability. Weed Sci. 1989, 37, 211–216. Available online: https://www.jstor.org/stable/4044846 (accessed on 31 January 2023). [CrossRef]
- Boyette, C.; Walker, H. Evaluation of Fusarium lateritium as a Biological Herbicide for Controlling Velvetleaf (Abutilon theophrasti) and Prickly Sida (Sida spinosa). Weed Sci. 1986, 34, 106–109. [Google Scholar] [CrossRef]
- Chee-Sanford, J.C.; Williams, M.M.; Davis, A.S.; Sims, G.K. Do microorganisms influence seed-bank dynamics? Weed Sci. 2006, 54, 575–587. [Google Scholar] [CrossRef]
- Meir, S.; Larroche, C.; Al-Ahmad, H.; Gressel, J. Fungal transformation of Colletotrichum coccodes with bacterial oahA to suppress defenses of Abutilon theophrasti. Crop. Prot. 2009, 28, 749–755. [Google Scholar] [CrossRef]
- Gressel, J.; Michaeli, D.; Kampel, V.; Amsellem, Z.; Warshawsky, A. Ultralow calcium requirements of fungi facilitate use of calcium regulating agents to suppress host calcium-dependent defenses, synergizing infection by a mycoherbicide. J. Agric. Food Chem. 2002, 50, 6353–6360. [Google Scholar] [CrossRef]
- Ahn, B.; Paulitz, T.; Jabaji-Hare, S.; Watson, A. Enhancement of Colletotrichum coccodes virulence by inhibitors of plant defense mechanisms. Biocontrol Sci. Technol. 2005, 15, 299–308. [Google Scholar] [CrossRef]
- Dutton, M.V.; Evans, C.S. Oxalate production by fungi: Its role in pathogenicity and ecology in the soil environment. Can. J. Microbiol. 1996, 42, 881–895. [Google Scholar] [CrossRef]
- Briere, S.C.; Watson, A.K.; Hallett, S.G. Oxalic acid production and mycelia biomass yield of Sclerotinia minor for the formulation enhancement of a granular turf bioherbicide. Biocontrol Sci. Technol. 2000, 10, 281–289. [Google Scholar] [CrossRef]
- Watson, A.K.; Ahn, E.S. Better Control of Weeds with Enhanced Bioherbicides. Enhancing Biocontrol Agents and Handling Risks; Vurro, M., Gressel, J., Butt, T., Harman, G., Pilgeram, A., Leger, R.S., Nuss, D., Eds.; IOS Press: Amsterdam, The Netherlands, 2001; pp. 106–113. [Google Scholar]
- Syed, S.; Buddolla, V.; Lian, B. Oxalate Carbonate Pathway—Conversion and Fixation of Soil Carbon—A Potential Scenario for Sustainability. Front. Plant Sci. 2020, 11, 591297. [Google Scholar] [CrossRef]
- Jennings, J.C.; Apel-Birkhold, P.C.; Bailey, B.A.; Anderson, J.D. Induction of Ethylene Biosynthesis and Necrosis in Weed Leaves by a Fusarium oxysporum Protein. Weed Sci. 2000, 48, 7–14. Available online: http://www.jstor.org/stable/4046151 (accessed on 31 January 2023). [CrossRef]
- Amsellem, Z.; Cohen, B.C.; Gressel, J. Engineering hypervirulence in a mycoherbicidal fungus for efficient weed control. Nat. Biotechnol. 2002, 10, 1035–1039. [Google Scholar] [CrossRef]
- Šantrić, L.; Radivojević, L.; Gajić Umiljendić, J.; Sarić-Krsmanović, M. Allelopathic effects of actinobacterial isolates on seed germination and early seedling growth of velvetleaf (Abutilon theophrasti Medik.). Pestic. Phytomed. 2022, 37, 41–48. [Google Scholar] [CrossRef]
- Kremer, R.J.; Begonia, M.F.T.; Stanley, L.; Lanham, E.T. Characterization of rhizobacteria associated with weed seedlings. Appl. Environ. Microbiol. 1990, 56, 1649–1655. [Google Scholar] [CrossRef] [PubMed]
- Owen, A.; Zdor, R. Effect of cyanogenic rhizobacteria on the growth of velvetleaf (Abutilon theophrasti) and corn (Zea mays L.) in autoclaved soil and the influence of supplemental glycine. Soil Biol. Biochem. 2001, 33, 801–809. [Google Scholar] [CrossRef]
- Siegien, I.; Bogatek, R. Cyanide action in plants-from toxic to regulatory. Acta Physiol. Plant. 2006, 28, 483–497. [Google Scholar] [CrossRef]
- Silva-Avalos, J.; Richmond, M.G.; Nagappan, O.; Kunz, D.A. Degradation of the metal-cyano complex tetracyanonickelate(II) by cyanide-utilizing bacterial isolates. Appl. Environ. Microbiol. 1990, 56, 3664–3670. [Google Scholar] [CrossRef]
- Harding, D.P.; Raizada, M.N. Controlling weeds with fungi, bacteria and viruses: A review. Front. Plant Sci. 2015, 6, 659. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; DiTommaso, A.; Kao-Kniffin, J. Opportunities for Microbiome Suppression of Weeds Using Regenerative Agricultural Technologies. Front. Soil Sci. 2022, 2, 838595. [Google Scholar] [CrossRef]
- Chee-Sanford, J.C. Weed Seeds as Nutritional Resources for Soil Ascomycota and Characterization of Specific Associations Between Plant and Fungal Species. Biol. Fertil. Soils 2008, 44, 763–771. [Google Scholar] [CrossRef]
- Verdeguer, M.; Sánchez-Moreiras, A.M.; Araniti, F. Phytotoxic Effects and Mechanism of Action of Essential Oils and Terpenoids. Plants 2020, 9, 1571. [Google Scholar] [CrossRef]
- Preusche, M.; Vahl, M.; Riediger, J.; Ulbrich, A.; Schulz, M. Modulating Expression Levels of TCP Transcription Factors by Mentha x piperita Volatiles—An Allelopathic Tool to Influence Leaf Growth? Plants 2022, 11, 3078. [Google Scholar] [CrossRef]
- Vaughn, S.; Spencer, G. Monoterpenes as parent structures for herbicides. Weed Sci. 1993, 41, 114–119. [Google Scholar] [CrossRef] [Green Version]
- Wolf, R.B.; Spencer, G.F.; Kwolek, W.F. Inhibition of Velvetleaf (Abutilon theophrasti) Germination and Growth by Benzyl Isothiocyanate, a Natural Toxicant. Weed Sci. 1984, 32, 612–615. [Google Scholar] [CrossRef]
- Tollsten, L.; Bergstrom, G. Headspace volatiles of whole plants and macerated plant parts of Brassica and Sinapis. Phytochemistry 1988, 27, 4013–4018. [Google Scholar] [CrossRef]
- Shettle, N.; Balke, E.N. Plant Growth Response to Several Allelopathic Chemicals. Weed Sci. 1983, 31, 293–298. [Google Scholar] [CrossRef]
- Rose, S.J.; Burnside, O.C.; Specht, J.E.; Swisher, B.A. Competition and Allelopathy Between Soybeans and Weeds. Agron. J. 1984, 76, 523–528. [Google Scholar] [CrossRef]
- Dayan, F.E.; Howell, J.L.; Weidenhamer, J.D. Dynamic root exudation of sorgoleone and its in planta mechanism of action. J. Exp. Bot. 2009, 60, 2107–2117. [Google Scholar] [CrossRef]
- Godlewska, K.; Ronga, D.; Michalak, I. Plant extracts—Importance in sustainable agriculture. Ital. J. Agron. 2021, 16, 1851. [Google Scholar] [CrossRef]
- Lebeis, S.L.; Paredes, S.H.; Lundberg, D.S.; Breakfield, N.; Gehring, J.; McDonald, M.; Malfatti, S.; Glavina del Rio, T.; Jones, C.D.; Tringe, S.G.; et al. Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Science 2015, 349, 860–864. [Google Scholar] [CrossRef]
- Siebers, M.; Rohr, T.; Ventura, M.; Schütz, V.; Thies, S.; Kovacic, F.; Jaeger, K.E.; Berg, M.; Dörmann, P.; Schulz, M. Disruption of microbial community composition and identification of plant growth promoting microorganisms after exposure of soil to rapeseed-derived glucosinolates. PLoS ONE 2018, 13, e0200160. [Google Scholar] [CrossRef]
- Hansen, J.C.; Schillinger, W.F.; Sullivan, T.S.; Paulitz, T.C. Soil microbial biomass and fungi reduced with canola introduced into long-term monoculture wheat rotations. Front. Microbiol. 2019, 10, 1488. [Google Scholar] [CrossRef]
- Hu, P.; Hollister, E.B.; Somenahally, A.C.; Hons, F.M.; Gentry, T.J. Soil bacterial and fungal communities respond differently to various isothiocyanates added for biofumigation. Front. Microbiol. 2015, 5, 729. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wu, F. Vanillic acid changed cucumber (Cucumis sativus L.) seedling rhizosphere total bacterial, Pseudomonas and Bacillus spp. communities. Sci. Rep. 2018, 8, 4929. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, J.; Pan, D.; Xin, G.; Jin, X.; Chen, S.; Wu, F. p-Coumaric can alter the composition of cucumber rhizosphere microbial communities and induce negative plant-microbial interactions. Biol. Fert. Soils 2018, 54, 363–372. [Google Scholar] [CrossRef]
- Wang, C.-M.; Li, T.-C.; Jhan, Y.-L.; Weng, J.-H.; Chou, C.-H. The Impact of Microbial Biotransformation of Catechin in Enhancing the Allelopathic Effects of Rhododendron formosanum. PLoS ONE 2013, 8, e85162. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Robert, C.A.M.; Cadot, S.; Zhang, X.; Ye, M.; Li, B.; Manzo, D.; Chervet, N.; Steinger, T.; Van der Heijden, M.G.A.; et al. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Comm. 2018, 9, 2738. [Google Scholar] [CrossRef]
- Kudjordjie, E.N.; Sapkota, R.; Steffensen, S.K.; Fomsgaard, I.S.; Nicolaisen, M. Maize synthesized benzoxazinoids affect the host associated microbiome. Microbiome 2019, 7, 59. [Google Scholar] [CrossRef]
- Schütz, V.; Frindte, K.; Cui, J.; Zhang, P.; Hacquard, S.; Schulze-Lefert, P.; Knief, C.; Schulz, M.; Dörmann, P. Differential Impact of Plant Secondary Metabolites on the Soil Microbiota. Front. Microbiol. 2021, 12, 666010. [Google Scholar] [CrossRef]
- Lehman, R.M.; Cambardella, C.A.; Stott, D.E.; Acosta-Martinez, V.; Manter, D.K.; Buyer, J.S.; Maul, J.E.; Smith, J.L.; Collins, H.P.; Halvorson, J.J.; et al. Understanding and Enhancing Soil Biological Health: The Solution for Reversing Soil Degradation. Sustainability 2015, 7, 988–1027. [Google Scholar] [CrossRef]
- McGivern, B.B.; Tfaily, M.M.; Borton, M.A.; Kosina, S.M.; Daly, R.A.; Nicora, C.D.; Purvine, S.O.; Wong, A.R.; Lipton, M.S.; Hoyt, D.W.; et al. Decrypting bacterial polyphenol metabolism in an anoxic wetland soil. Nat. Commun. 2021, 12, 2466. [Google Scholar] [CrossRef]
- Macías, F.A.; Marín, D.; Oliveros-Bastidas, A.; Castellano, D.; Simonet, A.M.; Molinillo, J.M. Structure-activity relationships (SAR) studies of benzoxazinones, their degradation products and analogues. Phytotoxicity on standard target species (STS). J. Agric. Food Chem. 2005, 53, 538–548. [Google Scholar] [CrossRef]
- Krogh, S.; Mensz, S.; Nielsen, S.; Mortensen, A.; Christophersen, C.; Fomsgaard, I. Fate of Benzoxazinone Allelochemicals in Soil after Incorporation of Wheat and Rye Sprouts. J. Agric. Food Chem. 2006, 54, 1064–1074. [Google Scholar] [CrossRef] [PubMed]
- Schulz, M.; Marocco, A.; Tabaglio, V.; Macias, F.A.; Molinillo, J.M.G. Benzoxazinoids in rye allelopathy—From discovery to application in sustainable weed control and organic farming. J. Chem. Ecol. 2013, 39, 154–174. [Google Scholar] [CrossRef] [PubMed]
- La Hovary, C.; Baerson, S.R.; Williamson, J.D.; Danehower, D.A.; Ma, G.; Pan, Z.; Mask, M.M.; Burton, J.D. Phytotoxicity and benzoxazinone concentration in field grown cereal rye (Secale cereale L.). Int. J. Agron. 2016, 2016, 6463826. [Google Scholar] [CrossRef]
- Hussain, M.I.; Araniti, F.; Schulz, M.; Baerson, S.; Vieites-Álvarez, Y.; Rempelos, L.; Bilsborrow, P.; Durán, A.G.; Chinchilla, N.; Macías, F.A.; et al. Benzoxazinoids in wheat allelopathy—From discovery to application for sustainable weed management. Environ. Exp. Bot. 2022, 202, 104997. [Google Scholar] [CrossRef]
- Fiorini, A.; Maris, S.C.; Abalos, D.; Amaducci, S.; Tabaglio, V. Combining no-till with rye (Secale cereale L.) cover crop mitigates nitrous oxide emissions without decreasing yield. Soil Tillage Res. 2020, 196, 104442. [Google Scholar] [CrossRef]
- Gavazzi, C.; Schulz, M.; Marocco, A.; Tabaglio, V. Sustainable weed control by allelochemicals from rye cover crops: From the greenhouse to field evidence. Allelopath. J. 2010, 25, 259–274. [Google Scholar]
- Tabaglio, V.; Marocco, A.; Schulz, M. Allelopathic cover crop of rye for integrated weed control in sustainable agroecosystems. Ital. J. Agron. 2013, 8, 35–40. [Google Scholar] [CrossRef]
- Boselli, R.; Anders, N.; Fiorini, A.; Ganimede, C.; Faccini, N.; Marocco, A.; Schulz, M.; Tabaglio, V. Improving weed control in sustainable agroecosystems: Role of cultivar and termination timing of rye cover crop. Ital. J. Agron. 2021, 16, 1807. [Google Scholar] [CrossRef]
- Reberg-Horton, S.C.; Burton, J.D.; Danehower, D.A.; Ma, G.; Monks, D.W.; Murphy, J.P.; Ranells, N.N.; Williamson, J.D.; Creamer, N.G. Changes over time in the allelochemical content of ten cultivars of rye (Secale cereale L.). J. Chem. Ecol. 2005, 31, 179–193. [Google Scholar] [CrossRef]
- Tabaglio, V.; Gavazzi, C.; Schulz, M.; Marocco, A. Alternative weed control using the allelopathic effect of natural benzoxazinoids from rye mulch. Agron. Sustain. Dev. 2008, 28, 397–401. [Google Scholar] [CrossRef]
- Ganimede, C.; Marocco, A.; Schulz, M.; Tabaglio, V. Investigation of Endophytic Fungi Associated to Abutilon theophrasti Root. In Proceedings of the 7th World Congress on Allelopathy, Vigo, Spain, 28 July–1 August 2014. [Google Scholar]
- DeMers, M. Alternaria alternata as endophyte and pathogen. Microbiology 2022, 168, 001153. [Google Scholar] [CrossRef]
- Toghueo, R.M.K. Bioprospecting endophytic fungi from Fusarium genus as sources of bioactive metabolites. Mycology 2019, 11, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prudence, S.; Newitt, J.; Worsley, S.F.; Macey, M.C.; Murrell, J.C.; Lehtovirta-Morley, L.E.; Hutchings, M.I. Soil, senescence and exudate utilisation: Characterisation of the Paragon var. spring bread wheat root microbiome. Environ. Microbiome 2021, 16, 12. [Google Scholar] [CrossRef]
- Schulz, M.; Sicker, D.; Schackow, O.; Hennig, L.; Yurkov, A.; Siebers, M.; Hofmann, D.; Disko, U.; Ganimede, C.; Mondani, L.; et al. Interspecies-Cooperations of Abutilon theophrasti with Root Colonizing Microorganisms Disarm BOA-OH Allelochemicals. Plant Signal. Behav. 2017, 12, e1358843. [Google Scholar] [CrossRef]
- Haghi Kia, S. Unraveling Abutilon theophrasti’s Tolerance to the Allelochemical Benzoxazolinone. Master’s Thesis, Rheinische Friedrich-Wilhelms-Universität, Bonn, Germany, 2013. [Google Scholar]
- Kia, S.H.; Schulz, M.; Ayah, E.; Schouten, A.; Müllenborn, C.; Paetz, C.; Schneider, B.; Hofmann, D.; Disko, U.; Tabaglio, V.; et al. Abutilon theophrasti’s defense against the allelochemical benzoxazolin-2(3H)-one: Support by Actinomucor elegans. J. Chem. Ecol. 2014, 40, 1286–1298. [Google Scholar] [CrossRef] [PubMed]
- Kia, S.H.; Wewer, V.; Hölzl, G.; Schulz, M. Assessing the Effect of Benzoxazolinone on the Plant–Associated Zygomycete Actinomucor elegans and Its Possible Role in a Weed’s Tolerance to Allelochemicals. In Proceedings of the 10th International Mycological Congress (Imc10), Bangkok, Thailand, 3–8 August 2014; Available online: http://www.imc10.com (accessed on 31 January 2023).
- Nadane, A. Effect of Velvetleaf Root Exudates on Indole-3-Acetic Acid Production by Rhizobium rubi AT3-4RS/6. Ph.D. Thesis, Andrews University, Berrien Springs, MI, USA, 2012. Available online: https://digitalcommons.andrews.edu/honors/24 (accessed on 31 January 2023).
- Schulz, M. (University of Bonn, Bonn, Germany); Hofmann, D. (Forschungszentrum Jülich GmbH, Jülich, Germany). Personal communication, 2022. [Google Scholar]
- Schulz, M.; Sicker, D.; Schackow, O.; Hennig, L.; Hofmann, D.; Disko, U.; Ventura, M.; Basyuk, K. 6-Hydroxy-5-nitrobenzo[d]oxazol- 2(3H)-one—A degradable derivative of natural 6-hydroxybenzoxazolin-2(3H)-one produced by Pantoea ananatis. Commun. Integr. Biol. 2017, 10, e1302633. [Google Scholar] [CrossRef] [PubMed]
- Schulz, M.; Marocco, A.; Tabaglio, V. BOA detoxification of four summer weeds during germination and seedling growth. J. Chem. Ecol. 2012, 38, 933–946. [Google Scholar] [CrossRef]
- Laschke, L.; Schütz, V.; Schackow, O.; Sicker, D.; Hennig, L.; Hofmann, D.; Dörmann, P.; Schulz, M. Survival of Plants during Short-Term BOA-OH Exposure: ROS Related Gene Expression and Detoxification Reactions Are Accompanied with Fast Membrane Lipid Repair in Root Tips. J. Chem. Ecol. 2022, 48, 219–239. [Google Scholar] [CrossRef]
- Sterling, T.M.; Houtz, R.L.; Putnam, A.R. Phytotoxic Exudates from Velvetleaf (Abutilon theophrasti) Glandular Trichomes. Am. J. Bot. 1987, 74, 543–550. [Google Scholar] [CrossRef]
- Radivojević, L.; Sarić-Krsmanović, M.; Umiljendić, J.G.; Šantrić, L. Allelopathic effects of invasive weed species Abutilon theophrasti Medik., Ambrosia artemisiifolia L., Datura stramonium L. and Xanthium strumarium L. on tomato. Pestic. Fitomed. 2019, 34, 183–191. [Google Scholar] [CrossRef]
- Tian, M.; Li, Q.; Zhao, W.; Qiao, B.; Shi, S.; Yu, M.; Li, X.; Li, C.; Zhao, C. Potential Allelopathic Interference of Abutilon theophrasti Medik. Powder/Extract on Seed Germination, Seedling Growth and Root System Activity of Maize, Wheat and Soybean. Agronomy 2022, 12, 844. [Google Scholar] [CrossRef]
- Novak, N.; Novak, M.; Barić, K.; Šćepanović, M.; Ivić, D. Allelopathic potential of segetal and ruderal invasive alien plants. J. Cent. Eur. Agric. 2018, 19, 408–422. [Google Scholar] [CrossRef] [Green Version]
- Paszkowski, W.L.; Kremer, R.J. Biological activity and tentative identification of flavonoid components in velvetleaf (Abutilon theophrasti Medik.) seed coats. J. Chem. Ecol. 1988, 14, 1573–1582. [Google Scholar] [CrossRef] [PubMed]
- Balah, M.A.; Nassar, M.I. Allelopathic Constituents from Abutilon theophrasti Aerial Parts to Other Weeds. Res. J. Agric. Sci. 2011, 7, 243–250. [Google Scholar]
- Tian, C.; Yang, C.; Zhang, D.; Han, L.; Liu, Y.; Guo, W.; Fan, R.; Liu, M. Antibacterial and antioxidant properties of various solvent extracts of Abutilon theophrasti Medic. leaves. Pak. J. Pharm. Sci. 2017, 3, 767–772. [Google Scholar]
- Hwang, J.H.; Jin, Q.; Woo, E.R.; Lee, D.G. Antifungal property of hibicuslide C and its membrane-active mechanism in Candida albicans. Biochimie 2013, 95, 1917–1922. [Google Scholar] [CrossRef]
- Mamadalieva, N.Z.F.; Sharopov, F.; Girault, J.-P.; Wink, M.; Lafont, R. Phytochemical analysis and bioactivity of the aerial parts of Abutilon theophrasti (Malvaceae), a medicinal weed. Nat. Prod. Res. 2014, 28, 1777–1779. [Google Scholar] [CrossRef]
- Itoh, A.; Fujii, K.; Tomatsu, S.; Takao, C.; Tanahashi, T.; Nagakura, N.; Chen, C.-C. Six secoiridoid glucosides from Adina racemosa. J. Nat. Prod. 2003, 66, 1212–1216. [Google Scholar] [CrossRef]
- Murai, Y.; Kashimura, S.; Tamezawa, S.; Hashimoto, T.; Takaoka, S.; Asakawa, Y.; Kiguchi, K.; Murai, F.; Tagawa, M. Absolute Configuration of (6S,9S)-Roseoside from Polygonum hydropiper. Planta Med. 2001, 67, 480–481. [Google Scholar] [CrossRef]
- Rumalla, C.S.; Ali, Z.; Weerasooriya, A.D.; Smillie, T.J.; Khan, I.J. A new triterpene glycoside from Centella erecta. Fitoterapia 2010, 81, 751–754. [Google Scholar] [CrossRef]
- Zhang, S.; Gai, Z.; Gui, T.; Chen, J.; Chen, Q.; Li, Y. Antioxidant Effects of Protocatechuic Acid and Protocatechuic Aldehyde: Old Wine in a New Bottle. Evid. Based Complement Alternat. Med. 2021, 8, 6139308. [Google Scholar] [CrossRef]
- Sterling, T.M.; Balke, N.E. Use of Soybean (Glycine max) and Velvetleaf (Abutilon theophrasti) Suspension-Cultured Cells to Study Bentazon Metabolism. Weed Sci. 1988, 36, 558–565. Available online: https://www.jstor.org/stable/4044806 (accessed on 31 January 2023). [CrossRef]
- Reese, A.T.; Chadaideh, K.S.; Caroline, E.; Diggins, C.E.; Schell, L.D.; Beckel, M.; Callahan, P.; Ryan, R.; Thompson, E.M.; Carmody, R.N. Effects of domestication on the gut microbiota parallel those of human industrialization. eLife 2021, 10, e60197. [Google Scholar] [CrossRef]
- Soldan, R.; Fusi, M.; Cardinale, M.; Daffonchio, D.; Preston, G.M. The effect of plant domestication on host control of the microbiota. Commun. Biol. 2021, 4, 936. [Google Scholar] [CrossRef] [PubMed]
- Raaijmakers, J.M.; Kiers, E.T. Rewilding plant microbiomes. Science 2022, 378, 599–600. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Cernava, T. The plant microbiota signature of the Anthropocene as a challenge for microbiome research. Microbiome 2022, 10, 54. [Google Scholar] [CrossRef]
- García-Campa, L.; Guerrero, S.; Lamelas, L.; Meijon, M.; Hasbún, R.; Cañal, M.J.; Valledor, L. Chloroplast proteomics reveals transgenerational cross-stress priming in Pinus radiata. Environ. Exp. Bot. 2022, 202, 105009. [Google Scholar] [CrossRef]
- Lagage, V.; Uphoff, S. Pulses and delays, anticipation and memory: Seeing bacterial stress responses from a single-cell perspective. FEMS Microbiol. Rev. 2020, 44, 565–571. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
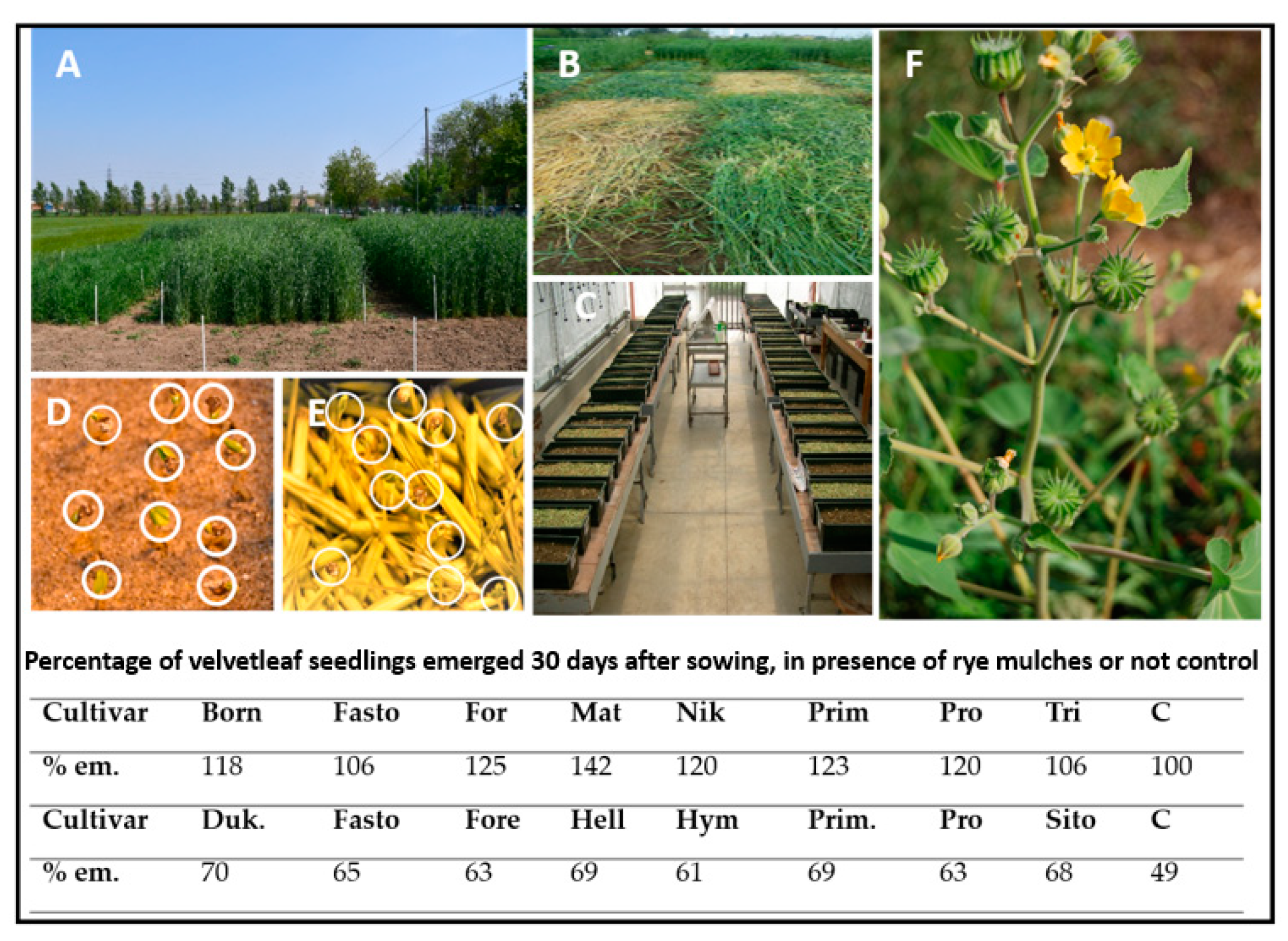{kind=link}
{kind=link}
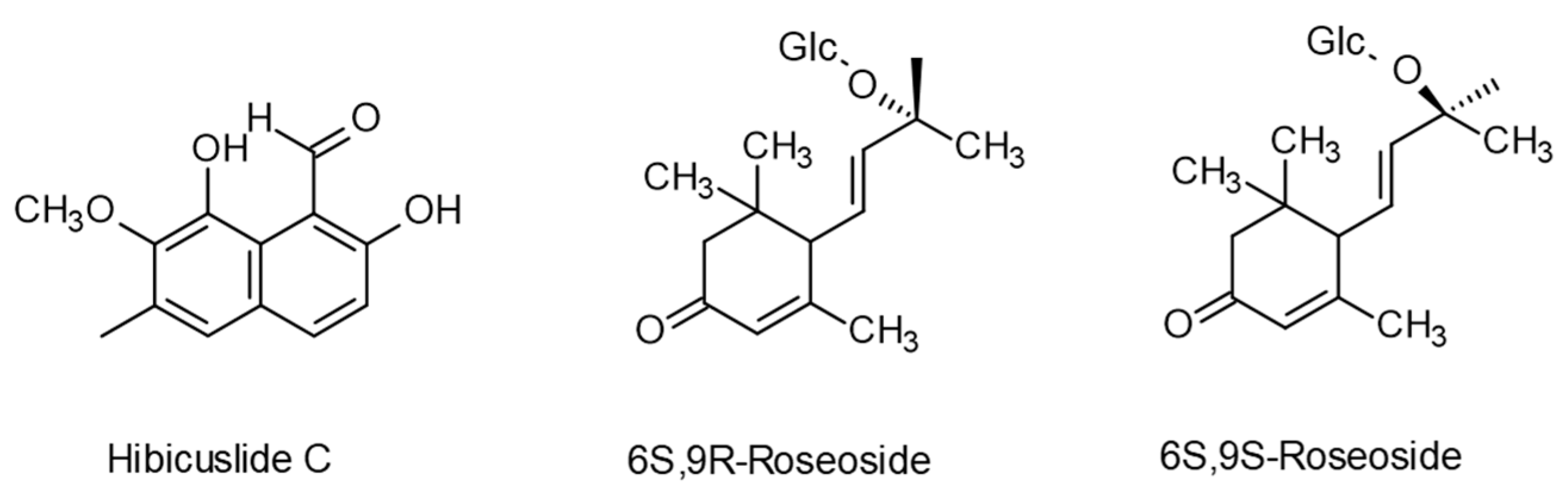{kind=link}
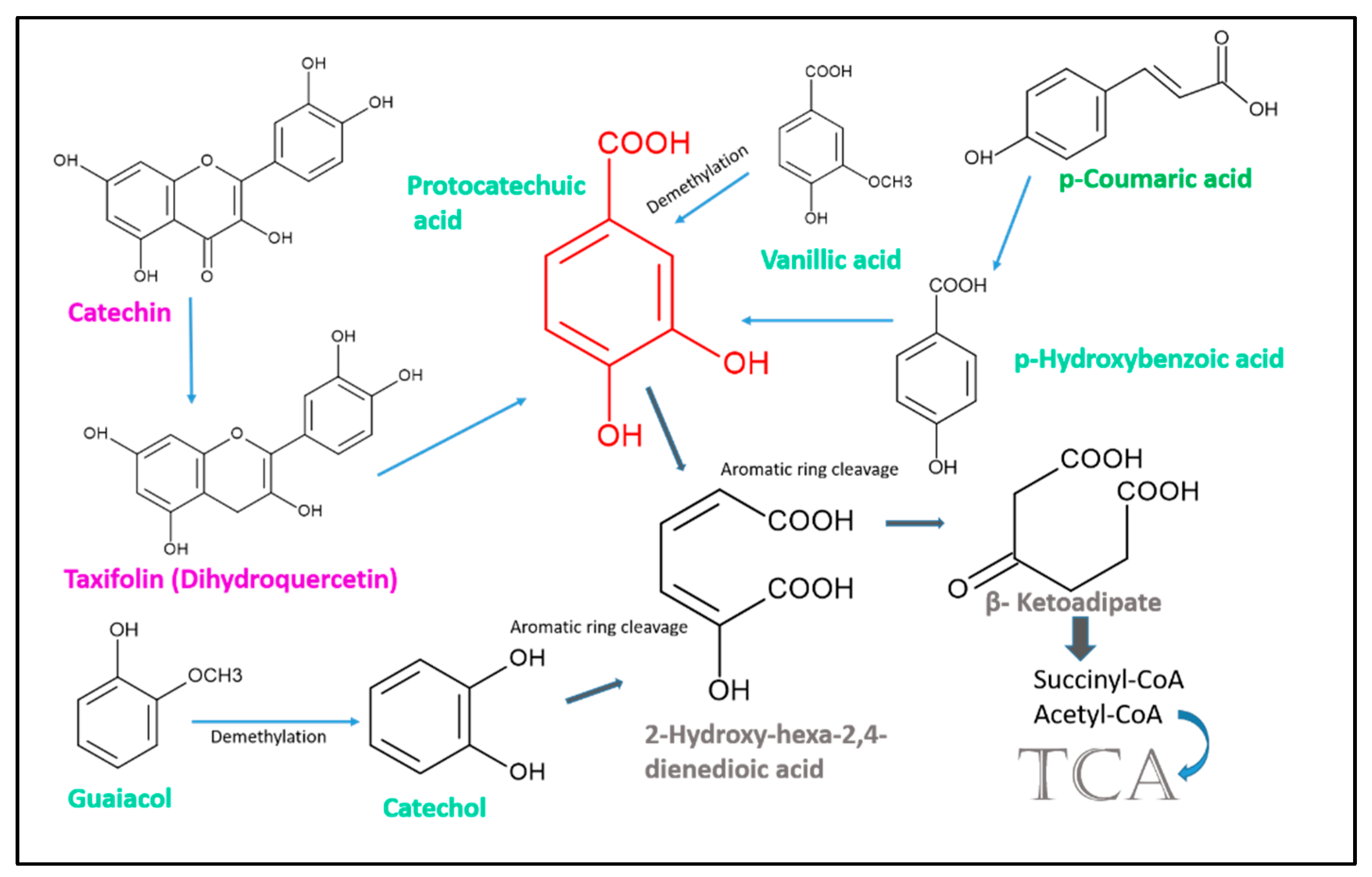{kind=link}
| Microorganisms | Mode of Action | Note | Reference |
|---|---|---|---|
| Fungi | |||
| Alternaria abutilonis | leaf spot | [2] | |
| Cercospora abutilonis Cerospora spec. complex? | Leaf spot | species needs additional identification | [2] |
| Cercospora althaeina Sac. | Leaf rot | [2] | |
| Cladosporium herbarum | secondary leaf spot | [2] | |
| Colletotrichum coccodes | foliar pathogen, plant death; suppression of plant defenses | enhancement of virulence by mannose and oxalic acid, NEP1 for hypervirulent transgenic strain | [2,39,40,45] |
| Colletotrichum malvarum | leaf spot | [2] | |
| Fusarium lateritium | growth suppression | [2,35] | |
| Fusarium oxysporum | 24-kDa protein Nep1 induces necrosis | [44] | |
| Macrophomina phaseoli | stem rot | [2] | |
| Phyllosticta althaeina Sacc. | Leaf rot | [2] | |
| Phymatotrichum omnivorum | root rot | [2] | |
| Puccinia heterospora | rust | possible mycoherbicide | [2] |
| Verticillium dahliae, V. nigrescens | wilt | suggested as a biocontrol agent, unspecific | [2] |
| Bacteria | |||
| Acidovorax delafieldii ATH2-2RS/1 | inhibits growth | [48] | |
| Pseudomonas putida ATH-1RI/9 | inhibits growth | [48] | |
| Pseudomonas spec isolates Pp001 and Pf239 | Reduction of seedlings emergence weaken seedlings | [32] | |
| Streptomyces isolates | inhibits germination, growth | [46] |
| Microorganisms | Seed Surface | Within Seeds | Location | Activity | Reference |
|---|---|---|---|---|---|
| Fungi | |||||
| Actinomucor elegans, strain AF157119 | x x | 6 7 | protects Abutilon seedlings | [53,89,90,91] | |
| Alternaria alternata | x | x | 1, 2, 3 | [30,86] | |
| Aspergillus flavus | x | 7 | [53] | ||
| Cephaliophora tropica strains | x | 8 | [53] | ||
| Cladosporium cladosporioides | x | 1, 2 | [30] | ||
| Cordyceps sinensis | x | 7, 8 | [53] | ||
| Epicoccumpurpurascens | x | 1, 2 | inhibits seed-borne bacteria | [30] | |
| Fusarium spp. | x | x | 1, 2, 3, 4, 5 | moderate antagonism toward test fungi | [30,86] |
| Madurella mycetomatis | x | 8 | [53] | ||
| Bacteria | |||||
| Alcaligenes spec. isolates | 1, 2 | highly antagonistic to seed fungi | [30] | ||
| Bacillus spec. isolates | 1, 2 | [30] | |||
| Bacillus subtilis | 1, 2 | [30] | |||
| Flavobacterium spec. | 1, 2 | [30] | |||
| Pseudomonas spec. isolates | 1, 2 | [30] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tabaglio, V.; Fiorini, A.; Sterling, T.M.; Schulz, M. Abutilon theophrasti’s Resilience against Allelochemical-Based Weed Management in Sustainable Agriculture – Due to Collection of Highly Advantageous Microorganisms? Plants 2023, 12, 700. https://doi.org/10.3390/plants12040700
Tabaglio V, Fiorini A, Sterling TM, Schulz M. Abutilon theophrasti’s Resilience against Allelochemical-Based Weed Management in Sustainable Agriculture – Due to Collection of Highly Advantageous Microorganisms? Plants. 2023; 12(4):700. https://doi.org/10.3390/plants12040700
Chicago/Turabian StyleTabaglio, Vincenzo, Andrea Fiorini, Tracy M. Sterling, and Margot Schulz. 2023. "Abutilon theophrasti’s Resilience against Allelochemical-Based Weed Management in Sustainable Agriculture – Due to Collection of Highly Advantageous Microorganisms?" Plants 12, no. 4: 700. https://doi.org/10.3390/plants12040700
APA StyleTabaglio, V., Fiorini, A., Sterling, T. M., & Schulz, M. (2023). Abutilon theophrasti’s Resilience against Allelochemical-Based Weed Management in Sustainable Agriculture – Due to Collection of Highly Advantageous Microorganisms? Plants, 12(4), 700. https://doi.org/10.3390/plants12040700