
The Enhanced Affinity of WRKY Reinforces Drought Tolerance in Solanum lycopersicum L.: An Innovative Bioinformatics Study
 , ,
, ,  , , , , ,
, , , , ,  and
and 
Abstract
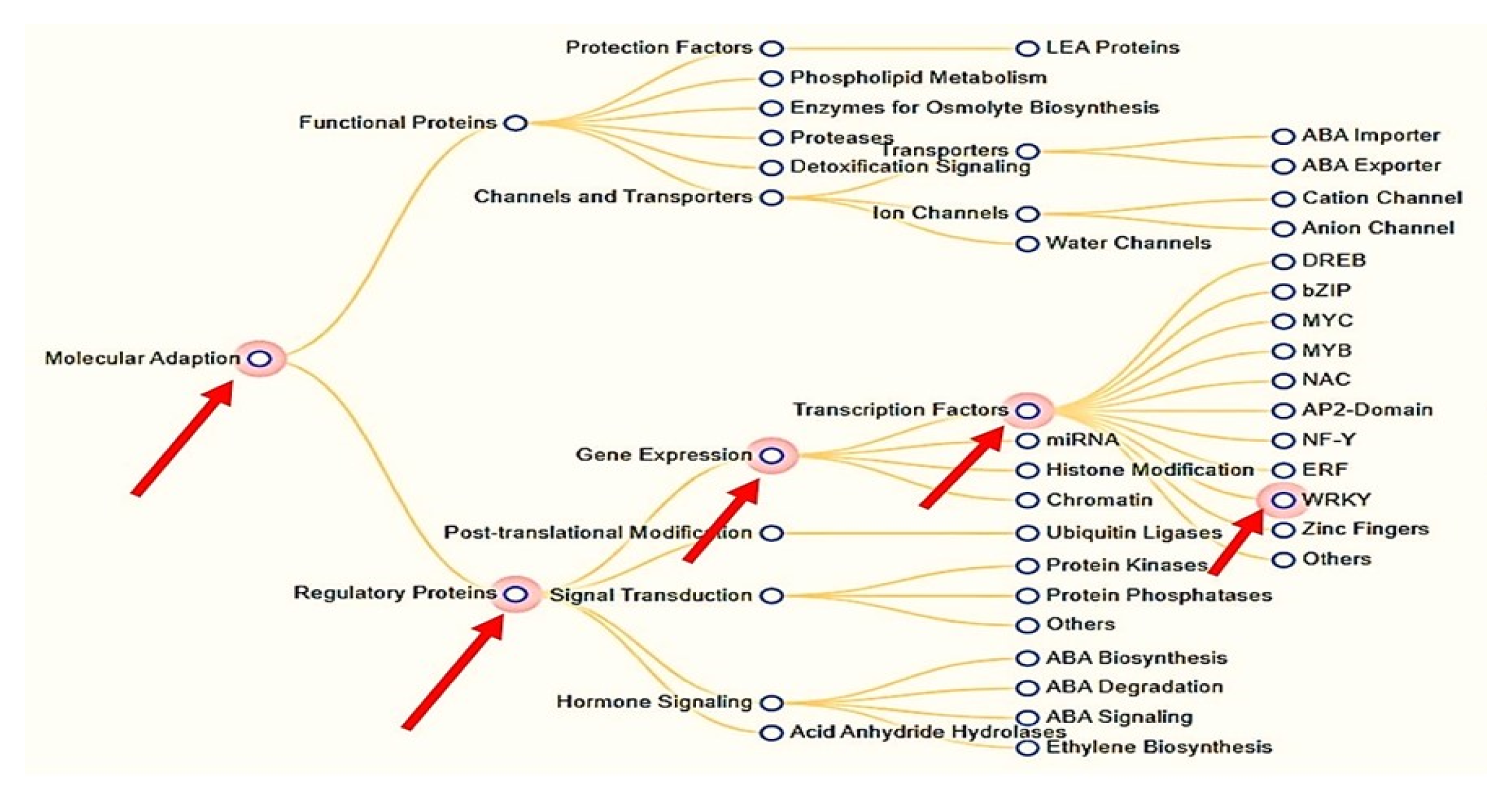
:1. Introduction
2. Results
2.1. Virtual Screening of Compounds
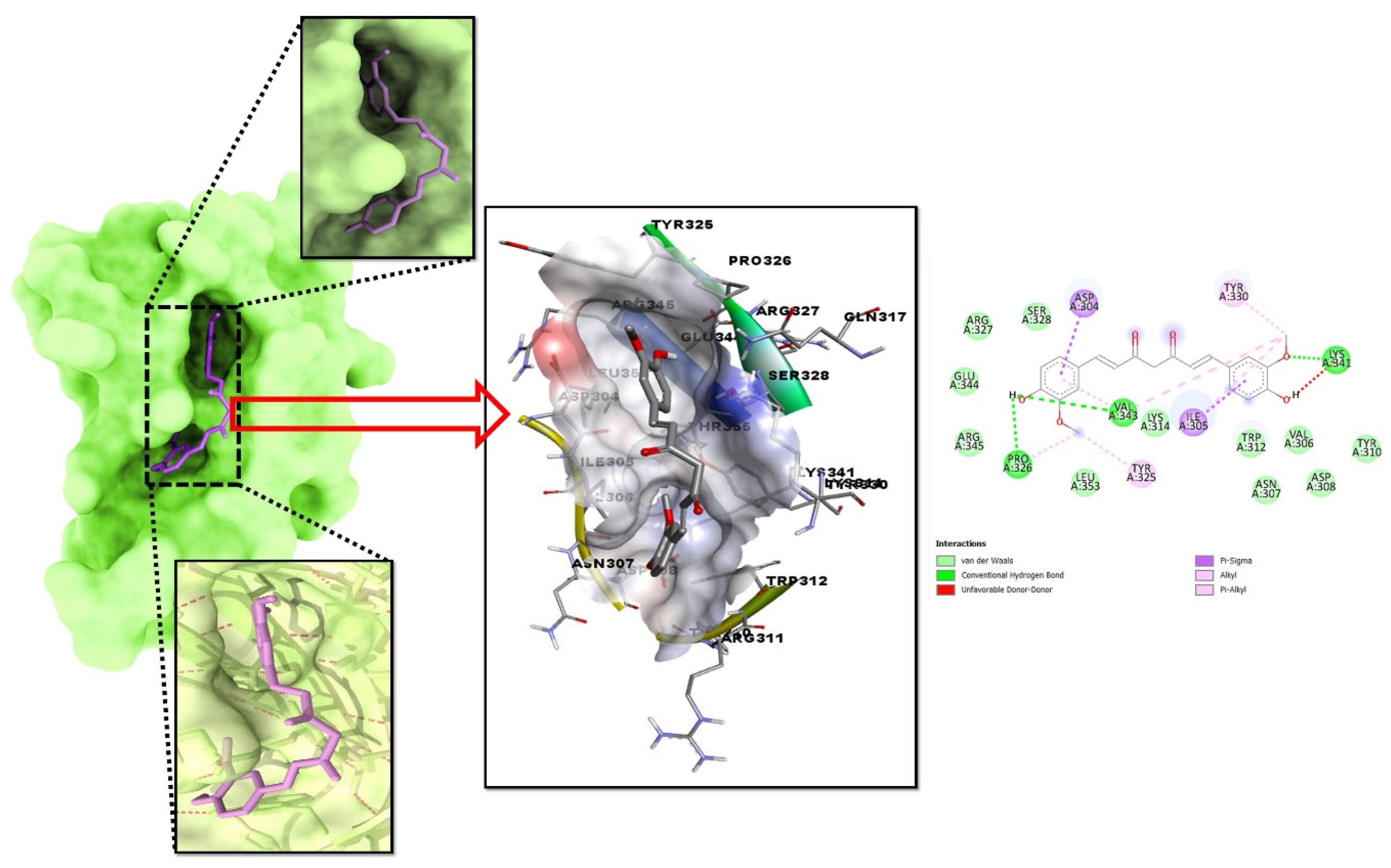
2.2. Molecular Docking Investigation
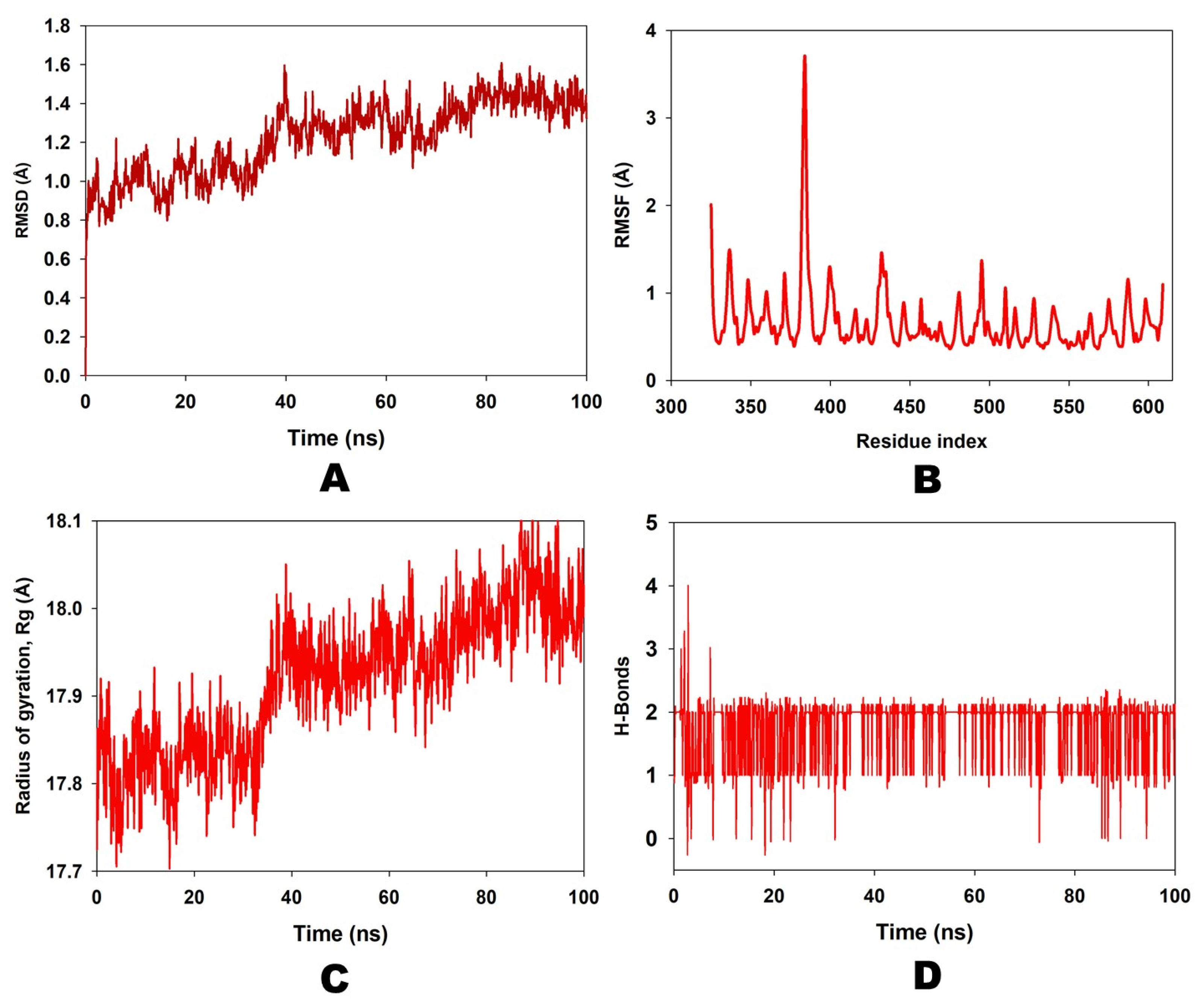
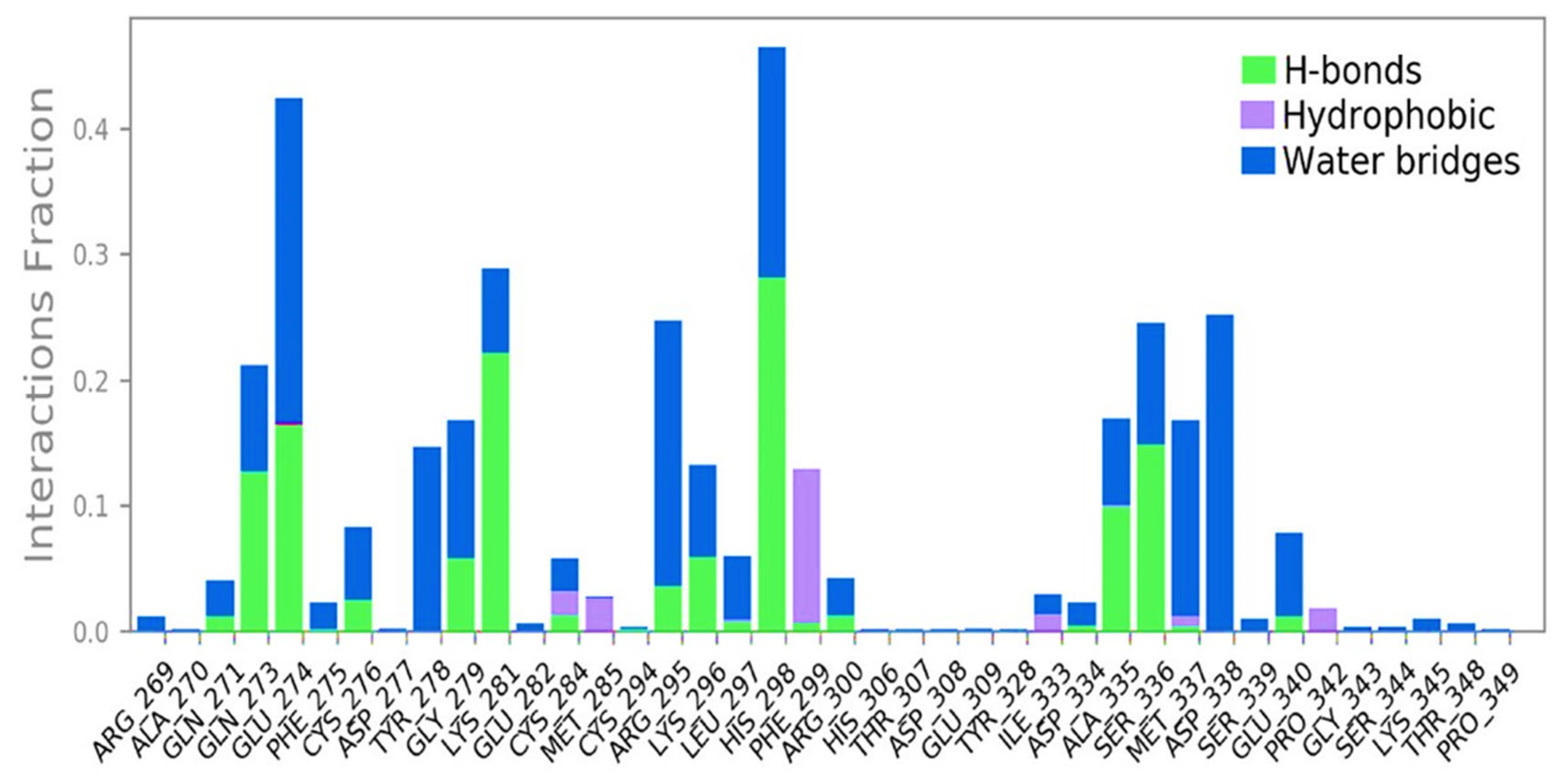
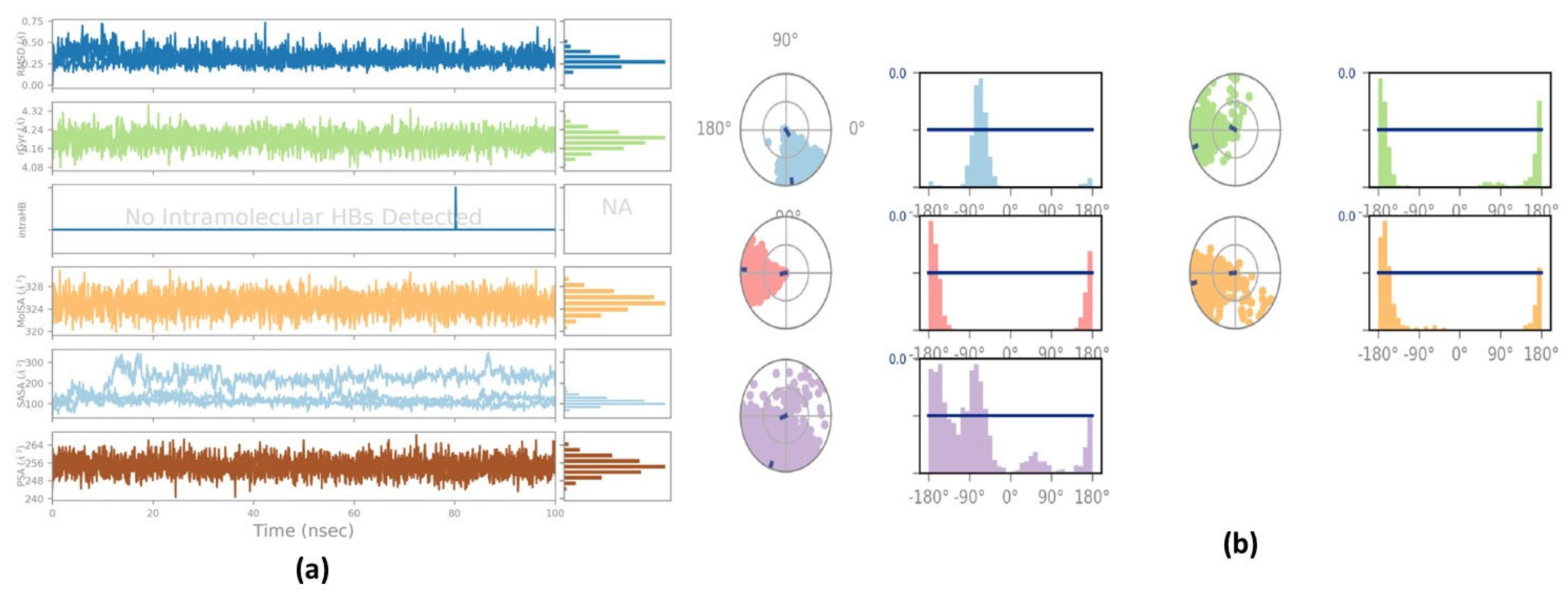
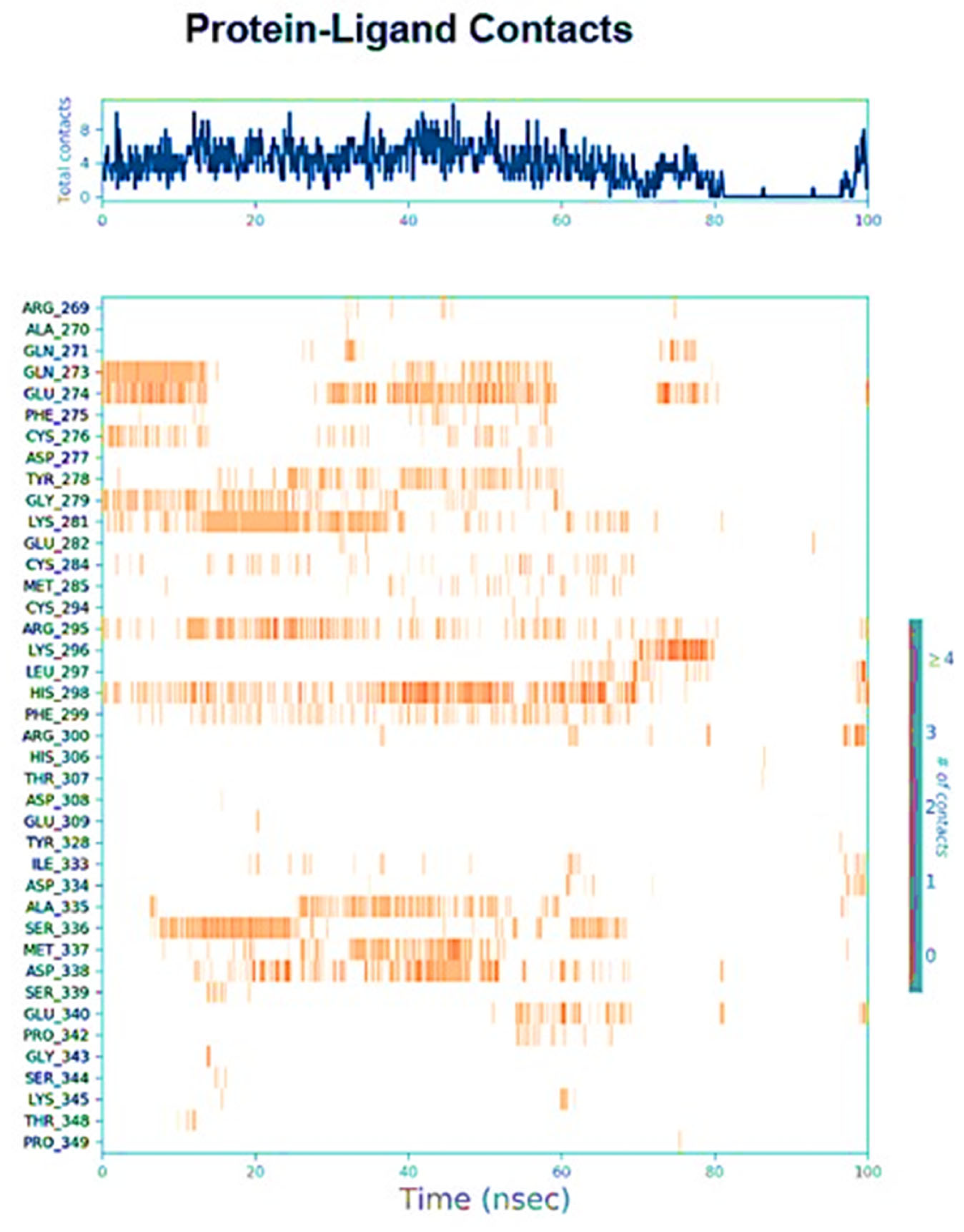
2.3. Molecular Dynamic Simulation Study
2.4. Molecular Mechanics Generalized Born Surface Area (MM-GBSA) Calculations
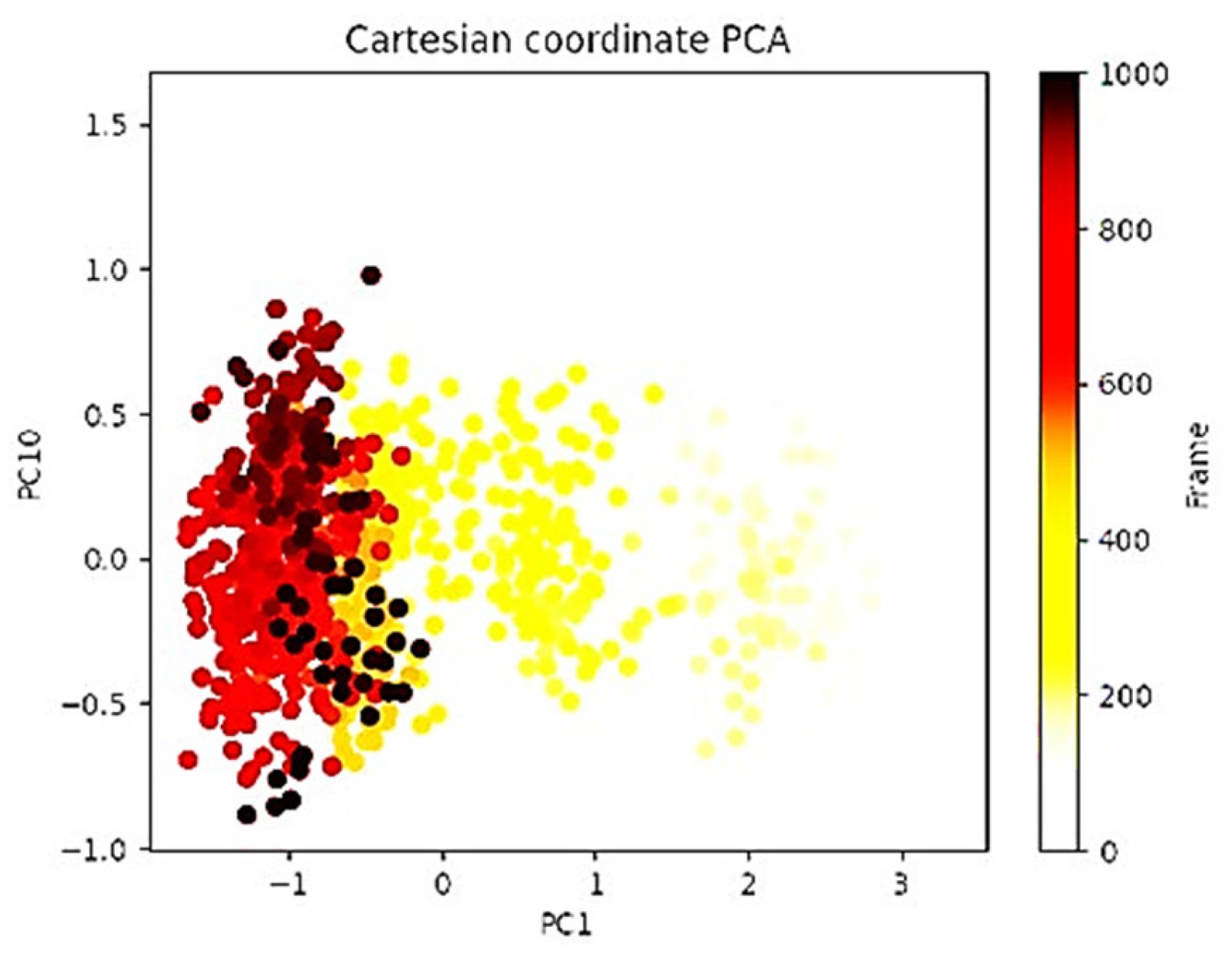
2.5. Principal Component Analysis
3. Materials and Methods
3.1. Potential Target Preparation
3.2. Virtual Screening of Selected Compounds
3.3. Molecular Docking Studies
3.4. Molecular Dynamic Simulations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conti, V.; Mareri, L.; Faleri, C.; Nepi, M.; Romi, M.; Cai, G.; Cantini, C. Drought Stress Affects the Response of Italian Local Tomato (Solanumlycopersicum L.) Varieties in a Genotype-Dependent Manner. Plants 2019, 8, 336. [Google Scholar] [CrossRef] [PubMed]
- Perveen, K.; Alfagham, A.T.; Debnath, S.; Bukhari, N.A.; Wei, D.; Alshaikh, N.A.; Alwadai, A.S. Enriching drought resistance in Solanum lycopersicum using Abscisic acid as drought enhancer derived from Lygodium japonicum: A new-fangled computational approach. Front. Plant Sci. 2023, 14. [Google Scholar] [CrossRef]
- Ikinci, A.; Bolat, I.; Ercisli, S.; Kodad, O. Influence of rootstocks on growth, yield, fruit quality and leaf mineral element contents of pear cv.‘SantaMaria’in semi-arid conditions. Biol. Res. 2014, 47, 71. [Google Scholar] [CrossRef] [PubMed]
- Kayacetin, F. Assessment of safflower genotypes for individual and combined effects of drought and salinity stress at early seedling growth stages. Turk. J. Agric. For. 2022, 46, 601–612. [Google Scholar] [CrossRef]
- KAYMAK, H.Ç.; Sevim, M.; METİN, Ö. Graphene oxide: A promising material for the germination of melon seeds under salinity stress. Turk. J. Agric. For. 2022, 46, 863–874. [Google Scholar] [CrossRef]
- Ximénez-Embún, M.G.; González-Guzmán, M.; Arbona, V.; Gómez-Cadenas, A.; Ortego, F.; Castañera, P. Plant-mediated effects of water deficit on the performance of Tetranychusevansi on tomato drought-adapted accessions. Front. Plant Sci. 2018, 9, 1490. [Google Scholar] [CrossRef]
- Li, D.; Zaman, W.; Lu, J.; Niu, Q.; Zhang, X.; Ayaz, A.; Saqib, S.; Yang, B.; Zhang, J.; Zhao, H.; et al. Natural lupeol level variation among castor accessions and the upregulation of lupeol synthesis in response to light. Ind. Crops Products 2023, 192, 116090. [Google Scholar] [CrossRef]
- Khoso, M.A.; Hussain, A.; Ritonga, F.N.; Ali, Q.; Channa, M.M.; Alshegaihi, R.M.; Meng, Q.; Ali, M.; Zaman, W.; Brohi, R.D.; et al. WRKY transcription factors (TFs): Molecular switches to regulate drought, temperature, and salinity stresses in plants. Front. Plant Sci. 2022, 13, 1039329. [Google Scholar] [CrossRef]
- Kçhler, F.E. Kçhler’s Medizinal-Pflanzen in Naturge-Treuen Abbildungen Mit Kurz Erläuterndem Texte: Atlas Zur Pharmacopoea Germanica; Gera-Untermhaus: Gera, Germany, 1887. [Google Scholar]
- Ammon, H.; Wahl, M.A. Pharmacology of Curcuma longa. Planta Med. 1991, 57, 1–7. [Google Scholar] [CrossRef]
- Abrahamian, P.; Klein, J.M.; Jones, J.B.; Vallad, G.E.; Melanson, R.A. First report of bacterial spot of tomato caused by Xanthomonas perforans in Mississippi. Plant Dis. 2019, 103, 147. [Google Scholar] [CrossRef]
- Rushton, P.J.; Torres, J.T.; Parniske, M.; Wernert, P.; Hahlbrock, K.; Somssich, I.E. Interaction of elicitor induced DNA-binding proteins with elicitor response elements in the promoters of parsley PR1 genes. EMBO J. 1996, 15, 5690–5700. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Jeffrey, R.S.; Dawn, N.; Qingxi, J.S. AWRKY gene from creosote bush encodes an activator of the Curcumin signaling pathway. J. Biol. Chem. 2004, 279, 55770–55779. [Google Scholar] [CrossRef] [PubMed]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Sundaram, C.; Malani, N.; Ichikawa, H. Curcumin: The Indian solid gold. Adv. Exp. Med. Biol. 2007, 595, 1–75. [Google Scholar]
- Tilak, J.; Banerjee, M.; Mohan, H.; Devasagayam, T.P.A. Antioxidant availability of turmeric in relation to its medicinal and culinary uses. Phytother Res. 2004, 18, 798–804. [Google Scholar] [CrossRef]
- Guo, Y.; Cai, Z.; Gan, S. Transcriptome of Arabidopsis leaf senescence. Plant Cell Environ. 2004, 27, 521–549. [Google Scholar] [CrossRef]
- Xie, Z.; Ruas, P.; Shen, Q.J. Regulatory networks of the phytohormone Curcumin. Vitam. Horm. 2005, 72, 235–269. [Google Scholar]
- Alexandrova, K.S.; Conger, B.V. Isolation of two somatic embryogenesis-related genes from orchard grass (Dactylisglomerata). Plant Sci. 2002, 162, 301–307. [Google Scholar] [CrossRef]
- Chen, C.; Chen, Z. Isolation and characterization of two pathogen and salicylic acid-induced genes encoding WRKY DNA-binding proteins from tobacco. Plant Mol. Biol. 2000, 42, 387–396. [Google Scholar] [CrossRef]
- Cormack, R.S.; Eulgem, T.; Rushton, P.J.; Köchner, P.; Hahlbrock, K.; Somssich, I. Leucine zipper containing WRKY proteins widen the spectrum of immediate early elicitor-induced WRKY transcription factors in parsley. Biochim. Biophys. Acta 2002, 1576, 92–100. [Google Scholar] [CrossRef]
- Xu, X.P.; Chen, C.; Fan, B.; Chen, Z. Physical and functional interactions between pathogen-induced Arabidopsis WRKY18, WRKY40, and WRKY60 transcription factors. Plant Cell 2006, 18, 1310–1326. [Google Scholar]
- Krishna, R.; Ansari, W.A.; Soumia, P.S.; Yadav, A.; Jaiswal, D.K.; Kumar, S.; Singh, A.K.; Singh, M.; Verma, J.P. Biotechnological Interventions in Tomato (Solanumlycopersicum) for Drought Stress Tolerance: Achievements and Future Prospects. Biotechnology 2022, 11, 48. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.-H.; Lee, J.-T.; Charng, Y.-Y.; Chan, M.-T. Tomato Plants Ectopically Expressing Arabidopsis CBF1 Show Enhanced Resistance to Water Deficit Stress. Plant Physiol. 2002, 130, 618–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, L.; Santos, R.D.; Oliva, G.; Andricopulo, A. Molecular Docking and Structure-Based Drug Design Strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [PubMed]
- Meng, X.Y.; Zhang, H.X.; Mezei, M.; Cui, M. Molecular Docking: A powerful approach for structure-based drug discovery. Curr. Comput. Aided Drug Des. 2011, 7, 146–157. [Google Scholar]
- Chow, E.; Rendleman, C.A.; Bowers, K.J.; Dror, R.O.; Hughes, D.H.; Gullingsrud, J.; Sacerdoti, F.D.; Shaw, D.E. Desmond Performance on a Cluster of Multicore Processors. DE Shaw Research Technical Report DESRES. DESRES/TR-2008-01. Available online: http://deshawresearch.com (accessed on 12 January 2023).
- Jorgensen, W.L.; Maxwell, D.S.; Tirado-Rives, J. Development and testing of the OPLS all-atom force field on conformational energetics and properties of organic liquids. J. Am. Chem. Soc. 1996, 118, 11225–11236. [Google Scholar] [CrossRef]
- Shivakumar, D.; Williams, J.; Wu, Y.; Damm, W.; Shelley, J.; Sherman, W. Prediction of absolute solvation free energies using molecular dynamics free energy perturbation and the OPLS force field. J. Chem. Theory Comput. 2010, 6, 1509–1519. [Google Scholar] [CrossRef]
- Martyna, G.J.; Klein, M.L.; Tuckerman, M. Nose-Hoover chains-the canonical ensemble via continuous dynamics. J. Chem. Phys. 1992, 97, 2635–2643. [Google Scholar]
- Martyna, G.J.; Tobias, D.J.; Klein, M.L. Constant pressure molecular dynamics algorithms. J. Chem. Phys. 1994, 101, 4177–4189. [Google Scholar] [CrossRef]
- Alamri, A.H.; Debnath, S.; Alqahtani, T.; Alqahtani, A.; Alshehri, S.A.; Ghosh, A. Enhancing plant-derived smart nano inhibitor in targeting mammalian target of rapamycin (mTOR) in breast cancer using Curcuma longa-derived compound curcumin. Environ. Sci. Pollut. Res. 2023, 2023, 1–8. [Google Scholar] [CrossRef]
- Goodsell, D.S.; Morris, G.M.; Olson, A.J. Automated docking of flexible ligands: Applications of AutoDock. J. MolRecognit. 1996, 9, 1–5. [Google Scholar] [CrossRef]
- Toukmaji, A.Y.; Board, J.A., Jr. Ewald summation techniques in perspective: A survey. Comput. Phys. Commun. 1996, 95, 73–92. [Google Scholar] [CrossRef]
- Bowers, K.J.; Chow, D.E.; Xu, H.; Dror, R.O.; Eastwood, M.P.; Gregersen, B.A.; Klepeis, J.L.; Kolossvary, I.; Moraes, M.A.; Sacerdoti, F.D.; et al. Scalable algorithms for molecular dynamics simulations oncommodity clusters. In Proceedings of the 2006 ACM/IEEE Conference on Supercomputing, Tampa, FL, USA, 1–17 November 2006; p. 43. [Google Scholar]
- Steuer, B.; Stuhlfauth, T.; Fock, H.P. The efficiency of water use in water stressed plants is increased due to LN induced stomatal closure. Photosynth. Res. 1988, 18, 327–336. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl No. | Compound ID (CID) | Binding Energy (kcal/mol) |
|---|---|---|
| 1. | CID_100332 | −5.6 |
| 2. | CID_101389368 | −5.5 |
| 3. | CID_119034 | −6.4 |
| 4. | CID_15559069 | −7.1 |
| 5. | CID_241572 | −7.0 |
| 6. | CID_969516 | −11.43 |
| 7. | CID_3981577 | −6.9 |
| 8. | CID_5280343 | −5.3 |
| 9. | CID_5280443 | −7.0 |
| 10. | CID_5280445 | −6.2 |
| Energies (kcal/mol) | Curcumin |
|---|---|
| ΔGbind | −35.56 ± 2.23 |
| ΔGbindLipo | −11.63 ± 1.6 |
| ΔGbindvdW | −10.22 ± 2.3 |
| ΔGbindCoulomb | −6.22 ± 1.0 |
| ΔGbindHbond | −0.45 ± 0.2 |
| ΔGbindSolvGB | 9.36 ± 2.1 |
| ΔGbindCovalent | 2.75 ± 1.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Debnath, S.; Kant, A.; Bhowmick, P.; Malakar, A.; Purkaystha, S.; Jena, B.K.; Mudgal, G.; Rahimi, M.; Helal, M.M.U.; Hasan, R.; et al. The Enhanced Affinity of WRKY Reinforces Drought Tolerance in Solanum lycopersicum L.: An Innovative Bioinformatics Study. Plants 2023, 12, 762. https://doi.org/10.3390/plants12040762
Debnath S, Kant A, Bhowmick P, Malakar A, Purkaystha S, Jena BK, Mudgal G, Rahimi M, Helal MMU, Hasan R, et al. The Enhanced Affinity of WRKY Reinforces Drought Tolerance in Solanum lycopersicum L.: An Innovative Bioinformatics Study. Plants. 2023; 12(4):762. https://doi.org/10.3390/plants12040762
Chicago/Turabian StyleDebnath, Sandip, Achal Kant, Pradipta Bhowmick, Ayushman Malakar, Shampa Purkaystha, Binod Kumar Jena, Gaurav Mudgal, Mehdi Rahimi, Md Mostofa Uddin Helal, Rakibul Hasan, and et al. 2023. "The Enhanced Affinity of WRKY Reinforces Drought Tolerance in Solanum lycopersicum L.: An Innovative Bioinformatics Study" Plants 12, no. 4: 762. https://doi.org/10.3390/plants12040762
APA StyleDebnath, S., Kant, A., Bhowmick, P., Malakar, A., Purkaystha, S., Jena, B. K., Mudgal, G., Rahimi, M., Helal, M. M. U., Hasan, R., Chen, J. -T., & Azam, F. (2023). The Enhanced Affinity of WRKY Reinforces Drought Tolerance in Solanum lycopersicum L.: An Innovative Bioinformatics Study. Plants, 12(4), 762. https://doi.org/10.3390/plants12040762