
Screening and Verification of Reference Genes for Analysis of Gene Expression in Garlic (Allium sativum L.) under Cold and Drought Stress

Abstract
:1. Introduction
2. Results
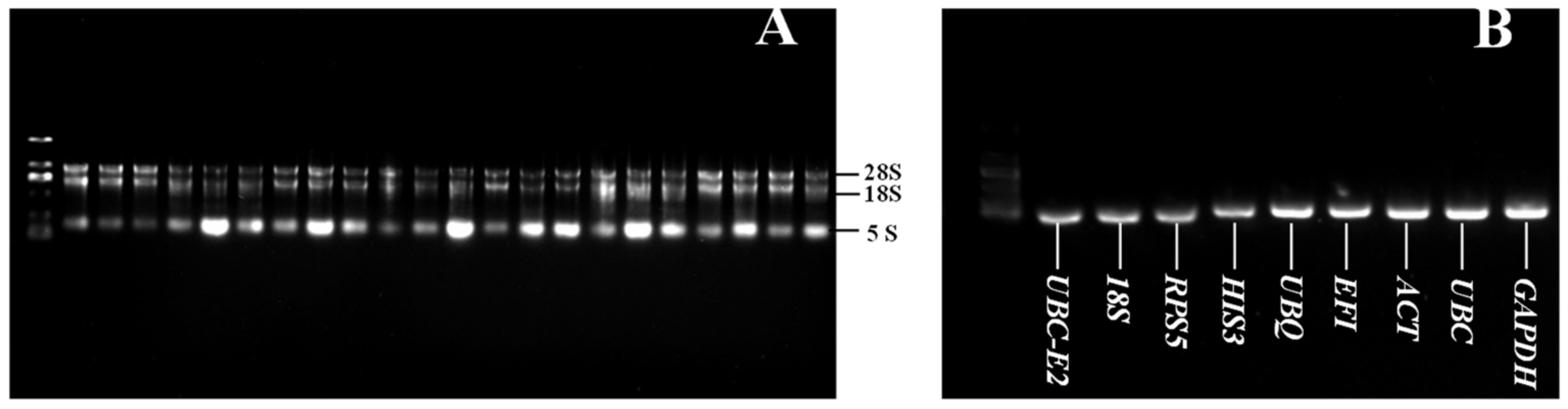
2.1. RNA Quality Detection and Primer Specificity Analysis of Reference Genes
2.2. Assessment of Primer Amplification Efficiency and Specificity
2.3. RT-qPCR Analysis of Candidate Reference Genes in Garlic
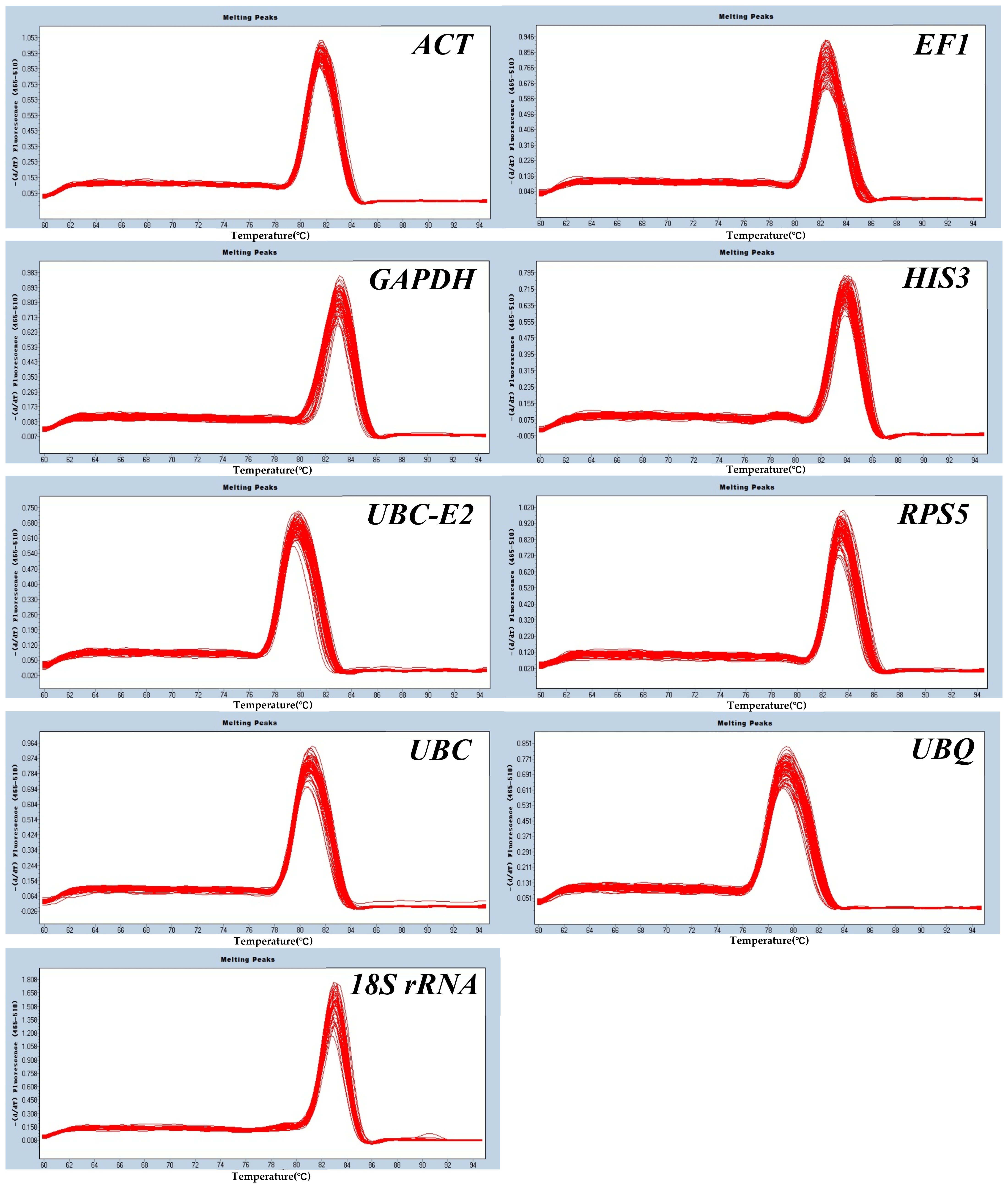
2.3.1. Melting Curve Analysis
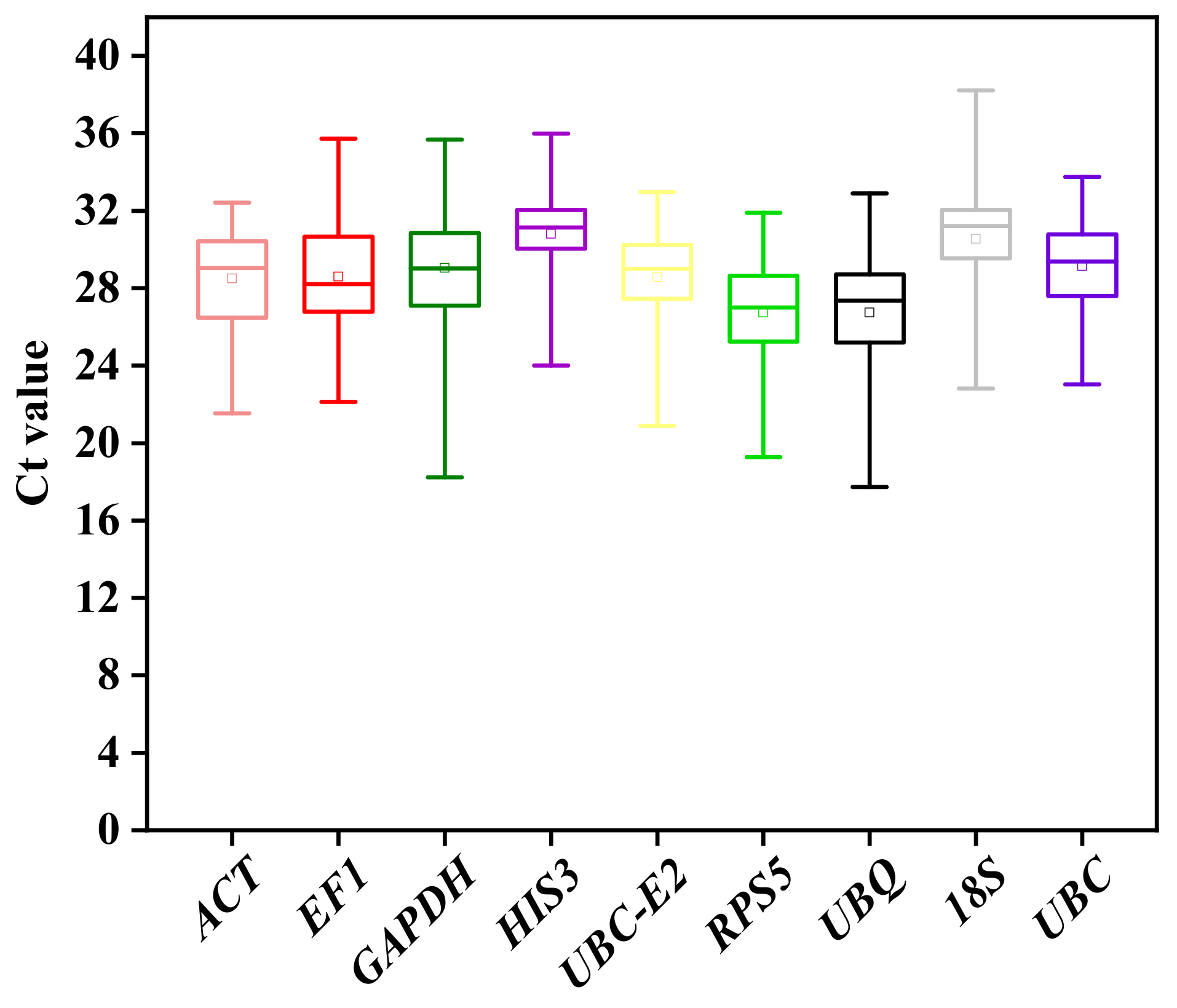
2.3.2. Expression Profile Analysis of Candidate Reference Genes
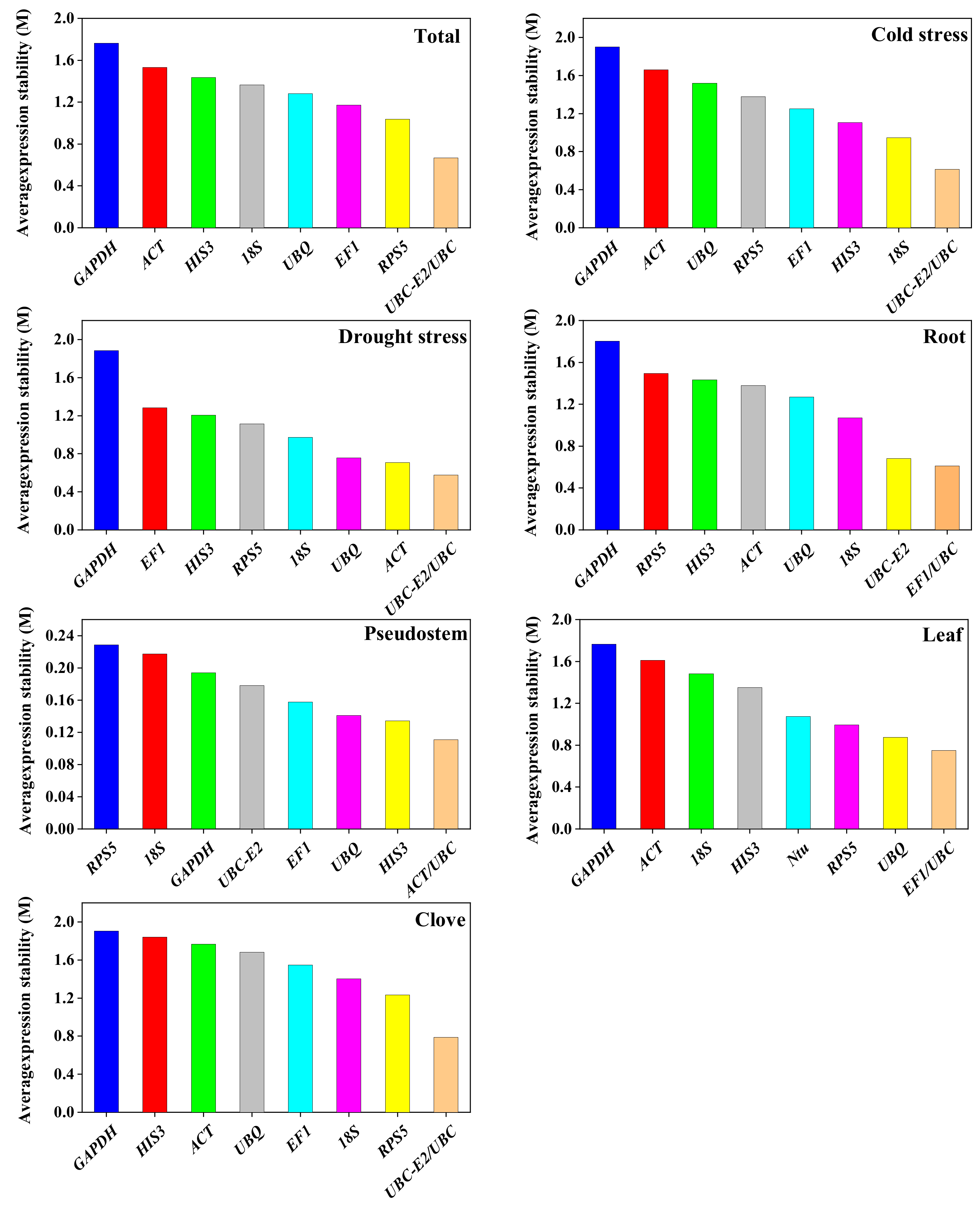
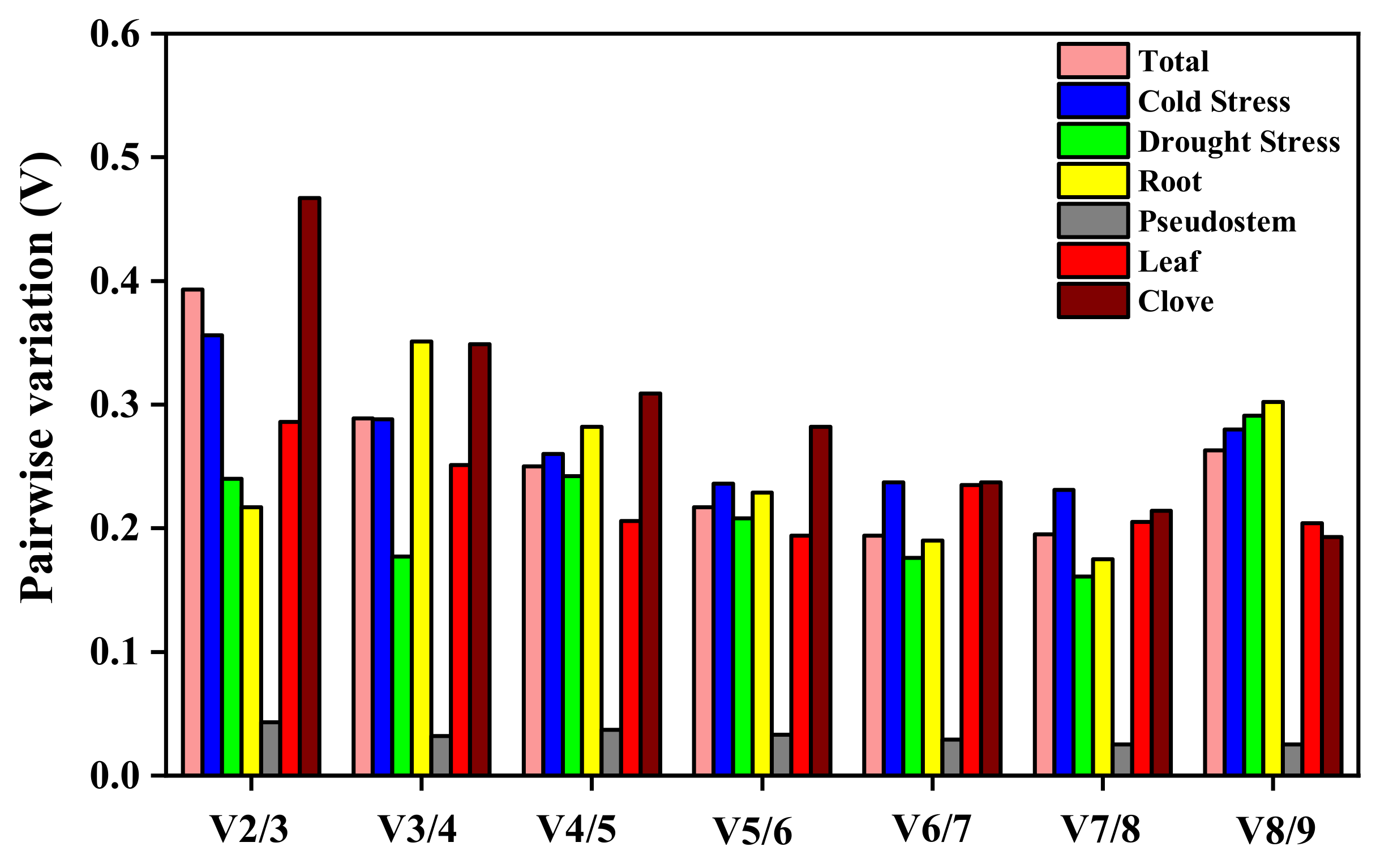
2.4. Expression Stability Analysis of the Reference Genes
2.4.1. geNorm Analysis
2.4.2. NormFinder Analysis
2.4.3. BestKeeper Analysis
2.4.4. ReFinder Analysis
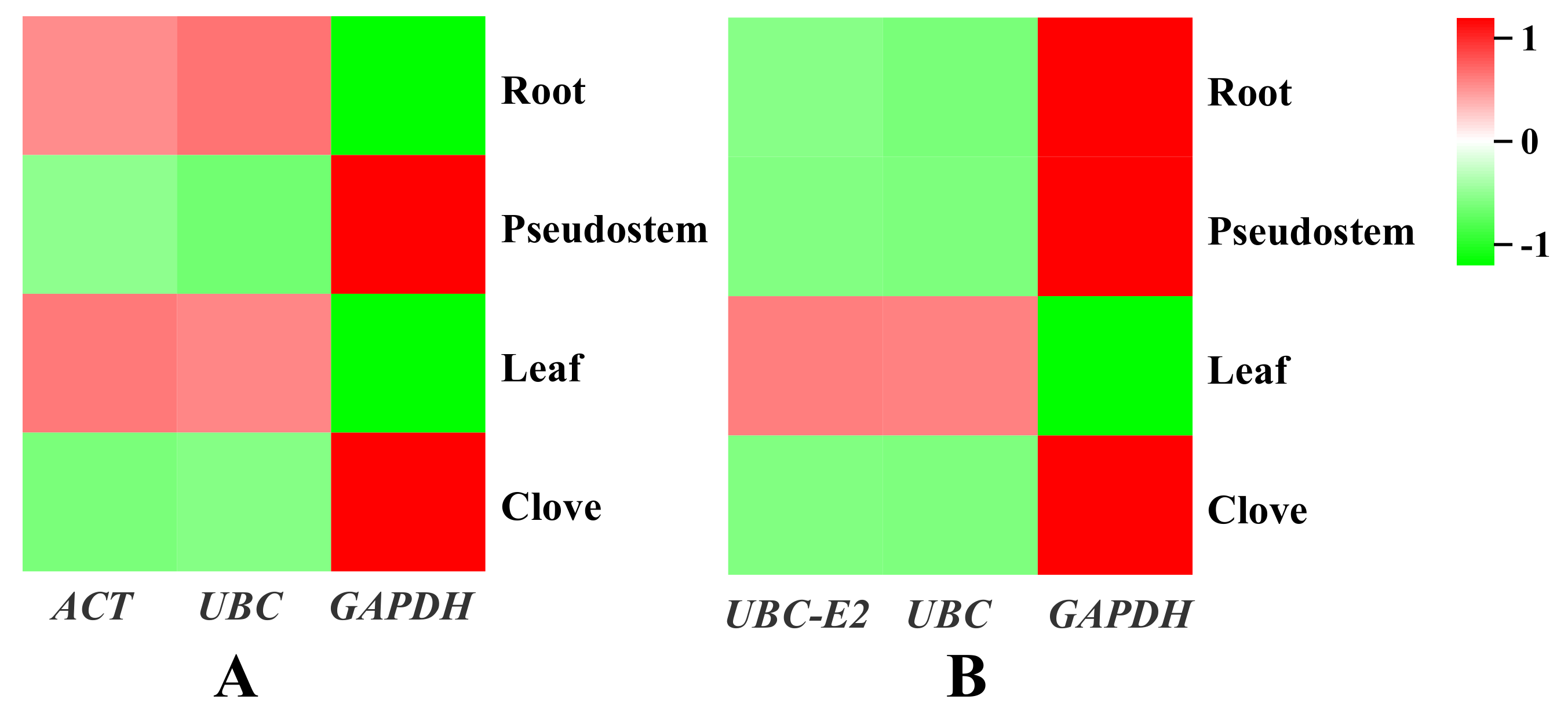
2.5. Stability Verification of CANDIDATE Reference Genes
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material
5.2. Total RNA Isolation and cDNA Synthesis
5.3. Primer Design
5.4. PCR and RT-qPCR Analysis of Reference Genes
5.5. Analysis of Reference Gene Expression Stability
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ried, K.; Frank, O.; Stocks, N. Aged garlic extract reduces blood pressure in hypertensives: A dose–response trial. Eur. J. Clin. Nutr. 2013, 67, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed]
- Farahani, H.A.; Valadabadi, S.A.; Daneshian, J.; Shiranirad, A.H.; Khalvati, M.A. Medicinal and aromatic plants farm-ing under drought conditions. J. Hortic. For. 2009, 1, 086–092. [Google Scholar]
- Ji, L.; Zhou, P.; Zhu, Y.; Liu, F.; Li, R.; Qiu, Y. Proteomic Analysis of Rice Seedlings Under Cold Stress. Protein J. 2017, 36, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.-R.; Liu, B.; Li, W.-Y.; He, Y.-M.; Qi, K.-B.; Wang, H.-B.; Wang, J.-F. Over-expression of a cold-induced plasma membrane protein gene (MpRCI) from plantain enhances low temperature-resistance in transgenic tobacco. Environ. Exp. Bot. 2009, 65, 395–402. [Google Scholar] [CrossRef]
- Khan, A.A.; Vincent, J.F. Mechanical damage induced by controlled freezing in apple and potato. J. Texture Stud. 1996, 27, 143–157. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Ding, H.; Qin, H.; Hou, J.; Huang, X.; Xie, Y.; Guo, T. Physiological Responses and Yield of Wheat Plants in Zinc-Mediated Alleviation of Drought Stress. Front. Plant Sci. 2017, 8, 860. [Google Scholar] [CrossRef]
- Razi, K.; Muneer, S. Drought stress-induced physiological mechanisms, signaling pathways and molecular response of chloroplasts in common vegetable crops. Crit. Rev. Biotechnol. 2021, 41, 669–691. [Google Scholar] [CrossRef]
- Wu, D.; Yang, C.; Zhang, X.; Hou, X.; Zhang, S.; Dai, X.; Zhang, X.; Igarashi, Y.; Luo, F. Algicidal effect of tryptoline against Microcystis aeruginosa: Excess reactive oxygen species production mediated by photosynthesis. Sci. Total. Environ. 2022, 806, 150719. [Google Scholar] [CrossRef]
- Taji, T.; Ohsumi, C.; Iuchi, S.; Seki, M.; Kasuga, M.; Kobayashi, M.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Important roles of drought- and cold-inducible genes for galactinol synthase in stress tolerance in Arabidopsis thaliana. Plant J. 2002, 29, 417–426. [Google Scholar] [CrossRef]
- Bartels, D.; Sunkar, R. Drought and Salt Tolerance in Plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Seki, M.; Umezawa, T.; Urano, K.; Shinozaki, K. Regulatory metabolic networks in drought stress responses. Curr. Opin. Plant Biol. 2007, 10, 296–302. [Google Scholar] [CrossRef]
- Jiang, Y.; Watkins, E.; Liu, S.; Yu, X.; Luo, N. Antioxidative Responses and Candidate Gene Expression in Prairie Junegrass under Drought Stress. J. Am. Soc. Hortic. Sci. 2010, 135, 303–309. [Google Scholar] [CrossRef]
- Lü, X.-P.; Gao, H.-J.; Zhang, L.; Wang, Y.-P.; Shao, K.-Z.; Zhao, Q.; Zhang, J.-L. Dynamic responses of Haloxylon ammodendron to various degrees of simulated drought stress. Plant Physiol. Biochem. 2019, 139, 121–131. [Google Scholar] [CrossRef]
- Huang, L.; Hong, Y.; Zhang, H.; Li, D.; Song, F. Rice NAC transcription factor ONAC095 plays opposite roles in drought and cold stress tolerance. BMC Plant Biol. 2016, 16, 203. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Ahmad, M. Overexpression of AtWRKY30 Transcription Factor Enhances Heat and Drought Stress Tolerance in Wheat (Triticum aestivum L.). Genes 2019, 10, 163. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef]
- Jogawat, A.; Yadav, B.; Chhaya; Lakra, N.; Singh, A.K.; Narayan, O.P. Crosstalk between phytohormones and secondary metabolites in the drought stress tolerance of crop plants: A review. Physiol. Plant. 2021, 172, 1106–1132. [Google Scholar] [CrossRef]
- Kulcheski, F.R.; Marcelino-Guimaraes, F.C.; Nepomuceno, A.L.; Abdelnoor, R.V.; Margis, R. The use of microRNAs as reference genes for quantitative polymerase chain reaction in soybean. Anal. Biochem. 2010, 406, 185–192. [Google Scholar] [CrossRef]
- Wen, L.; Tan, B.; Guo, W.-W. Estimating transgene copy number in precocious trifoliate orange by TaqMan real-time PCR. Plant Cell, Tissue Organ Cult. (PCTOC) 2012, 109, 363–371. [Google Scholar] [CrossRef]
- Zhang, C.; Lei, Z.; Li, Y.; Yi, L.; Shang, Y. Identification of Tps2 Used as an Endogenous Reference Gene in Qualitative and Real-time Quantitative PCR Detection of Flammulina velutipes. Food Anal. Methods 2021, 14, 2152–2160. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.Q.; Fortunato, A.S.; Rodrigues, W.P.; Partelli, F.L.; Campostrini, E.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C.; Ribeiro-Barros, A.I. Selection and Validation of Reference Genes for Accurate RT-qPCR Data Normalization in Coffea spp. under a Climate Changes Context of Interacting Elevated [CO2] and Temperature. Front. Plant Sci. 2017, 8, 307. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.; Hands, R.E.; Bustin, S.A. Quantification of mRNA using real-time RT-PCR. Nat. Protoc. 2006, 1, 1559–1582. [Google Scholar] [CrossRef]
- Libault, M.; Thibivilliers, S.; Bilgin, D.D.; Radwan, O.; Benitez, M.; Clough, S.J.; Stacey, G. Identification of Four Soybean Reference Genes for Gene Expression Normalization. Plant Genome 2008, 1. [Google Scholar] [CrossRef]
- Thomas, C.; Meyer, D.; Wolff, M.; Himber, C.; Alioua, M.; Steinmetz, A. Molecular characterization and spatial expression of the sunflower ABP1 gene. Plant Mol. Biol. 2003, 52, 1025–1036. [Google Scholar] [CrossRef]
- Sikand, K.; Singh, J.; Ebron, J.S.; Shukla, G.C. Housekeeping Gene Selection Advisory: Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) and β-Actin Are Targets of miR-644a. PLoS ONE 2012, 7, e47510. [Google Scholar] [CrossRef]
- Radonić, A.; Thulke, S.; Mackay, I.M.; Landt, O.; Siegert, W.; Nitsche, A. Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2004, 313, 856–862. [Google Scholar] [CrossRef]
- Ding, Y.; Sun, H.; Zhang, R.; Yang, Q.; Liu, Y.; Zang, X.; Zhang, X. Selection of reference gene from Gracilaria lemaneiformis under temperature stress. J. Appl. Phycol. 2015, 27, 1365–1372. [Google Scholar] [CrossRef]
- Yi, S.; Lu, H.; Tian, C.; Xu, T.; Song, C.; Wang, W.; Wei, P.; Gu, F.; Liu, D.; Cai, Y.; et al. Selection of Suitable Reference Genes for Gene Expression Normalization Studies in Dendrobium huoshanense. Genes 2022, 13, 1486. [Google Scholar] [CrossRef]
- Yang, Q.; Yin, J.; Li, G.; Qi, L.; Yang, F.; Wang, R.; Li, G. Reference gene selection for qRT-PCR in Caragana korshinskii Kom. under different stress conditions. Mol. Biol. Rep. 2014, 41, 2325–2334. [Google Scholar] [CrossRef]
- Wang, Q.; Ishikawa, T.; Michiue, T.; Zhu, B.L.; Guan, D.W.; Maeda, H. Stability of endogenous reference genes in postmortem human brains for normalization of quantitative real-time PCR data: Comprehensive evaluation using geNorm, NormFinder, and BestKeeper. Int. J. Leg. Med. 2012, 126, 943–952. [Google Scholar] [CrossRef]
- Zhang, D.; Li, J.; Li, B.; Li, C.; Chen, X.; Ouyang, K. Internal Reference Gene Selection under Different Hormone Stresses in Multipurpose Timber Yielding Tree Neolamarckia cadamba. Forests 2020, 11, 1014. [Google Scholar] [CrossRef]
- Bu, J.; Zhao, J.; Liu, M. Expression Stabilities of Candidate Reference Genes for RT-qPCR in Chinese Jujube (Ziziphus jujuba Mill.) under a Variety of Conditions. PloS ONE 2016, 11, e0154212. [Google Scholar] [CrossRef]
- Liu, M.; Wu, Z.; Jiang, F. Selection and validation of garlic reference genes for quantitative real-time PCR normalization. Plant Cell Tissue Organ Cult. (PCTOC) 2015, 122, 435–444. [Google Scholar] [CrossRef]
- Wang, G.; Tian, C.; Wang, Y.; Wan, F.; Hu, L.; Xiong, A.; Tian, J. Selection of reliable reference genes for quantitative RT-PCR in garlic under salt stress. Peerj 2019, 7, e7319. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Xie, F.; Sun, G.; Stiller, J.; Zhang, B. Genome-Wide Functional Analysis of the Cotton Transcriptome by Creating an Integrated EST Database. PLoS ONE 2011, 6, e26980. [Google Scholar] [CrossRef]
- Jiang, B.; Xie, D.; Liu, W.; Peng, Q.; He, X. De Novo Assembly and Characterization of the Transcriptome, and Development of SSR Markers in Wax Gourd (Benicasa hispida). PLoS ONE 2013, 8, e71054. [Google Scholar] [CrossRef]
- Udvardi, M.K.; Czechowski, T.; Scheible, W.-R. Eleven Golden Rules of Quantitative RT-PCR. Plant Cell 2008, 20, 1736–1737. [Google Scholar] [CrossRef]
- Gutierrez, L.; Mauriat, M.; Guénin, S.; Pelloux, J.; Lefebvre, J.-F.; Louvet, R.; Rusterucci, C.; Moritz, T.; Guerineau, F.; Bellini, C.; et al. The lack of a systematic validation of reference genes: A serious pitfall undervalued in reverse transcription-polymerase chain reaction (RT-PCR) analysis in plants. Plant Biotechnol. J. 2008, 6, 609–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Liu, D.; Li, Y.-C.; Sui, C.; Chen, G.-D.; Tang, Z.-K.; Yang, C.-M.; Hou, D.-B.; Wei, J.-H. Validation of reference genes for expression analysis in three Bupleurum species. Biotechnol. Biotechnol. Equip. 2019, 33, 154–161. [Google Scholar] [CrossRef]
- Huggett, J.F.; Dheda, K.; Bustin, S.; Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun. 2005, 6, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Marum, L.; Miguel, A.; Ricardo, C.P.; Miguel, C. Reference Gene Selection for Quantitative Real-time PCR Normalization in Quercus suber. PLoS ONE 2012, 7, e35113. [Google Scholar] [CrossRef]
- Qi, S.; Yang, L.; Wen, X.; Hong, Y.; Song, X.; Zhang, M.; Dai, S. Reference Gene Selection for RT-qPCR Analysis of Flower Development in Chrysanthemum morifolium and Chrysanthemum lavandulifolium. Front. Plant Sci. 2016, 7, 287. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef]
- Hong, S.-Y.; Seo, P.J.; Yang, M.-S.; Xiang, F.; Park, C.-M. Exploring valid reference genes for gene expression studies in Brachypodium distachyonby real-time PCR. BMC Plant Biol. 2008, 8, 112. [Google Scholar] [CrossRef]
- Wang, G.; Guo, Z.; Wang, X.; Guan, S.L.; Gao, H.; Qi, K.; Gu, C.; Zhang, S. Identification and testing of reference genes for qRT-PCR analysis during pear fruit development. Biologia 2022, 77, 2763–2777. [Google Scholar] [CrossRef]
- Wu, J.; Fan, H.; Hu, Y.; Guo, H.; Lin, H.; Jiao, Y.; Lu, Z.; Du, S.; Liu, X.; Shahid, M.Q. Identification of stable pollen development related reference genes for accurate qRT-PCR analysis and morphological variations in autotetraploid and diploid rice. PLoS ONE 2021, 16, e0253244. [Google Scholar] [CrossRef]
- Shen, C.; Li, J.; Zhang, X.; Wei, C.; Wu, Y. Transcriptome-based identification and validation of optimal reference genes for quantitative real-time PCR normalisation in Psathyrostachys huashanica. Plant Gene 2021, 27, 100306. [Google Scholar] [CrossRef]
- Schmidt, G.W.; Delaney, S.K. Stable internal reference genes for normalization of real-time RT-PCR in tobacco (Nicotiana tabacum) during development and abiotic stress. Mol. Genet. Genom. 2010, 283, 233–241. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Liu, Q.; Tong, H.; Zhang, T.; Gu, C.; Liu, L.; Huang, S.; Yuan, H. Selection and validation of appropriate reference genes for RT-qPCR analysis of flowering stages and different genotypes of Iris germanica L. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Wang, G.-L.; Tian, C.; Jiang, Q.; Xu, Z.-S.; Wang, F.; Xiong, A.-S. Comparison of nine reference genes for real-time quantitative PCR in roots and leaves during five developmental stages in carrot (Daucus carota L.). J. Hortic. Sci. Biotechnol. 2016, 91, 264–270. [Google Scholar] [CrossRef]
- Tang, X.; Wang, H.; Shao, C.; Shao, H. Reference Gene Selection for qPCR Normalization of Kosteletzkya virginica under Salt Stress. BioMed Res. Int. 2015, 2015, 1–8. [Google Scholar] [CrossRef]
- Stürzenbaum, S.R.; Kille, P. Control genes in quantitative molecular biological techniques: The variability of invariance. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol. 2001, 130, 281–289. [Google Scholar] [CrossRef]
- Wang, M.; Wang, Q.; Zhang, B. Evaluation and selection of reliable reference genes for gene expression under abiotic stress in cotton (Gossypium hirsutum L.). Gene 2013, 530, 44–50. [Google Scholar] [CrossRef]
- Jin, Y.; Liu, F.; Huang, W.; Sun, Q.; Huang, X. Identification of reliable reference genes for qRT-PCR in the ephemeral plant Arabidopsis pumila based on full-length transcriptome data. Sci. Rep. 2019, 9, 8408. [Google Scholar] [CrossRef]
- Dudziak, K.; Zapalska, M.; Börner, A.; Szczerba, H.; Kowalczyk, K.; Nowak, M. Analysis of wheat gene expression related to the oxidative stress response and signal transduction under short-term osmotic stress. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Liu, J.; Wang, J.; Lee, S.; Wen, R. Copper-caused oxidative stress triggers the activation of antioxidant enzymes via ZmMPK3 in maize leaves. PLoS ONE 2018, 13, e0203612. [Google Scholar] [CrossRef]
- Gupta, S.; Dong, Y.; Dijkwel, P.P.; Mueller-Roeber, B.; Gechev, T.S. Genome-Wide Analysis of ROS Antioxidant Genes in Resurrection Species Suggest an Involvement of Distinct ROS Detoxification Systems during Desiccation. Int. J. Mol. Sci. 2019, 20, 3101. [Google Scholar] [CrossRef]
- Verma, D.; Upadhyay, S.K.; Singh, K. Characterization of APX and APX-R gene family in Brassica juncea and B. rapa for tolerance against abiotic stresses. Plant Cell Rep. 2021, 41, 571–592. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Yang, H.Y.; Choi, J.-I. Study of functional verification to abiotic stress through antioxidant gene transformation of Pyropia yezoensis (Bangiales, Rhodophyta) APX and MnSOD in Chlamydomonas. J. Microbiol. Biotechnol. 2018, 28, 1217–1224. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Liu, M.; Shi, J.; Zheng, G.; Wang, Y.; Wang, J.; Chen, Y.; Lu, C.; Yin, W. Reference gene selection for qPCR in Ammopiptanthus mongolicus under abiotic stresses and expression analysis of seven ROS-scavenging enzyme genes. Plant Cell Rep. 2012, 31, 1245–1254. [Google Scholar] [CrossRef] [PubMed]
- Wani, M.A.; Jan, N.; Qazi, H.A.; Andrabi, K.I.; John, R. Cold stress induces biochemical changes, fatty acid profile, antioxidant system and gene expression in Capsella bursa pastoris L. Acta Physiol. Plant. 2018, 40, 167. [Google Scholar] [CrossRef]
- Fones, H.; Preston, G.M. Reactive oxygen and oxidative stress tolerance in plant pathogenic Pseudomonas. FEMS Microbiol. Lett. 2012, 327, 1–8. [Google Scholar] [CrossRef]
- Petriccione, M.; Mastrobuoni, F.; Zampella, L.; Scortichini, M. Reference gene selection for normalization of RT-qPCR gene expression data from Actinidia deliciosa leaves infected with Pseudomonas syringae pv. actinidiae. Sci. Rep. 2015, 5, 16961. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper–Excel–based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeep Finder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genename | Primer Sequence (Forward/Reverse) | Amplicon Length (bp) | Tm (℃) | E (%) | Slope | R2 |
|---|---|---|---|---|---|---|
| ACT | ATTAGTGTCGCCATTCTT | 117 | 54 | 91.25413 | −3.551 | 0.999 |
| Actin | TTGACGCACATTACCATC | |||||
| 18S rRNA(18 S) | CGCTGGTGGCGTAGTTGT | 101 | 58 | 109.1765 | −3.12 | 0.996 |
| 18S ribosomal RNA | TGGGAAGGGTGGTTTGTG | |||||
| HIS3 | CCCGTCACAGAGGAAAGA | 121 | 63 | 157.4409 | −2.435 | 0.936 |
| HistoneH3 | GAGCAGCAGGGATAAGCA | |||||
| GAPDH Glyceraldehyde-3-phosphatedehydrogenase | CCCTGGCAAAGGTGAT | 106 | 53 | 96.06391 | −3.42 | 0.999 |
| AAGGCAGTTGGTGGTG | ||||||
| RPS5 | TACCGACCAGAACCCTA | 103 | 53 | 90.62968 | −3.569 | 0.998 |
| RibosomalproteinS5 | CTGCCTGACGCCTAAC | |||||
| UBC-E2 | CGGTTTGTATGAATGTGC | 97 | 54 | 97.83189 | −3.375 | 0.992 |
| Ubiquitin-conjugating enzyme E2 | TTAGGGTAAGAAAGGAGTTG | |||||
| UBC | TTCGGGTTCGGTTTGTAT | 105 | 56 | 83.23965 | −3.802 | 0.996 |
| Ubiquitin-conjugating enzyme | TTAGGGTAAGAAAGGAGTTGAG | |||||
| UBQ | GGAAGATGGCAGAACG | 139 | 50 | 94.88171 | −3.451 | 0.999 |
| Polyubiquitin | GCACAAGATGAAGGGTA | |||||
| EF1 | GCATAAAGAAGGAGGGT | 147 | 56 | 85.79506 | −3.717 | 0.992 |
| elongationfactor1 | CTGGTTCGGCAGTAAG |
| Rank | Total | Cold Stress | Drought Stress | Root | Pseudostem | Leaf | Clove | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | |
| 1 | UBC-E2 | 0.035 | HIS3 | 0.48 | EF1 | 0.566 | ACT | 0.582 | UBQ | 0.063 | UBC | 0.482 | UBC-E2 | 0.7 |
| 2 | EF1 | 0.041 | UBC-E2 | 0.633 | ACT | 0.579 | 18S rRNA | 0.645 | HIS3 | 0.072 | EF1 | 0.527 | UBC | 0.722 |
| 3 | HIS3 | 0.042 | UBC | 0.692 | UBC | 0.607 | EF1 | 0.72 | ACT | 0.072 | UBC-E2 | 0.564 | RPS5 | 0.73 |
| 4 | UBQ | 0.046 | EF1 | 0.832 | UBC-E2 | 0.65 | HIS3 | 0.744 | UBC-E2 | 0.073 | UBQ | 0.637 | EF1 | 0.772 |
| 5 | 18S rRNA | 0.047 | UBQ | 0.901 | RPS5 | 0.657 | UBC-E2 | 0.802 | UBC | 0.083 | RPS5 | 0.657 | UBQ | 0.837 |
| 6 | ACT | 0.05 | ACT | 0.923 | UBQ | 0.658 | UBC | 0.81 | GAPDH | 0.085 | 18S rRNA | 0.784 | 18S rRNA | 0.857 |
| 7 | RPS5 | 0.051 | 18S rRNA | 1.03 | HIS3 | 0.689 | UBQ | 0.923 | EF1 | 0.096 | HIS3 | 0.882 | HIS3 | 0.985 |
| 8 | UBC | 0.062 | RPS5 | 1.132 | 18S rRNA | 0.772 | RPS5 | 1.052 | 18S rRNA | 0.108 | GAPDH | 0.943 | GAPDH | 1.006 |
| 9 | GAPDH | 0.069 | GAPDH | 1.341 | GAPDH | 1.525 | GAPDH | 1.589 | RPS5 | 0.113 | ACT | 1.09 | ACT | 1.009 |
| Rank | Total | Cold Stress | Drought Stress | Root | Pseudostem | Leaf | Clove | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | Gene | SD | CV | |
| 1 | HIS3 | 0.773 | 2.451 | UBC-E2 | 0.787 | 2.751 | HIS3 | 0.711 | 2.271 | ACT | 0.309 | 1.07 | 18S rRNA | 0.367 | 1.142 | 18S rRNA | 0.344 | 1.073 | UBC-E2 | 0.742 | 2.45 |
| 2 | 18S rRNA | 0.845 | 2.676 | HIS3 | 0.927 | 2.921 | RPS5 | 0.752 | 2.824 | HIS3 | 0.744 | 2.375 | HIS3 | 0.554 | 1.762 | GAPDH | 0.617 | 2.273 | 18S rRNA | 0.784 | 2.478 |
| 3 | UBC-E2 | 1.112 | 3.775 | UBC | 0.933 | 3.23 | 18S rRNA | 0.801 | 2.564 | 18S rRNA | 0.83 | 2.721 | UBC-E2 | 0.621 | 2.137 | HIS3 | 0.625 | 1.982 | UBC | 0.846 | 2.761 |
| 4 | UBC | 1.26 | 4.218 | UBQ | 0.945 | 3.524 | EF1 | 0.858 | 3.047 | UBQ | 1.373 | 5.065 | GAPDH | 0.659 | 2.39 | RPS5 | 0.956 | 3.389 | UBQ | 1.17 | 4.149 |
| 5 | RPS5 | 1.284 | 4.648 | 18S rRNA | 0.97 | 3.081 | ACT | 0.885 | 2.944 | GAPDH | 1.525 | 5.046 | UBC | 0.961 | 3.218 | UBC-E2 | 1.079 | 3.623 | HIS3 | 1.267 | 3.98 |
| 6 | UBQ | 1.321 | 4.747 | RPS5 | 1.329 | 4.887 | UBC-E2 | 0.994 | 3.419 | RPS5 | 1.531 | 5.717 | RPS5 | 1.274 | 4.647 | UBC | 1.12 | 3.726 | RPS5 | 1.311 | 4.672 |
| 7 | ACT | 1.484 | 5.066 | EF1 | 1.513 | 5.357 | UBC | 1.023 | 3.494 | EF1 | 1.588 | 5.592 | UBQ | 1.295 | 4.564 | EF1 | 1.287 | 4.385 | GAPDH | 1.391 | 4.584 |
| 8 | GAPDH | 1.67 | 5.794 | ACT | 1.677 | 6.149 | UBQ | 1.145 | 4.227 | UBC-E2 | 1.609 | 5.6 | ACT | 2.037 | 6.956 | UBQ | 1.413 | 5.113 | ACT | 1.397 | 4.719 |
| 9 | EF1 | 1.712 | 5.84 | GAPDH | 1.883 | 6.604 | GAPDH | 2.046 | 7.101 | UBC | 1.723 | 5.968 | EF1 | 2.042 | 7.092 | ACT | 1.951 | 6.66 | EF1 | 1.596 | 5.195 |
| Rank | Total | Cold Stress | Drought Stress | Root | Pseudostem | Leaf | Clove | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genes | Rank | Genes | Rank | Genes | Rank | Genes | Rank | Genes | Rank | Genes | Rank | Genes | Rank | |
| 1 | UBC | 1.41 | UBC | 1.68 | ACT | 1.57 | EF1 | 1.97 | HIS3 | 1.41 | UBC | 1.50 | UBC-E2 | 1.00 |
| 2 | UBC-E2 | 1.86 | UBC-E2 | 1.73 | UBC | 2.30 | ACT | 2.34 | UBC | 1.86 | EF1 | 2.30 | UBC | 1.68 |
| 3 | RPS5 | 3.94 | HIS3 | 1.86 | UBC-E2 | 2.71 | 18S rRNA | 3.13 | UBC-E2 | 2.06 | UBQ | 3.83 | RPS5 | 3.71 |
| 4 | HIS3 | 3.96 | 18S rRNA | 4.36 | HIS3 | 4.30 | UBC | 3.20 | GAPDH | 4.23 | RPS5 | 4.00 | 18S rRNA | 4.16 |
| 5 | UBQ | 4.05 | EF1 | 4.95 | 18S rRNA | 4.61 | UBC-E2 | 3.46 | 18S rRNA | 4.82 | HIS3 | 4.30. | EF1 | 5.03 |
| 6 | 18S rRNA | 4.36 | UBQ | 5.38 | UBQ | 4.76 | HIS3 | 5.12 | UBQ | 5.48 | GAPDH | 4.56 | UBQ | 5.96 |
| 7 | EF1 | 6.05 | RPS5 | 6.96 | RPS5 | 5.18 | UBQ | 5.18 | ACT | 7.24 | UBC-E2 | 5.23 | HIS3 | 6.05 |
| 8 | ACT | 7.74 | ACT | 7.74 | EF1 | 7.11 | RPS5 | 8.00 | EF1 | 8.24 | 18S rRNA | 6.26 | ACT | 7.20 |
| 9 | GAPDH | 9.00 | GAPDH | 9.00 | GAPDH | 9.00 | GAPDH | 9.00 | RPS5 | 8.45 | ACT | 9.00 | GAPDH | 9.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Guo, C.; Yang, S.; Zhong, Q.; Tian, J. Screening and Verification of Reference Genes for Analysis of Gene Expression in Garlic (Allium sativum L.) under Cold and Drought Stress. Plants 2023, 12, 763. https://doi.org/10.3390/plants12040763
Wang Q, Guo C, Yang S, Zhong Q, Tian J. Screening and Verification of Reference Genes for Analysis of Gene Expression in Garlic (Allium sativum L.) under Cold and Drought Stress. Plants. 2023; 12(4):763. https://doi.org/10.3390/plants12040763
Chicago/Turabian StyleWang, Qizhang, Chunqian Guo, Shipeng Yang, Qiwen Zhong, and Jie Tian. 2023. "Screening and Verification of Reference Genes for Analysis of Gene Expression in Garlic (Allium sativum L.) under Cold and Drought Stress" Plants 12, no. 4: 763. https://doi.org/10.3390/plants12040763
APA StyleWang, Q., Guo, C., Yang, S., Zhong, Q., & Tian, J. (2023). Screening and Verification of Reference Genes for Analysis of Gene Expression in Garlic (Allium sativum L.) under Cold and Drought Stress. Plants, 12(4), 763. https://doi.org/10.3390/plants12040763