
Cd Phytoextraction Potential in Halophyte Salicornia fruticosa: Salinity Impact

 ,
,  and
and
Abstract
:1. Introduction
2. Results
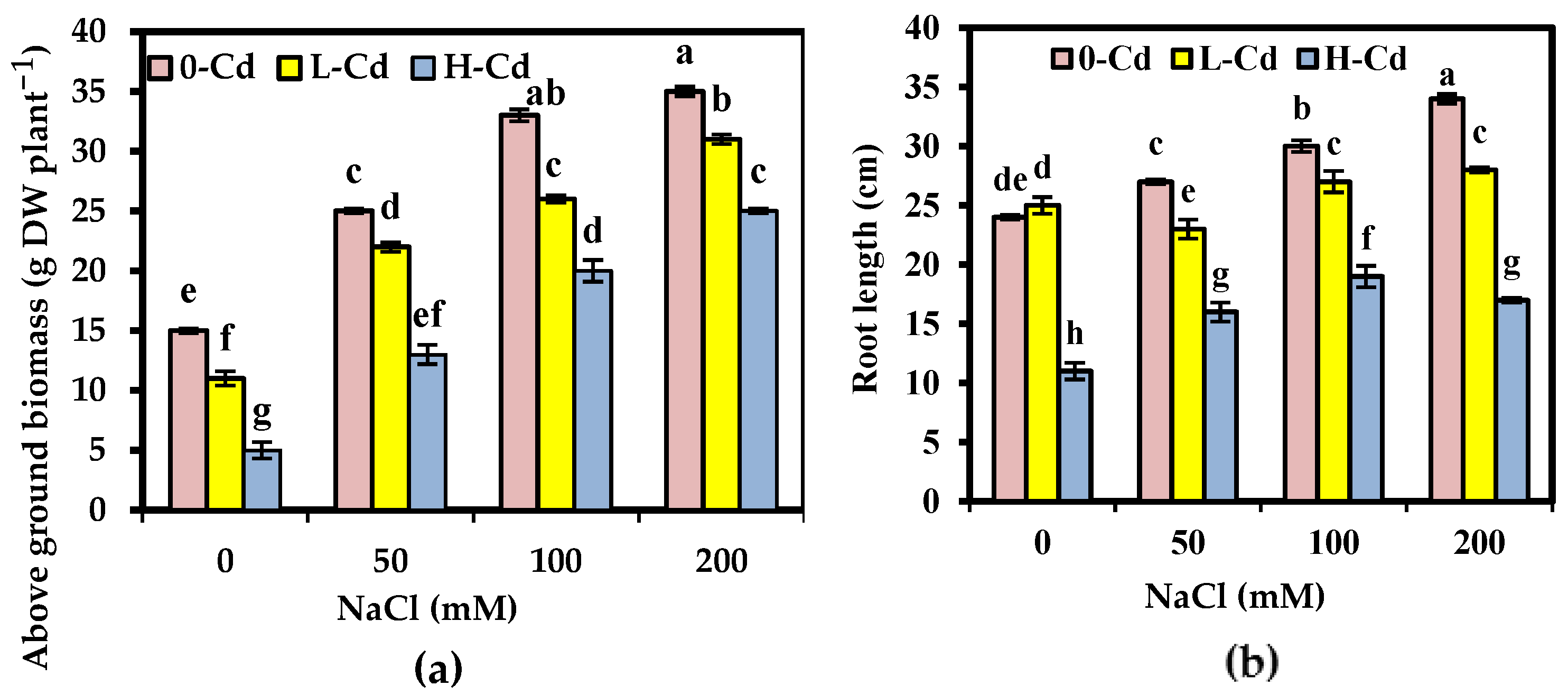
2.1. Phenotypic Criteria Affected by Interaction between Cd Stress and Salinity in S. fruticosa
2.2. Water Relation Indices Affected by Cd Stress and Salinity Co-Occurrence in S. fruticosa
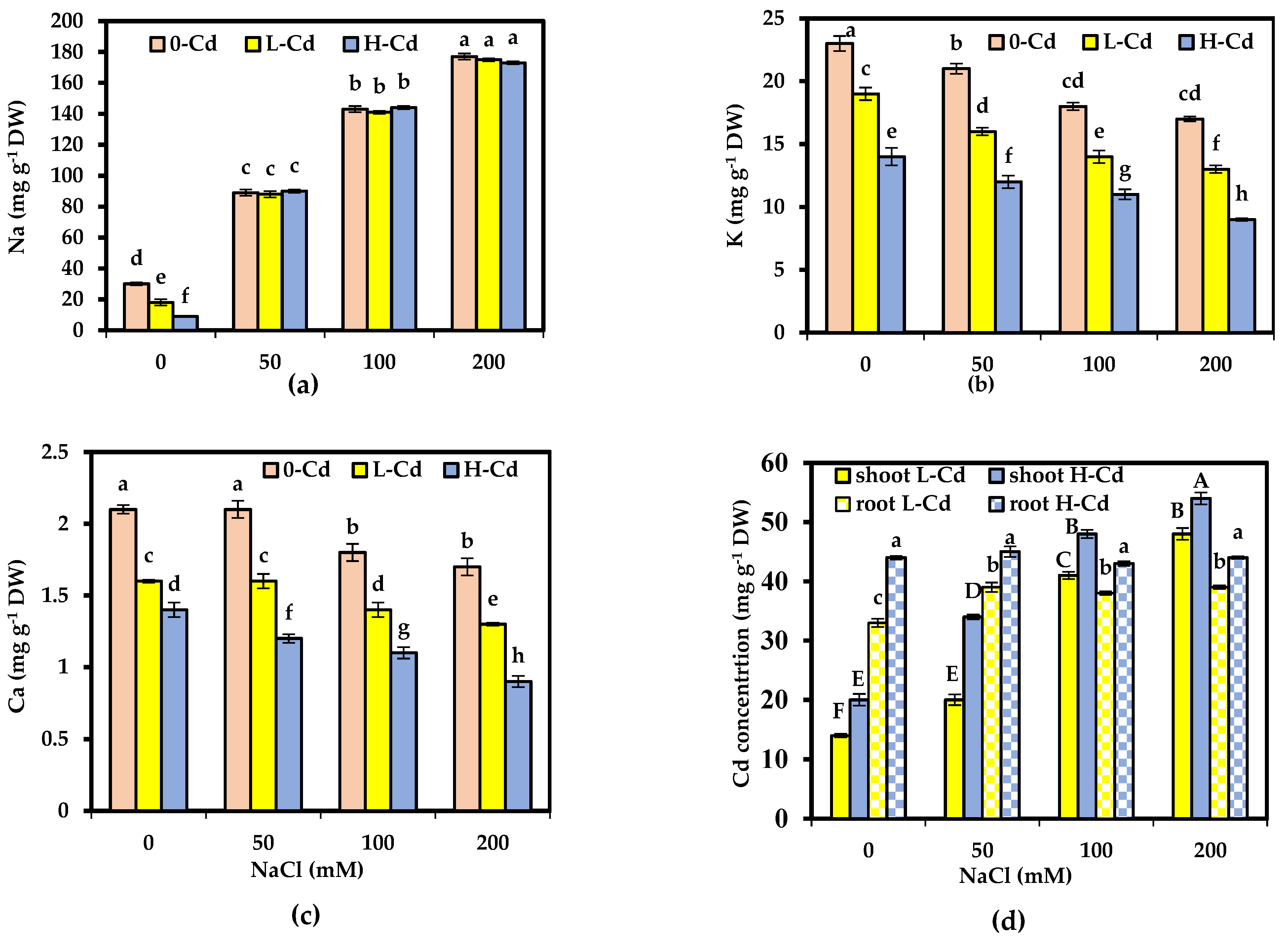
2.3. Mineral Composition Affected by Cd Stress and Salinity Co-Occurrence in S. fruticosa
2.4. Phytoremediation Parameters Affected by Cd Stress and Salinity Co-Occurrence in S. fruticosa
2.5. Non-Enzymatic Antioxidant Indices as Affected by Cd Stress and Salinity Co-Occurrence in S. fruticosa
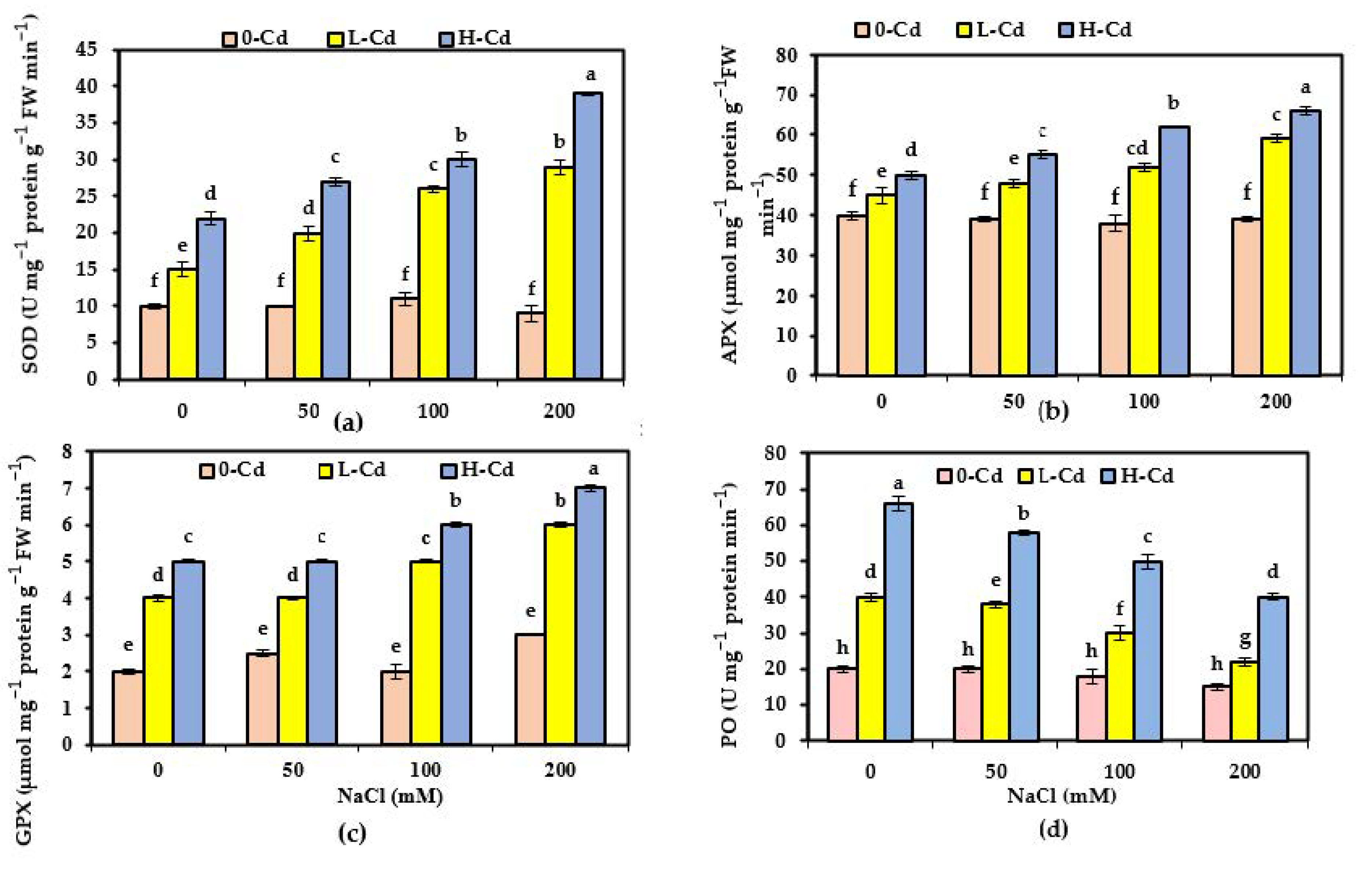
2.6. Alternations in the Capacities of Enzymatic Antioxidant of S. fruticosa as Affected by Cd Stress and Salinity Co-Occurrence
3. Discussion
3.1. Phenotypic Criteria Affected by Interaction between Cd Stress and Salinity in S. fruticosa
3.2. Water Relation Indices Affected by Cd Stress and Salinity Co-Occurrence in S. fruticosa
3.3. Mineral Composition Affected by Cd Stress and Salinity Co-Occurrence in S. fruticosa
3.4. Phytoremediation Parameters Affected by Cd Stress and Salinity Co-Occurrence in S. fruticosa
3.5. Non-Enzymatic Antioxidant Indices as Affected by Cd Stress and Salinity Co-Occurrence in S. fruticosa
3.6. Enzymatic Antioxidant Capacities as Affected by Cd Stress and Salinity Co-Occurrence in S. fruticosa
4. Materials and Methods
4.1. Plant Material Collection
4.2. Hydroponic Culturing
4.3. Phenotypic Criteria
4.4. Shoot Succulence Degree (SSD)
4.5. Transpiration Rate
4.6. Total Osmotic Potential (TOP) Determination
4.7. Proline Determination
4.8. Enzymatic and Non-Enzymatic Antioxidant Capacities
4.9. Cation Assay
4.10. Cd Accumulation Characteristics
- Bioconcentration factor (BCF) and enrichment factor = Cd concentration in the plant/Cd concentration in external medium;
- Translocation factor (TF) = Cd concentration in the shoot/Cd concentration in the root;
- Cd absorption efficiency (AE) = Cd accumulation in the whole plant/root biomass.
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60. [Google Scholar] [CrossRef] [PubMed]
- Eissa, M.A.; Abeed, A.H. Growth and biochemical changes in quail bush (Atriplex lentiformis (Torr.) S. Wats) under Cd stress. Environ. Sci. Pollut. Res. 2019, 26, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Abeed, A.H.A.; Salama, F.M. Attenuating Effect of an Extract of Cd-Hyperaccumulator Solanum nigrum on the Growth and Physio-chemical Changes of Datura innoxia Under Cd Stress. J. Soil Sci. Plant Nutr. 2022, 22, 1–15. [Google Scholar] [CrossRef]
- Ullah, A.; Heng, S.; Munis, M.F.H.; Fahad, S.; Yang, X. Phytoremediation of heavy metals assisted by plant growth promoting (PGP) bacteria: A review. Environ. Exp. Bot. 2015, 117, 28–40. [Google Scholar] [CrossRef]
- Younis, A.M.; Ismail, I.S.; Mohamedein, L.I.; Ahmed, S.F. spatial variation and environmental risk assessment of heavy metal in the surficial sediments along the Egyptian Red Sea coast. Catrina. Int. J. Environ. Sci. 2015, 10, 45–52. [Google Scholar] [CrossRef]
- Pourret, O.; Hursthouse, A. It’s Time to Replace the Term “Heavy Metals” with “Potentially Toxic Elements” When Reporting Environmental Research. Int. J. Environ. Res. Public Health 2019, 16, 4446. [Google Scholar] [CrossRef]
- Fifield, F.W.; Haines, P.J. Environmental Analytical Chemistry; Wiley-Blackwell: Hoboken, NJ, USA, 2000. [Google Scholar]
- El-Sorogy, A.S.; Tawfik, M.; Almadani, S.A.; Attiah, A. Assessment of toxic metals in coastal sediments of the Rosetta area, Mediterranean Sea, Egypt. Environ. Earth Sci. 2016, 75, 1–11. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency (USEPA). Quality Criteria for Water; EPA 440/5-86-001; Office of Water Regulations and Standards: Washington, DC, USA, 1986.
- Khallaf, E.A.; Galal, M.; Authman, M. Assessment of heavy metals pollution and their effect on Oreochromis niloticus in aquatic drainage canals. J. Egypt. Ger. Soc. Zoo. 1998, 26, 39–74. [Google Scholar]
- Lefèvre, I.; Marchal, G.; Meerts, P.; Corréal, E.; Lutts, S. Chloride salinity reduces cadmium accumulation by the Mediterranean halophyte species Atriplex halimus L. Environ. Exp. Bot. 2009, 65, 142–152. [Google Scholar] [CrossRef]
- Kumar, V.; Shahi, S.K.; Singh, S. Bioremediation: An eco-sustainable approach for restoration of contaminated sites. In Microbial Bioprospecting for Sustainable Development; Springer: Singapore, 2018; pp. 115–136. [Google Scholar] [CrossRef]
- Marques, A.P.G.C.; Rangel, A.O.S.S.; Castro, P.M.L. Remediation of heavy metal contaminated soils: Phytoremediation as a potentially promising clean-up technology. Crit. Rev. Environ. Sci. Technol. 2009, 39, 622–654. [Google Scholar] [CrossRef]
- Sharma, R.; Wungrampha, S.; Singh, V.; Pareek, A.; Sharma, M.K. Halophytes as bioenergy crops. Front. Plant Sci. 2016, 7, 1372. [Google Scholar] [CrossRef] [PubMed]
- Ghnaya, T.; Slama, I.; Messedi, D.; Grignon, C.; Ghorbel, M.H.; Abdelly, C. Cd-induced growth reduction in the halophyte Sesuvium portulacastrum is significantly improved by NaCl. J. Plant Res. 2007, 120, 309–316. [Google Scholar] [CrossRef]
- Christofilopoulos, S.; Syranidou, E.; Gkavrou, G.; Manousaki, E.; Kalogerakis, N. The role of halophyte Juncus acutus L. in the remediation of mixed contamination in a hydroponic greenhouse experiment. J. Chem. Technol. Biotechnol. 2016, 91, 1665–1674. [Google Scholar] [CrossRef]
- Rahman, M.M.; Mostofa, M.G.; Keya, S.S.; Siddiqui, M.N.; Ansary, M.M.U.; Das, A.K.; Rahman, M.A.; Tran, L.S.-P. Adaptive mechanisms of halophytes and their potential in improving salinity tolerance in plants. Int. J. Mol. Sci. 2021, 22, 10733. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Guo, J.; Shabala, S.; Wang, B. Reproductive physiology of halophytes: Current standing. Front. Plant Sci. 2019, 9, 1954. [Google Scholar] [CrossRef]
- Smýkalová, I.; Zámečníková, B. The relationship between salinity and cadmium stress in barley. Biol. Plant. 2003, 46, 269–273. [Google Scholar] [CrossRef]
- Sepehr, M.F.; Ghorbanli, M.; Khavari Nejad, R.A. Effects of cadmium and salinity on growth, photosynthesis and ionic contents of Zea mays. Asian J. Plant Sci. 2003, 2, 196–201. [Google Scholar] [CrossRef]
- Patel, M.; Parida, A.K. Salinity alleviates the arsenic toxicity in the facultative halophyte Salvadora persica L. by the modulations of physiological, biochemical, and ROS scavenging attributes. J. Hazard. Mater. 2021, 401, 123368. [Google Scholar] [CrossRef]
- Borin, M.; Malagoli, M. Ecology, functioning and management of wetland systems. Environ. Sci. Pollut. Res. 2015, 22, 2357–2359. [Google Scholar] [CrossRef]
- Marco, P.; Carvajal, M.; del Carmen Martinez-Ballesta, M. Efficient leaf solute partioning in Salicornia fruticosa allows growth under salinity. Environ. Exp. Bot. 2019, 157, 177–186. [Google Scholar] [CrossRef]
- Han, R.-M.; Lefèvre, I.; Albacete, A.; Pérez-Alfocéa, F.; Barba-Espin, G.; Diaz-Vivancos, P.; Quinet, M.; Ruan, C.-J.; Hernandez, J.-A.; Cantero-Nazvarro, E.; et al. Antioxidant enzyme activities and hormonal status in response to Cd stress in the wetland halophyte Kosteletzkya virginica under saline conditions. Physiol. Plant. 2013, 147, 352–368. [Google Scholar] [CrossRef]
- Zhang, S.; Ni, X.; Arif, M.; Yuan, Z.; Li, L.; Li, C. Salinity influences Cd accumulation and distribution characteristics in two contrasting halophytes, Suaeda glauca and Limonium aureum. Ecotoxicol. Environ. Saf. 2020, 191, 110230. [Google Scholar] [CrossRef] [PubMed]
- Sruthi, P.; Shackira, A.M.; Puthur, J.T. Heavy metal detoxification mechanisms in halophytes: An overview. Wetl. Ecol. Manag. 2017, 25, 129–148. [Google Scholar] [CrossRef]
- Lim, S.D.; Mayer, J.A.; Yim, W.C.; Cushman, J.C. Plant tissue succulence engineering improves water-use efficiency, water-deficit stress attenuation and salinity tolerance in Arabidopsis. Plant J. 2020, 103, 1049–1072. [Google Scholar] [CrossRef] [PubMed]
- Hamed, K.B.; Ellouzi, H.; Talbi, O.Z.; Hessini, K.; Slama, I.; Ghnaya, T.; Bosch, S.M.; Savouré, A.; Abdelly, C. Physiological response of halophytes to multiple stresses. Funct. Plant Biol. 2013, 40, 883–896. [Google Scholar] [CrossRef]
- Benzarti, M.; Rejeb, K.B.; Messedi, D.; Mna, A.B.; Hessini, K.; Ksontini, M.; Abdelly, C.; Debez, A. Effect of high salinity on Atriplex portulacoides: Growth, leaf water relations and solute accumulation in relation with osmotic adjustment. S. Afr. J. Bot. 2014, 95, 70–77. [Google Scholar] [CrossRef]
- Guo, Q.; Meng, L.; Zhang, Y.N.; Mao, P.C.; Tian, X.X.; Li, S.S.; Zhang, L. Antioxidative systems, metal ion homeostasis and Cd distribution in Iris lactea exposed to Cd stress. Ecotoxicol. Environ. Saf. 2017, 139, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Wiszniewska, A.; Koźmińska, A.; Hanus-Fajerska, E.; Dziurka, M.; Dziurka, K. Insight into mechanisms of multiple stresses tolerance in a halophyte Aster tripolium subjected to salinity and heavy metal stress. Ecotoxicol. Environ. Saf. 2019, 180, 12–22. [Google Scholar] [CrossRef]
- Samiei, L.; Pahnehkolayi, M.D.; Karimian, Z.; Nabati, J. Morpho-Physiological Responses of Halophyte Climacoptera crassa to Salinity and Heavy Metal Stresses in In Vitro Condition. S. Afr. J. Bot. 2020, 131, 468–474. [Google Scholar] [CrossRef]
- Parida, A.K.; Jha, B. Antioxidative defense potential to salinity in the euhalophyte Salicornia brachiata. J. Plant Growth Regul. 2010, 29, 137–148. [Google Scholar] [CrossRef]
- Vaněk, T.; Moťková, K.; Podlipná, R. Accumulation of cadmium by halophytic and non-halophytic Juncus species. Theor. Exp. Plant Physiol. 2016, 28, 415–423. [Google Scholar] [CrossRef]
- Wali, M.; Gunsè, B.; Llugany, M.; Corrales, I.; Abdelly, C.; Poschenrieder, C.; Ghnaya, T. High salinity helps the halophyte Sesuvium portulacastrum in defense against Cd toxicity by maintaining redox balance and photosynthesis. Planta 2016, 244, 333–346. [Google Scholar] [CrossRef]
- Clemens, S. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochimie 2006, 88, 1707–1719. [Google Scholar] [CrossRef] [PubMed]
- Lutts, S.; Lefèvre, I. How can we take advantage of halophyte properties to cope with heavy metal toxicity in salt-affected areas? Ann. Bot. 2015, 115, 509–528. [Google Scholar] [CrossRef] [PubMed]
- Wali, M.; Fourati, E.; Hmaeid, N.; Ghabriche, R.; Poschenrieder, C.; Abdelly, C.; Ghnaya, T. NaCl alleviates Cd toxicity by changing its chemical forms of accumulation in the halophyte Sesuvium portulacastrum. Environ. Sci. Pollut. Res. 2015, 22, 10769–10777. [Google Scholar] [CrossRef]
- Ozgur, R.; Uzilday, B.; Sekmen, A.H.; Turkan, I. Reactive oxygen species regulation and antioxidant defence in halophytes. Funct. Plant Biol. 2013, 40, 832–847. [Google Scholar] [CrossRef]
- Zhou, M.; Han, R.; Ghnaya, T.; Lutts, S. Salinity influences the interactive effects of cadmium and zinc on ethylene and polyamine synthesis in the halophyte plant species Kosteletzkya pentacarpos. Chemosphere 2018, 209, 892–900. [Google Scholar] [CrossRef]
- Nimptsch, J.; Wunderlin, D.A.; Dollan, A.; Pflugmacher, S. Antioxidant and biotransformation enzymes in Myriophyllum quitense as biomarkers of heavy metal exposure and eutrophication in Suquía River basin (Cordoba, Argentina). Chemosphere 2005, 61, 147–157. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water culture method for growing plant without soil. Calif. Agric. Exp. Stn. Cir. 1950, 347, 32. [Google Scholar]
- Qi, C.H.; Chen, M.; Song, J.; Wang, B.S. Increase in aquaporin activity is involved in leaf succulence of the euhalophyte Suaeda salsa, under salinity. Plant Sci. 2009, 176, 200–205. [Google Scholar] [CrossRef]
- Llanes, A.; Pinamonti, P.; Iparraguirre, J.; Bertazza, G.; Luna, V. Abscisic acid alters carbohydrate accumulation induced by differential response to sodium salts in the halophyte Prosopis strombulifera. Plant Biosyst. 2020, 154, 337–347. [Google Scholar] [CrossRef]
- Abeed, A.; Dawood, M. Comparative impact of different isoosmotic solutions on osmotic adjustment in Gossypium barbadense. J. Glob. Nest 2020, 22, 75–84. [Google Scholar] [CrossRef]
- Walter, H. Hydratur der Pflanze und ihre physiologischökologische Bedeutung. In Untersuchungen uX ber den Osmotischen Wert; Fischer, G., Ed.; Springer: Jena, Germany, 1931. [Google Scholar]
- Bates, L.S.; Walds, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Jagota, S.K.; Dani, H.M. Anew colorimetric technique for the estimation of vitamin C using Folin phenol reagent. Anal. Biochem. 1982, 127, 178–182. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Suzuki, T.; Fujita, M. Polyamine and nitric oxide crosstalk: Antagonistic effects on cadmium toxicity in mung bean plants through upregulating the metal detoxification, antioxidant defense and methylglyoxal detoxification systems. Ecotoxicol. Environ. Saf. 2016, 126, 245–255. [Google Scholar] [CrossRef]
- Flohé, L.; Günzler, W.A. Methods in Enzymology. In Assays of Glutathione Peroxidase; Packer, L., Ed.; Academic Press: New York, NY, USA, 1984; pp. 114–121. [Google Scholar] [CrossRef]
- Abeed, A.H.A.; Ali, M.; Ali, E.F.; Majrashi, A.; Eissa, M.A. Induction of Catharanthus roseus Secondary Metabolites When Calotropis procera Was Used as Bio-Stimulant. Plants 2021, 10, 1623. [Google Scholar] [CrossRef]
- Abeed, A.H.; Eissa, M.A.; Abdel-Wahab, D.A. Effect of Exogenously Applied Jasmonic Acid and Kinetin on Drought Tolerance of Wheat Cultivars Based on Morpho-Physiological Evaluation. J. Soil Sci. Plant Nutr. 2020, 21, 131–144. [Google Scholar] [CrossRef]
- Ghanati, F.; Morita, A.; Yokota, H. Induction of suberin and increase of lignin content by excess boron in Tabacco cells. Soil Sci. Plant Nut. 2002, 48, 357–364. [Google Scholar] [CrossRef]
- Li, J.; Chang, Y.; Al-Huqail, A.A.; Ding, Z.; Al-Harbi, M.S.; Ali, E.F.; Abeed, A.H.A.; Rekaby, S.A.; Eissa, M.A.; Ghoneim, A.M.; et al. Effect of manure and compost on the phytostabilization potential of heavy metals by the halophytic plant wavy-leaved saltbush. Plants 2021, 10, 2176. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, B.; Xia, H. Effect of cadmium on growth, photosynthesis, mineral nutrition and metal accumulation of bana grass and vetiver grass. Ecotoxicol. Environ. Saf. 2014, 106, 102–108. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Treatments | Transpiration Rate (mL Transpired H2O g−1 FW Day−1) | Shoot Succulence Degree (g g−1) | TOP (MPa) | |
|---|---|---|---|---|
| 0 mM NaCl | 0-Cd | 30 ± 0.50 a | 5.02 ± 0.04 a | −24.7 ± 0.32 f |
| L-Cd | 22 ± 0.40 b | 5.00 ± 0.05 a | −25.5 ± 0.22 e | |
| H-Cd | 6 ± 0.08 d | 3.45 ± 0.06 c | −28.3 ± 0.15 a | |
| 50 mM NaCl | 0-Cd | 28 ± 0.60 a | 5.41 ± 0.04 a | −24.7 ± 0.25 f |
| L-Cd | 20 ± 0.90 b | 5.11 ± 0.03 a | −25.1 ± 0.13 e | |
| H-Cd | 11 ± 0.40 c | 3.99 ± 0.05 b | −27.8 ± 0.14 b | |
| 100 mM NaCl | 0-Cd | 30 ± 0.90 a | 5.33 ± 0.04 a | −24.9 ± 0.44 f |
| L-Cd | 21 ± 0.60 b | 4.99 ± 0.03 a | −24.8 ± 0.24 f | |
| H-Cd | 19 ± 0.70 b | 4.11 ± 0.03 b | −26.4 ± 0.13 c | |
| 200 mM NaCl | 0-Cd | 29 ± 0.70 a | 5.34 ± 0.02 a | −24.8 ± 0.12 f |
| L-Cd | 20 ± 0.60 b | 5.11 ± 0.02 a | −24.7 ± 0.25 f | |
| H-Cd | 22 ± 0.60 b | 4.28 ± 0.01 b | −25.8 ± 0.44 d | |
| Treatments | Accumulated Cd (μg Plant−1 DW) | BCF | TF | AE (µg g−1) | Phytoremediation Strategy | ||
|---|---|---|---|---|---|---|---|
| Root | Shoot | ||||||
| 0 mM NaCl | L-Cd | 12.3 ± 0.10 c | 4.4 ± 0.2 h | 1.1± 0.07 f | 0.42 ± 0.01 f | 221 ± 3.5 g | Phytostabilization |
| H-Cd | 15.0 ± 0.20 a | 6.1 ± 0.2 g | 1.4 ± 0.08 f | 0.45 ± 0.02 f | 404 ± 5.0 e | ||
| 50 mM NaCl | L-Cd | 11.0 ± 0.10 d | 8.9 ± 0.3 f | 5.3 ± 0.10 e | 0.51 ± 0.03 e | 340 ± 4.8 f | Phytostabilization |
| H-Cd | 13.0 ± 0.20 b | 16.0 ± 0.4 e | 6.8 ± 0.20 d | 0.76 ± 0.05 d | 611 ± 3.2 c | ||
| 100 mM NaCl | L-Cd | 2.5 ± 0.05 e | 23.0 ± 0.5 d | 7.4 ± 0.10 c | 1.08 ± 0.01 c | 552 ± 3.0 d | Phytoextraction |
| H-Cd | 2.9 ± 0.04 e | 43.0 ± 0.6 c | 9.9 ± 0.20 b | 1.12 ± 0.06 b | 907 ± 4.1 b | ||
| 200 mM NaCl | L-Cd | 1.4 ± 0.07 f | 55.0 ± 0.9 b | 11.3 ± 0.30 a | 1.23 ± 0.07 a | 690 ± 4.4 c | Phytoextraction |
| H-Cd | 1.9 ± 0.05 f | 62.0 ± 0.8 a | 12.1 ± 0.30 a | 1.23 ± 0.07 a | 1303 ± 6.6 a | ||
| Treatments | ASA (µmol g−1 FW) | GSH (nmol g−1 FW) | PCs (µmol g–1 DW) | Proline (μmol g−1 FW) | |
|---|---|---|---|---|---|
| 0 mM NaCl | 0-Cd | 2.60 ± 0.05 e | 110 ± 2.1 k | 10.91 ± 0.9 g | 5.2 ± 0.07 i |
| L-Cd | 3.50 ± 0.04 d | 170 ± 1.5 h | 28.62 ± 0.4 f | 11.6 ± 0.41 e | |
| H-Cd | 1.40 ± 0.01 h | 201 ± 1.0 g | 37.33 ± 0.3 a | 21.0 ± 0.52 a | |
| 50 mM NaCl | 0-Cd | 2.51 ± 0.01 e | 113 ± 2.0 j | 10.51 ± 0.5 g | 5.1 ± 0.01 i |
| L-Cd | 3.90 ± 0.02 d | 210 ± 1.4 f | 27.03 ± 0.4 f | 9.8 ± 0.21 f | |
| H-Cd | 1.60 ± 0.01 h | 260 ± 2.3 e | 35.11 ± 0.2 b | 19.3 ± 0.30 b | |
| 100 mM NaCl | 0-Cd | 2.61 ± 0.03 e | 116 ± 1.1 i | 11.01 ± 0.5 g | 4.9 ± 0.01 i |
| L-Cd | 4.51 ± 0.03 b | 269 ± 2.3 d | 22.24 ± 0.4 e | 8.5 ± 0.11 g | |
| H-Cd | 2.01 ± 0.01 f | 304 ± 3.0 b | 31.81 ± 0.8 c | 16.6 ± 0.20 c | |
| 200 mM NaCl | 0-Cd | 2.54 ± 0.01 e | 118 ±1.1 i | 10.70 ± 0.2 g | 5.0 ± 0.09 i |
| L-Cd | 5.12 ± 0.03 a | 297 ± 1.6 c | 20.54 ± 0.2 d | 7.2 ± 0.07 h | |
| H-Cd | 2.55 ± 0.01 e | 366 ± 2.2 a | 30.25 ± 0.7 c | 15.4 ± 0.13 d | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salama, F.M.; AL-Huqail, A.A.; Ali, M.; Abeed, A.H.A. Cd Phytoextraction Potential in Halophyte Salicornia fruticosa: Salinity Impact. Plants 2022, 11, 2556. https://doi.org/10.3390/plants11192556
Salama FM, AL-Huqail AA, Ali M, Abeed AHA. Cd Phytoextraction Potential in Halophyte Salicornia fruticosa: Salinity Impact. Plants. 2022; 11(19):2556. https://doi.org/10.3390/plants11192556
Chicago/Turabian StyleSalama, Fawzy Mahmoud, Arwa Abdulkreem AL-Huqail, Mohammed Ali, and Amany H. A. Abeed. 2022. "Cd Phytoextraction Potential in Halophyte Salicornia fruticosa: Salinity Impact" Plants 11, no. 19: 2556. https://doi.org/10.3390/plants11192556
APA StyleSalama, F. M., AL-Huqail, A. A., Ali, M., & Abeed, A. H. A. (2022). Cd Phytoextraction Potential in Halophyte Salicornia fruticosa: Salinity Impact. Plants, 11(19), 2556. https://doi.org/10.3390/plants11192556