
Polyethylene Glycol and Sorbitol-Mediated In Vitro Screening for Drought Stress as an Efficient and Rapid Tool to Reach the Tolerant Cucumis melo L. Genotypes

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Morphological Traits
2.2. Photosynthetic Pigments
2.3. Proline Content
2.4. MDA Content
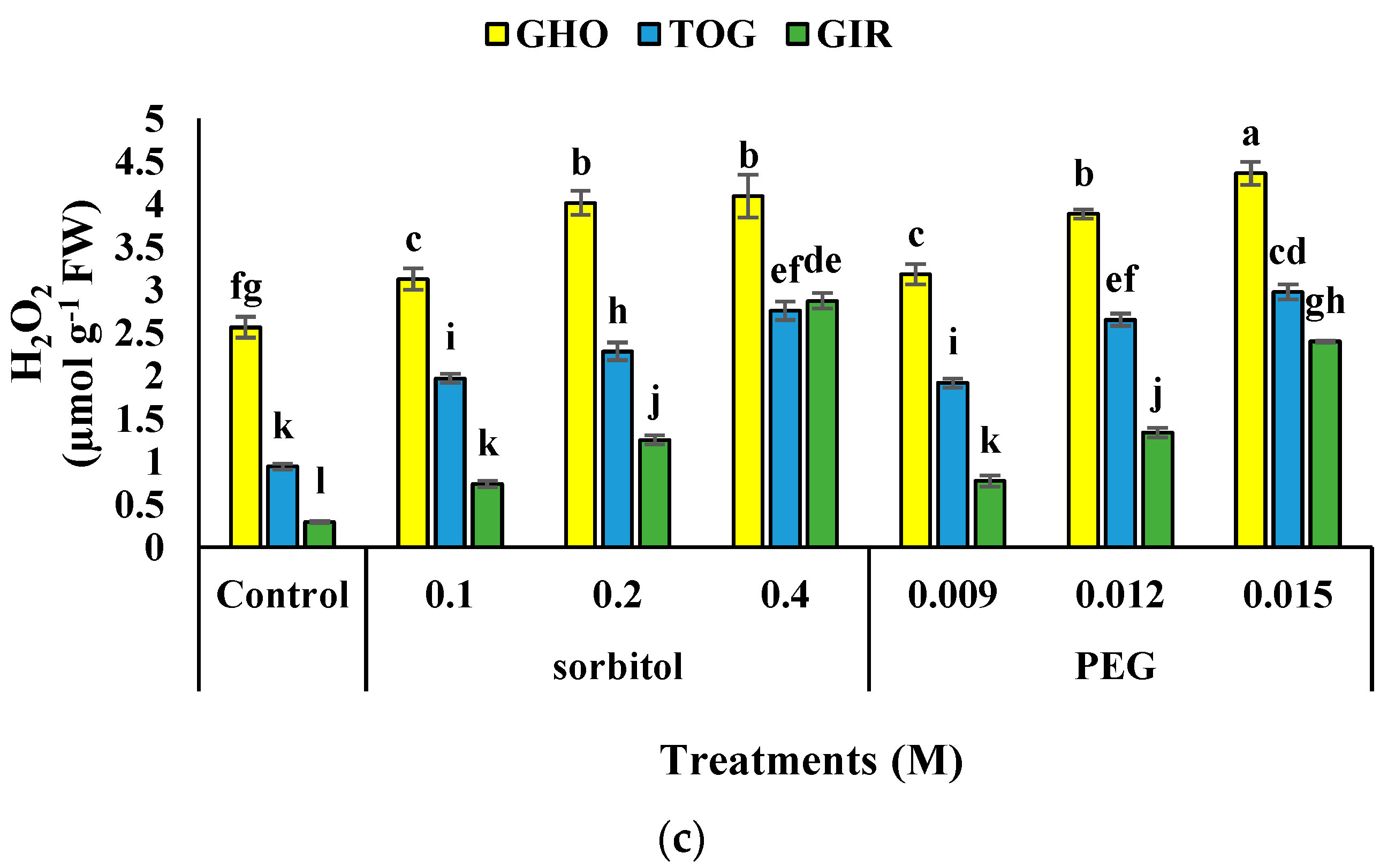
2.5. Hydrogen Peroxide Content
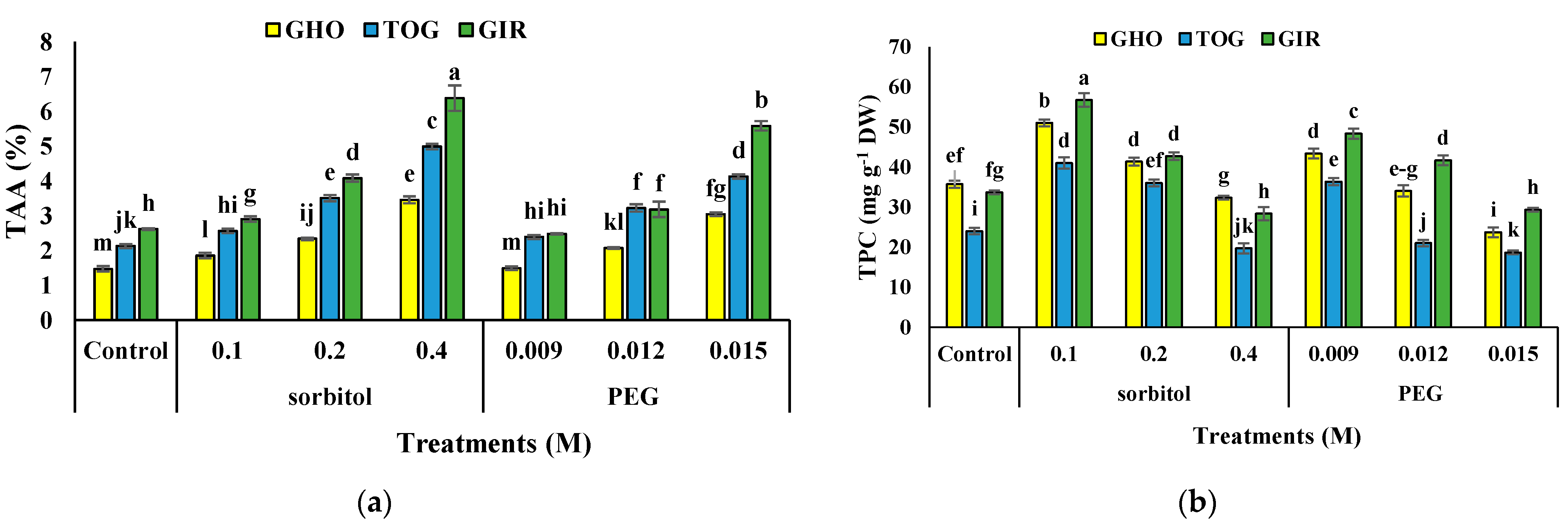
2.6. TAA
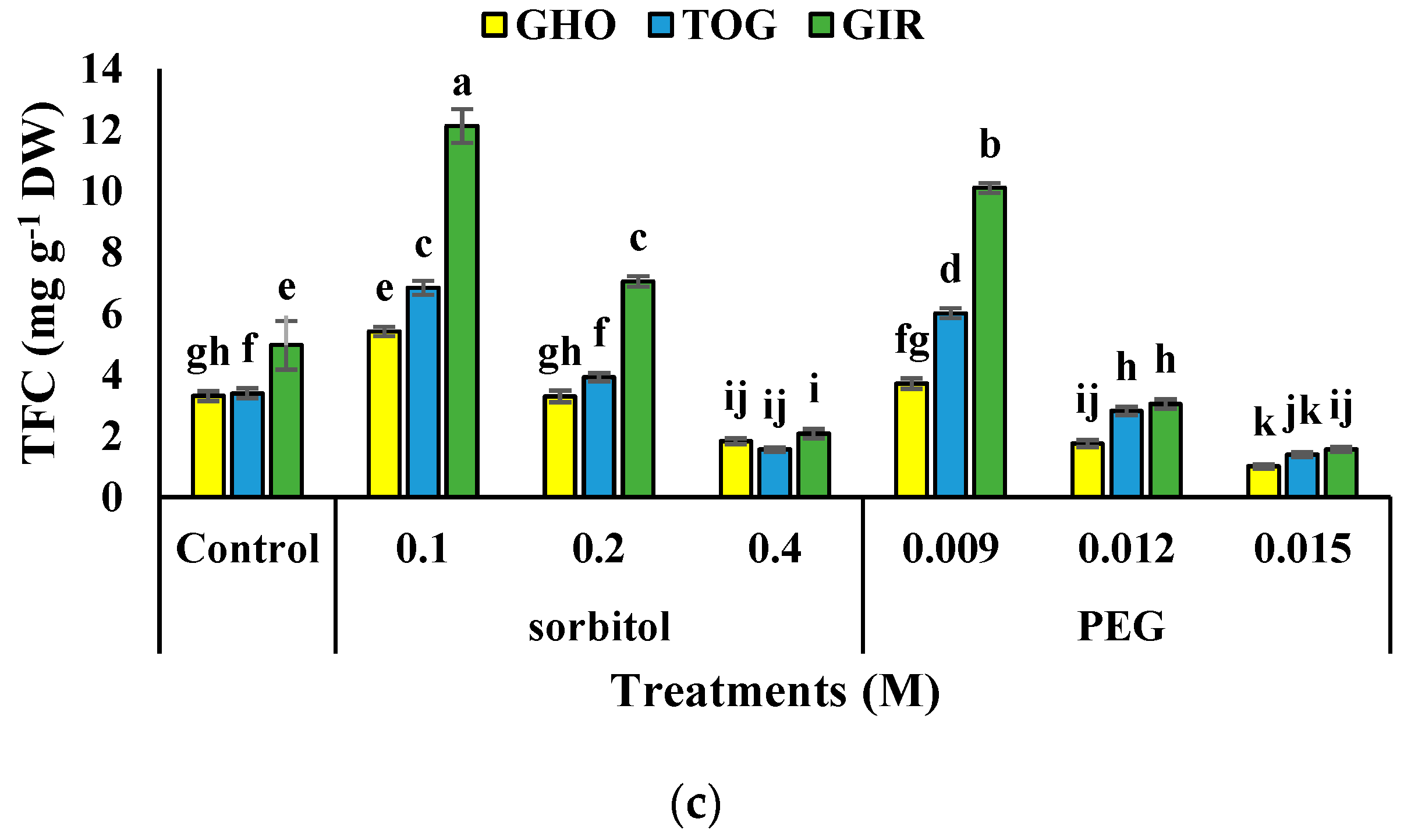
2.7. TPC and TFC
2.8. TSP Content
2.9. Antioxidant Enzyme Activity
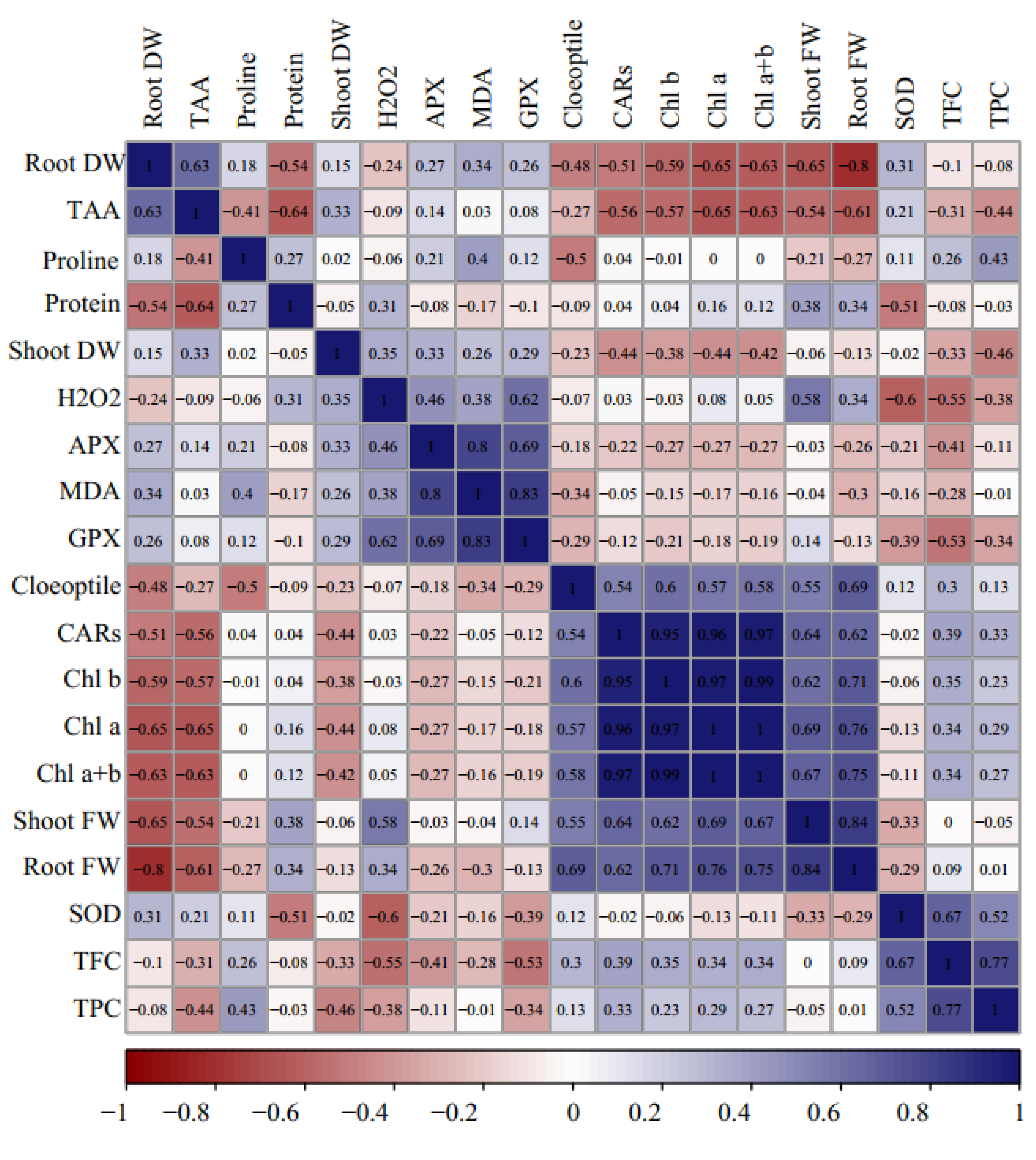
2.10. Correlations, Principal Component Analysis, and Loading Biplot Analysis
3. Discussion
4. Materials and Methods
4.1. Morphological Traits
4.2. Photosynthesis Pigments
4.3. Proline Content
4.4. Malondialdehyde Content (MDA)
4.5. H2O2 Content
4.6. Total Antioxidant Activity (TAA)
4.7. Total Phenolics Content (TPC)
4.8. Total Flavonoids Content (TFC)
4.9. Total Soluble Protein (TSP) Content
4.10. Antioxidant Enzymes Activity
4.11. Statistical Experiments
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Ethics Approval and Consent to Participate
References
- Ouzounidou, G.; Giannakoula, A.; Ilias, I.; Zamanidis, P. Alleviation of drought and salinity stresses on growth, physiology, biochemistry and quality of two Cucumis sativus L. cultivars by Si application. Braz. J. Bot. 2016, 39, 531–539. [Google Scholar] [CrossRef]
- Eskandari Dameneh, H.; Gholami, H.; Telfer, M.W.; Comino, J.R.; Collins, A.L.; Jansen, J.D. Desertification of Iran in the early twenty-first century: Assessment using climate and vegetation indices. Sci. Rep. 2021, 11, 20548. [Google Scholar] [CrossRef] [PubMed]
- Kıran, S.; Furtana, G.B.; Talhouni, M.; Ellialtıoğlu, Ş.Ş. Drought stress mitigation with humic acid in two Cucumis melo L. genotypes differ in their drought tolerance. Bragantia 2019, 78, 490–497. [Google Scholar] [CrossRef] [Green Version]
- Zahedi, S.M.; Hosseini, M.S.; Meybodi, N.D.H.; Abadía, J.; Germ, M.; Gholami, R.; Abdelrahman, M. Evaluation of drought tolerance in three commercial pomegranate cultivars using photosynthetic pigments, yield parameters and biochemical traits as biomarkers. Agric. Water Manag. 2022, 261, 107357. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Burritt, D.J.; Tran, L.-S.P. The use of metabolomic quantitative trait locus mapping and osmotic adjustment traits for the improvement of crop yields under environmental stresses. Semin. Cell Dev. Biol. 2018, 83, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Pourghayoumi, M.; Bakhshi, D.; Rahemi, M.; Kamgar-Haghighi, A.A.; Aalami, A. The physiological responses of various pomegranate cultivars to drought stress and recovery in order to screen for drought tolerance. Sci. Hortic. 2017, 217, 164–172. [Google Scholar] [CrossRef]
- Kavas, M.; Baloğlu, M.C.; Akca, O.; Köse, F.S.; Gökçay, D. Effect of drought stress on oxidative damage and antioxidant enzyme activity in melon seedlings. Turk. J. Biol. 2013, 37, 491–498. [Google Scholar] [CrossRef]
- Munger, H.; Robinson, R. Nomenclature of Cucumis melo L. Cucurbit. Genet. Coop Rep. 1991, 14, 43–44. [Google Scholar]
- Rai, M.K.; Kalia, R.K.; Singh, R.; Gangola, M.P.; Dhawan, A. Developing stress tolerant plants through in vitro selection—An overview of the recent progress. Environ. Exp. Bot. 2011, 71, 89–98. [Google Scholar] [CrossRef]
- Dwivedi, S.L.; Ceccarelli, S.; Blair, M.W.; Upadhyaya, H.D.; Are, A.K.; Ortiz, R. Landrace germplasm for improving yield and abiotic stress adaptation. Trends Plant Sci. 2016, 21, 31–42. [Google Scholar] [CrossRef]
- Errabii, T.; Gandonou, C.B.; Essalmani, H.; Abrini, J.; Idaomar, M.; Skali-Senhaji, N. Growth, proline and ion accumulation in sugarcane callus cultures under drought-induced osmotic stress and its subsequent relief. Afr. J. Biotechnol. 2006, 5, 1488–1493. [Google Scholar]
- Munns, R.; Husain, S.; Rivelli, A.R.; James, R.A.; Condon, A.; Lindsay, M.P.; Lagudah, E.S.; Schachtman, D.P.; Hare, R.A. Avenues for increasing salt tolerance of crops, and the role of physiologically based selection traits. In Progress in Plant Nutrition: Plenary Lectures of the XIV International Plant Nutrition Colloquium; Springer Nature: Hannover, The Netherlands, 2002; pp. 93–105. [Google Scholar]
- Gopal, J.; Iwama, K. In vitro screening of potato against water-stress mediated through sorbitol and polyethylene glycol. Plant Cell Rep. 2007, 26, 693–700. [Google Scholar] [CrossRef] [PubMed]
- D’souza, A.A.; Shegokar, R. Polyethylene glycol (PEG): A versatile polymer for pharmaceutical applications. Expert Opin. Drug Deliv. 2016, 13, 1257–1275. [Google Scholar] [CrossRef] [PubMed]
- Siaga, E.; Maharijaya, A.; Rahayu, M.S. Plant Growth of Eggplant (Solanum melongena L.) In Vitro in Drought Stress Polyethylene Glycol (PEG). BIOVALENTIA Biol. Res. J. 2016, 2, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Cammenga, H.; Figura, L.; Zielasko, B. Thermal behaviour of some sugar alcohols. J. Therm. Anal. Calorim. 1996, 47, 427–434. [Google Scholar] [CrossRef]
- Ansari, W.A.; Atri, N.; Ahmad, J.; Qureshi, M.I.; Singh, B.; Kumar, R.; Rai, V.; Pandey, S. Drought mediated physiological and molecular changes in muskmelon (Cucumis melo L.). PLoS ONE 2019, 14, e0222647. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol 2009, 11, 100–105. [Google Scholar]
- Abu-Romman, S.; Suwwan, M.; Al-Shadiadeh, A.; Hasan, H. Effects of osmotic stress on cucumber (Cucumis sativus L.) microshoots cultured on proliferation medium. World Appl. Sci. J. 2012, 20, 177–181. [Google Scholar]
- Hellal, F.; El-Shabrawi, H.; Abd El-Hady, M.; Khatab, I.; El-Sayed, S.; Abdelly, C. Influence of PEG induced drought stress on molecular and biochemical constituents and seedling growth of Egyptian barley cultivars. J. Genet. Eng. Biotechnol. 2018, 16, 203–212. [Google Scholar] [CrossRef]
- Sairam, R.; Saxena, D. Oxidative stress and antioxidants in wheat genotypes: Possible mechanism of water stress tolerance. J. Agron. Crop Sci. 2000, 184, 55–61. [Google Scholar] [CrossRef]
- Shivakrishna, P.; Reddy, K.A.; Rao, D.M. Effect of PEG-6000 imposed drought stress on RNA content, relative water content (RWC), and chlorophyll content in peanut leaves and roots. Saudi J. Biol. Sci. 2018, 25, 285–289. [Google Scholar]
- Sun, Y.; Wang, H.; Liu, S.; Peng, X. Exogenous application of hydrogen peroxide alleviates drought stress in cucumber seedlings. S. Afr. J. Bot. 2016, 106, 23–28. [Google Scholar] [CrossRef]
- Per, T.S.; Khan, N.A.; Reddy, P.S.; Masood, A.; Hasanuzzaman, M.; Khan, M.I.R.; Anjum, N.A. Approaches in modulating proline metabolism in plants for salt and drought stress tolerance: Phytohormones, mineral nutrients and transgenics. Plant Physiol. Biochem. 2017, 115, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Jariteh, M.; Ebrahimzadeh, H.; Niknam, V.; Mirmasoumi, M.; Vahdati, K. Developmental changes of protein, proline and some antioxidant enzymes activities in somatic and zygotic embryos of Persian walnut (Juglans regia L.). Plant Cell Tissue Organ Cult. (PCTOC) 2015, 122, 101–115. [Google Scholar] [CrossRef]
- Gharibi, S.; Tabatabaei, B.E.S.; Saeidi, G.; Goli, S.A.H. Effect of drought stress on total phenolic, lipid peroxidation, and antioxidant activity of Achillea species. Appl. Biochem. Biotechnol. 2016, 178, 796–809. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Shirani Bidabadi, S.; Sharifi, P. Strigolactone and methyl Jasmonate-induced antioxidant defense and the composition alterations of different active compounds in Dracocephalum kotschyi Boiss under drought stress. J. Plant Growth Regul. 2021, 40, 878–889. [Google Scholar] [CrossRef]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Differential responses of phenolic compounds of Brassica napus under drought stress. Iran. J. Plant Physiol. 2018, 8, 2417–2425. [Google Scholar]
- Josipović, A.; Sudar, R.; Sudarić, A.; Jurković, V.; Matoša Kočar, M.; Markulj Kulundžić, A. Total phenolic and total flavonoid content variability of soybean genotypes in eastern Croatia. Croat. J. Food Sci. Technol. 2016, 8, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Anjum, s.A.; Xie, X.; Wang, L.C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Sebnem, K. Effects of drought and salt stresses on growth, stomatal conductance, leaf water and osmotic potentials of melon genotypes (Cucumis melo L.). Afr. J. Agric. Res. 2012, 7, 775–781. [Google Scholar]
- Ghassemi-Golezani, K.; Farhadi, N.; Nikpour-Rashidabad, N. Responses of in vitro-cultured Allium hirtifolium to exogenous sodium nitroprusside under PEG-imposed drought stress. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 133, 237–248. [Google Scholar] [CrossRef]
- Sharma, P.; Gujral, H.S.; Singh, B. Antioxidant activity of barley as affected by extrusion cooking. Food Chem. 2012, 131, 1406–1413. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Behboudi, F.; Tahmasebi Sarvestani, Z.; Kassaee, M.Z.; Modares Sanavi, S.; Sorooshzadeh, A. Improving growth and yield of wheat under drought stress via application of SiO2 nanoparticles. J. Agric. Sci. Technol. 2018, 20, 1479–1492. [Google Scholar]
- Khudsar, T.; Iqbal, M. Cadmium-induced changes in leaf epidermes, photosynthetic rate and pigment concentrations in Cajanus cajan. Biol. Plant. 2001, 44, 59–64. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2009; pp. 153–188. [Google Scholar]
- Reid, D.S. Water Activity. In Water Activity in Foods; Wiley Online Library: Hoboken, NJ, USA, 2020; pp. 13–26. [Google Scholar]
- Michel, B.E.; Kaufmann, M.R. The osmotic potential of polyethylene glycol 6000. Plant Physiol. 1973, 51, 914–916. [Google Scholar] [CrossRef]
- Armon, D. Copper enzymes in isolated chloroplast. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Bates, L.; Waldren, R.P.; Tare, I.D. Rapid Determination of Free Proline for Water Stress Studies. Plant Soil 1973, 33, 205–208. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Saxena, R.; Singh, S. Chromium induced lipid peroxidation in the plants of Pistia stratiotes L.: Role of antioxidants and antioxidant enzymes. Chemosphere 2005, 58, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Chiou, M.J.; Wang, Y.D.; Kuo, C.M.; Chen, J.C.; Chen, J.Y. Functional analysis of mitogen-activated protein kinase-3 (MAPK3) and its regulation of the promoter region in zebrafish. DNA Cell Biol. 2007, 26, 781–790. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.-C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 45–52. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Treatment | Concentration (M) | Character | ||||
|---|---|---|---|---|---|---|---|
| Coleoptile Length (mm) | Shoot FW (g Plantlet−1) | Shoot DW (%) | Root FW (g Plantlet−1) | Root DW (%) | |||
| GHO | Control | 0 | 39.82 ± 4.18 b | 0.913 ± 0.04 c | 1.360 ± 0.38 e | 0.293 ± 0.03 b | 4.131 ± 1.11 i–k |
| Sorbitol | 0.1 | 30.24 ± 0.87 f–i | 0.596 ± 0.03 d | 1.850 ± 0.33 d,e | 0.213 ± 0.02 c,d | 5.113 ± 0.28 g–k | |
| 0.2 | 25.88 ± 0.90 i–l | 0.516 ± 0.11 d,e | 5.103 ± 2.27 b,c | 0.136 ± 0.04 e–g | 8.387 ± 1.72 b–e | ||
| 0.4 | 25.57 ± 0.77 j–l | 0.420 ± 0.02 f,g | 7.270 ± 1.74 a | 0.09 ± 0.02 f–h | 9.037 ± 1.41 b | ||
| PEG | 0.009 | 31.65 ± 2.12 d–g | 0.59 ± 0.11 d | 1.757 ± 0.44 e | 0.206 ± 0.01 c,d | 4.723 ± 0.66 h–k | |
| 0.012 | 29.18 ± 1.37 f–j | 0.396 ± 0.06 f,g | 2.330 ± 0.32 d,e | 0.213 ± 0.01 c,d | 6.473 ± 0.39 e–h | ||
| 0.015 | 24.23 ± 0.68 k,l | 0.37 ± 0.04 g,h | 2.873 ± 0.35 d,e | 0.11 ± 0.06 d,e | 6.547 ± 0.41 e–h | ||
| TOG | Control | 0 | 44.50 ± 4.30 a | 1.173 ± 0.09 a | 2.583 ± 0.39 c–e | 0.446 ± 0.07 a | 1.523 ± 0.13 l |
| Sorbitol | 0.1 | 35.64 ± 2.41 c,d | 1.053 ± 0.01 b | 3.613 ± 1.69 c,d | 0.233 ± 0.01 c | 3.843 ± 1.36 k | |
| 0.2 | 34.95 ± 1.90 c–e | 0.500 ± 0.01 d–f | 6.330 ± 0.49 a,b | 0.200 ± 0.02 c,d | 5.273 ± 0.22 g–k | ||
| 0.4 | 31.31 ± 2.16 d–g | 0.400 ± 0.05 f,g | 7.383 ± 0.86 a | 0.070 ± 0.04 h | 8.917 ± 0.88 b,c | ||
| PEG | 0.009 | 27.24 ± 0.72 g–k | 0.413 ± 0.01 e,f | 2.680 ± 0.08 d,e | 0.126 ± 0.01 e–h | 6.257 ± 0.78 f–i | |
| 0.012 | 26.59 ± 1.84 h–k | 0.316 ± 0.14 g–i | 3.563 ± 0.89c,d | 0.120 ± 0.01 e–h | 6.91 ± 0.07 f–i | ||
| 0.015 | 22.23 ± 0.81 l | 0.320 ± 0.03 g–i | 5.657 ± 0.88 b | 0.096 ± 0.01 f–h | 7.060 ± 0.99 c–g | ||
| GIR | Control | 0 | 37.12 ± 2.45 b,c | 0.263 ± 0.05 h,i | 1.630 ± 0.29 e | 0.206 ± 0.02 c,d | 4.263 ± 0.27 j,k |
| Sorbitol | 0.1 | 32.96 ± 0.20 c–f | 0.263 ± 0.04 h,i | 1.840 ± 0.37 d,e | 0.140 ± 0.01 e,f | 8.620 ± 1.19 b–d | |
| 0.2 | 31.52 ± 0.84 d–g | 0.243 ± 0.01 i–k | 2.297 ± 0.91 c–e | 0.110 ± 0.01e–h | 9.083 ± 0.60 b | ||
| 0.4 | 29.43 ± 2.00 f–j | 0.183 ± 0.01 j,k | 5.637 ± 0.02 b,c | 0.073 ± 0.01 h | 11.44 ± 1.35 a | ||
| PEG | 0.009 | 34.99 ± 2.01 c–e | 0.233 ± 0.02 i,j | 2.150 ± 0.16 d,e | 0.163 ± 0.02 d,e | 5.863 ± 0.23 f–j | |
| 0.012 | 30.60 ± 0.44 e–h | 0.170 ± 0.04 j,k | 2.397 ± 0.15 c–e | 0.133 ± 0.03 e–g | 6.803 ± 0.48 d–g | ||
| 0.015 | 28.19 ± 0.82 g–k | 0.133 ± 0.01 k | 3.553 ± 0.13 c,d | 0.090 ± 0.01 f–h | 7.567 ± 0.53 b–f | ||
| S.O.V. | |||||||
| Genotype | 42.210 ** | 0.901 ** | 17.612 ** | 0.022 ** | 12.828 ** | ||
| Drought | 199.411 ** | 0.310 ** | 17.694 ** | 0.046 ** | 38.264 ** | ||
| Genotype × Drought | 38.582 ** | 0.095 ** | 6.582 ** | 0.011 ** | 8.639 ** | ||
| Error | 5.664 | 0.004 | 1.107 | 0.001 | 1.108 | ||
| C.V. (%) | 7.64 | 14.56 | 29.91 | 19.19 | 15.57 | ||
| Genotype | Treatment | Concentration (M) | Character | |||
|---|---|---|---|---|---|---|
| Chl a (mg kg−1 FW) | Chl b (mg kg−1 FW) | Chl a + b (mg kg−1 FW) | CARs (mg kg−1 FW) | |||
| GHO | Control | 0 | 30.60 ± 0.44 a | 12.61 ± 0.07 b,c | 43.22 ± 0.51 a | 7.543 ± 0.66 a |
| Sorbitol | 0.1 | 23.36 ± 1.90 c | 9.743 ± 0.55 e | 33.10 ± 2.44 c | 5.423 ± 0.27 c | |
| 0.2 | 18.12 ± 1.05 d | 6.650 ± 0.44 f | 24.77 ± 1.48 d | 4.407 ± 0.32 d,e | ||
| 0.4 | 14.24 ± 2.03 e,f | 5.913 ± 0.74 f–h | 20.16 ± 2.78 e,f | 3.723 ± 0.60 e,f | ||
| PEG | 0.009 | 18.27 ± 1.04 d | 6.433 ± 0.06 f,g | 24.70 ± 0.99 d | 4.640 ± 0.37 c,d | |
| 0.012 | 16.40 ± 0.29 d,e | 6.400 ± 1.37 f,g | 22.80 ± 1.56 d,e | 3.343 ± 0.05 f,g | ||
| 0.015 | 13.92 ± 0.19 f,g | 5.953 ± 0.08 f–h | 19.87 ± 0.27 e–g | 2.960 ± 0.12 f,g | ||
| OG | Control | 0 | 31.72 ± 1.66 a | 13.38 ± 0.74 b | 45.10 ± 2.39 a | 6.907 ± 0.71 a,b |
| Sorbitol | 0.1 | 27.33 ± 0.10 b | 11.20 ± 0.04 d | 38.54 ± 0.14 b | 6.463 ± 0.84 b | |
| 0.2 | 12.52 ± 1.13 f–h | 5.343 ± 1.76 f–i | 17.86 ± 2.08 f–h | 2.970 ± 0.29 f,g | ||
| 0.4 | 9.21 ± 0.65 i,j | 4.937 ± 0.43 g–j | 13.27 ± 2.03 j–l | 2.687 ± 0.53 g–i | ||
| EG | 0.009 | 11.87 ± 0.87 f–i | 5.247 ± 0.42 f–i | 17.11 ± 1.26 f–i | 2.770 ± 0.24 g,h | |
| 0.012 | 11.63 ± 1.28 g–i | 4.737 ± 0.47 h–j | 16.36 ± 1.75 g–j | 2.643 ± 0.29 g–i | ||
| 0.015 | 10.09 ± 1.27 h,i | 4.017 ± 0.62 i–k | 14.11 ± 1.53 i–l | 2.627 ± 0.39 g–i | ||
| GIR | Control | 0 | 30.58 ± 1.91 a | 15.32 ± 0.81 a | 45.89 ± 2.67 a | 7.200 ± 0.33 a,b |
| Sorbitol | 0.1 | 26.73 ± 1.26 b | 11.70 ± 0.60 c,d | 38.43 ± 1.85 b | 6.670 ± 0.37 a,b | |
| 0.2 | 9.563 ± 1.50 i,j | 4.263 ± 0.40 i–k | 13.59 ± 0.81 i–l | 2.750 ± 0.18 g,h | ||
| 0.4 | 9.327 ± 0.53 i,j | 3.707 ± 0.53 j–l | 11.46 ± 0.36 k–m | 2.583 ± 0.33 g–i | ||
| PEG | 0.009 | 11.17 ± 0.13 h,i | 4.753 ± 0.22 h–j | 15.93 ± 0.09 h–j | 2.553 ± 0.09 g–i | |
| 0.012 | 7.300 ± 0.52 j,k | 3.010 ± 0.28 j–l | 10.31 ± 0.79 l,m | 1.823 ± 0.31 h,i | ||
| 0.015 | 6.643 ± 1.00 k | 2.650 ± 0.28 l | 9.293 ± 1.28 m | 1.747 ± 0.23 i | ||
| S.O.V. | ||||||
| Genotype | 126.906 ** | 7.460 ** | 195.443 ** | 5.202 ** | ||
| Drought | 624.092 ** | 125.034 ** | 1305.071 ** | 31.152 ** | ||
| Genotype × Drought | 23.462 ** | 4.756 ** | 44.513 ** | 1.588 ** | ||
| Error | 1.983 | 0.658 | 3.885 | 0.251 | ||
| C.V. (%) | 8.50 | 11.51 | 8.35 | 12.46 | ||
| Mean Square | |||||||
|---|---|---|---|---|---|---|---|
| S.O.V. | df | Proline | MDA | H2O2 | TAA | TFC | TPC |
| Genotype | 2 | 193.397 ** | 65.835 ** | 26.620 ** | 14.484 ** | 47.989 ** | 829.397 ** |
| Drought | 6 | 227.600 ** | 27.153 ** | 4.726 ** | 10.926 ** | 57.348 ** | 763.995 ** |
| Genotype × Drought | 12 | 126.294 ** | 9.013 ** | 0.266 ** | 0.540 ** | 6.179 ** | 26.860 ** |
| Error | 42 | 2.174 | 0.291 | 0.015 | 0.020 | 0.095 | 1.762 |
| C.V. (%) | 8.52 | 10.72 | 5.10 | 4.54 | 7.38 | 3.77 | |
| Mean Square | |||||
|---|---|---|---|---|---|
| df | TSP Content | APX Specialactivity | GPX Special Activity | SOD Special Activity | |
| Genotype | 2 | 2.212 ** | 0.802 ** | 4133.235 ** | 2.256 ** |
| Drought | 6 | 1.387 ** | 0.627 ** | 341.848 ** | 0.584 ** |
| Genotype × Drought | 12 | 0.146 ** | 0.265 ** | 62.531 ** | 0.141 ** |
| Error | 42 | 0.034 | 0.066 | 4.619 | 0.017 |
| C.V. (%) | 13.916 | 61.09 | 7.29 | 15.07 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mehmandar, M.N.; Rasouli, F.; Giglou, M.T.; Zahedi, S.M.; Hassanpouraghdam, M.B.; Aazami, M.A.; Tajaragh, R.P.; Ryant, P.; Mlcek, J. Polyethylene Glycol and Sorbitol-Mediated In Vitro Screening for Drought Stress as an Efficient and Rapid Tool to Reach the Tolerant Cucumis melo L. Genotypes. Plants 2023, 12, 870. https://doi.org/10.3390/plants12040870
Mehmandar MN, Rasouli F, Giglou MT, Zahedi SM, Hassanpouraghdam MB, Aazami MA, Tajaragh RP, Ryant P, Mlcek J. Polyethylene Glycol and Sorbitol-Mediated In Vitro Screening for Drought Stress as an Efficient and Rapid Tool to Reach the Tolerant Cucumis melo L. Genotypes. Plants. 2023; 12(4):870. https://doi.org/10.3390/plants12040870
Chicago/Turabian StyleMehmandar, Maryam Nekoee, Farzad Rasouli, Mousa Torabi Giglou, Seyed Morteza Zahedi, Mohammad Bagher Hassanpouraghdam, Mohammad Ali Aazami, Rana Panahi Tajaragh, Pavel Ryant, and Jiri Mlcek. 2023. "Polyethylene Glycol and Sorbitol-Mediated In Vitro Screening for Drought Stress as an Efficient and Rapid Tool to Reach the Tolerant Cucumis melo L. Genotypes" Plants 12, no. 4: 870. https://doi.org/10.3390/plants12040870
APA StyleMehmandar, M. N., Rasouli, F., Giglou, M. T., Zahedi, S. M., Hassanpouraghdam, M. B., Aazami, M. A., Tajaragh, R. P., Ryant, P., & Mlcek, J. (2023). Polyethylene Glycol and Sorbitol-Mediated In Vitro Screening for Drought Stress as an Efficient and Rapid Tool to Reach the Tolerant Cucumis melo L. Genotypes. Plants, 12(4), 870. https://doi.org/10.3390/plants12040870