
Impact of a DANA Event on the Thermal Response of Nectarine Trees

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
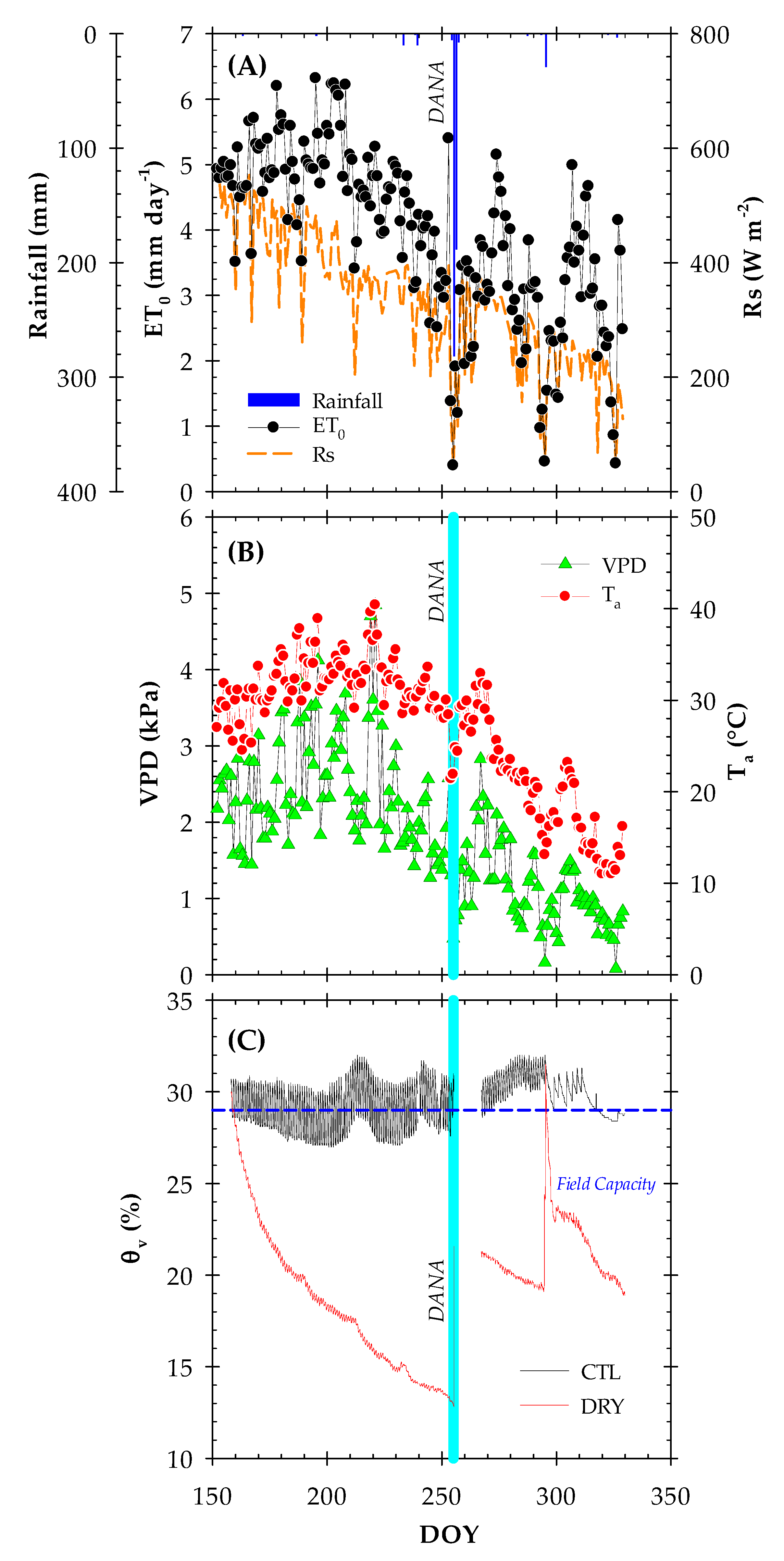
2.1. Environmental Conditions and Soil Water Content
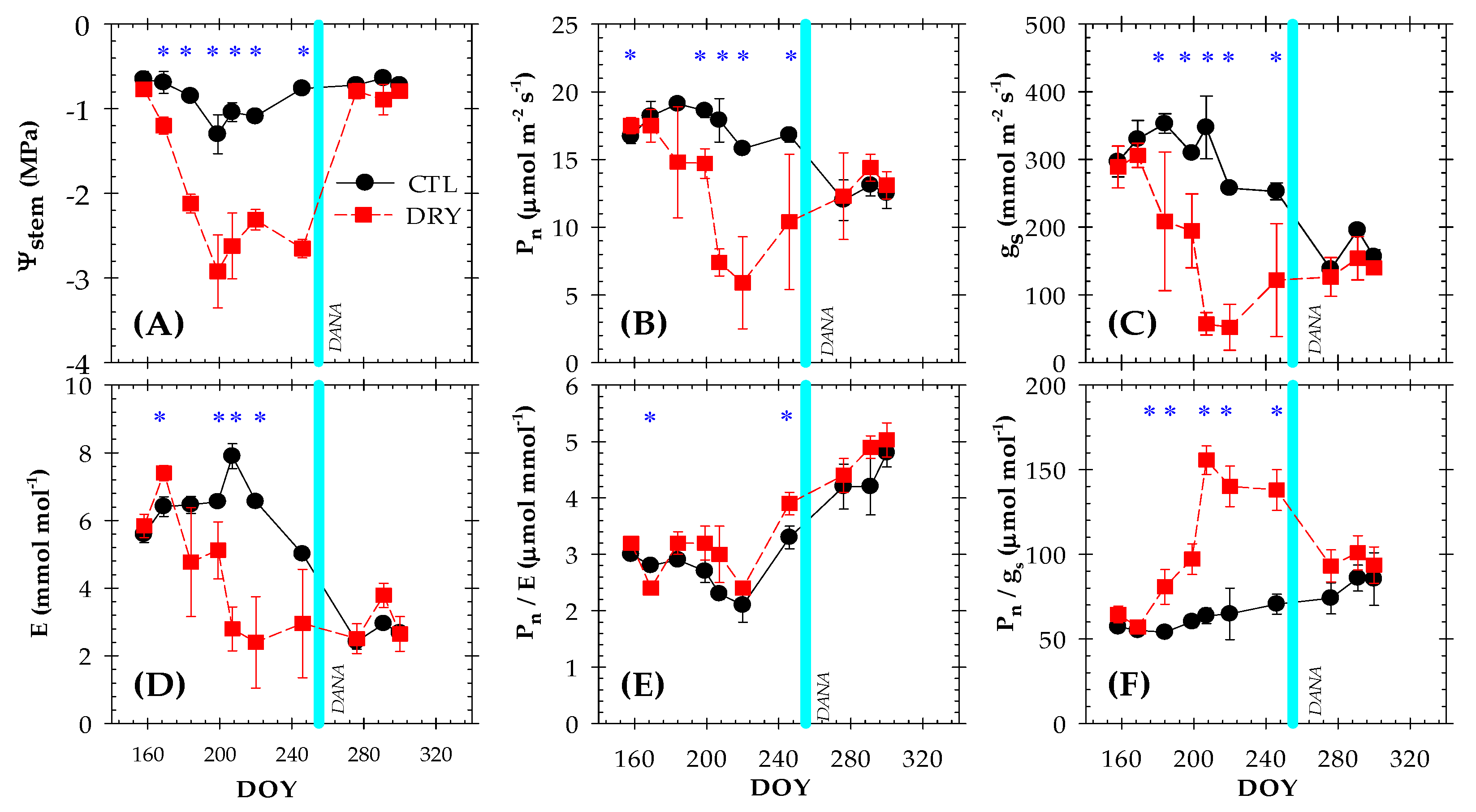
2.2. Plant Water Status and Leaf Gas Exchange
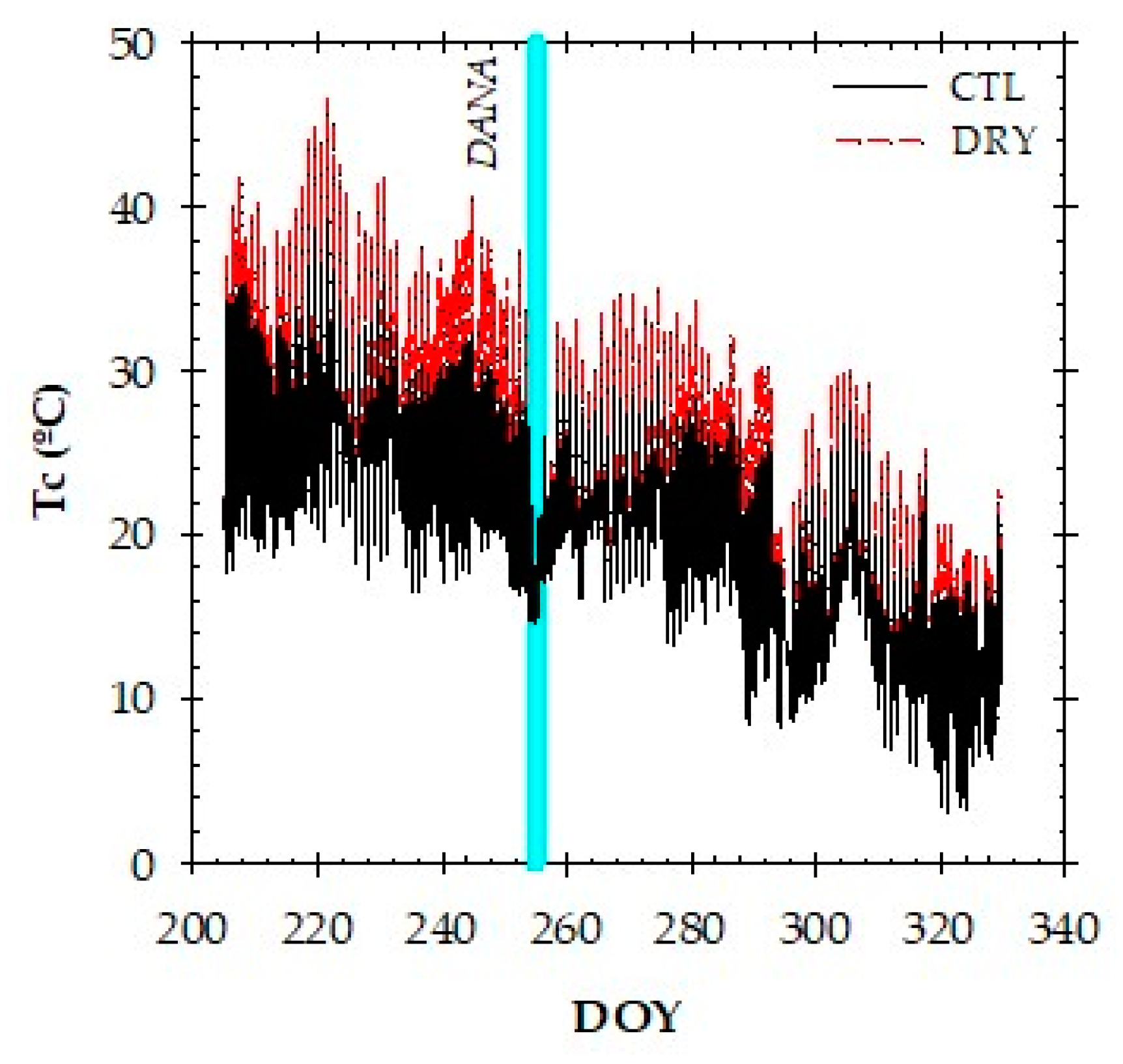
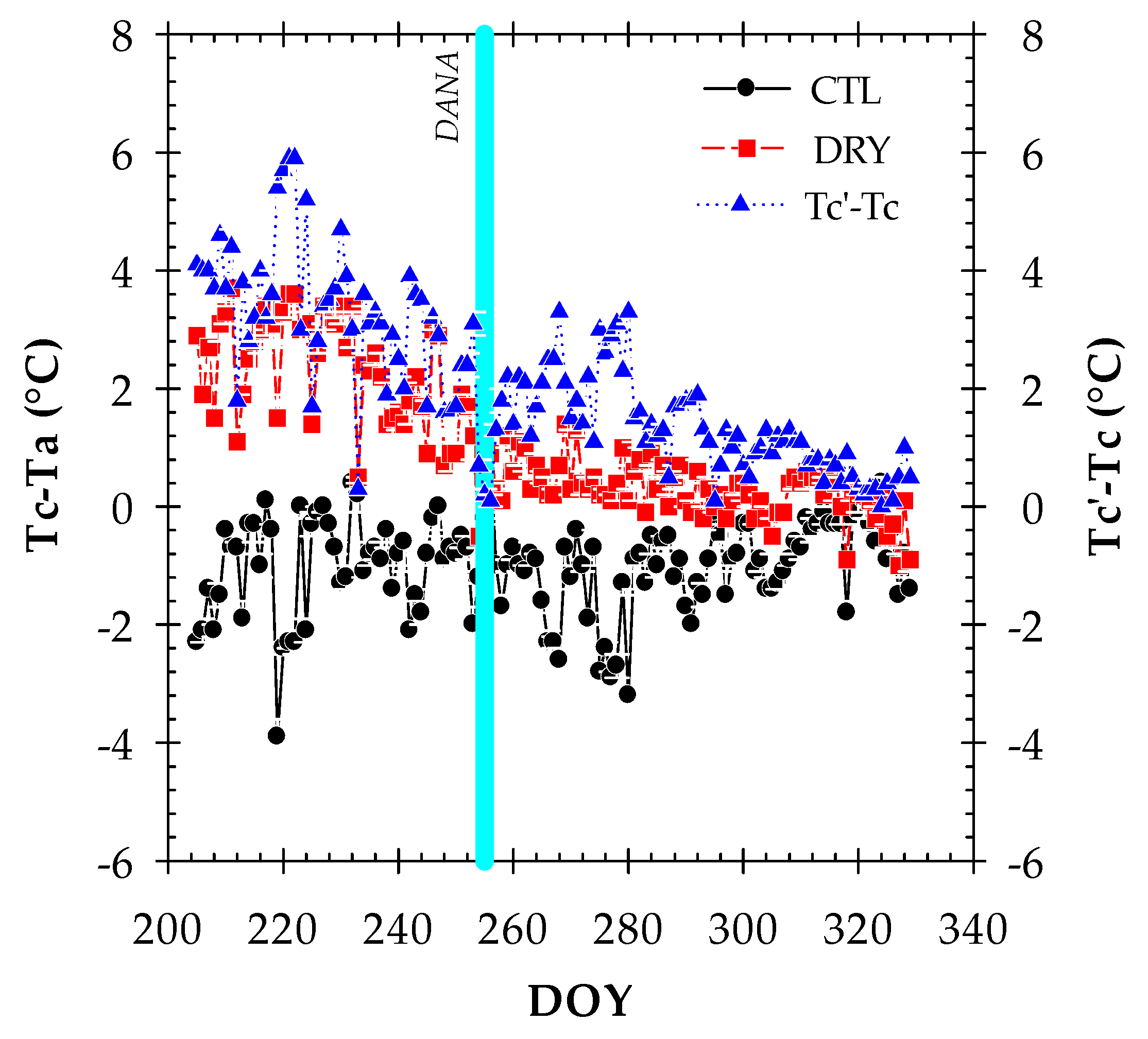
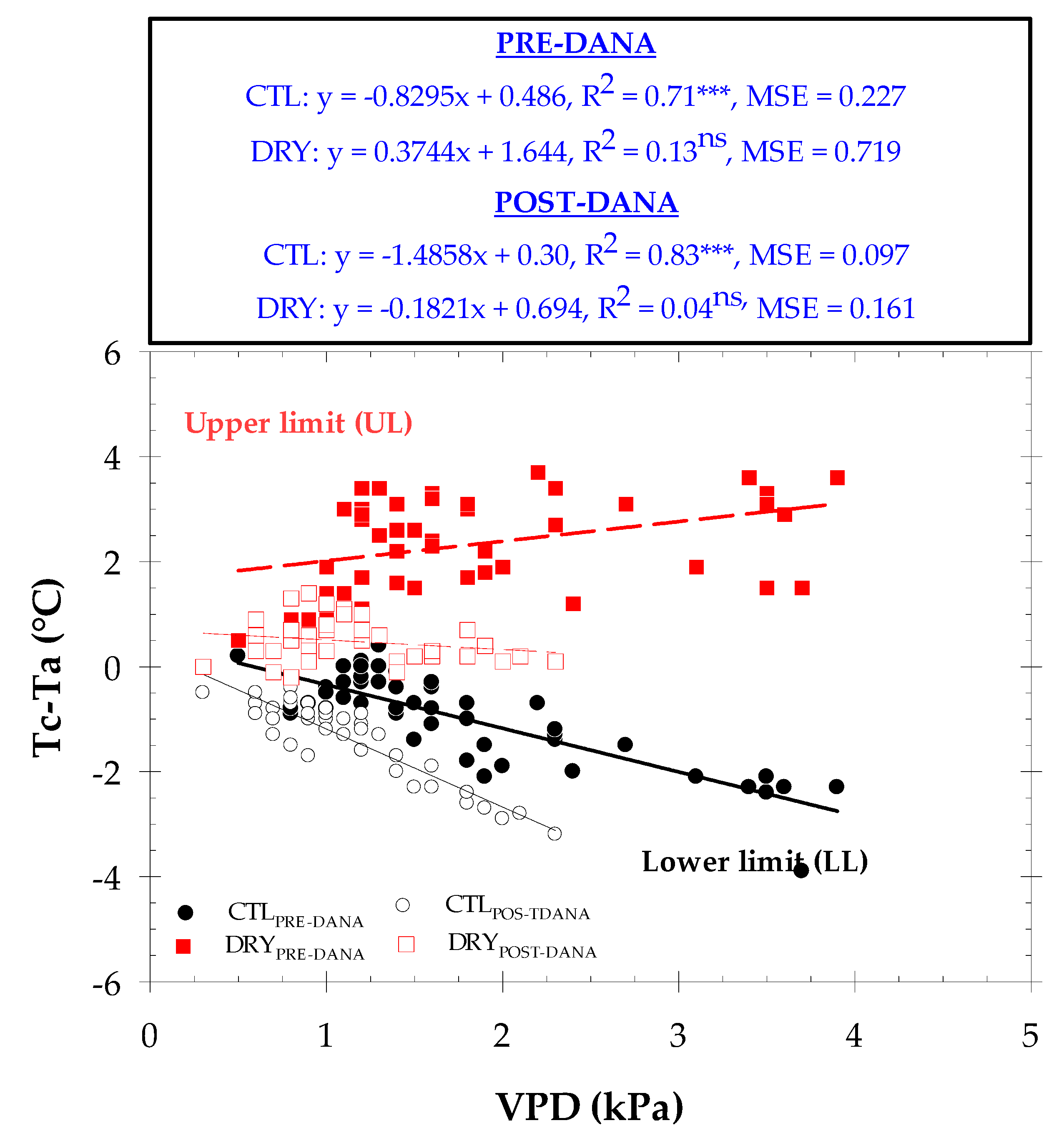
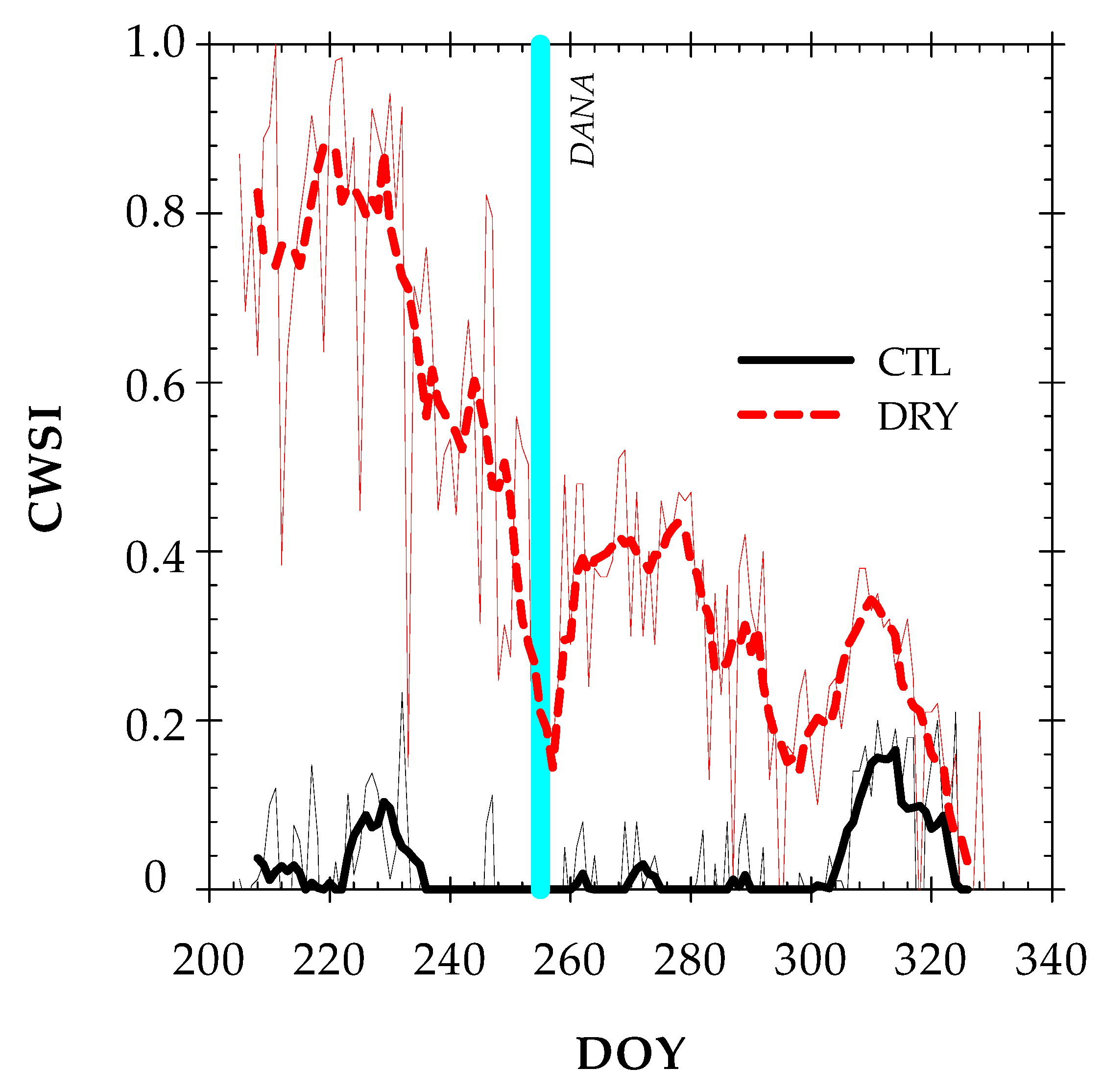
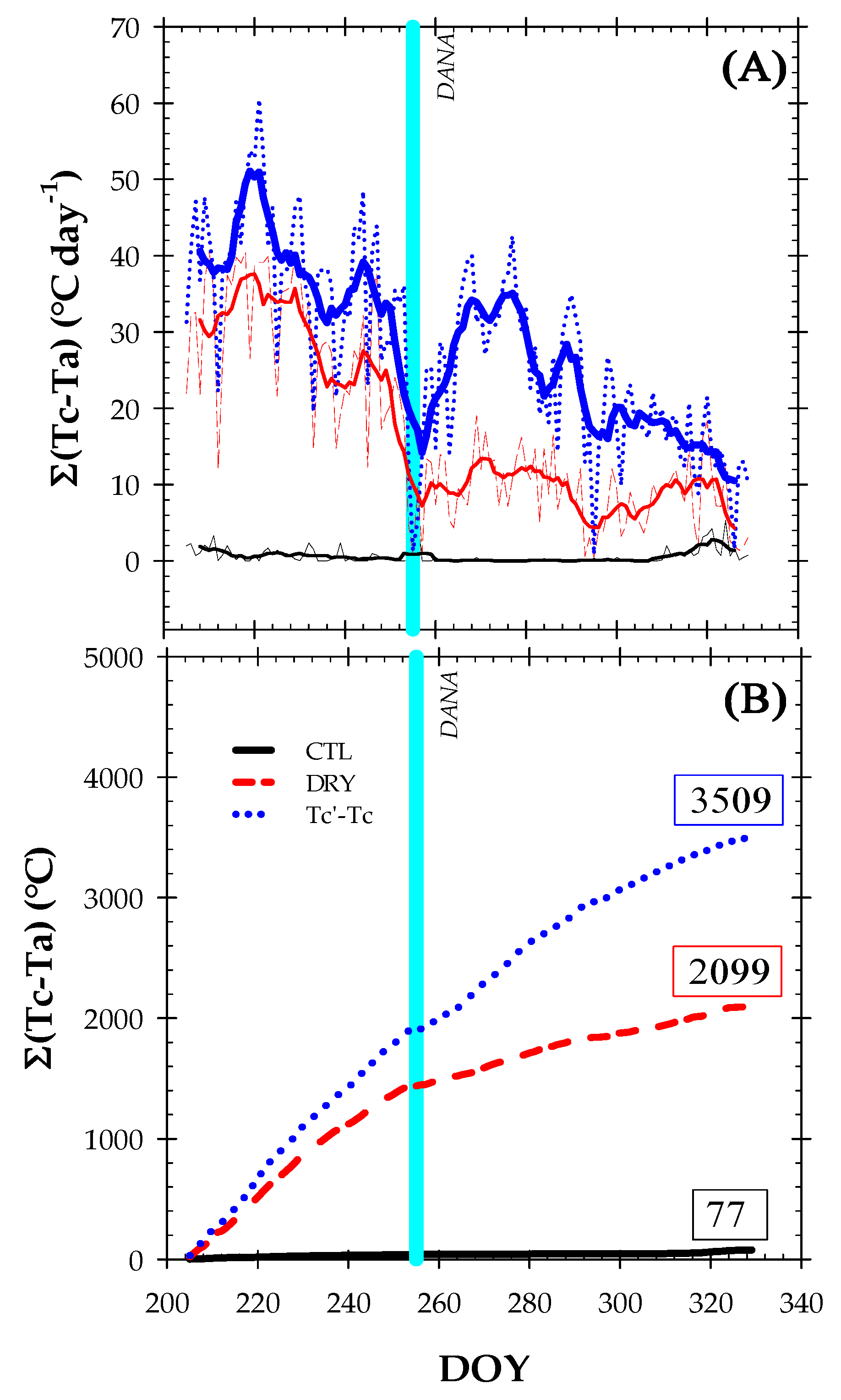
2.3. Canopy Temperature and Thermal Indexes
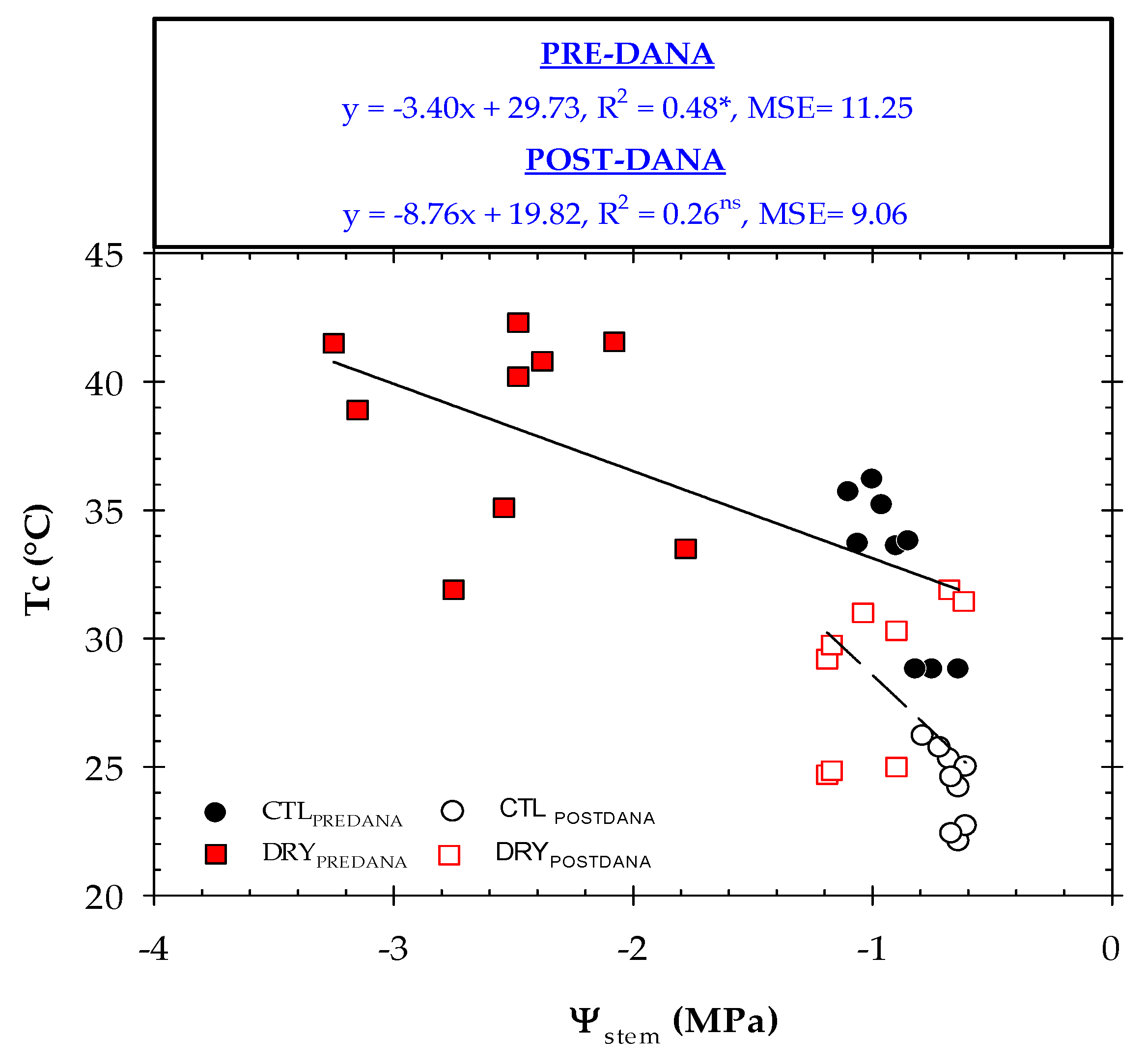
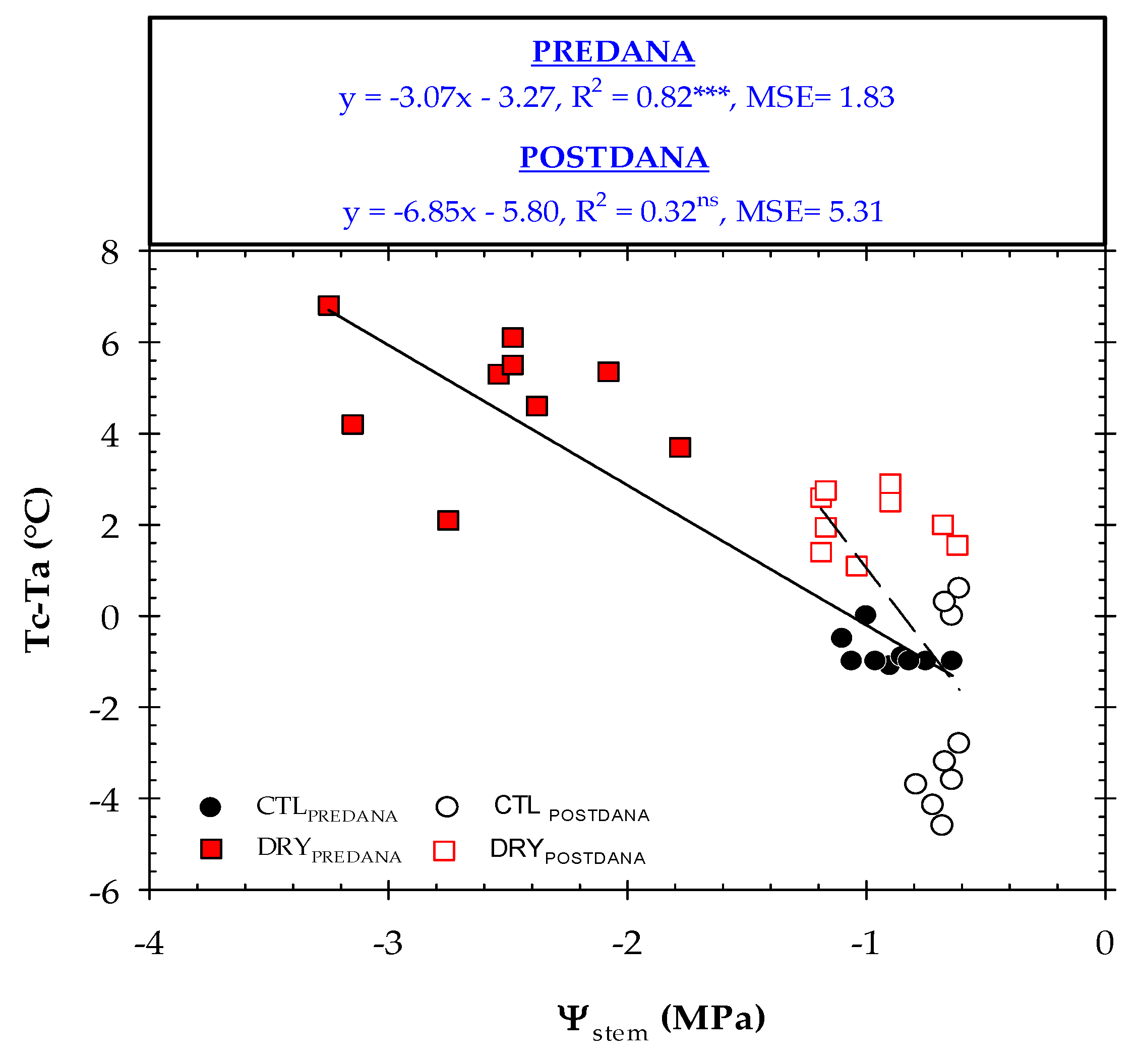
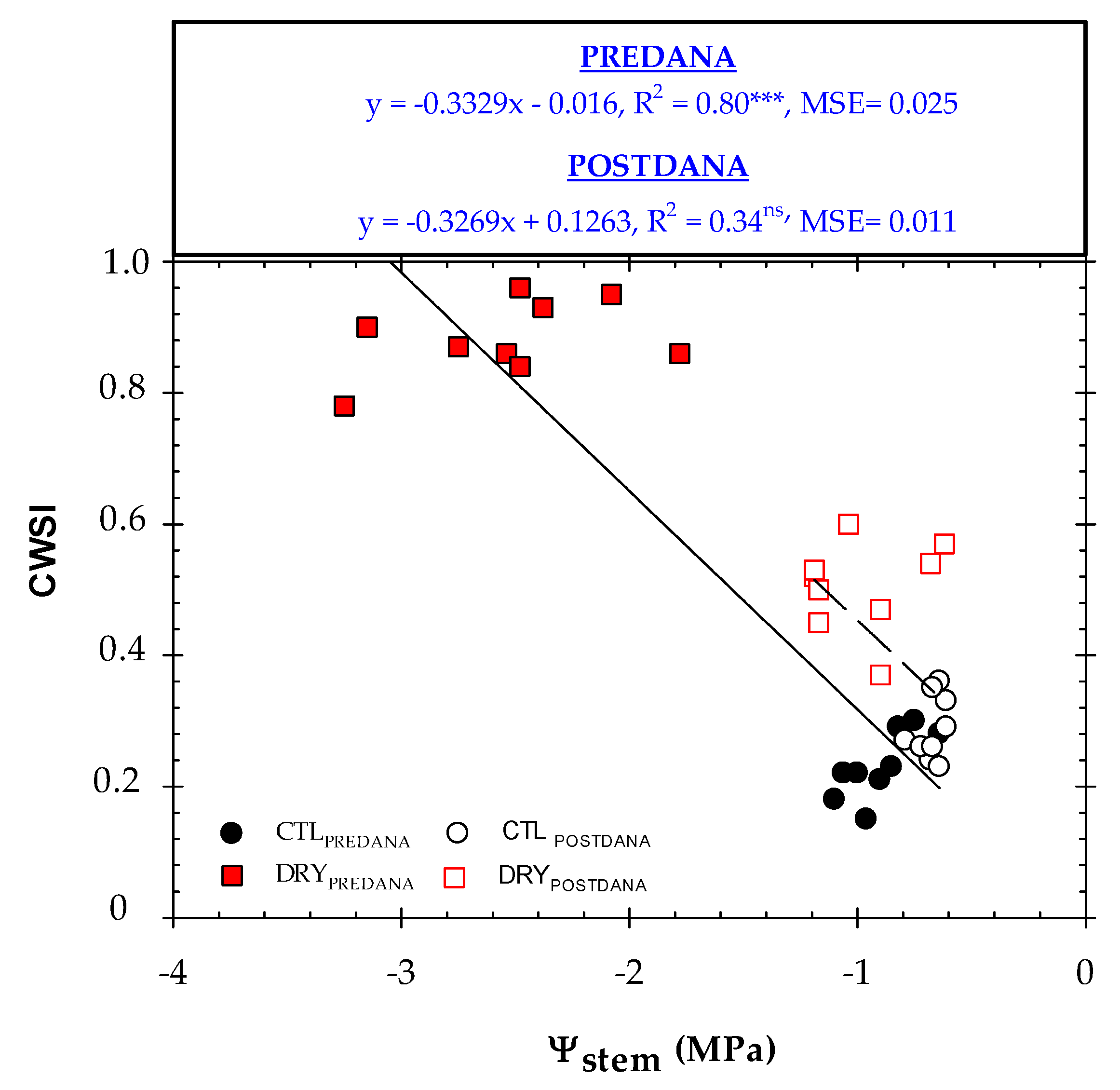
2.4. Thermal Indexes vs. Plant Water Status
3. Materials and Methods
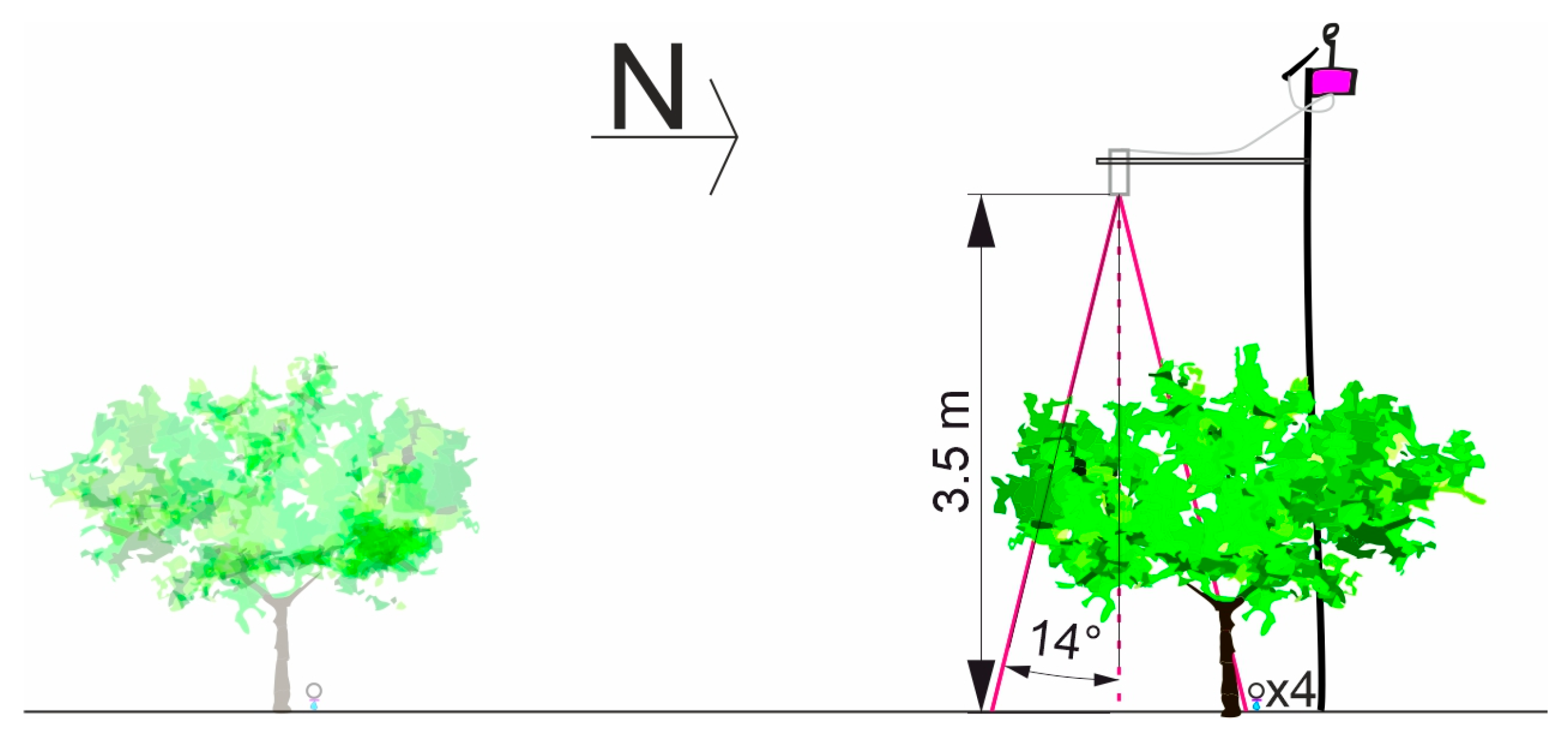
3.1. Experimental Conditions and DANA Event
3.2. Irrigation Treatments
- -
- Full-irrigated treatment (Control, CTL): during the whole season trees were irrigated based on 100% of the crop evapotranspiration (ETc) to ensure non-limiting soil water conditions. The ETc was estimated following the FAO approach by multiplying the crop reference evapotranspiration (ET0), using the Penman–Monteith equation [55], by the crop coefficients (Kc) obtained by [56] at the same location for Prunus persica sp. Irrigation was scheduled weekly, and water was applied daily during the night as needed.
- -
- Non-irrigated treatment (DRY): irrigation was suspended from June to November (DOY 158–329), coinciding with the postharvest period of our early-maturing cultivar. Before and after these dates, irrigation rates were the same as for the CTL treatment.
3.3. Continuous Measurements
- (i)
- Tc − Ta: the difference between the canopy and the surrounding air temperature.
- (ii)
- CWSI (Crop Water Stress Index): calculated empirically according to the equation described by Idso et al. [11]:
- (i)
- Tc’ − Tc: the difference between the canopy temperature of DRY and CTL treatments, respectively.
- (ii)
- ΣTc − Ta: the daily integral thermal index for daylight values of Tc − Ta > 0.
- (iii)
- ΣTc’ − Tc: the daily integral thermal index for daylight values of Tc − ‘Tc > 0
3.4. Discrete Measurements
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Michaelides, S.; Karacostas, T.; Sánchez, J.; Retalis, A.; Pytharoulis, I.; Homar, V.; Romero, R.; Zanis, P.; Giannakopoulos, C.; Bühl, J.; et al. Reviews and perspectives of high impact atmospheric processes in the Mediterranean. Atmos. Res. 2018, 208, 4–44. [Google Scholar] [CrossRef]
- Pytharoulis, I. Analysis of a Mediterranean tropical-like cyclone and its sensitivity to the sea surface temperatures. Atmos. Res. 2018, 208, 167–179. [Google Scholar] [CrossRef]
- García-Ayllon, S.; Radke, J. Geostatistical analysis of the spatial correlation between territorial anthropization and flooding vulnerability: Application to the DANA phenomenon in a Mediterranean watershed. Appl. Sci. 2021, 11, 809. [Google Scholar] [CrossRef]
- Cuenca Hidrográfica del Segura (CHS). Available online: www.chsegura.es (accessed on 12 December 2021).
- Paxian, A.; Hertig, E.; Seubert, S.; Vogt, G.; Jacobeit, J.; Paeth, H. Present-day and future Mediterranean precipitation extremes assessed by different statistical approaches. Clim. Dyn. 2015, 44, 845–860. [Google Scholar] [CrossRef]
- Chien, S.; Tanpipat, V. Remote Sensing of Natural Disasters. In Earth System Monitoring; Orcutt, J., Ed.; Springer: New York, NY, USA, 2013; pp. 1–517. [Google Scholar] [CrossRef]
- Munawar, H.S.; Hammad, A.W.A.; Waller, S.T. Remote Sensing Methods for Flood Prediction: A Review. Sensors 2022, 22, 960. [Google Scholar] [CrossRef]
- Blaya-Ros, P.J.; Blanco, V.; Domingo, R.; Soto-Valles, F.; Torres-Sánchez, R. Feasibility of low-cost thermal imaging for monitoring water stress in young and mature sweet cherry trees. Appl. Sci. 2020, 10, 5461. [Google Scholar] [CrossRef]
- Jones, H.G. Application of Thermal Imaging and Infrared Sensing in Plant Physiology and Ecophysiology. Adv. Plant Ecophysiol. Tech. 2004, 41, 107–163. [Google Scholar] [CrossRef]
- Mira-García, A.B.; Conejero, W.; Vera, J.; Ruiz-Sánchez, M.C. Water status and thermal response of lime trees to irrigation and shade screen. Agric. Water Manag. 2022, 272, 107843. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, D.; Gartung, J.L. Influence of irrigation scheduling using thermometry on peach tree water status and yield under different irrigation systems. Agronomy 2007, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Idso, S.B.; Jackson, R.D.; Pinter, P.J.; Reginato, R.J.; Hatfield, J.L. Normalizing the stress-degree-day parameter for environmental variability. Agric. Meteorol. 1981, 24, 45–55. [Google Scholar] [CrossRef]
- Jackson, R.D.; Idso, S.B.; Reginato, R.J.; Pinter, P.J. Canopy temperature as a crop water stress indicator. Water Resour. Res. 1981, 17, 1133–1138. [Google Scholar] [CrossRef]
- Fuchs, M.; Tanner, C.B. Infrared thermometry of vegetation. Agron. J. 1966, 58, 297–601. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Rubio, A.E.; Viñuela, I.; Hernández, A.; Gutiérrez-Gordillo, S.; Rodríguez-Pleguezuelo, C.R.; Durán-Zuazo, V.H. Thermal imaging at plant level to assess the crop-water status in almond trees (cv. Guara) under deficit irrigation strategies. Agric. Water Manag. 2018, 208, 176–186. [Google Scholar] [CrossRef]
- González-Dugo, V.; Lopez-Lopez, M.; Espadafor, M.; Orgaz, F.; Testi, L.; Zarco-Tejada, P.; Lorite, I.J.; Fereres, E. Transpiration from canopy temperature: Implications for the assessment of crop yield in almond orchards. Eur. J. Agron. 2019, 105, 78–85. [Google Scholar] [CrossRef]
- Wang, D.; Gartung, J. Infrared canopy temperature of early-ripening peach trees under postharvest deficit irrigation. Agric. Water Manag. 2010, 97, 1787–1794. [Google Scholar] [CrossRef]
- Bellvert, J.; Marsal, J.; Girona, J.; Gonzalez-Dugo, V.; Fereres, E.; Ustin, S.L.; Zarco-Tejada, P.J. Airborne thermal imagery to detect the seasonal evolution of crop water status in peach, nectarine and Saturn peach orchards. Remote Sens. 2016, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Giuliani, R.; Flore, J.A. Potential use of infra-red thermometry for the detection of water stress in apple trees. Acta Hortic. 2000, 537, 383–392. [Google Scholar] [CrossRef]
- Testi, L.; Goldhamer, D.; Iniesta, F.; Salinas, M. Crop water stress index is a sensitive water stress indicator in pistachio trees. Irrig. Sci. 2008, 26, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Lima, R.S.N.; García-Tejero, I.; Lopes, T.S.; Costa, J.M.; Vaz, M.; Durán-Zuazo, V.H.; Chaves, M.; Glenn, D.M.; Campostrini, E. Linking thermal imaging to physiological indicators in Carica papaya L. under different watering regimes. Agric. Water Manag. 2016, 164, 148–157. [Google Scholar] [CrossRef]
- Conesa, M.R.; Conejero, W.; Vera, J.; Ruiz-Sánchez, M.C. Effects of postharvest water deficits on the physiological behavior of early-maturing nectarine trees. Plants 2020, 9, 1104. [Google Scholar] [CrossRef]
- McCutchan, H.; Shackel, K. Stem-water potential as a sensitive indicator of water stress in prune trees Prunus domestica L. cv. French). J. Am. Soc. Hortic. Sci. 1992, 117, 607–611. [Google Scholar] [CrossRef] [Green Version]
- Vera, J.; Conejero, W.; Conesa, M.R.; Ruiz-Sánchez, M.C. Irrigation factor approach based on soil water content: A nectarine orchard case study. Water 2019, 11, 589. [Google Scholar] [CrossRef] [Green Version]
- Mira-García, A.B.; Vera, J.; Conejero, W.; Conesa, M.R.; Ruiz-Sánchez, M.C. Evapotranspiration in young lime trees with automated irrigation. Sci. Hortic. 2021, 288, 110396. [Google Scholar] [CrossRef]
- Vera, J.; Conejero, W.; Mira-García, A.B.; Conesa, M.R.; Ruiz-Sánchez, M.C. Towards irrigation automation based on dielectric soil sensors. J. Hortic. Sci. Biotech. 2021, 96, 696–707. [Google Scholar] [CrossRef]
- Abrisqueta, I.; Conejero, W.; Valdés-Vela, M.; Vera, J.; Ortuño, M.F.; Ruiz-Sánchez, M.C. Stem water potential estimation of drip-irrigated early-maturing peach trees under Mediterranean conditions. Comput. Electron. Agric. 2015, 114, 7–13. [Google Scholar] [CrossRef]
- Rahmati, M.; Davarynejad, G.H.; Génard, M.; Bannayan, M.; Azizi, M.; Vercambre, G. Peach water relations, gas exchange, growth and shoot mortality under water deficit in semi-arid weather conditions. PLoS ONE 2015, 10, e0120246. [Google Scholar] [CrossRef] [Green Version]
- De la Rosa, J.M.; Conesa, M.R.; Domingo, R.; Aguayo, E.; Falagán, E.; Pérez-Pastor, A. Combined effects of deficit irrigation and crop level on early nectarine trees. Agric. Water Manag. 2016, 170, 120–132. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Cuesta, J.M.; Ortuño, M.F.; González-Dugo, V.; Zarco-Tejada, P.J.; Parra, M.; Rubio-Asensio, J.S.; Intrigliolo, D.S. Assessment of peach trees water status and leaf gas exchange using on-the-ground versus airborne-based thermal imagery. Agric. Water Manag. 2022, 267, 107628. [Google Scholar] [CrossRef]
- Girona, J.; Gelly, M.; Mata, M.; Arbonés, A.; Rufat, J.; Marsal, J. Peach tree response to single and combined deficit irrigation regimes in deep soils. Agric. Water Manag. 2005, 72, 97–108. [Google Scholar] [CrossRef]
- Naor, A.; Stern, R.; Flaishman, M.; Gal, Y.; Peres, M. Effects pf post-harvest water stress on autumnal bloom and subsequent-season productivity in mid-season ‘Spadona’ pear. J. Hortic. Sci. Biotech. 2006, 81, 3651–3700. [Google Scholar] [CrossRef]
- Shackel, K.; Lampinen, B.; Sibbett, S.; Olson, W. The relation of midday stem water potential to the growth and physiology of fruit trees under water limited conditions. Acta Hortic. 2000, 537, 425–430. [Google Scholar] [CrossRef]
- Álvarez, S.; Navarro, A.; Nicolás, E.; Sánchez-Blanco, M.J. Transpiration, photosynthetic responses, tissue water relations and dry mass partitioning in Callistemon plants during drought conditions. Sci. Hortic. 2011, 129, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Chaves, M.M. Effects of water deficits on carbon assimilation. J. Exp. Bot. 1991, 42, 1–16. [Google Scholar] [CrossRef]
- Gulías, J.; Cifre, J.; Jonasson, S.; Medrano, H.; Flexas, J. Seasonal and inter-annual variations of gas exchange in thirteen woody species along a climatic gradient in the Mediterranean island of Mallorca. Flora Morphol. Distrib. Funct. Ecol. Plants 2009, 204, 169–181. [Google Scholar] [CrossRef]
- Delucia, E.H. Effect of low root temperature on net photosynthesis, stomatal conductance and carbohydrate concentration in Engelmann soruce (Picea engelmannii Parry ex Engelm.) seedlings. Tree Physiol. 1986, 2, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Pineda, M.; Barón, M.; Pérez-Bueno, M.L. Thermal Imaging for Plant Stress Detection and Phenotyping. Remote Sens. 2021, 13, 68. [Google Scholar] [CrossRef]
- González-Dugo, M.P.; Moran, M.S.; Mateos, L.; Bryant, R. Canopy temperature variability as an indicator of crop water stress severity. Irrig. Sci. 2006, 24, 233–240. [Google Scholar] [CrossRef]
- Giménez-Gallego, J.; González-Teruel, J.D.; Soto-Valles, F.; Jiménez-Buendía, M.; Navarro-Hellín, H.; Torres-Sánchez, R. Intelligent thermal image-based sensor for affordable measurement of crop canopy temperature. Comput. Electron. Agric. 2021, 188, 106319. [Google Scholar] [CrossRef]
- Massai, R.; Remorini, D.; Casula, F. Leaf temperature measured on peach trees growing in different climatic and soil water conditions. Acta Hortic. 2000, 537, 399–406. [Google Scholar] [CrossRef]
- Egea, G.; Padilla-Díaz, C.M.; Martinez-Guanter, J.; Fernández, J.E.; Pérez-Ruiz, M. Assessing a crop water stress index derived from aerial thermal imaging and infrared thermometry in super-high-density olive orchards. Agric. Water Manag. 2017, 187, 210–221. [Google Scholar] [CrossRef] [Green Version]
- Remorini, D.; Massai, R. Comparison of water status indicators for young peach trees. Irrig. Sci. 2003, 22, 39–46. [Google Scholar] [CrossRef]
- González-Dugo, V.; Zarco-Tejada, P.J.; Fereres, E. Applicability and limitations of using the crop water stress index as an indicator of water deficits in citrus orchards. Agric. For. Meteorol. 2014, 198, 94–104. [Google Scholar] [CrossRef]
- Yang, M.; Gao, P.; Zhou, P.; Xie, J.; Sun, D.; Han, X.; Wang, W. Simulating canopy temperature using a random forest model to calculate the crop water stress index of chinese brassica. Agronomy 2021, 11, 2244. [Google Scholar] [CrossRef]
- Romero-Trigueros, C.; Bayona, J.M.; Nortes, P.A.; Alarcón, J.J.; Nicolás, E. Determination of crop water stress index by infrared thermometry in grapefruit trees irrigated with saline reclaimed water combined with deficit irrigation. Remote Sens. 2019, 11, 757. [Google Scholar] [CrossRef] [Green Version]
- Agam, N.; Cohen, Y.; Berni, J.A.J.; Alchanatis, V.; Kool, D.; Dag, A.; Yermiyahu, U.; Ben-Gal, A. An insight to the performance of crop water stress index for olive trees. Agric. Water Manag. 2013, 118, 79–86. [Google Scholar] [CrossRef]
- Jackson, R.D. Canopy Temperature and Crop Water Stress, Advances in Irrigation; Academic Press, Inc.: Cambridge, MA, USA, 1982; pp. 43–85. [Google Scholar] [CrossRef]
- Maes, W.H.; Steppe, K. Estimating evapotranspiration and drought stress with ground-based thermal remote sensing in agriculture: A review. J. Exp. Bot. 2012, 63, 4671–4712. [Google Scholar] [CrossRef] [Green Version]
- Conesa, M.R.; Conejero, W.; Vera, J.; Ramírez-Cuesta, J.M.; Ruiz-Sánchez, M.C. Terrestrial and remote indexes to assess moderate deficit irrigation in early-maturing nectarine trees. Agronomy 2019, 9, 630. [Google Scholar] [CrossRef] [Green Version]
- López-López, M.; Espadafor, M.; Testi, L.; Lorite, I.J.; Orgaz, F.; Fereres, E. Water use of irrigated almond trees when subjected to water deficits. Agric. Water Manag. 2018, 195, 84–93. [Google Scholar] [CrossRef]
- Saxton, K.E.; Rawls, W.J.; Romberger, J.S.; Papendick, R.I. Estimating generalized soil-water characteristics from texture. Soil Sci. Soc. Am. J. 1986, 50, 1031–1035. [Google Scholar] [CrossRef]
- Vera, J.; de la Peña, J.M. FERTIGA: Programa de Fertirrigación de Frutales; CEBAS-CSIC: Murcia, Spain, 1994; p. 69. [Google Scholar]
- Mounzer, O.; Conejero, W.; Nicolás, E.; Abrisqueta, I.; García-Orellana, Y.; Tapia, L.; Vera, J.; Abrisqueta, J.M.; Ruiz-Sánchez, M.C. Growth pattern and phenological stages of early-maturing peach trees under a Mediterranean climate. HortScience 2008, 43, 1813–1818. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.G.; Pereira, J.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guide-Lines for Computing Crop Water Requirements; FAO Irrigation and DrainagePaper; Food and Agriculture Organization of the United Nations: Rome, Italy, 1998; Volume 56, p. 300. [Google Scholar]
- Abrisqueta, I.; Abrisqueta, J.; Tapia, L.M.; Munguía, J.; Conejero, W.; Vera, J.; Ruiz-Sánchez, M.C. Basal crop coefficients for early-season peach trees. Agric. Water Manag. 2013, 121, 158–163. [Google Scholar] [CrossRef]
- Balsalobre, N.; Koptsyukh, E.; Ruiz-Sánchez, M.C.; Vera, J.; Conejero, W.; Nicolás, M.J. Riego deficitario y raíces de nectarinos. In Proceedings of the V Congreso IDIES “I+D en Institutos de Enseñanza Secundaria”, Murcia, Spain, 26 June 2018; ISBN 978-84-09-03063-7. [Google Scholar]
- Abrisqueta, I.; Vera, J.; Tapia, L.M.; Abrisqueta, J.M.; Ruiz-Sánchez, M.C. Soil water content criteria for peach trees water stress detection during the postharvest period. Agric. Water Manag. 2012, 104, 62–67. [Google Scholar] [CrossRef]
- Evett, S.R.; Tolk, J.A.; Howell, T.A. Soil profile water content determination: Sensor accuracy, axial response, calibration, temperature, dependence and precision. Vadose Zone J. 2006, 5, 894–907. [Google Scholar] [CrossRef]
- Abrisqueta, I.; Conejero, W.; López-Martínez, L.; Vera, J.; Ruiz-Sánchez, M.C. Root and aerial growth in early-maturing peach trees under two crop load treatments. Span. J. Agric. Res. 2017, 15, e0803. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, T.C. Measurement of tree water status. In Irrigation of Agricultural Crops; Steward, B.A., Nielsen, D.R., Eds.; Agronomy Monograph No. 30; American Society of Agronomy: Madison, WI, USA, 1990; pp. 243–279. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tc − Ta vs. VPD | TOTAL | PRE-DANA | POST-DANA | ||||
|---|---|---|---|---|---|---|---|
| CTL Treatment | TIME (h) (GTM + 0) | R2 | p-Value | R2 | p-Value | R2 | p-Value |
| 08:00–10:00 | 0.45 | *** | 0.72 | *** | 0.84 | *** | |
| 10:00–12:00 | 0.20 | ns | 0.19 | ns | 0.79 | *** | |
| 12:00–14:00 | 0.09 | ns | 0.14 | ns | 0.82 | *** | |
| 14:00–16:00 | 0.00 | ns | 0.03 | ns | 0.72 | *** | |
| 16:00–18:00 | 0.18 | ns | 0.07 | ns | 0.27 | ns | |
| 18:00–20:00 | 0.02 | ns | 0.01 | ns | 0.25 | ns | |
| Average | 0.01 | ns | 0.04 | ns | 0.01 | ns | |
| DRY treatment | |||||||
| 08:00–10:00 | 0.35 | *** | 0.14 | ns | 0.05 | ns | |
| 10:00–12:00 | 0.43 | *** | 0.20 | ns | 0.05 | ns | |
| 12:00–14:00 | 0.33 | *** | 0.04 | ns | 0.10 | ns | |
| 14:00–16:00 | 0.33 | *** | 0.15 | ns | 0.02 | ns | |
| 16:00–18:00 | 0.41 | *** | 0.40 | ** | 0.11 | ns | |
| 18:00–20:00 | 0.08 | ns | 0.09 | ns | 0.36 | ** | |
| Average | 0.39 | *** | 0.26 | * | 0.36 | ** | |
| DRY–CTL treatments | Tc’ − Tc vs. VPD | ||||||
| 08:00–10:00 | 0.80 | *** | 0.77 | *** | 0.85 | *** | |
| 10:00–12:00 | 0.74 | *** | 0.56 | *** | 0.84 | *** | |
| 12:00–14:00 | 0.56 | *** | 0.15 | ns | 0.83 | *** | |
| 14:00–16:00 | 0.46 | *** | 0.12 | ns | 0.74 | *** | |
| 16:00–18:00 | 0.20 | ns | 0.26 | ns | 0.33 | * | |
| 18:00–20:00 | 0.00 | ns | 0.01 | ns | 0.61 | *** | |
| Average | 0.52 | *** | 0.31 | ** | 0.45 | *** | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conesa, M.R.; Conejero, W.; Vera, J.; Mira-García, A.B.; Ruiz-Sánchez, M.C. Impact of a DANA Event on the Thermal Response of Nectarine Trees. Plants 2023, 12, 907. https://doi.org/10.3390/plants12040907
Conesa MR, Conejero W, Vera J, Mira-García AB, Ruiz-Sánchez MC. Impact of a DANA Event on the Thermal Response of Nectarine Trees. Plants. 2023; 12(4):907. https://doi.org/10.3390/plants12040907
Chicago/Turabian StyleConesa, María R., Wenceslao Conejero, Juan Vera, Ana Belén Mira-García, and María Carmen Ruiz-Sánchez. 2023. "Impact of a DANA Event on the Thermal Response of Nectarine Trees" Plants 12, no. 4: 907. https://doi.org/10.3390/plants12040907
APA StyleConesa, M. R., Conejero, W., Vera, J., Mira-García, A. B., & Ruiz-Sánchez, M. C. (2023). Impact of a DANA Event on the Thermal Response of Nectarine Trees. Plants, 12(4), 907. https://doi.org/10.3390/plants12040907