

Encephalartos natalensis, Their Nutrient-Cycling Microbes and Enzymes: A Story of Successful Trade-Offs


Abstract
:1. Introduction
2. Results
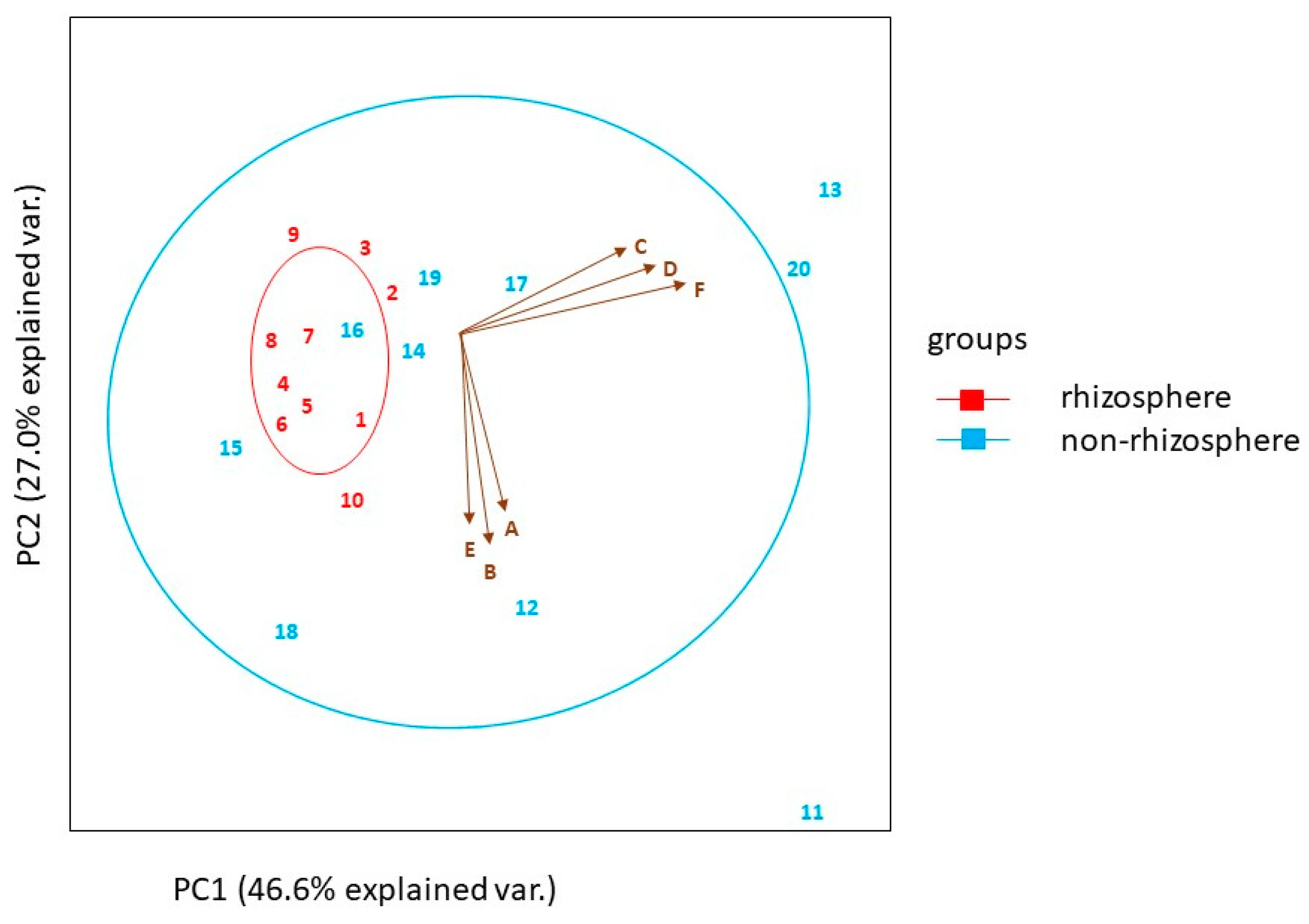
2.1. Soil Characteristics
Soil Nutrients
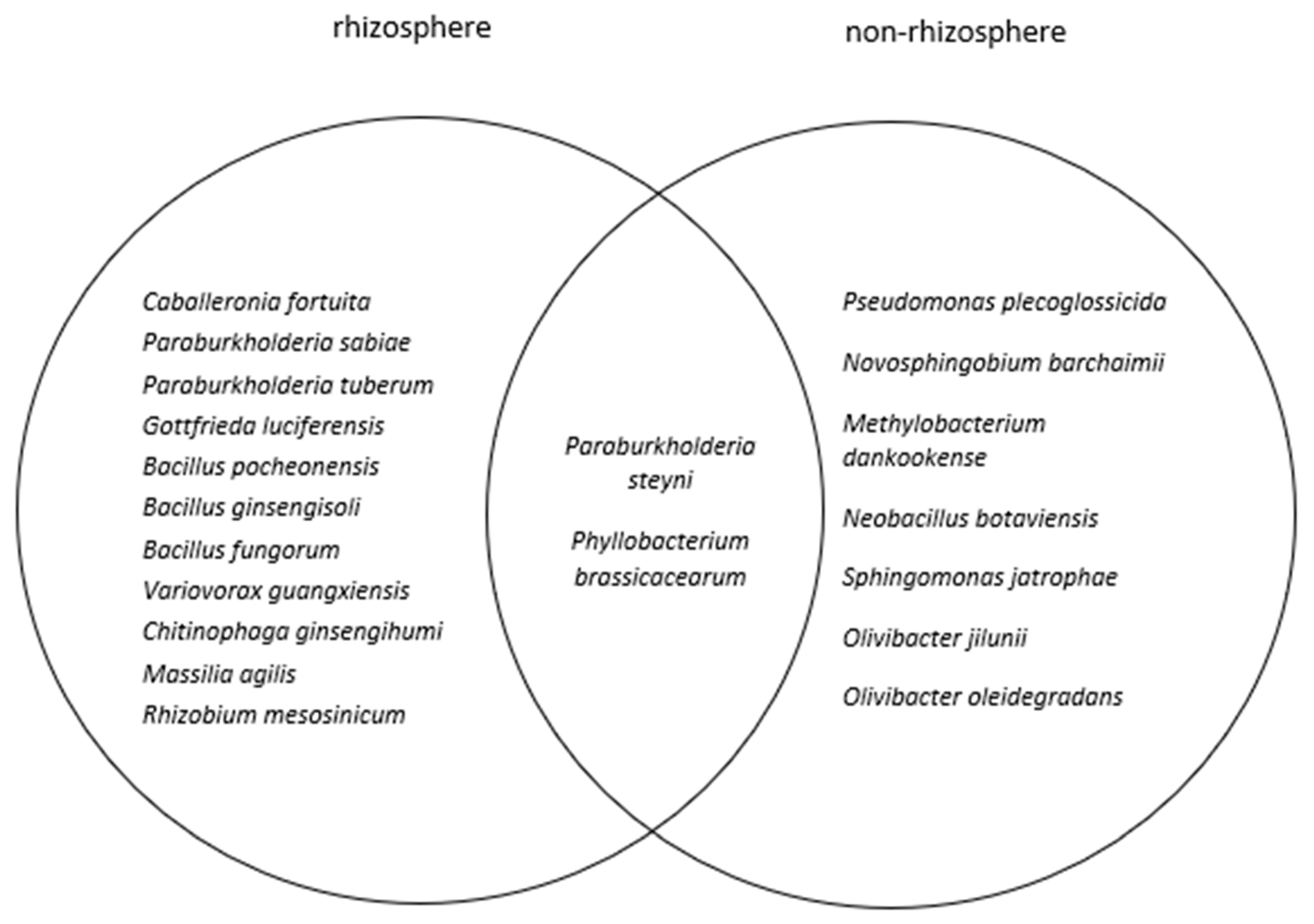
2.2. Bacterial Identification
2.2.1. Bacterial Identification of E. natalensis Coralloid Roots
2.2.2. Bacterial Identification of E. natalensis Rhizosphere
2.2.3. Bacterial Identification of E. natalensis Non-Rhizosphere Control Soils
2.3. Soil Enzyme Activities
3. Discussion
4. Materials and Methods
4.1. Study Sites and Target Species
4.2. Soil Sampling and Soil Nutrition Analysis
4.3. Soil Serial Dilutions and Bacterial Extraction
4.4. Coralloid Roots Surface Sterilization and Bacterial Extraction
4.5. Coralloid Roots, Rhizosphere and Non-Rhizosphere Soils Bacterial Amplification, Sequencing, and Identification
4.6. Soil Enzymatic Studies
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brenner, E.D.; Stevenson, D.W.; Twigg, R.W. Cycads: Evolutionary innovations and the role of plant-derived neurotoxins. Trends Plant Sci. 2003, 8, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Mamay, S.H. Cycads: Fossil evidence of late paleozoic origin. Science 1969, 164, 295–296. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, J.S. (Ed.) Regional overview: Africa. In Status Survey and Conservation Action Plan, Cycad; IUCN/SSC Cycad Specialist Group: Cambridge, UK; The World Conservation Union: Fontainebleau, France, 2003. [Google Scholar]
- Ma, Y.; Jiang, H.; Wang, B.; Zhou, G.; Yu, S.; Peng, S.; Hao, Y.; Wei, X.; Liu, J.; Yu, Z. Carbon Storage of cycad and other gymnosperm ecosystems in China: Implications to evolutionary trends. Pol. J. Ecol. 2009, 57, 635–646. [Google Scholar]
- Alvarez-Yepiz, J.C.; Cueva, A.; Dovciak, M.; Teece, M.; Enrico, A.; Yepez, E.A. Ontogenic resource-use strategies in a rare long-lived cycad along environmental gradients. Conserv. Physiol. 2014, 2, 2–4. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Yepiz, J.C.; Dovčiak, M. Enhancing ecosystem function through conservation: Threatened plants increase local carbon storage in tropical dry forests. Trop. Conserv. Sci. 2015, 8, 999–1008. [Google Scholar] [CrossRef] [Green Version]
- Berkelmann, D.; Schneider, D.; Engelhaupt, M.; Heinemann, M.; Christel, S.; Wijayanti, M. How rainforest conversion to agricultural systems in Sumatra (Indonesia) affects active soil bacterial communities. Front. Microbiol. 2018, 9, 2381. [Google Scholar] [CrossRef] [Green Version]
- Hill, K.D.; Chase, M.W.; Stevenson, D.W.; Hills, H.G.; Schutzman, B. The families and genera of cycads: A molecular phylogenetic analysis of Cycadophyta based on nuclear and plastid DNA sequences. Int. J. Plant Sci. 2003, 164, 933–948. [Google Scholar] [CrossRef]
- Rousseau, P. A Molecular Systematics Study of the African Endemic Cycads. Master’s Thesis, Department of Botany and Plant Biotechnology, University of Johannesburg, Johannesburg, South Africa, 2012. [Google Scholar]
- Mankga, L.T.; Yessoufou, K. Factors driving the global decline of cycad diversity. AoB Plants 2017, 9, plx022. [Google Scholar] [CrossRef]
- Donaldson, J.S. South African Encephalartos species. Non-detrimental findings workshop case studies on succulents and cycads, study 4 on Encephalartos. In NDF Workshop Case Studies: Case Study; Elsevier: Amsterdam, The Netherlands, 2008; Volume 4. [Google Scholar]
- Bamigboye, S.O. Acceleration of cycads diversification towards the end of Pliocene in Africa. J. Ecosyst. Ecography 2013, 3, 4. [Google Scholar]
- Costa, J.L.; Paulsrud, P.; Lindblad, P. Cyanobiont diversity within coralloid roots of selected cycad species. FEMS Microbiol. Ecol. 1999, 28, 85–91. [Google Scholar] [CrossRef]
- Lobakova, E.; Orazova, M.K.; Dobrovolskaya, T. The structure of cyanobacterial communities formed during the degradation of apogeotropic roots of cycads. Microbiology 2003, 72, 634–637. [Google Scholar] [CrossRef]
- Gehringer, M.M.; Pengelly, J.J.; Cuddy, W.S.; Fieker, C.; Forster, P.I.; Neilan, B.A. Host selection of symbiotic cyanobacteria in 31 species of the Australian cycad genus: Macrozamia (Zamiaceae). Mol. Plant-Microbe Interact. 2010, 23, 811–822. [Google Scholar] [CrossRef] [Green Version]
- Surez-Moo, P.D.J.; Vovides, A.P.; Griffith, M.P.; Barona-Gómez, F.; Cibrián-Jaramillo, A. Unlocking a high bacterial diversity in the coralloid root microbiome from the cycad genus Dioon. PLoS ONE 2019, 14, e0211271. [Google Scholar] [CrossRef] [Green Version]
- Sasse, J.; Martinoia, E.; Northern, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Warshan, D. Cyanobacteria in Symbiosis with Boreal Forest Feathermosses: From Genome Evolution and Gene Regulation to Impact on the Ecosystem. Ph.D. Thesis, Stockholm University, Stockholm, Sweden, 2017. [Google Scholar]
- Lucas, N.; Bienaime, C.; Belloy, C.; Queneudec, M.; Silvestre, F.; Nava-Saucedo, J.E. Polymer biodegradation: Mechanisms and estimation techniques—A review. Chemosphere 2008, 73, 429–442. [Google Scholar] [CrossRef]
- Hill, J.M.; Roberts, J.; Loeb, E.; Khan, A.; MacLellan, A.; Hill, R.W. L-asparaginase therapy for leukemia and other malignant neoplasms: Remission in human leukemia. JAMA 1967, 202, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Mega, T.; Ikenaka, T.; Matsushima, Y. Studies on N-acetyl-β-D-glucosaminidase of Aspergillus oryzae: II. Substrate specificity of the enzyme. J. Biochem. 1972, 71, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Turner, B.L.; Baxter, R.; Whitton, B.A. Seasonal phosphatase activity in three characteristic soils of the English uplands polluted by long-term atmospheric nitrogen deposition. Environ. Pollut. 2002, 120, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Henriksson, G.; Sild, V.; Szabó, I.G.; Pettersson, G.; Johansson, G. Substrate specificity of cellobiose dehydrogenase from Phanerochaete chrysosporium. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1998, 1383, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Lindström, K.; Mousavi, S.A. Rhizobium and Other N fixing Symbioses. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Chichester, UK, 2010. [Google Scholar]
- Santi, C.; Bogusz, D.; Franche, C. Biological nitrogen fixation in non-legume plants. Ann. Bot. 2013, 111, 743–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raimondo, D.; von Staden, L.; Foden, W.; Victor, J.E.; Helme, N.A.; Turner, R.C.; Kamundi, D.A.; Manyama, P.A. Red List of South African Plants. Strelitzia 25; South African National Biodiversity Institute: Pretoria, South Africa, 2009. [Google Scholar]
- Rasheed, M.; Naseer, T.; Hassan, A.; Hassan, F.; Hayat, R.; Jilani, G.; Vaseer, S.G.; Ali, M.B. Isolation of nodule associated bacteria for promotion of lentil growth. Pak. J. Agric. Res. 2020, 33, 170–179. [Google Scholar] [CrossRef]
- Magadlela, A.; Beukes, C.; Venter, F.; Steenkamp, E.; Valentine, A. Does P deficiency affect nodule bacterial composition and N source utilization in a legume from nutrient-poor Mediterranean-type ecosystems? Soil Biol. Biochem. 2017, 104, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Matiwane, S.E.; Aremu, A.O.; Valentine, A.J.; Magadlela, A. Nutritional status of KwaZulu-Natal soils affects microbe symbiosis, nitrogen utilization and growth of Vigna radiata (L.) R. Walczak. S. Afr. J. Bot. 2019, 126, 115–120. [Google Scholar] [CrossRef]
- Manson, A.D.; Roberts, V.G. Analytical Methods Used by the Soil Fertility and Analytical Services Section; Republic of South Africa: Pietermaritzburg, South Africa, 2000.
- Ahsan, N.; Marian, M.; Suga, H.; Shimizu, M. Lysinibacillus xylanilyticus strain GIC41 as a potential plant biostimulant. Microbes Environ. 2021, 36, ME21047. [Google Scholar] [CrossRef] [PubMed]
- Jyolsna, K.S.; Bharathi, N.; Ali, L.R.; Paari, K.A. Impact of Lysinibacillus macroides, a potential plant growth promoting rhizobacteria on growth, yield and nutritional value of tomato Plant (Solanum lycopersicum L. f1 hybrid Sachriya). Plant Sci. Today 2021, 8, 365–372. [Google Scholar] [CrossRef]
- Jackson, C.R.; Tyler, H.L.; Millar, J.J. Determination of microbial extracellular enzyme activity in waters, soils, and sediments using high throughput microplate assays. J. Vis. Exp. 2013, 80, e50399. [Google Scholar]
- Bruckner, A.; Wright, J.; Kampichler, C.; Bauer, R.; Kandeler, E. A method of preparing mesocosms for assessing complex biotic processes in soils. Biol. Fertil. Soils 1995, 19, 257–262. [Google Scholar] [CrossRef]
- Hernandez, J.P.; de-Bashan, L.E.; Rodriguez, D.J.; Rodriguez, Y.; Bashan, Y. Growth promotion of the freshwater microalga Chlorella vulgaris by the nitrogen-fixing, plant growth-promoting bacterium Bacillus pumilus from arid zone soils. Eur. J. Soil Biol. 2009, 45, 88–93. [Google Scholar] [CrossRef]
- Hashem, A.; Tabassum, B.; Abd_Allah, E.F. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic al stress. Saudi J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef]
- Grady, E.N.; MacDonald, J.; Liu, L.; Richman, A.; Yuan, Z.C. Current knowledge and perspectives of Paenibacillus: A review. Microb. Cell Factories 2016, 15, 203. [Google Scholar] [CrossRef] [Green Version]
- von der Weid, I.; Duarte, G.F.; van Elsas, J.D.; Seldin, L. Paenibacillus brasilensis sp. nov., a novel nitrogen-fixing species isolated from the maize rhizosphere in Brazil. Int. J. Syst. Evol. Microbiol. 2002, 52, 2147–2153. [Google Scholar] [PubMed] [Green Version]
- Mayer, E.; Dörr de Quadros, P.; Fulthorpe, R. Plantibacter flavus, Curtobacterium herbarum, Paenibacillus taichungensis, and Rhizobium selenitireducens endophytes provide host-specific growth promotion of Arabidopsis thaliana, basil, lettuce, and bok choy plants. Appl. Environ. Microbiol. 2019, 85, e00383-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oggerin, M.; Arahal, D.R.; Rubio, V.; Marín, I. Identification of Beijerinckia fluminensis strains CIP 106281T and UQM 1685T as Rhizobium radiobacter strains, and proposal of Beijerinckia doebereinerae sp. nov. to accommodate Beijerinckia fluminensis LMG 2819. Int. J. Syst. Evol. Microbiol. 2009, 59, 2323–2328. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Oger, P. Caballeronia mineralivorans sp. nov., isolated from oak-Scleroderma citrinum mycorrhizosphere. Syst. Appl. Microbiol. 2017, 40, 345–351. [Google Scholar] [CrossRef]
- Hetz, S.A.; Poehlein, A.; Horn, M.A. Whole-genome sequences of two new Caballeronia strains isolated from cryoturbated peat circles of the permafrost-affected Eastern European Tundra. Microbiol. Resour. Announc. 2020, 9, e00731-20. [Google Scholar] [CrossRef]
- Guo, Y.Z.; Ding, X.M.; Yao, L.; Xu, D.M.; Zhao, Y.J.; Feng, F.Y.; Meng, J.Y. Isolation and Identification of Massilia sp. B260 and Its Effect on Seedling Raising. Biotechnol. Bull. 2019, 35, 144. [Google Scholar]
- Tapia-García, E.Y.; Arroyo-Herrera, I.; Rojas-Rojas, F.U.; Ibarra, J.A.; Vásquez-Murrieta, M.S.; Martínez-Aguilar, L.; López-Lara, I.M.; Whitman, W.B.; De Los Santos, P.E. Paraburkholderia lycopersici sp. nov., a nitrogen-fixing species isolated from rhizoplane of Lycopersicon esculentum Mill. var. Saladette in Mexico. Syst. Appl. Microbiol. 2020, 43, 126133. [Google Scholar] [CrossRef]
- Too, C.C.; Ong, K.S.; Yule, C.M.; Keller, A. Putative roles of bacteria in the carbon and nitrogen cycles in a tropical peat swamp forest. Basic Appl. Ecol. 2021, 52, 109–123. [Google Scholar] [CrossRef]
- Soares Neto, C.B.; Ribeiro, P.R.; Fernandes-Júnior, P.I.; de Andrade, L.R.; Zilli, J.E.; Mendes, I.C.; do Vale, H.M.; James, E.K.; dos Reis Junior, F.B. Paraburkholderia atlantica is the main rhizobial symbiont of Mimosa spp. in ultramafic soils in the Brazilian Cerrado biome. Plant Soil 2022, 10, 465–479. [Google Scholar] [CrossRef]
- Beneduzi, A.; Peres, D.; Vargas, L.K.; Bodanese-Zanettini, M.H.; Passaglia, L.M. Evaluation of genetic diversity and plant growth promoting activities of nitrogen-fixing bacilli isolated from rice fields in South Brazil. Appl. Soil Ecol. 2008, 39, 311–320. [Google Scholar] [CrossRef]
- Sun, B.; Bai, Z.; Bao, L.; Xue, L.; Zhang, S.; Wei, Y.; Zhang, Z.; Zhuang, G.; Zhuang, X. Bacillus subtilis biofertilizer mitigating agricultural ammonia emission and shifting soil nitrogen cycling microbiomes. Environ. Int. 2020, 144, 105989. [Google Scholar] [CrossRef] [PubMed]
- Seldin, L.; Dubnau, D. Deoxyribonucleic acid homology among Bacillus polymyxa, Bacillus macerans, Bacillus azotofixans, and other nitrogen-fixing Bacillus strains. Int. J. Syst. Evol. Microbiol. 1985, 35, 151–154. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.M.; Trinh, N.H.; Kim, J. Proposal of three novel species of soil bacteria, Variovorax ureilyticus, Variovorax rhizosphaerae, and Variovorax robiniae, in the family Comamonadaceae. J. Microbiol. 2018, 56, 485–492. [Google Scholar] [CrossRef]
- Jin, C.Z.; Jin, L.; Kang, D.H.; Liu, M.J.; Lee, J.M.; Park, D.J.; Kim, C.J. Description of desferrioxamine-producing bacterium Chitinophaga agrisoli sp. nov., isolated from soil. Antonie Van Leeuwenhoek 2021, 114, 741–750. [Google Scholar] [CrossRef]
- Anderson, C.R.; Hamonts, K.; Clough, T.J.; Condron, L.M. Biochar does not affect soil N-transformations or microbial community structure under ruminant urine patches but does alter relative proportions of nitrogen cycling bacteria. Agric. Ecosyst. Environ. 2014, 191, 63–72. [Google Scholar] [CrossRef]
- Gao, Z.H.; Ruan, S.L.; Huang, Y.X.; Lv, Y.Y.; Qiu, L.H. Paraburkholderia phosphatilytica sp. nov., a phosphate-solubilizing bacterium isolated from forest soil. Int. J. Syst. Evol. Microbiol. 2019, 69, 196–202. [Google Scholar] [CrossRef]
- Pandey, A.; Trivedi, P.; Kumar, B.; Palni, L.M. Characterization of a phosphate solubilizing and antagonistic strain of Pseudomonas putida (B0) isolated from a sub-alpine location in the Indian Central Himalaya. Curr. Microbiol. 2006, 53, 102–107. [Google Scholar] [CrossRef]
- Toyofuku, M.; Nomura, N.; Kuno, E.; Tashiro, Y.; Nakajima, T.; Uchiyama, H. Influence of the Pseudomonas quinolone signal on denitrification in Pseudomonas aeruginosa. J. Bacteriol. 2008, 190, 7947–7956. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Hernández, L.; Coutino-Megchun, J.T.; Rincon-Molina, C.I.; Ruíz-Valdiviezo, V.M.; Culebro-Ricaldi, J.M.; Cruz-Rodríguez, R.I.; Palomeque-Dominguez, H.H.; Rincón-Rosales, R. Endophytic bacteria from root nodules of Ormosia macrocalyx with potential as plant growth promoters and antifungal activity. J. Environ. Biol. 2018, 39, 997–1005. [Google Scholar] [CrossRef]
- Dourado, M.N.; Aparecida Camargo Neves, A.; Santos, D.S.; Araújo, W.L. Biotechnological and agronomic potential of endophytic, pink-pigmented methylotrophic Methylobacterium spp. BioMed Res. Int. 2015, 4, 2–5. [Google Scholar]
- Pascual, J.A.; Ros, M.; Martínez, J.; Carmona, F.; Bernabé, A.; Torres, R.; Lucena, T.; Aznar, R.; Arahal, D.R.; Fernández, F. Methylobacterium symbioticum sp. nov., a new species isolated from spores of Glomus iranicum var. tenuihypharum. Curr. Microbiol. 2020, 77, 2031–2041. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Lee, M.H.; Jeong, J.C.; Kim, D.H.; Kim, C.Y.; Kim, S.W.; Lee, J. Neobacillus endophyticus sp. nov., an endophytic bacterium isolated from Selaginella involvens roots. Int. J. Syst. Evol. Microbiol. 2021, 71, 004581. [Google Scholar] [CrossRef] [PubMed]
- Marler, T.E.; Krishnapillai, M.V. Cycas micronesica trees alter local soil traits. Forests 2018, 9, 565. [Google Scholar] [CrossRef] [Green Version]
- Marler, T.E.; Calonje, M. Two cycad species affect the carbon, nitrogen, and phosphorus content of soils. Horticulturae 2020, 6, 24. [Google Scholar] [CrossRef] [Green Version]
- Rayne, N.; Aula, L. Livestock manure and the impacts on soil health: A review. Soil Syst. 2020, 4, 64. [Google Scholar] [CrossRef]
- Ano, A.O.; Ubochi, C.I. Neutralization of soil acidity by animal manure: Mechanism of reaction. Afr. J. Biotechnol. 2007, 2007, 364–368. [Google Scholar]
- Özdoğan, D.K.; Akçelik, N.; Akçelik, M. Genetic diversity and characterization of plant growth-promoting effects of bacteria isolated from rhizospheric soils. Curr. Microbiol. 2022, 79, 132. [Google Scholar] [CrossRef]
- Bell, T.; Newman, J.A.; Silverman, B.W.; Turner, S.L.; Lilley, A.K. The contribution of species richness and composition to bacterial services. Nature 2005, 436, 1157–1160. [Google Scholar] [CrossRef]
- Glick, B.R. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Nacke, H.; Thu¨rmer, A.; Wollherr, A.; Will, C.; Hodac, L. Pyrosequencing-based assessment of bacterial community structure along different management types in German forest and grassland soils. PLoS ONE 2011, 6, e17000. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Meena, M.K.; Datta, S. Isolation, characterization of plant growth promoting bacteria from the plant Chlorophytum borivilianum and in-vitro screening for activity of nitrogen fixation, phosphate solubilization and IAA production. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 1082–1090. [Google Scholar]
- Adeniyan, O.N.; Ojo, A.O.; Akinbode, O.A.; Adediran, J.A. Comparative study of different organic manures and NPK fertilizer for improvement of soil chemical properties and dry matter yield of maize in two different soils. J. Soil Sci. Environ. Manag. 2011, 2, 9–13. [Google Scholar]
- Akinrinlola, R.J.; Yuen, G.Y.; Drijber, R.A.; Adesemoye, A.O. Evaluation of Bacillus strains for plant growth promotion and predictability of efficacy by in vitro physiological traits. Int. J. Microbiol. 2018, 5, 6–9. [Google Scholar] [CrossRef] [Green Version]
- Sunera, A.; Saqib, S.; Uddin, S.; Zaman, W.; Ullah, F.; Ayaz, A.; Asghar, M.; Rehman, S.; Munis, M.F.H. Characterization and phytostimulatory activity of bacteria isolated from tomato (Lycopersicin esculentum Mill.) rhizosphere. Microb. Pathog. 2020, 140, 103966. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.; Yeh, M.W.; Hsieh, Y.T.; Chung, W.C.; Lo, C.T.; Young, L.S. Diversity and functional characterization of bacterial endophytes dwelling in various rice (Oryza sativa L.) tissues, and their seed-borne dissemination into rhizosphere under gnotobiotic P-stress. Plant Soil 2015, 394, 177–197. [Google Scholar] [CrossRef]
- Hussain, A.; Ahmad, M.; Nafees, M.; Iqbal, Z.; Luqman, M.; Jamil, M.; Maqsood, A.; Mora-Poblete, F.; Ahmar, S.; Chen, J.T.; et al. Plant-growth-promoting Bacillus and Paenibacillus species improve the nutritional status of Triticum aestivum L. PLoS ONE 2020, 15, e0241130. [Google Scholar] [CrossRef]
- Al-Shwaiman, H.A.; Shahid, M.; Elgorban, A.M.; Siddique, K.H.; Syed, A. Beijerinckia fluminensis BFC-33, a novel multi-stress-tolerant soil bacterium: Deciphering the stress amelioration, phytopathogenic inhibition and growth promotion in Triticum aestivum (L.). Chemosphere 2022, 295, 133843. [Google Scholar] [CrossRef] [PubMed]
- Colin, Y.; Nicolitch, O.; Turpault, M.P.; Uroz, S. Mineral types and tree species determine the functional and taxonomic structures of forest soil bacterial communities. Appl. Environ. Microbiol. 2017, 83, e02684-16. [Google Scholar] [CrossRef] [Green Version]
- Nicolitch, O.; Feucherolles, M.; Churin, J.-L.; Fauchery, L.; Turpault, M.-P.; Uroz, S. A microcosm approach highlights the response of soil mineral weathering bacterial communities to an increase of K and Mg availability. Sci. Rep. 2019, 9, 14403. [Google Scholar] [CrossRef] [Green Version]
- Samuels, T.; Bryce, C.; Landenmark, H.; Marie-Loudon, C.; Nicholson, N.; Stevens, A.H. Microbial weathering of minerals and rocks in natural environments. In Biogeochemical Cycles: Ecological Drivers and Environmental Impact; Dontsova, K., Zsuzsanna, B., Le Roux, G., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA; American Geophysical Union: Washington, DC, USA, 2020; pp. 59–79. [Google Scholar]
- Sithole, N.; Pérez-Fernández, M.; Magadlela, A. Nutritional status of soils from KwaZulu-Natal modulates symbiotic interactions and plant performance in Vigna unguiculata L. (Walp). Appl. Soil Ecol. 2019, 142, 1–7. [Google Scholar] [CrossRef]
- Vyas, P.; Gulati, A. Organic acid production in vitro and plant growth promotion in maize under controlled environment by phosphate-solubilizing fluorescent Pseudomonas. BMC Microbiol. 2009, 9, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutschera, U. Plant-associated methylobacteria as co-evolved phytosymbionts: A hypothesis. Plant Signal. Behav. 2007, 2, 74–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omer, Z.S.; Tombolini, R.; Broberg, A.; Gerhardson, B. Indole-3-acetic acid production by pink-pigmented facultative methylotrophic bacteria. Plant Growth Regul. 2004, 43, 93–96. [Google Scholar] [CrossRef]
- Trotsenko, Y.A.; Ivanova, E.G.; Doronina, N.V. Aerobic methylotrophic bacteria as phytosymbionts. Mikrobiologiia 2001, 70, 623–632. [Google Scholar]
- Lee, S.H.; Kim, M.S.; Kim, J.G.; Kim, S.O. Use of soil enzymes as indicators for contaminated soil monitoring and sustainable management. Sustainability 2020, 12, 8209. [Google Scholar] [CrossRef]
- Magadlela, A.; Lembede, Z.; Egbewale, S.O.; Olaniran, A.O. The metabolic potential of soil microorganisms and enzymes in phosphorus-deficient KwaZulu-Natal grassland ecosystem soils. Appl. Soil Ecol. 2022, 181, 104647. [Google Scholar] [CrossRef]
- Rao, M.A.; Scelza, R.; Gianfreda, L. Soil enzymes. In Enzymes in Agricultural Sciences; OMICS Group eBooks: Foster City, CA, USA, 2014; Volume 2, pp. 10–24. [Google Scholar]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Fujita, K.; Kunito, T.; Matsushita, J.; Nakamura, K.; Moro, H.; Yoshida, S.; Toda, H.; Otsuka, S.; Nagaoka, K. The nitrogen supply rate regulates microbial resource allocation for the synthesis of nitrogen-acquiring enzymes. PLoS ONE 2018, 13, e0202086. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Liu, Y.; Lv, X.; Li, J.; Du, G.; Liu, L. Combinatorial pathway enzyme engineering and host engineering overcomes pyruvate overflow and enhances overproduction of N-acetylglucosamine in Bacillus subtilis. Microb. Cell Factories 2019, 18, 1. [Google Scholar] [CrossRef] [Green Version]
- Parham, J.A.; Deng, S.P. Detection, quantification, and characterization of β-glucosaminidase activity in soil. Soil Biol. Biochem. 2000, 32, 1183–1190. [Google Scholar] [CrossRef]
- Adetunji, A.T.; Lewu, F.B.; Mulidzi, R.; Ncube, B. The biological activities of β-glucosidase, phosphatase and urease as soil quality indicators: A review. J. Soil Sci. Plant Nutr. 2017, 17, 794–807. [Google Scholar] [CrossRef] [Green Version]
- Gavrilova, A.N.; Savchenka, N.I.; Shimko, N.A. Forms of phosphorus and phosphatase activity of chief soil type in the BSSR. Int. Congr. Soil Sci. 1974, 4, 281–288. [Google Scholar]
- Speir, T.W.; Ross, D.J. Soil phosphatase and sulphatase. In Soil Enzymes; Burns, R.G., Ed.; Academic Press: New York, NY, USA, 1978. [Google Scholar]
- Kitayama, K. The activities of soil and root acid phosphatase in the nine tropical rain forests that differ in phosphorus availability on Mount Kinabalu, Borneo. Plant Soil 2013, 367, 215–224. [Google Scholar] [CrossRef]
- Cenini, V.L.; Fornara, D.A.; McMullan, G.; Ternan, N.; Carolan, R.; Crawley, M.J.; Clément, J.C.; Lavorel, S. Linkages between extracellular enzyme activities and the carbon and nitrogen content of grassland soils. Soil Biol. Biochem. 2016, 96, 198–206. [Google Scholar] [CrossRef] [Green Version]
- Ekenler, M.; Tabatabai, M. β-Glucosaminidase activity of soils: Effect of cropping systems and its relationship to nitrogen mineralization. Biol. Fertil. Soils 2002, 36, 367–376. [Google Scholar] [CrossRef]
- Ekenler, M.; Tabatabai, M.A. β-Glucosaminidase activity as an index of nitrogen mineralization in soils. Commun. Soil Sci. Plant Anal. 2004, 35, 1081–1094. [Google Scholar] [CrossRef]
- Cheng, Y.; Elrys, A.S.; Merwad, A.R.; Zhang, H.; Chen, Z.; Zhang, J.; Cai, Z.; Müller, C. Global patterns and drivers of soil dissimilatory nitrate reduction to ammonium. Environ. Sci. Technol. 2022, 56, 3791–3800. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Rhizosphere | Non-Rhizosphere | |
|---|---|---|
| Primary nutrients (mg.kg−1) | ||
| Total nitrogen | 4187.01 ± 567.76 a | 3223.90 ± 487.21 b |
| Extractable phosphorus | 11.83 ± 3.66 a | 9.24 ± 0.94 a |
| Extractable potassium | 537.76 ± 50.27 a | 201.15 ± 56.66 b |
| Extractable intermediate nutrients (mg.kg−1) | ||
| Magnesium | 620.96 ± 44.48 a | 555.75 ± 85.87 a |
| Calcium | 4644.63 ± 829.93 a | 3542.67 ± 1121.89 b |
| Extractable micronutrients (mg.kg−1) | ||
| Zinc | 5.54 ± 0.54 a | 3.69 ± 0.57 b |
| Manganese | 50.84 ± 4.72 a | 75.94 ± 7.82 a |
| Copper | 74.83 ± 37.36 a | 89.77 ± 27.50 b |
| Soil relative acidity | ||
| pH | 5.75 ± 0.71 a | 5.14 ± 0.42 b |
| Exchange acidity (cmolc.kg−1) | 0.08 ± 0.07 a | 0.05 ± 0.02 a |
| Total cation exchange (cmolc.kg−1) | 29.61 ± 4.62 a | 22.62 ± 6.37 b |
| Parameter | ||
| Organic Carbon (%) | 5.24 | 4.21 |
| Family | Scientific Name | Accession Number | Similarity (%) | Function |
|---|---|---|---|---|
| Bacillaceae | Lysinibacillus xylanilyticus | NR_116698.1 | 99.93 | P solubilizing [30] N-fixing [30] |
| Lysinibacillus macrolides | NR_114920.1 | 99.06 | N-fixing [31] | |
| Lysinibacillus pakistanensis | NR_113166.1 | 99.02 | N-fixing [32] | |
| Bacillus thuringiensis | MG470721.1 | 99.72 | N-fixing [33] | |
| Bacillus pumilus | MN581190.1 | 99.93 | N-fixing [34] | |
| Bacillus safensis | CPO43404.1 | 99.58 | N-fixing [35] | |
| Paenibacillaceae | Paenibacillus peoriae | NR_117742.1 | 100 | P solubilizing [36] N-fixing [37] |
| Paenibacillus taichungensis | NR_044428.1 | 96.70 | N-fixing [38] | |
| Paenibacillus kribbensis | NR_025169.1 | 99.04 | N-fixing [38] | |
| Beijerinckiaceae | Beijerinckia fluminensis | NR_116306.1 | 99.76 | N-fixing [39] |
| Family | Scientific Name | Accession Number | Similarity (%) | Function |
|---|---|---|---|---|
| Burkholderiales | Caballeronia fortuita | NR_145600.1 | 99.09 | P solubilizing [40] N cycling [41] |
| Paraburkholderia steyni | NR_164972.1 | 99.19 | P solubilizing [42] N-fixing [43] N cycling [43] | |
| Paraburkholderia sabiae | NR_115261.1 | 99.28 | N-fixing [44] | |
| Paraburkholderia tuberum | NR_118081.1 | 99.83 | N-fixing [45] | |
| Bacillaceae | Gottfrieda luciferensis | NR_025511.1 | 98.41 | N cycling [46] |
| Bacillus pocheonensis | NR_041377.1 | 99.62 | N cycling [46] | |
| Bacillus ginsengisoli | NR_109068.1 | 98.93 | N cycling [47] | |
| Bacillus fungorum | NR_170494.1 | 90.10 | N-fixing [47] | |
| Comamonadaceae | Variovorax guangxiensis | NR_134828.1 | 99.30 | N cycling [48] |
| Chitinophagaceae | Chitinophaga ginsengihumi | NR_134000.1 | 99.19 | N cycling [49] |
| Phyllobacteriaceae | Phyllobacterium brassicacearum | NR_043190.1 | 91.3 | N cycling [50] |
| Oxalobacteraceae | Massilia agilis | NR_157770.1 | 98.51 | N-fixing [51] |
| Rhizobiaceae | Rhizobium mesosinicum | NR_043548.1 | 99.26 | N-fixing [52] |
| Family | Scientific Name | Accession Number | Similarity (%) | Function |
|---|---|---|---|---|
| Burkholderiales | Paraburkholderia steyni | NR_164972.1 | 98.41 | P solubilizing [45] N-fixing [43] N cycling [53] |
| Pseudomonadaceae | Pseudomonas plecoglossicida | NR_114226.1 | 98.38 | P solubilizing [44] N cycling [45] |
| Erythrobacteraceae | Novosphingobium barchaimii | NR_118314.1 | 99.65 | P solubilizing [54] |
| Methylobacteriaceae | Methylobacterium dankookense | NR_116545.1 | 99.77 | P solubilizing [55] N-fixing [56] |
| Bacillaceae | Neobacillus bataviensis | NR_114093.1 | 99.51 | N cycling [57] |
| Bacillus ginsengisoli | NR_109068.1 | 83.02 | N cycling [58] | |
| Sphingomonadaceae | Sphingomonas jatrophae | NR_159248.1 | 85.41 | N cycling [58] |
| Sphingobacteriaceae | Olivibacter jilunii | NR_109321.1 | 99.30 | N cycling [59] |
| Olivibacter oleidegradans | NR_108900.1 | 98.41 | N-fixing [59] | |
| Phyllobacteriaceae | Phyllobacterium brassicacearum | NR_043190.1 | 90.30 | N cycling [59] |
| Enzyme Activity | Rhizosphere | Non-Rhizosphere Soils |
|---|---|---|
| β-(D)-Glucosaminidase (nmolh−1g−1) | 20.47 ± 0.95 a | 20.44 ± 1.03 a |
| Nitrate reductase (µmolh−1g−1) | 4496 ± 2116.6 a | 3178.87 ± 1408.48 a |
| Acid phosphatase (nmolh−1g−1) | 13.13 ± 2.03 a | 13.50 ± 3.98 a |
| Alkaline phosphatase (nmolh−1g−1) | 15.88 ± 4.10 a | 19.47 ± 6.81 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndlovu, S.; Suinyuy, T.N.; Pérez-Fernández, M.A.; Magadlela, A. Encephalartos natalensis, Their Nutrient-Cycling Microbes and Enzymes: A Story of Successful Trade-Offs. Plants 2023, 12, 1034. https://doi.org/10.3390/plants12051034
Ndlovu S, Suinyuy TN, Pérez-Fernández MA, Magadlela A. Encephalartos natalensis, Their Nutrient-Cycling Microbes and Enzymes: A Story of Successful Trade-Offs. Plants. 2023; 12(5):1034. https://doi.org/10.3390/plants12051034
Chicago/Turabian StyleNdlovu, Siphelele, Terence N. Suinyuy, María A. Pérez-Fernández, and Anathi Magadlela. 2023. "Encephalartos natalensis, Their Nutrient-Cycling Microbes and Enzymes: A Story of Successful Trade-Offs" Plants 12, no. 5: 1034. https://doi.org/10.3390/plants12051034
APA StyleNdlovu, S., Suinyuy, T. N., Pérez-Fernández, M. A., & Magadlela, A. (2023). Encephalartos natalensis, Their Nutrient-Cycling Microbes and Enzymes: A Story of Successful Trade-Offs. Plants, 12(5), 1034. https://doi.org/10.3390/plants12051034