
Brassica napus Plants Gain Improved Salt-Stress Tolerance and Increased Storage Oil Biosynthesis by Interfering with CRL3BPM Activities

 , , , and
, , , and 
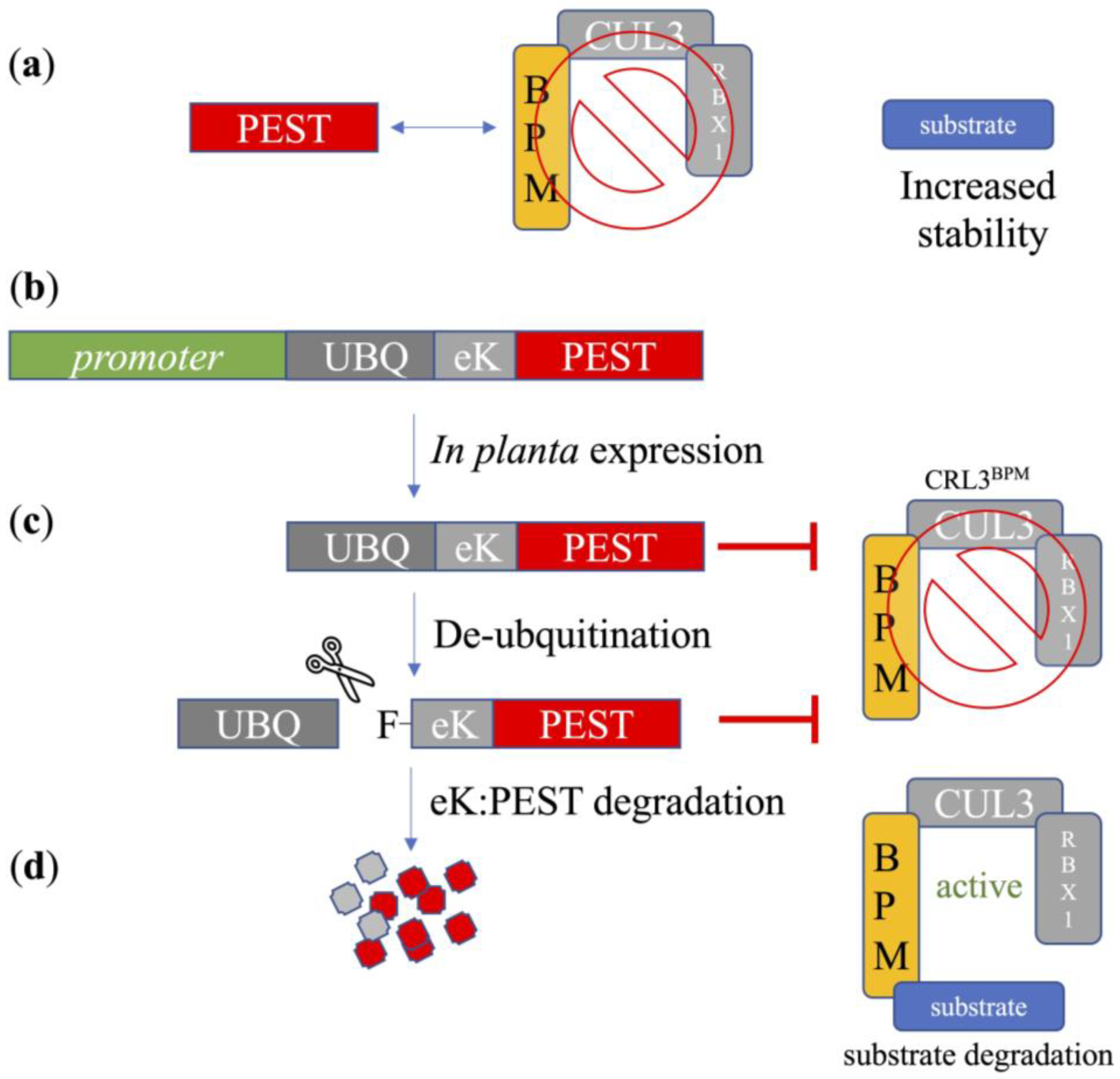{kind=link}
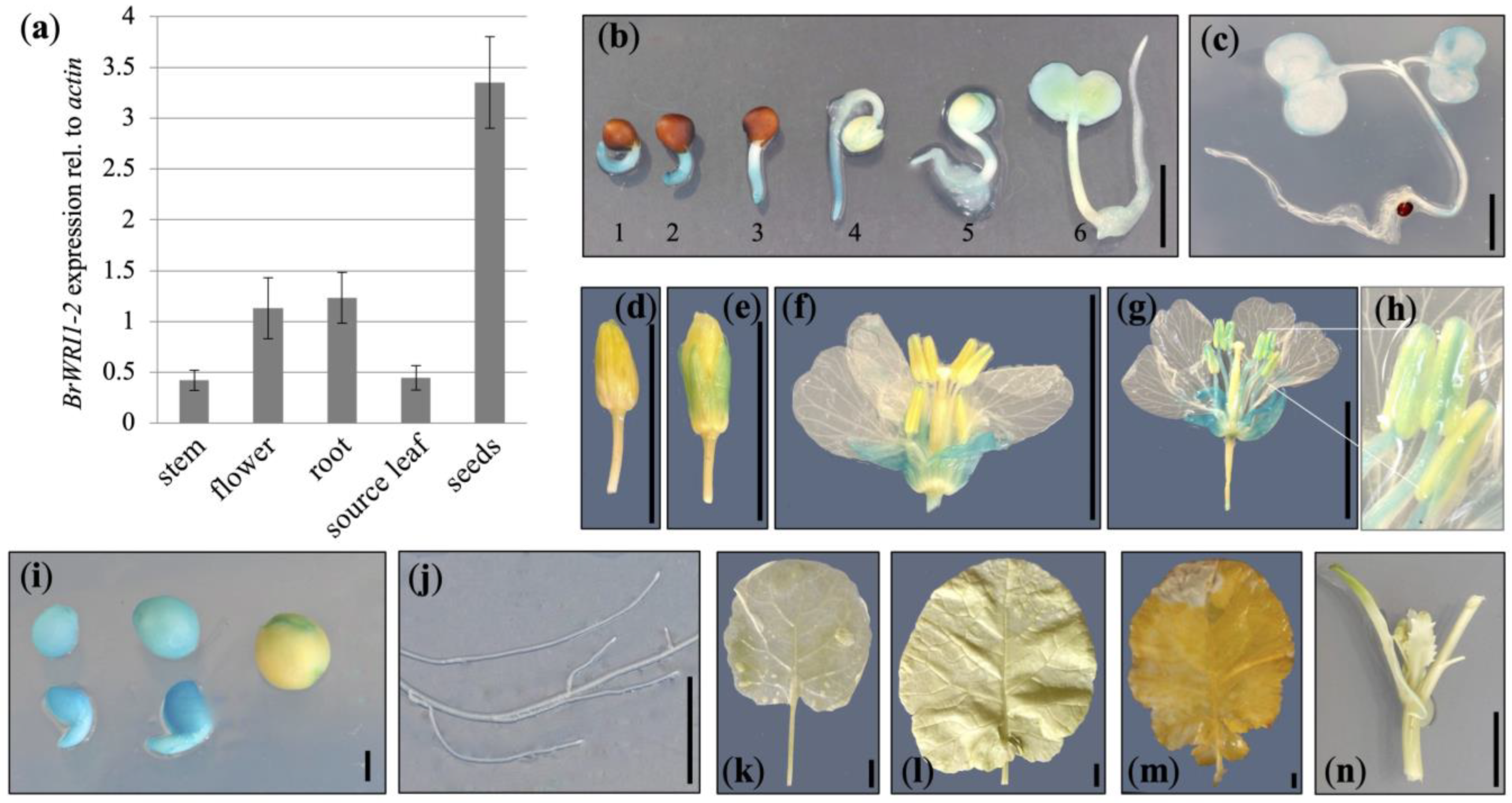{kind=link}
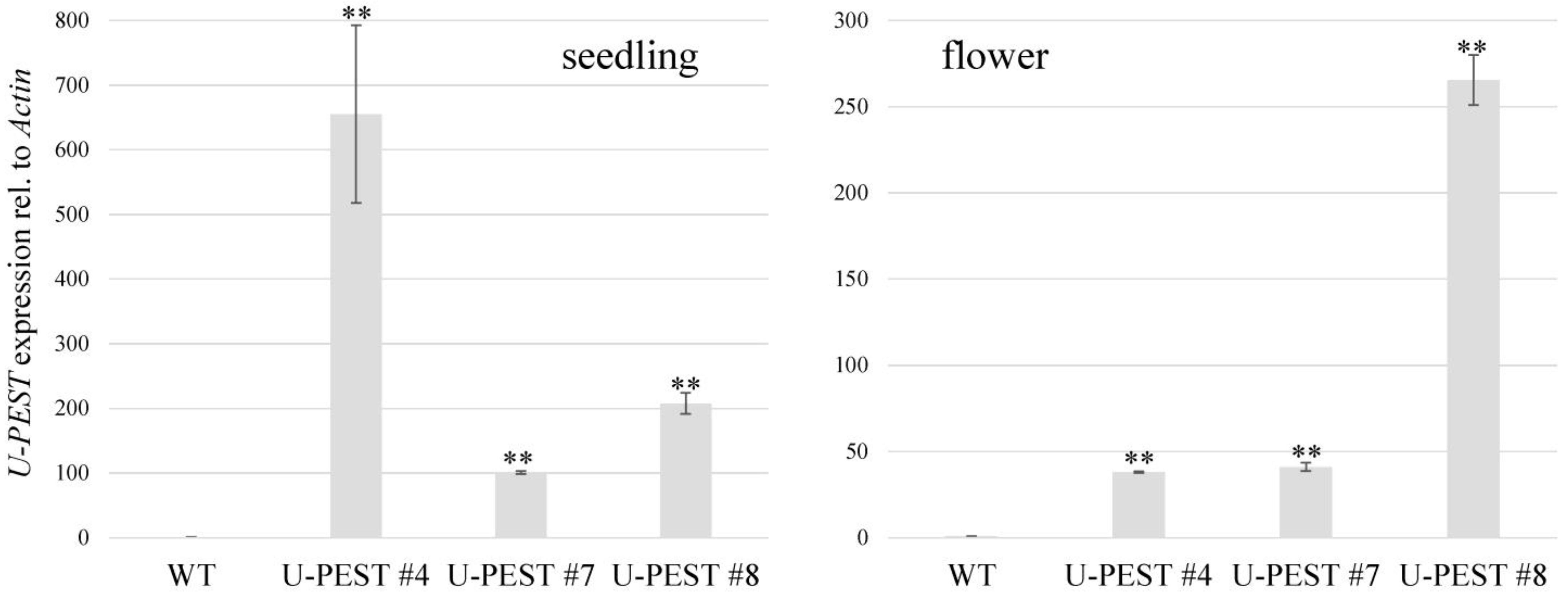{kind=link}
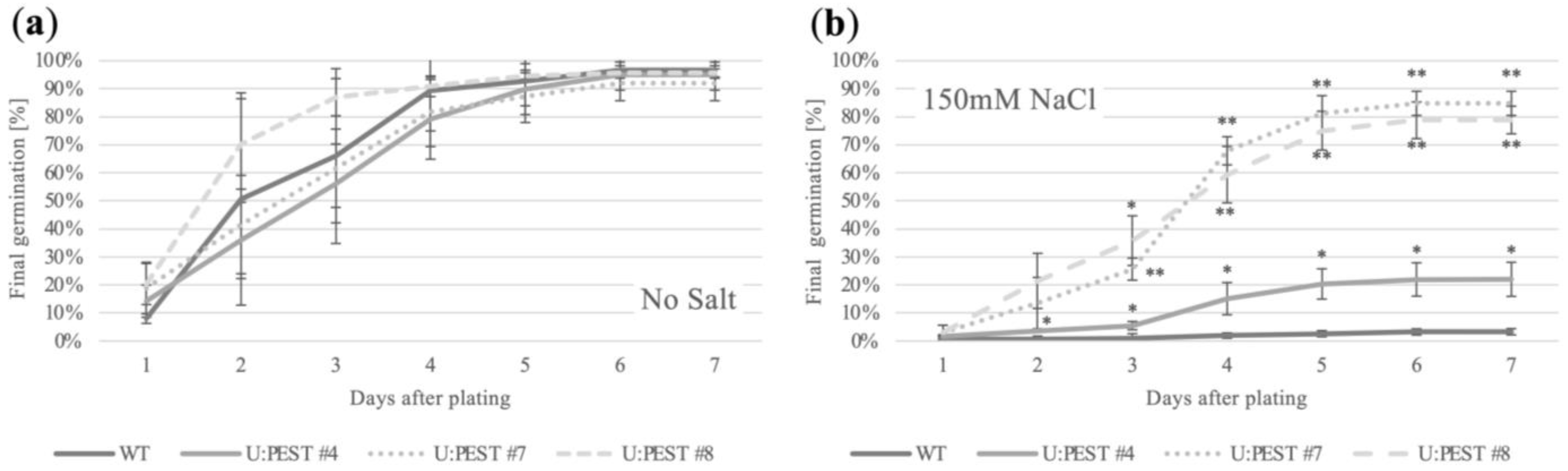{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification of a Suitable Promoter for U:PEST Expression
2.2. Generation of Transgenic B. napus Plants
2.3. proBrWRI1.2:U:PEST Seedlings Have Increased Salt-Stress Tolerance
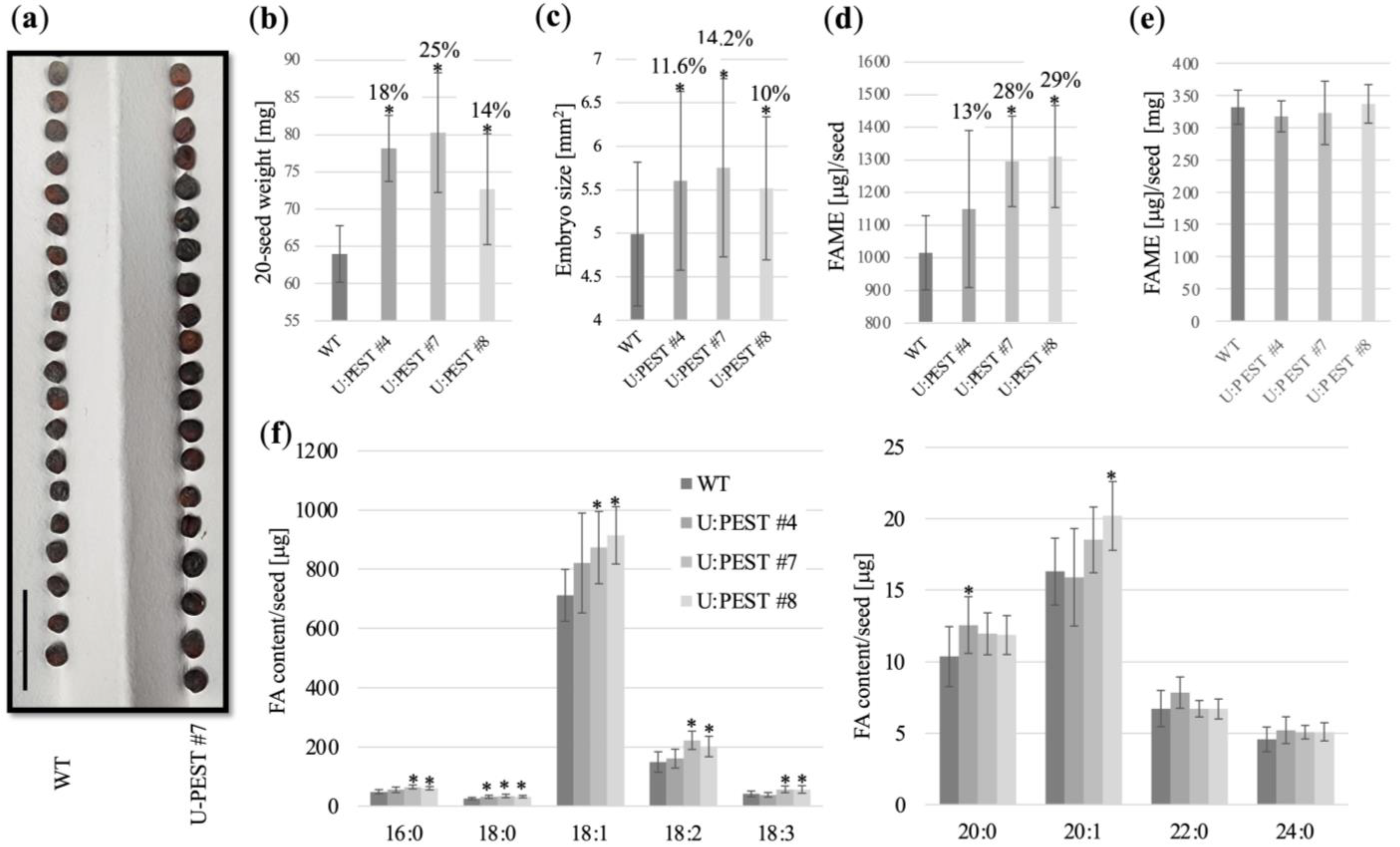
2.4. proBrWRI1.2:U:PEST Plants Have Larger Seeds and Increased Fatty Acid Content
3. Discussion
4. Materials & Methods
4.1. Plant Growth, Transformation, and Phenotypic Analysis
4.2. Generation of Expression Constructs
4.3. GUS Staining
4.4. mRNA Isolation and RT–qPCR Analysis
4.5. Fatty Acid Analysis
4.6. Imaging
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, A.; Mehta, S.; Yadav, S.; Nagar, G.; Ghosh, R.; Roy, A.; Chakraborty, A.; Singh, I.K. How to Cope with the Challenges of Environmental Stresses in the Era of Global Climate Change: An Update on ROS Stave off in Plants. Int. J. Mol. Sci. 2022, 23, 1995. [Google Scholar] [CrossRef] [PubMed]
- Thornton, P.K.; Ericksen, P.J.; Herrero, M.; Challinor, A.J. Climate variability and vulnerability to climate change: A review. Glob. Chang. Biol. 2014, 20, 3313–3328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.A.; Cai, X.M. Climate change impacts on global agricultural land availability. Environ. Res. Lett. 2011, 6, 014014. [Google Scholar] [CrossRef]
- Al-Saharin, R.; Mooney, S.; Hellmann, H. Plant E3 Ligases as Versatile Tools for Novel Drug Development and Plant Bioengineering. Drug Discov. Ser. 2021, 212–233. [Google Scholar] [CrossRef]
- Smalley, S.; Hellmann, H. Review: Exploring possible approaches using ubiquitylation and sumoylation pathways in modifying plant stress tolerance. Plant Sci. 2022, 319, 111275. [Google Scholar] [CrossRef] [PubMed]
- Miricescu, A.; Goslin, K.; Graciet, E. Ubiquitylation in plants: Signaling hub for the integration of environmental signals. J. Exp. Bot. 2018, 69, 4511–4527. [Google Scholar] [CrossRef] [Green Version]
- Sharma, B.; Joshi, D.; Yadav, P.K.; Gupta, A.K.; Bhatt, T.K. Role of Ubiquitin-Mediated Degradation System in Plant Biology. Front. Plant Sci. 2016, 7, 806. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Cano, E.; Zaltsman, A.; Citovsky, V. Assaying proteasomal degradation in a cell-free system in plants. J. Vis. Exp. 2014, 85, 51293. [Google Scholar] [CrossRef]
- Walsh, C.K.; Sadanandom, A. Ubiquitin chain topology in plant cell signaling: A new facet to an evergreen story. Front. Plant Sci. 2014, 5, 122. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Hellmann, H. Plant E3 ligases: Flexible enzymes in a sessile world. Mol. Plant 2013, 6, 1388–1404. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.M.; Gray, W.M.; Mooney, S.; Hellmann, H. Composition, roles, and regulation of cullin-based ubiquitin e3 ligases. Arabidopsis. Book 2014, 12, e0175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, H.; Bernhardt, A.; Dieterle, M.; Hano, P.; Mutlu, A.; Estelle, M.; Genschik, P.; Hellmann, H. Arabidopsis AtCUL3a and AtCUL3b form complexes with members of the BTB/POZ-MATH protein family. Plant Physiol. 2005, 137, 83–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gingerich, D.J.; Gagne, J.M.; Salter, D.W.; Hellmann, H.; Estelle, M.; Ma, L.; Vierstra, R.D. Cullins 3a and 3b assemble with members of the broad complex/tramtrack/bric-a-brac (BTB) protein family to form essential ubiquitin-protein ligases (E3s) in Arabidopsis. J. Biol. Chem. 2005, 280, 18810–18821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juranic, M.; Srilunchang, K.O.; Krohn, N.G.; Leljak-Levanic, D.; Sprunck, S.; Dresselhaus, T. Germline-specific MATH-BTB substrate adaptor MAB1 regulates spindle length and nuclei identity in maize. Plant Cell 2012, 24, 4974–4991. [Google Scholar] [CrossRef] [Green Version]
- Gingerich, D.J.; Hanada, K.; Shiu, S.H.; Vierstra, R.D. Large-scale, lineage-specific expansion of a bric-a-brac/tramtrack/broad complex ubiquitin-ligase gene family in rice. Plant Cell 2007, 19, 2329–2348. [Google Scholar] [CrossRef] [Green Version]
- Juranic, M.; Dresselhaus, T. Phylogenetic analysis of the expansion of the MATH-BTB gene family in the grasses. Plant Signal. Behav. 2014, 9, e28242. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Lee, J.H.; Weber, H.; Tohge, T.; Witt, S.; Roje, S.; Fernie, A.R.; Hellmann, H. Arabidopsis BPM proteins function as substrate adaptors to a cullin3-based E3 ligase to affect fatty acid metabolism in plants. Plant Cell 2013, 25, 2253–2264. [Google Scholar] [CrossRef] [Green Version]
- Weber, H.; Hellmann, H. Arabidopsis thaliana BTB/ POZ-MATH proteins interact with members of the ERF/AP2 transcription factor family. FEBS J. 2009, 276, 6624–6635. [Google Scholar] [CrossRef]
- Mooney, S.; Al-Saharin, R.; Choi, C.M.; Tucker, K.; Beathard, C.; Hellmann, H.A. Characterization of Brassica rapa RAP2.4-Related Proteins in Stress Response and as CUL3-Dependent E3 Ligase Substrates. Cells 2019, 8, 336. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, K.; Ohama, N.; Kidokoro, S.; Mizoi, J.; Takahashi, F.; Todaka, D.; Mogami, J.; Sato, H.; Qin, F.; Kim, J.S.; et al. BPM-CUL3 E3 ligase modulates thermotolerance by facilitating negative regulatory domain-mediated degradation of DREB2A in Arabidopsis. Proc. Nat. Acad. Sci. USA 2017, 114, E8528–E8536. [Google Scholar] [CrossRef] [Green Version]
- Chico, J.M.; Lechner, E.; Fernandez-Barbero, G.; Canibano, E.; Garcia-Casado, G.; Franco-Zorrilla, J.M.; Hammann, P.; Zamarreno, A.M.; Garcia-Mina, J.M.; Rubio, V.; et al. CUL3(BPM) E3 ubiquitin ligases regulate MYC2, MYC3, and MYC4 stability and JA responses. Proc. Nat. Acad. Sci. USA 2020, 117, 6205–6215. [Google Scholar] [CrossRef] [PubMed]
- Beathard, C.; Mooney, S.; Al-Saharin, R.; Goyer, A.; Hellmann, H. Characterization of Arabidopsis thaliana R2R3 S23 MYB Transcription Factors as Novel Targets of the Ubiquitin Proteasome-Pathway and Regulators of Salt Stress and Abscisic Acid Response. Front. Plant Sci. 2021, 12, 629208. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Bernhardt, A.; Lee, J.; Hellmann, H. Identification of Arabidopsis MYB56 as a novel substrate for CRL3(BPM) E3 ligases. Mol. Plant 2015, 8, 242–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lechner, E.; Leonhardt, N.; Eisler, H.; Parmentier, Y.; Alioua, M.; Jacquet, H.; Leung, J.; Genschik, P. MATH/BTB CRL3 receptors target the homeodomain-leucine zipper ATHB6 to modulate abscisic acid signaling. Dev. Cell 2011, 21, 1116–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skiljaica, A.; Lechner, E.; Jagic, M.; Majsec, K.; Malenica, N.; Genschik, P.; Bauer, N. The protein turnover of Arabidopsis BPM1 is involved in regulation of flowering time and abiotic stress response. Plant Mol. Biol. 2020, 102, 359–372. [Google Scholar] [CrossRef]
- Julian, J.; Coego, A.; Lozano-Juste, J.; Lechner, E.; Wu, Q.; Zhang, X.; Merilo, E.; Belda-Palazon, B.; Park, S.Y.; Cutler, S.R.; et al. The MATH-BTB BPM3 and BPM5 subunits of Cullin3-RING E3 ubiquitin ligases target PP2CA and other clade A PP2Cs for degradation. Proc. Nat. Acad. Sci. USA 2019, 116, 15725–15734. [Google Scholar] [CrossRef] [Green Version]
- Bauer, N.; Skiljaica, A.; Malenica, N.; Razdorov, G.; Klasic, M.; Juranic, M.; Mocibob, M.; Sprunck, S.; Dresselhaus, T.; Levanic, D.L. The MATH-BTB Protein TaMAB2 Accumulates in Ubiquitin-Containing Foci and Interacts with the Translation Initiation Machinery in Arabidopsis. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Jagic, M.; Vuk, T.; Skiljaica, A.; Markulin, L.; Bockor, V.V.; Tokic, M.; Miskec, K.; Razdorov, G.; Habazin, S.; Sostar, M.; et al. BPM1 regulates RdDM-mediated DNA methylation via a cullin 3 independent mechanism. Plant Cell Rep. 2022, 41, 2139–2157. [Google Scholar] [CrossRef]
- Zhuang, M.; Calabrese, M.F.; Liu, J.; Waddell, M.B.; Nourse, A.; Hammel, M.; Miller, D.J.; Walden, H.; Duda, D.M.; Seyedin, S.N.; et al. Structures of SPOP-substrate complexes: Insights into molecular architectures of BTB-Cul3 ubiquitin ligases. Mol. Cell 2009, 36, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Belizario, J.E.; Alves, J.; Garay-Malpartida, M.; Occhiucci, J.M. Coupling caspase cleavage and proteasomal degradation of proteins carrying PEST motif. Curr. Prot. Pep. Sci. 2008, 9, 210–220. [Google Scholar] [CrossRef]
- Al-Saharin, R.; Mooney, S.; Dissmeyer, N.; Hellmann, H. Using CRL3(BPM) E3 ligase substrate recognition sites as tools to impact plant development and stress tolerance in Arabidopsis thaliana. Plant Direct 2022, 6, e474. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Kong, Q.; Grix, M.; Mantyla, J.J.; Yang, Y.; Benning, C.; Ohlrogge, J.B. Deletion of a C-terminal intrinsically disordered region of WRINKLED1 affects its stability and enhances oil accumulation in Arabidopsis. Plant J. 2015, 83, 864–874. [Google Scholar] [CrossRef]
- Varshavsky, A. N-degron and C-degron pathways of protein degradation. Proc. Nat. Acad. Sci. USA 2019, 116, 358–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faden, F.; Mielke, S.; Dissmeyer, N. Modulating Protein Stability to Switch Toxic Protein Function on and off in Living Cells. Plant Physiol. 2019, 179, 929–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Plant genetics. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [Green Version]
- To, A.; Joubes, J.; Barthole, G.; Lecureuil, A.; Scagnelli, A.; Jasinski, S.; Lepiniec, L.; Baud, S. WRINKLED transcription factors orchestrate tissue-specific regulation of fatty acid biosynthesis in Arabidopsis. Plant Cell 2012, 24, 5007–5023. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Wang, C.; Sun, Y.; Qu, J.; Mao, H.; Chua, N.H. Overexpression of a Transcription Factor Increases Lipid Content in a Woody Perennial Jatropha curcas. Front. Plant Sci. 2018, 9, 1479. [Google Scholar] [CrossRef] [Green Version]
- Ivarson, E.; Leiva-Eriksson, N.; Ahlman, A.; Kanagarajan, S.; Bulow, L.; Zhu, L.H. Effects of Overexpression of WRI1 and Hemoglobin Genes on the Seed Oil Content of Lepidium campestre. Front. Plant Sci. 2016, 7, 2032. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Kong, Q.; Arondel, V.; Kilaru, A.; Bates, P.D.; Thrower, N.A.; Benning, C.; Ohlrogge, J.B. Wrinkled1, a ubiquitous regulator in oil accumulating tissues from Arabidopsis embryos to oil palm mesocarp. PLoS ONE 2013, 8, e68887. [Google Scholar] [CrossRef]
- Li, Q.; Shao, J.; Tang, S.; Shen, Q.; Wang, T.; Chen, W.; Hong, Y. Wrinkled1 accelerates flowering and regulates lipid homeostasis between oil accumulation and membrane lipid anabolism in Brassica napus. Front. Plant Sci. 2015, 6, 1015. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liang, W.; Shi, J.; Xu, J.; Zhang, D. MYB56 encoding a R2R3 MYB transcription factor regulates seed size in Arabidopsis thaliana. J. Integr. Plant Biol. 2013, 55, 1166–1178. [Google Scholar] [CrossRef] [PubMed]
- Shockey, J.; Mason, C.; Gilbert, M.; Cao, H.P.; Li, X.J.; Cahoon, E.; Dyer, J. Development and analysis of a highly flexible multi-gene expression system for metabolic engineering in Arabidopsis seeds and other plant tissues. Plant Mol. Biol. 2015, 89, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Estelle, M.A.; Somerville, C.R. Auxin Resistant Mutants of Arabidopsis with an Altered Morphology. Mol. Gen. Genet. MGG 1987, 206, 200–206. [Google Scholar] [CrossRef]
- Cardoza, V.; Stewart, C.N. Increased Agrobacterium-mediated transformation and rooting efficiencies in canola (Brassica napus L.) from hypocotyl segment explants. Plant Cell Rep. 2003, 21, 599–604. [Google Scholar] [CrossRef]
- Curtis, M.D.; Grossniklaus, U. A gateway cloning vector set for high-throughput functional analysis of genes in planta. Plant Physiol. 2003, 133, 462–469. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, R.A. The Gus Reporter Gene System. Nature 1989, 342, 837–838. [Google Scholar] [CrossRef]
- Niu, Y.; Wu, G.Z.; Ye, R.; Lin, W.H.; Shi, Q.M.; Xue, L.J.; Xu, X.D.; Li, Y.; Du, Y.G.; Xue, H.W. Global analysis of gene expression profiles in Brassica napus developing seeds reveals a conserved lipid metabolism regulation with Arabidopsis thaliana. Mol. Plant 2009, 2, 1107–1122. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wang, M.; Garneau, M.G.; Poudel, A.N.; Lamm, D.; Koo, A.J.; Bates, P.D.; Thelen, J.J. Overexpression of pea α-carboxyltransferase in Arabidopsis and camelina increases fatty acid synthesis leading to improved seed oil content. Plant J. 2022, 110, 1035–1046. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corbridge, E.; MacGregor, A.; Al-Saharin, R.; Garneau, M.G.; Smalley, S.; Mooney, S.; Roje, S.; Bates, P.D.; Hellmann, H. Brassica napus Plants Gain Improved Salt-Stress Tolerance and Increased Storage Oil Biosynthesis by Interfering with CRL3BPM Activities. Plants 2023, 12, 1085. https://doi.org/10.3390/plants12051085
Corbridge E, MacGregor A, Al-Saharin R, Garneau MG, Smalley S, Mooney S, Roje S, Bates PD, Hellmann H. Brassica napus Plants Gain Improved Salt-Stress Tolerance and Increased Storage Oil Biosynthesis by Interfering with CRL3BPM Activities. Plants. 2023; 12(5):1085. https://doi.org/10.3390/plants12051085
Chicago/Turabian StyleCorbridge, Emily, Alexandra MacGregor, Raed Al-Saharin, Matthew G. Garneau, Samuel Smalley, Sutton Mooney, Sanja Roje, Philip D. Bates, and Hanjo Hellmann. 2023. "Brassica napus Plants Gain Improved Salt-Stress Tolerance and Increased Storage Oil Biosynthesis by Interfering with CRL3BPM Activities" Plants 12, no. 5: 1085. https://doi.org/10.3390/plants12051085
APA StyleCorbridge, E., MacGregor, A., Al-Saharin, R., Garneau, M. G., Smalley, S., Mooney, S., Roje, S., Bates, P. D., & Hellmann, H. (2023). Brassica napus Plants Gain Improved Salt-Stress Tolerance and Increased Storage Oil Biosynthesis by Interfering with CRL3BPM Activities. Plants, 12(5), 1085. https://doi.org/10.3390/plants12051085