

Hypericum foliosum Quality Botanical and Chemical Markers and In Vitro Antioxidant and Anticancer Activities


 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Macroscopic Analysis
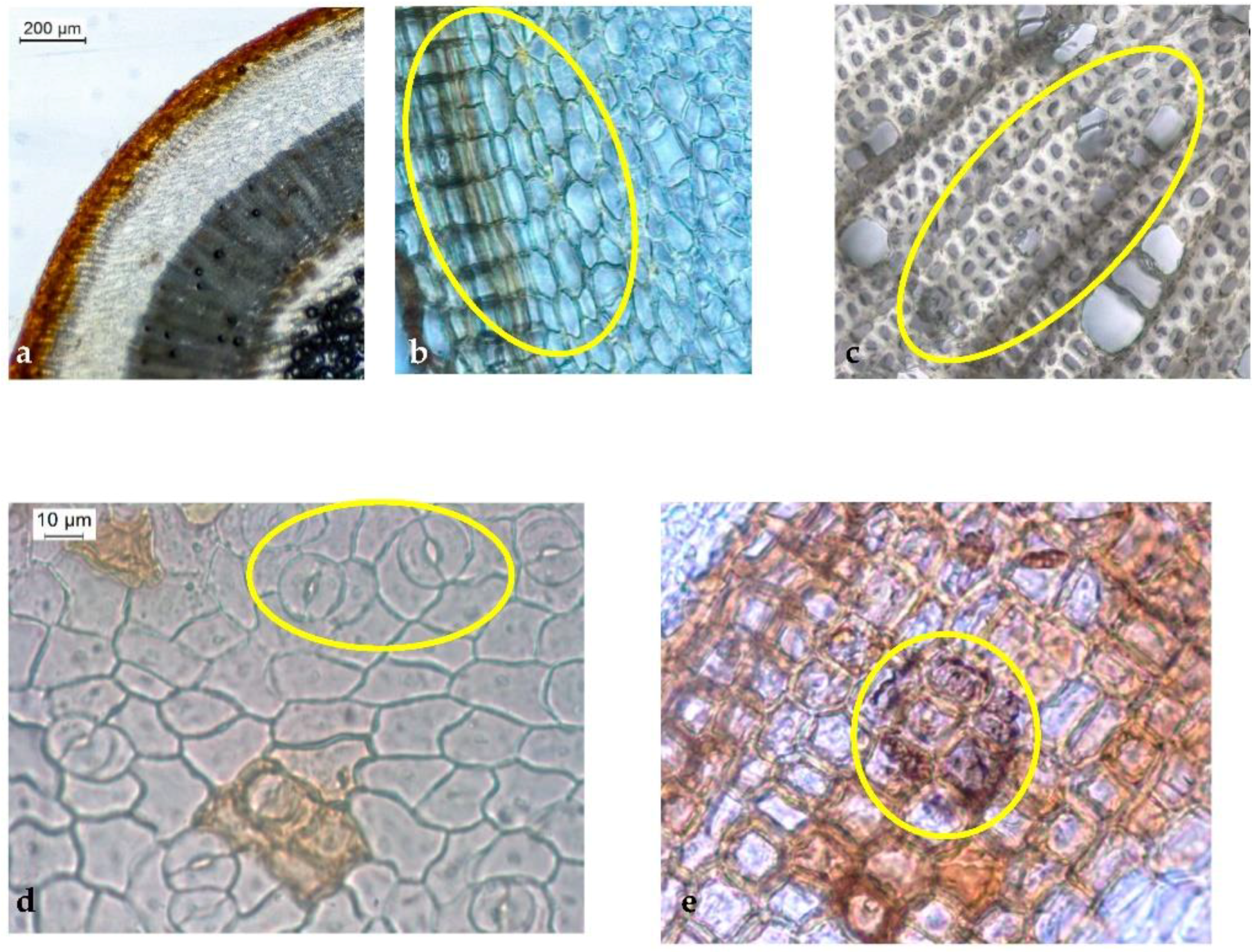
2.2. Microscopic Analysis
2.2.1. Stem
2.2.2. Leaf
2.2.3. Powder
2.3. Antioxidant Activity
2.4. In Vitro Anticancer Activity
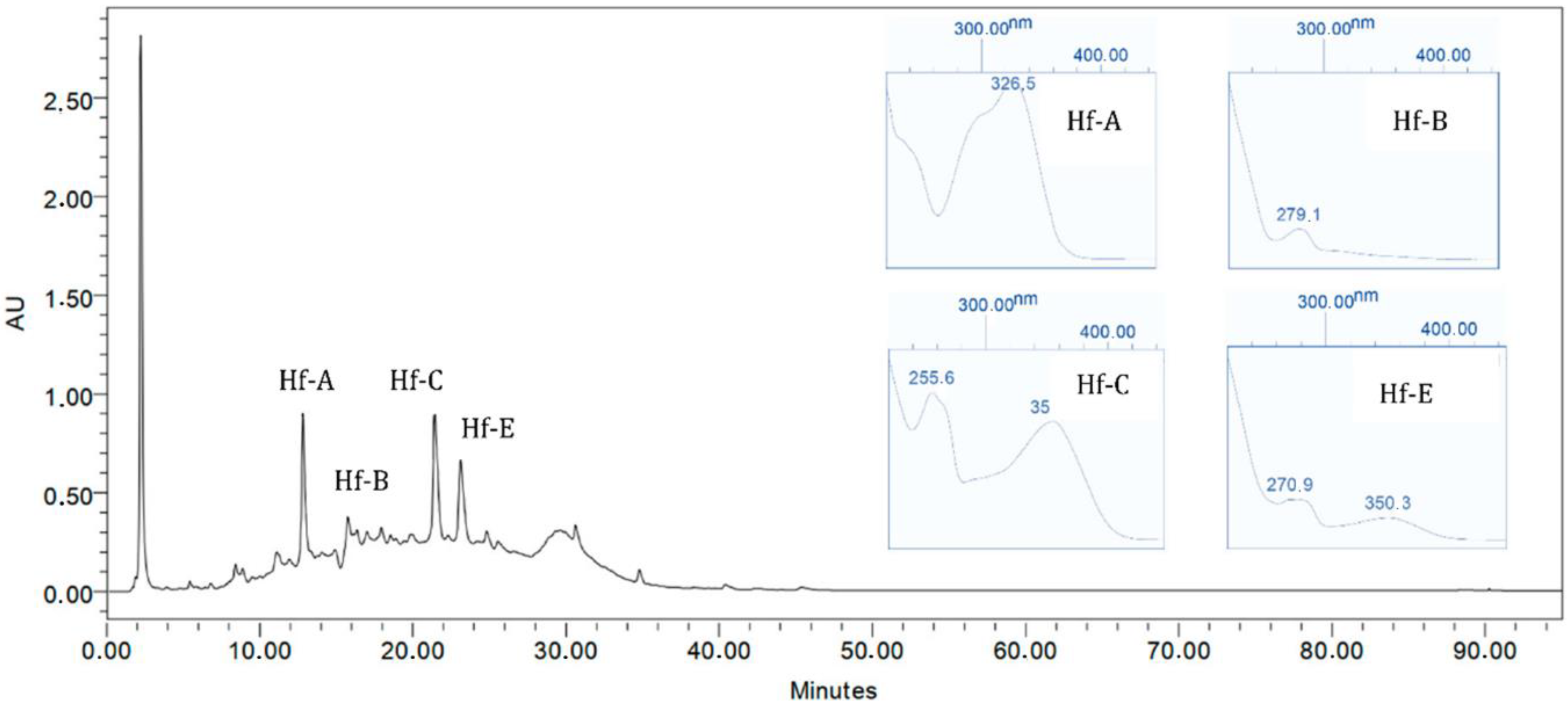
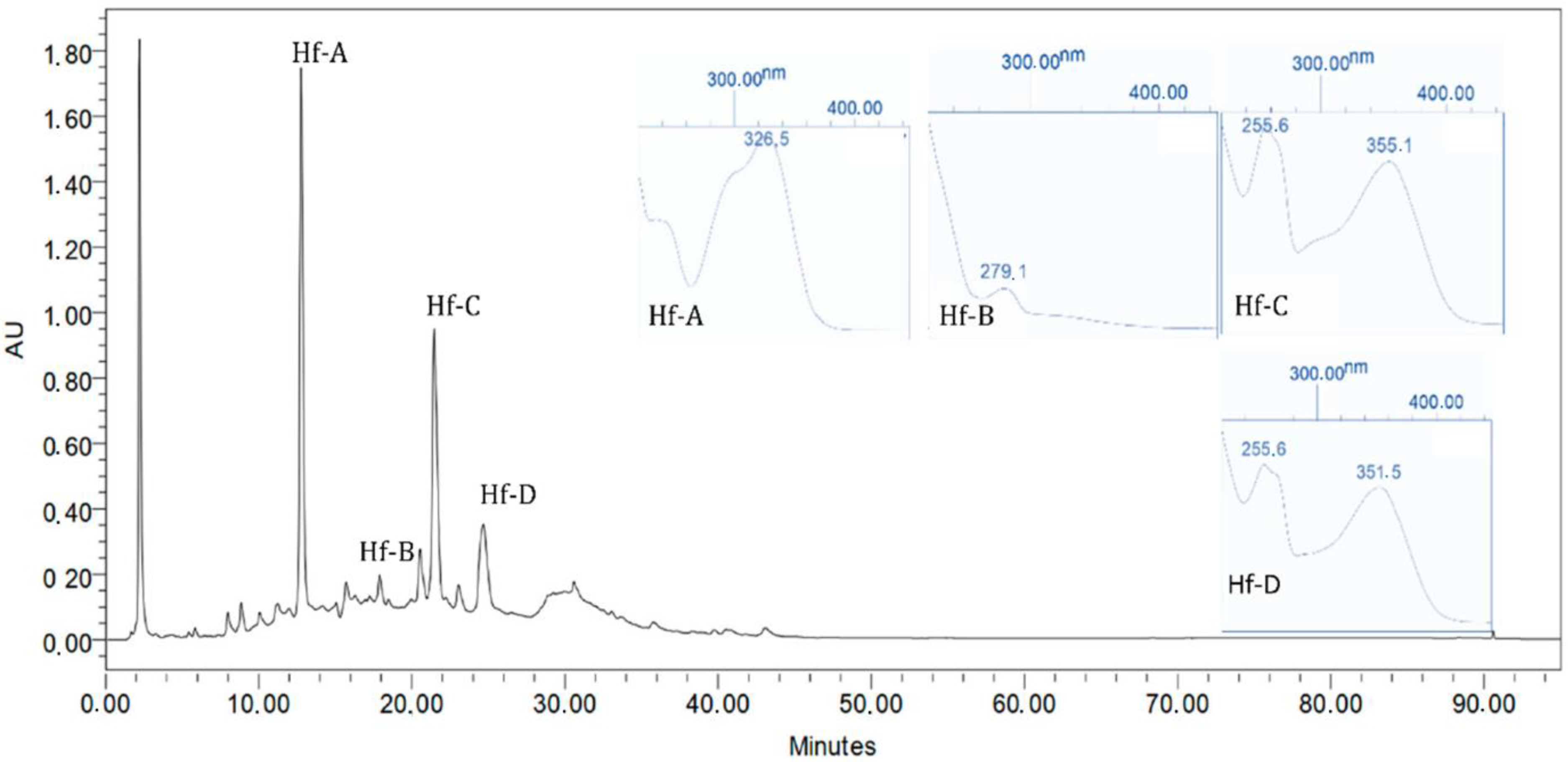
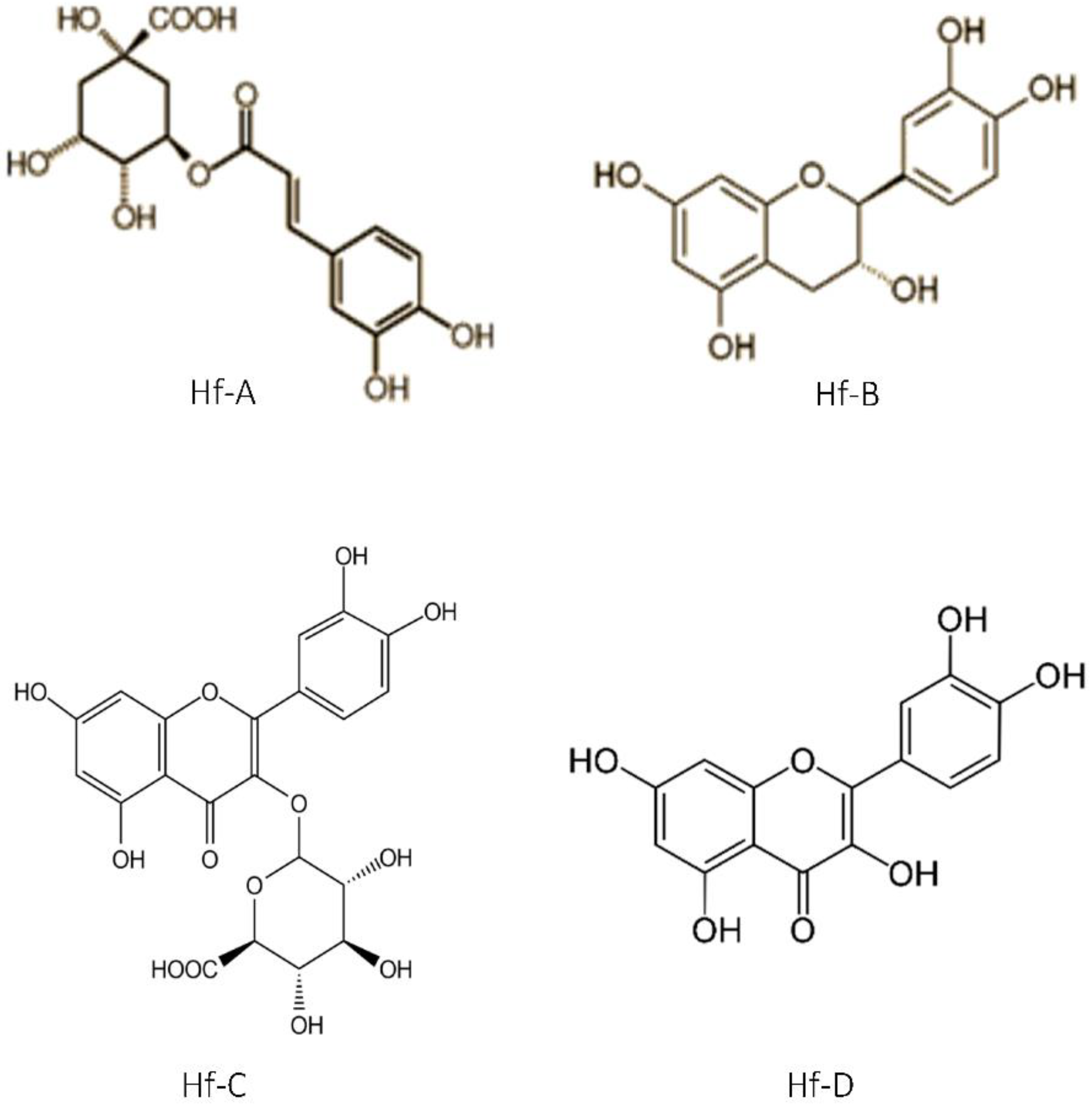
2.5. Chromatographic Profile
3. Discussion
4. Material and Methods
4.1. Plant Material
4.2. Macroscopic Analysis (MA)
4.3. Light Microscopy (LM)
4.4. Quantitative and Statistical Analysis
4.5. Extract Preparation
- The first portion was used for the whole-extract preparation (Hf. E), adding ten times the amount of cold ethanol (70%) and soaking it for 24 h. This procedure was repeated until the total raw material was exhausted. The obtained solution was filtered and evaporated under reduced pressure (T < 40 °C) in a rotary evaporator. The Hf. E total extract was kept in a desiccator at room temperature and protected from light;
- The second portion was used for the dichloromethane/methanol 1:1 (Hf. DM) and water (Hf. W) extracts. The raw material was completely covered with 1:1 DCM/methanol for 24 h. The Hf. DM extract was obtained after evaporating both solutions under pressure with a rotary vacuum flask evaporator;
- After the extraction with DCM/methanol and methanol, the final obtained residue was re-extracted with ultrapure water for 24 h. The obtained solution was freeze dried for 2~3 days and lyophilized (Hf. W).
4.6. Biochemical Antioxidant Assays
4.6.1. DPPH (2,2-Diphenyl-1-picrylhydrazyl) Assay for Free Radical Scavenging Activity
4.6.2. Ferric Reducing Antioxidant Power (FRAP) Assay for Reducing Power
4.6.3. Phosphomolybdic Acid (PA) Assay for Total Antioxidant Activity
4.7. Anticancer Activity
4.7.1. Tumor Cell Lines
4.7.2. MTT Assay
4.8. LC-UV/DAD Chromatographic Profile
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ramalhete, N.; Machado, A.; Serrano, R.; Gomes, E.T.; Mota-Filipe, H.; Silva, O. Comparative study on the in vivo antidepressant activities of the Portuguese Hypericum foliosum, Hypericum androsaemum and Hypericum perforatum medicinal plants. Ind. Crops Prod. 2016, 82, 29–36. [Google Scholar] [CrossRef]
- Santos, P.A.; Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G.; Scheffer, J.J. Composition of the essential oil of Hypericum foliosum Aiton from five Azorean islands. Flavour Fragr. J. 1999, 14, 283–286. [Google Scholar] [CrossRef]
- Ruhfel, B.R.; Bittrich, V.; Bove, C.P.; Gustafsson, M.H.; Philbrick, C.T.; Rutishauser, R.; Xi, Z.; Davis, C.C. Phylogeny of the clusioid clade (Malpighiales): Evidence from the plastid and mitochondrial genomes. Am. J. Bot. 2011, 98, 306–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nürk, N.M.; Crockett, S.L. Morphological and Phytochemical Diversity among Hypericum Species of the Mediterranean Basin. Med. Aromat. Plant Sci. Biotechnol. 2011, 5, 14–28. [Google Scholar] [PubMed]
- do Amaral Franco, J. Nova Flora de Portugal; Sociedade Astória: Lisboa, Portugal, 1971; Volume 1. [Google Scholar]
- Robson, S.L.C.N.K.B. Taxonomy and Chemotaxonomy of the Genus Hypericum. Med. Aromat. Plant Sci. Biotechnol. 2012, 5, 1–13. [Google Scholar]
- Rainha, N.; Lima, E.; Baptista, J. Comparison of the endemic Azorean Hypericum foliosum with other Hypericum species: Antioxidant activity and phenolic profile. Nat. Prod. Res. 2011, 25, 123–135. [Google Scholar] [CrossRef] [PubMed]
- EMA. Community Herbal Monograph Hypericum perforatum L., Herba (Well-Established Medicinal Use); European Medicines Agency—Committee on Herbal Medicinal Products (HPMC): Lisboa, Portugal, 2009.
- Cardile, A.; Zanre, V.; Campagnari, R.; Asson, F.; Addo, S.S.; Orlandi, E.; Menegazzi, M. Hyperforin Elicits Cytostatic/Cytotoxic Activity in Human Melanoma Cell Lines, Inhibiting Pro-Survival NF-kappaB, STAT3, AP1 Transcription Factors and the Expression of Functional Proteins Involved in Mitochondrial and Cytosolic Metabolism. Int. J. Mol. Sci. 2023, 24, 1263. [Google Scholar] [CrossRef]
- Deng, X.; Xia, J.; Hu, B.; Hou, X.C.; Pu, X.D.; Wu, L. Hyjapones A-D, trimethylated acyphloroglucinol meroterpenoids from Hypericum japonicum thunb. With anti-inflammatory activity. Phytochemistry 2022, 202, 113308. [Google Scholar] [CrossRef]
- Zhai, X.; Chen, Y.; Han, X.; Zhu, Y.; Li, X.; Zhang, Y.; Lu, Y. The protective effect of hypericin on postpartum depression rat model by inhibiting the NLRP3 inflammasome activation and regulating glucocorticoid metabolism. Int. Immunopharmacol. 2022, 105, 108560. [Google Scholar] [CrossRef]
- Rafailovska, E.; Tushevski, O.; Shijakova, K.; Simic, S.G.; Kjovkarovska, S.D.; Miova, B. Hypericum perforatum L. extract exerts insulinotropic effects and inhibits gluconeogenesis in diabetic rats by regulating AMPK expression and PKCepsilon concentration. J. Ethnopharmacol. 2023, 302, 115899. [Google Scholar] [CrossRef]
- Rainha, N.; Lima, E.; Baptista, J.; Rodrigues, C. Antioxidant properties, total phenolic, total carotenoid and chlorophyll content of anatomical parts of Hypericum foliosum. J. Med. Plants Res. 2011, 5, 1930–1940. [Google Scholar]
- Arruda, M.; Rainha, N.; Barreto, M.; Lima, E.; Baptista, J. Acetylcholinesterase inhibition properties of Hypericum foliosum Aiton. Planta Med. 2010, 76, P073. [Google Scholar] [CrossRef]
- Gibbons, S.; Moser, E.; Hausmann, S.; Stavri, M.; Smith, E.; Clennett, C. An anti-staphylococcal acylphloroglucinol from Hypericum foliosum. Phytochemistry 2005, 66, 1472–1475. [Google Scholar] [CrossRef] [PubMed]
- Moura, M. Conservation ofHypericum foliosum aiton, an endemic azorean species, by micropropagation. Vitr. Cell. Dev. Biol. 1998, 34, 244–248. [Google Scholar] [CrossRef]
- Caldeira, G.I.; Gouveia, L.P.; Serrano, R.; Silva, O.D. Hypericum Genus as a Natural Source for Biologically Active Compounds. Plants 2022, 11, 2509. [Google Scholar] [CrossRef]
- Bottega, S.; Garbar, F.; Pagni, A.M. Secretory Structures in Hypericum elodes L. (Hypericaceae). I. Preliminary Observations; Atti—Società Toscana di Scienze Naturali: Pisa, Italy, 1999. [Google Scholar]
- Perrone, R.; De Rosa, P.; De Castro, O.; Colombo, P. A further analysis of secretory structures of some taxa belonging to the genus Hypericum (Clusiaceae) in relation to the leaf vascular pattern. Turk. J. Bot. 2013, 37, 847–858. [Google Scholar] [CrossRef]
- Machado, A.M.P. Contribuição para o Conhecimento do Perfil Metabolómico e das Potencialidades Farmacológicas de Hypericum foliosum Aiton. Master’s Thesis, Faculdade de Farmácia da Universidade de Lisboa, Lisbon, Portugal, 2012. [Google Scholar]
- Serrano, R.; Ferreira, P.; Gomes, E.T.; Silva, O. The Use of SEM and Light Microscopy for the Characterization of Hypericum foliosum Aerial Part as a Medicinal Plant. Microsc. Microanal. 2008, 14, 107–108. [Google Scholar] [CrossRef] [Green Version]
- Tekin, M. Pharmacobotanical study of Hypericum thymopsis. Rev. Bras. Farmacogn. 2017, 27, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Fornasiero, R.B.; Bianchi, A.; Pinetti, A. Anatomical and Ultrastuctural Observations inHypericum perforatumL. Leaves. J. Herbs Spices Med. Plants 1998, 5, 21–33. [Google Scholar] [CrossRef]
- Ciccarelli, D.; Andreucci, A.C.; Pagni, A.M. The “black nodules” of Hypericum perforatum L. subsp. perforatum: Morphological,anatomical, and histochemical studies during the course of ontogenesis. Isr. J. Plant Sci. 2001, 49, 33–40. [Google Scholar]
- Ciccarelli, D. Translucent Glands and Secretory Canals in Hypericum perforatum L. (Hypericaceae): Morphological, Anatomical and Histochemical Studies During the Course of Ontogenesis. Ann. Bot. 2001, 88, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Perrone, R.; Rosa, P.; Castro, O.; Colombo, P. Leaf and stem anatomy in eight Hypericum species (Clusiaceae). Acta Bot. Croat. 2013, 72, 269–286. [Google Scholar] [CrossRef]
- Osinska, B.L.E. Shoot anatomy and secretory structures in Hypericum species (Hypericaceae). Bot. J. Linn. Soc. 2010, 163, 70–86. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Statistical Parameters, n = 40 | Min | Max | Mean | Median | ±SD | |
|---|---|---|---|---|---|---|
| Leaf Size | Length (cm) | 2.8 | 5.3 | 4.0 | 4.0 | 0.6 |
| Width (cm) | 1.1 | 2.8 | 1.9 | 1.9 | 0.4 | |
| Stem Size | Diameter (mm) | 2.6 | 4.3 | 3.5 | 3.4 | 0.4 |
| Internode distance (cm) | 0.8 | 4.0 | 2.0 | 1.8 | 0.7 | |
| Statistical Parameters N = 20 | Min | Max | Mean | Median | ±SD |
|---|---|---|---|---|---|
| Stem Surface | |||||
| Epidermal cell area (µm2) | 254.7 | 1053.1 | 639.8 | 653.8 | 165.0 |
| Transverse Section | |||||
| Phloem thickness (µm) | 17.1 | 95.4 | 48.0 | 47.9 | 14.9 |
| Xylem vessel area (µm2) | 210.5 | 1419.4 | 639.4 | 627.1 | 252.1 |
| Epidermal cell layers (number) | 2.0 | 8.0 | 4.4 | 4.0 | 1.4 |
| Medullary ray width (cell number) | 1.0 | 3.0 | 1.2 | 1.0 | 0.5 |
| Secretory canals—type A (diameter, µm) | 18.6 | 65.9 | 29.1 | 27.8 | 8.7 |
| Statistical Parameters N = 20 | Min | Max | Mean | Median | ±SD | |
|---|---|---|---|---|---|---|
| Leaf Surface | ||||||
| Adaxial epidermal cells | Area (µm2) | 79.3 | 802.3 | 317.4 | 369.6 | 168.8 |
| Number | 29.0 | 59.0 | 43.5 | 44.5 | 7.2 | |
| Abaxial epidermal cells | Area (µm2) | 187.1 | 650.9 | 382.4 | 356.2 | 154.4 |
| Number | 26.0 | 100.0 | 59.9 | 58.0 | 17.9 | |
| Abaxial stomatal index (SI) (%) | 5.1 | 18.8 | 11.5 | 11.9 | 3.6 | |
| Transverse Section | ||||||
| Mesophyll thickness (µm) | 94.3 | 177.9 | 126.9 | 124.3 | 18.9 | |
| Adaxial cuticle thickness (µm) | 1.7 | 6.9 | 4.0 | 3.9 | 1.7 | |
| Abaxial cuticle thickness (µm) | 1.5 | 6.8 | 3.6 | 3.2 | 1.4 | |
| Palisade parenchyma length (µm) | 28.6 | 49.1 | 36.6 | 36.2 | 5.1 | |
| Spongy parenchyma length (µm) | 56.9 | 144.7 | 89.1 | 87.7 | 17.8 | |
| Palisade parenchyma/spongy parenchyma ratio | 0.2 | 0.6 | 0.4 | 0.4 | 0.1 | |
| Translucent gland diameter (µm) | 18.1 | 66.4 | 44.7 | 43.1 | 11.2 | |
| Midrib thickness (µm) | 294.0 | 882.1 | 719.3 | 754.9 | 148.1 | |
| Phloem thickness (µm) | 40.5 | 92.5 | 63.8 | 62.3 | 15.6 | |
| Xylem thickness (µm) | 65.4 | 97.3 | 80.2 | 77.7 | 8.8 | |
| Secretory canals—type B diameter (µm) | 11.2 | 59.7 | 32.2 | 27.7 | 12.2 | |
| DPPH | FRAP | PA | |
|---|---|---|---|
| IC50 (µg/mL) ±SD | mg AAE/g dE ± SD | IC50 (µg/mL) ±SD | |
| Hf. E | 490.5 ± 20.6 | 351.9 ± 12.15 | 180.9 ± 6.2 |
| Hf. DM | 695.7 ± 29.0 | 349.5 ± 4.47 | 154.7 ± 5.3 |
| Hf. W | 902.2 ± 27.7 | 346.5 ± 3.31 | 411.4 ± 14.3 |
| Ascorbic Acid | 84.4 ± 0.7 | / | 49.1 ± 0.4 |
| MDA-Mb-436 | A549 | HCT8 | |
|---|---|---|---|
| IC50 (µg/mL) ± SD | IC50 (µg/mL) ± SD | IC50 (µg/mL) ± SD | |
| Hf. E | 108.6 ± 11.9 | 76.3 ± 9.1 | 119.9 V 10.5 |
| Hf. DM | 71.5 ± 4.0 | 27.3 ± 2.1 | 37.7 ± 5.5 |
| Hf.W | > 200 | >200 | >200 |
| Time (Min) | %H2O + 0.05%TFA | %MeCN | %MeOH |
|---|---|---|---|
| 0 | 98 | 2 | 0 |
| 12 | 85 | 15 | 0 |
| 25 | 70 | 20 | 0 |
| 35 | 40 | 50 | 10 |
| 40 | 31 | 54 | 10 |
| 49 | 21 | 64 | 15 |
| 55 | 15 | 70 | 15 |
| 80 | 5 | 80 | 15 |
| 81 | 98 | 2 | 0 |
| 90 | 90 | 2 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caldeira, G.I.; Zhang, G.; Gouveia, L.P.; Videira, M.; Serrano, R.; Silva, O. Hypericum foliosum Quality Botanical and Chemical Markers and In Vitro Antioxidant and Anticancer Activities. Plants 2023, 12, 1087. https://doi.org/10.3390/plants12051087
Caldeira GI, Zhang G, Gouveia LP, Videira M, Serrano R, Silva O. Hypericum foliosum Quality Botanical and Chemical Markers and In Vitro Antioxidant and Anticancer Activities. Plants. 2023; 12(5):1087. https://doi.org/10.3390/plants12051087
Chicago/Turabian StyleCaldeira, Gonçalo Infante, Guanghong Zhang, Luís Pleno Gouveia, Mafalda Videira, Rita Serrano, and Olga Silva. 2023. "Hypericum foliosum Quality Botanical and Chemical Markers and In Vitro Antioxidant and Anticancer Activities" Plants 12, no. 5: 1087. https://doi.org/10.3390/plants12051087
APA StyleCaldeira, G. I., Zhang, G., Gouveia, L. P., Videira, M., Serrano, R., & Silva, O. (2023). Hypericum foliosum Quality Botanical and Chemical Markers and In Vitro Antioxidant and Anticancer Activities. Plants, 12(5), 1087. https://doi.org/10.3390/plants12051087