
Anti-Oxidative and Anti-Aging Effects of Ethanol Extract of the Officinal Breynia (Breynia vitis-idaea) In Vitro

 , ,
, , 
Abstract
:1. Introduction
2. Results
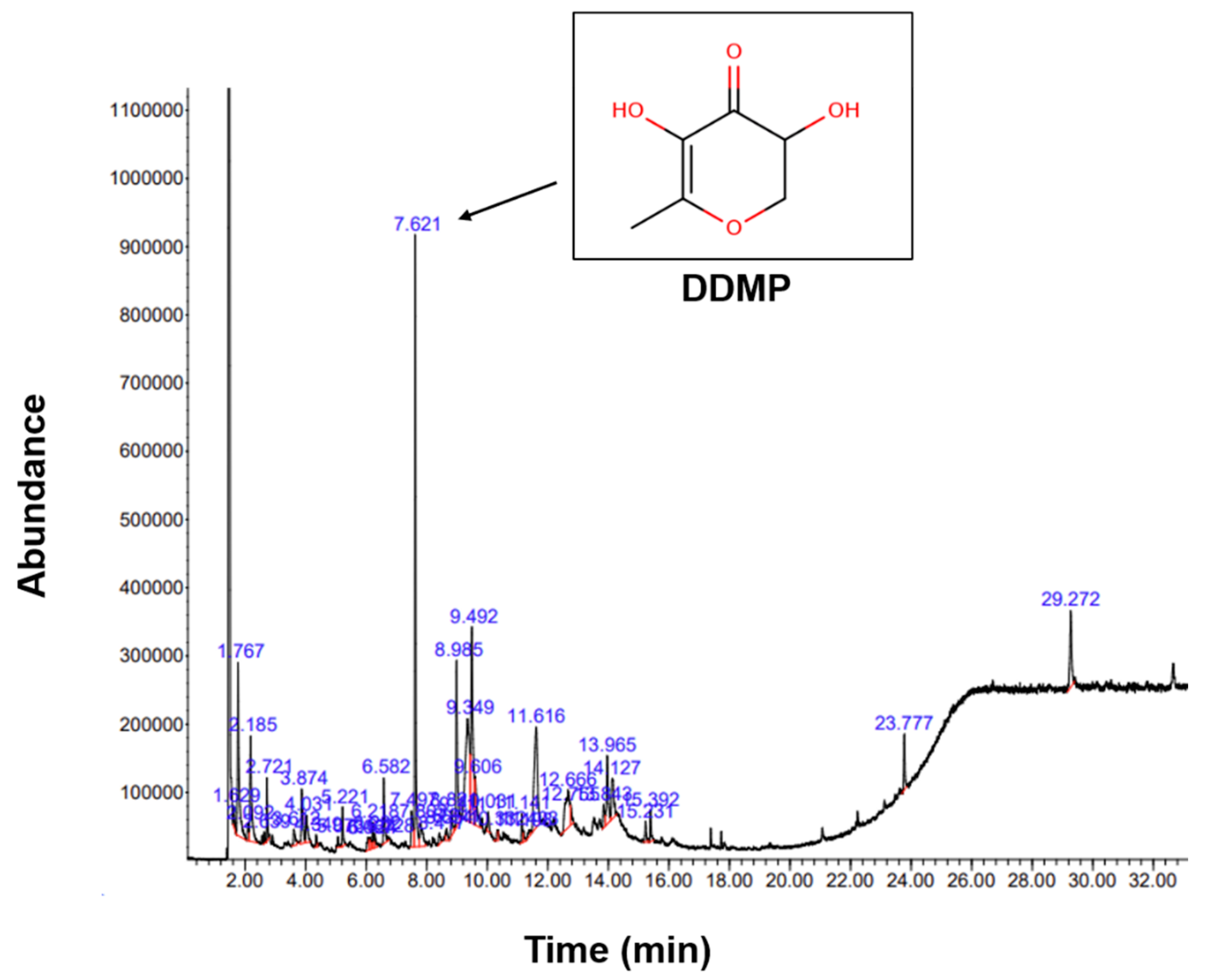
2.1. Phytochemical Components of Bv-EE
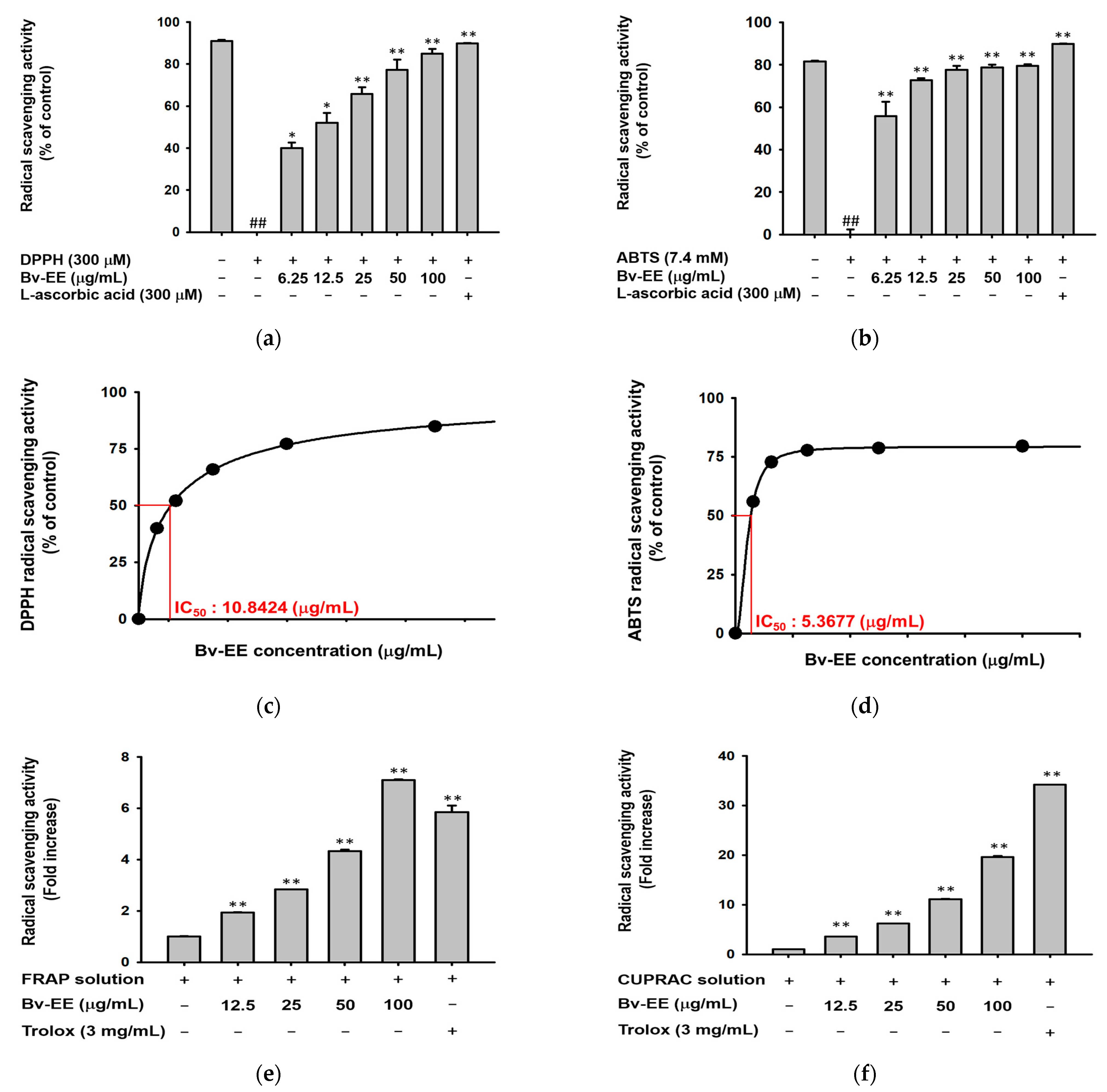
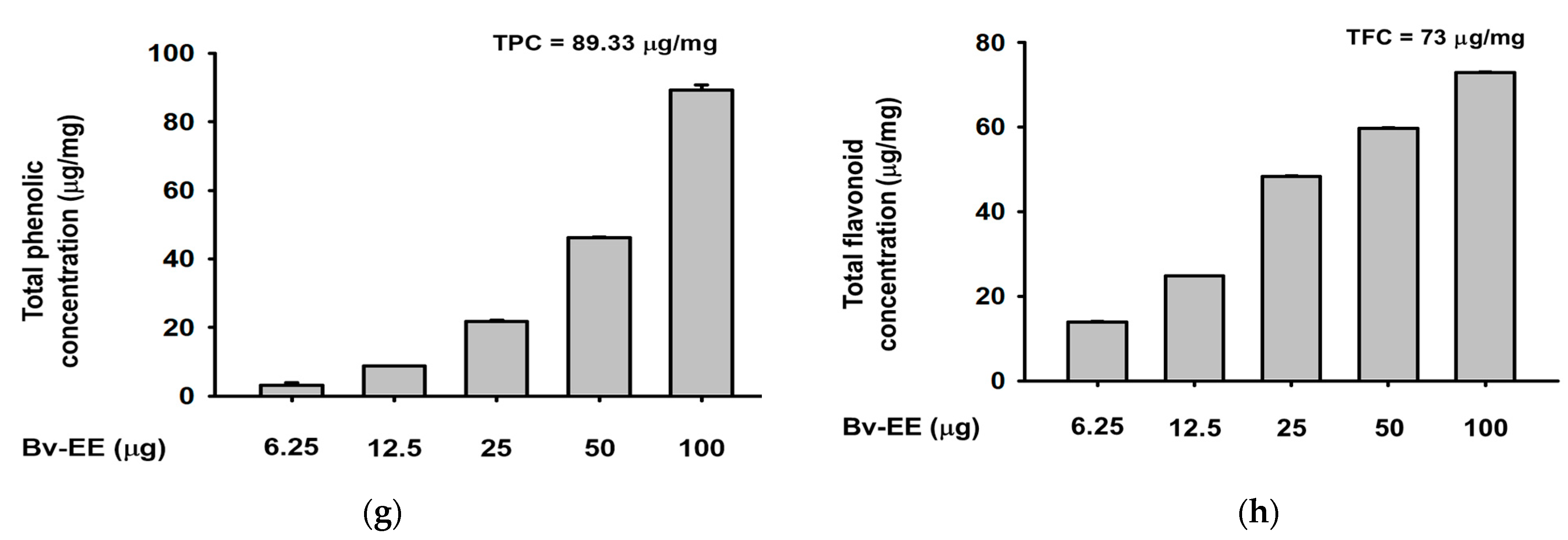
2.2. Bv-EE Exerts Reactive Oxygen Species Scavenging Activity
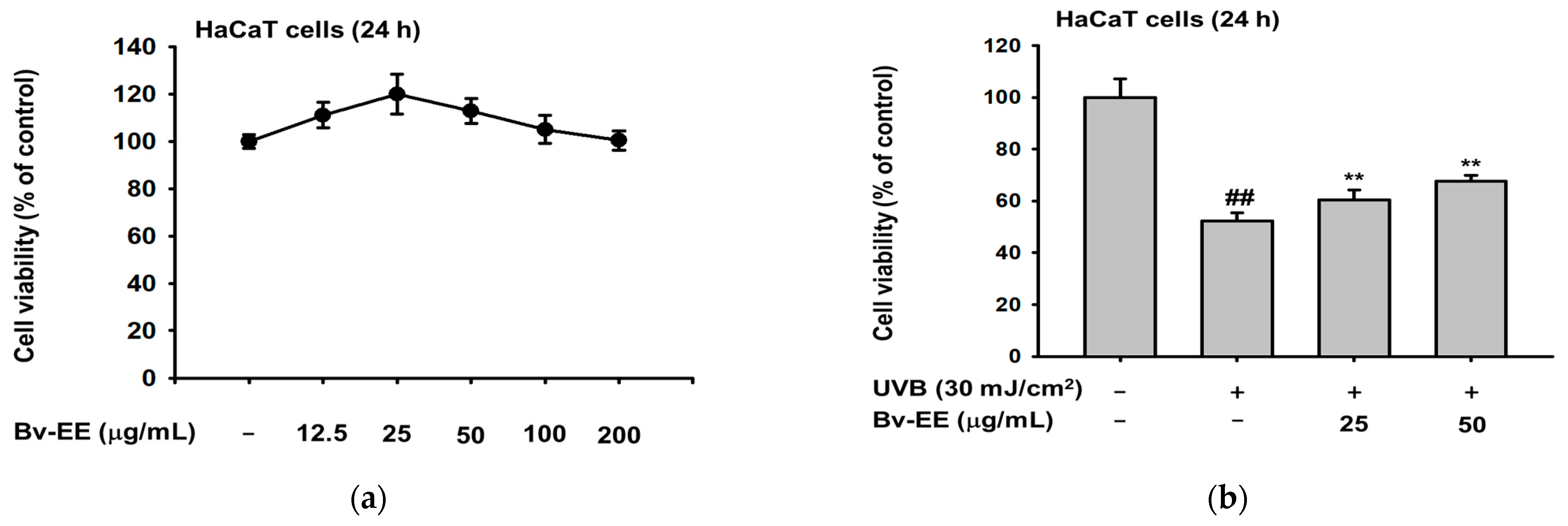
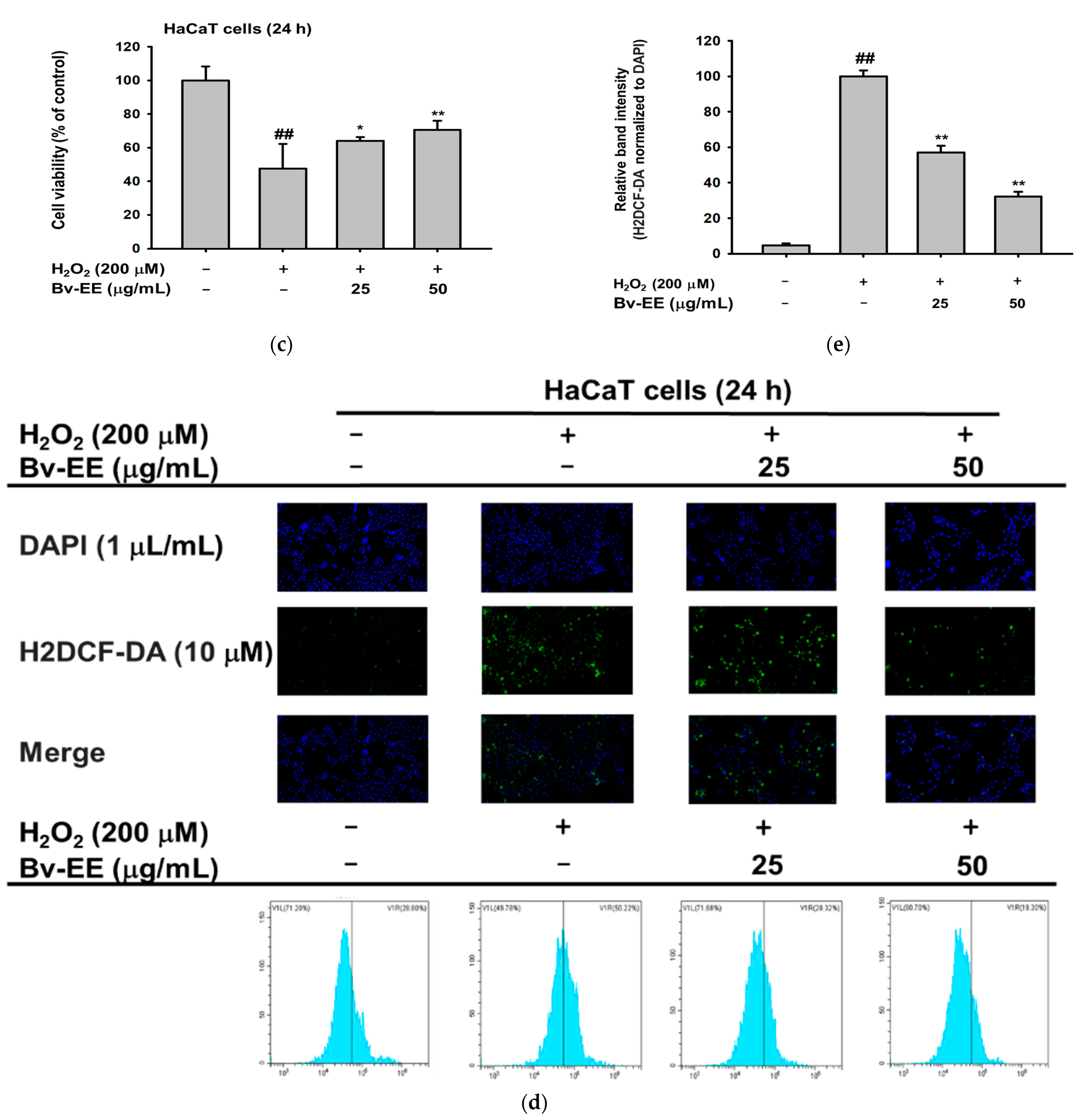
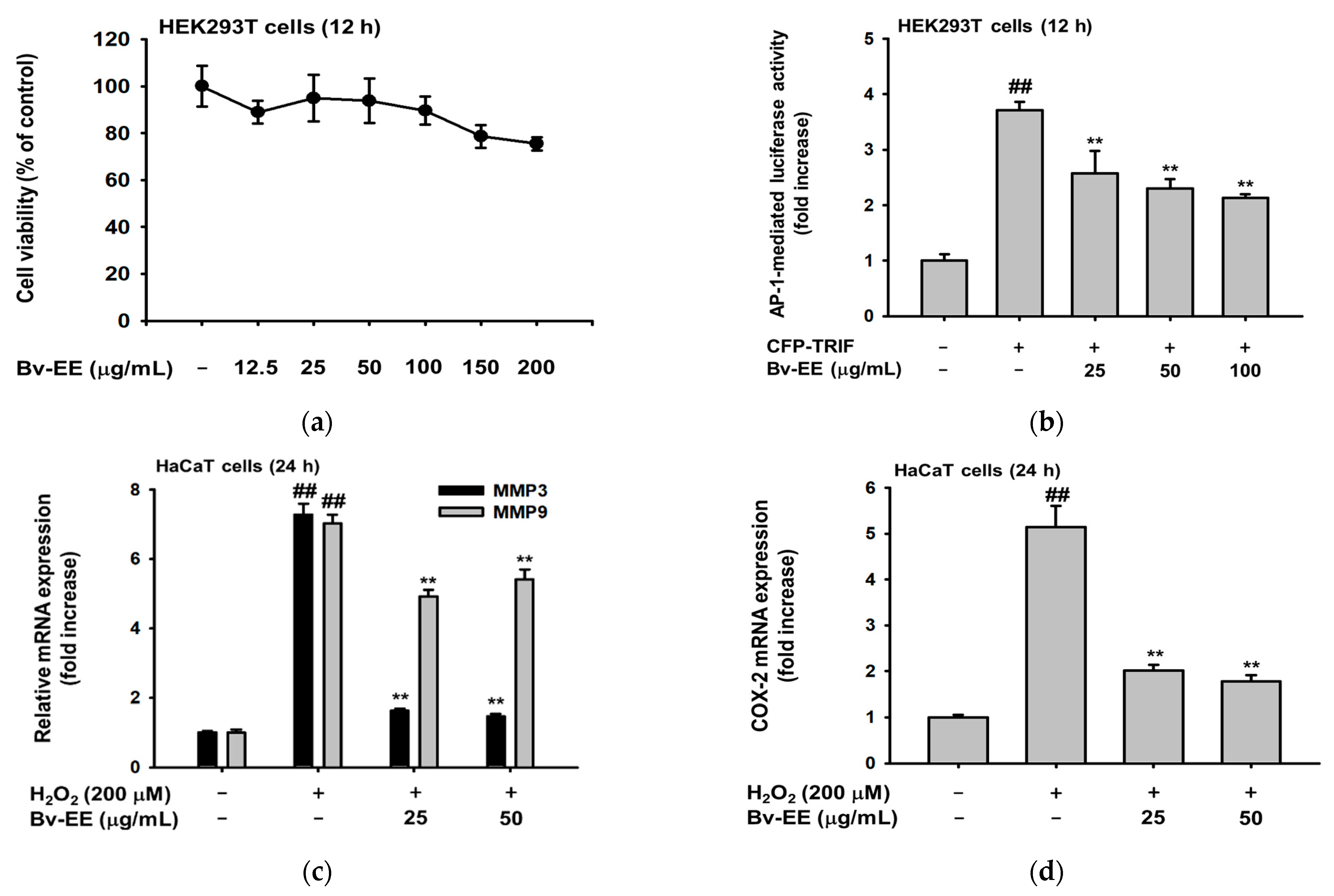
2.3. Bv-EE Prevents Cell Death Caused by UVB and H2O2 in HaCaT cells
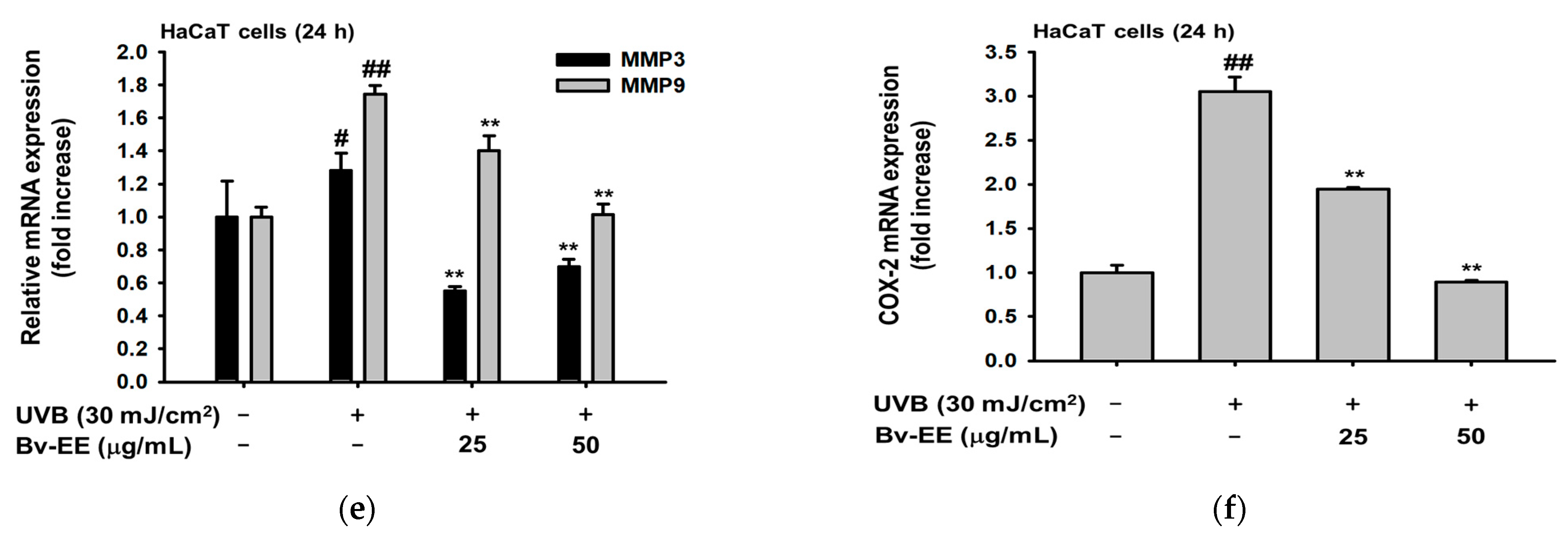
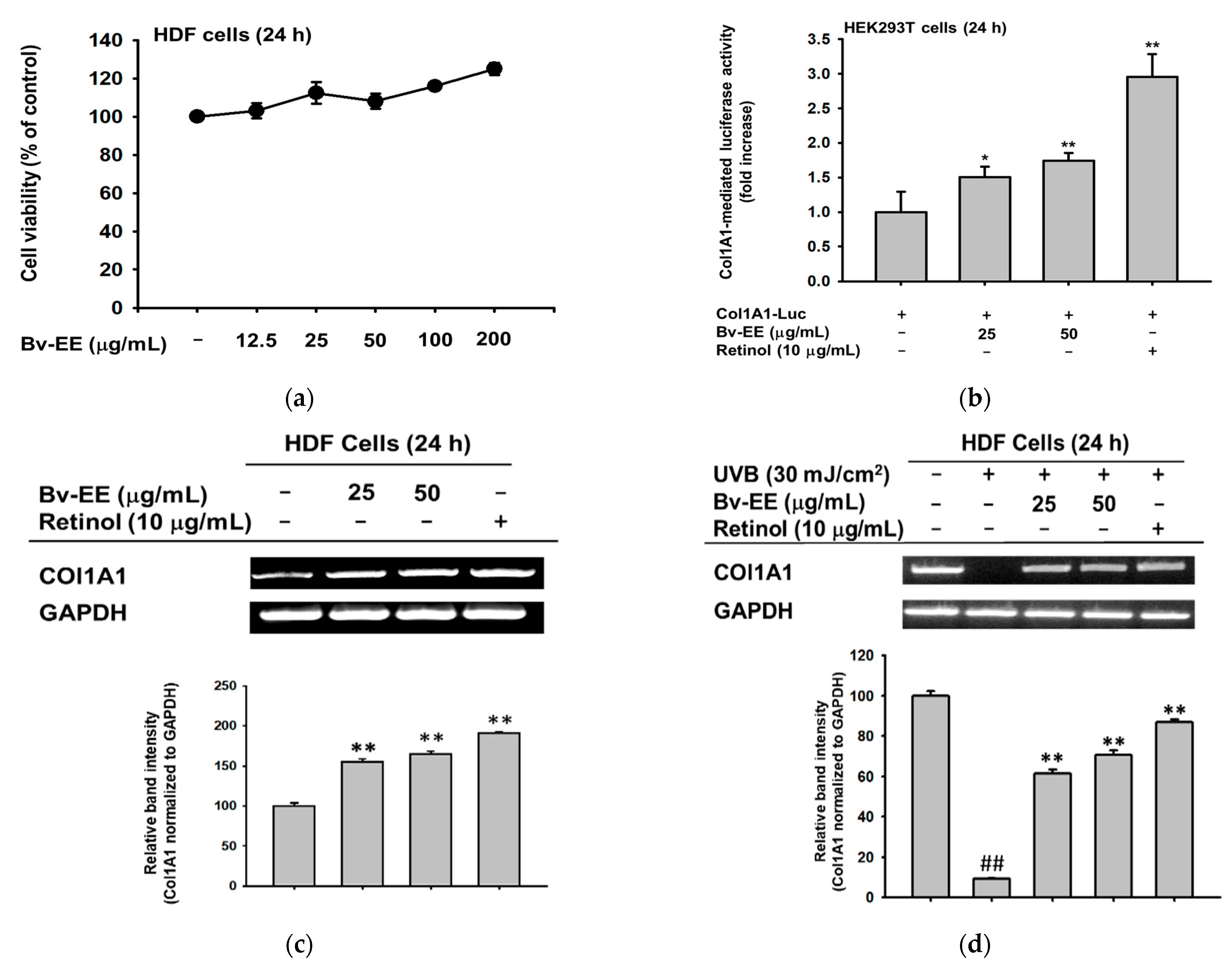
2.4. Bv-EE Inhibits AP-1 Promoter Activity and mRNA Expression of Aging Factors
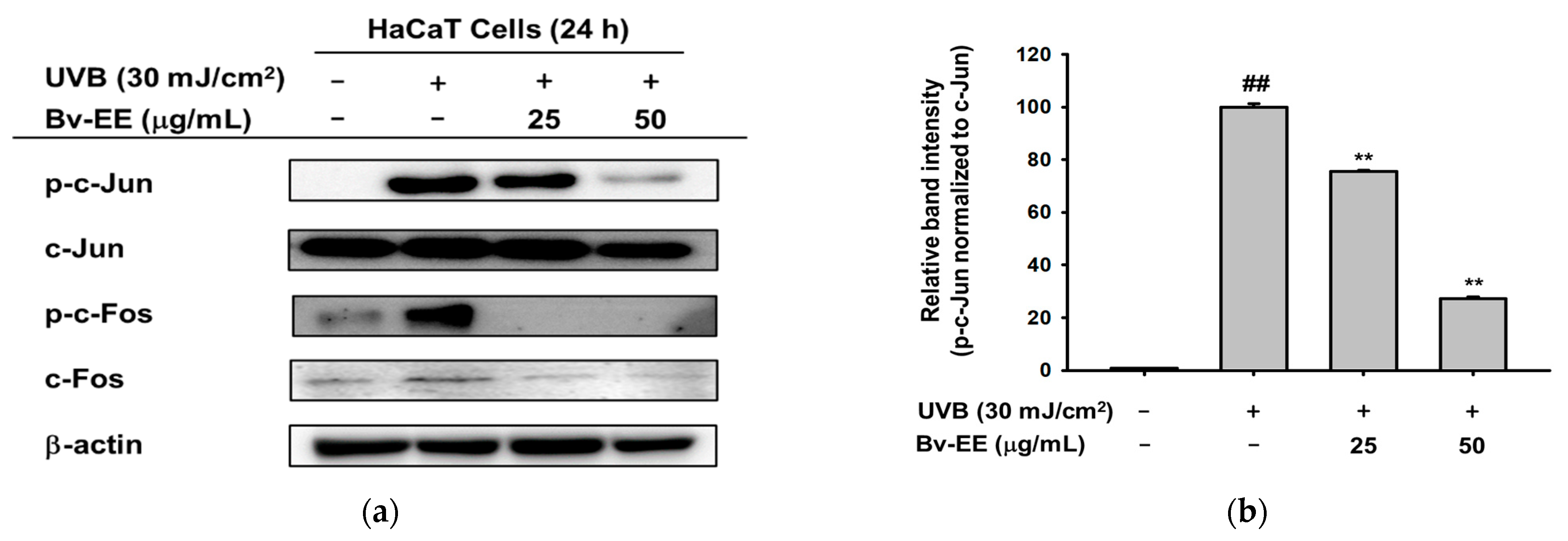
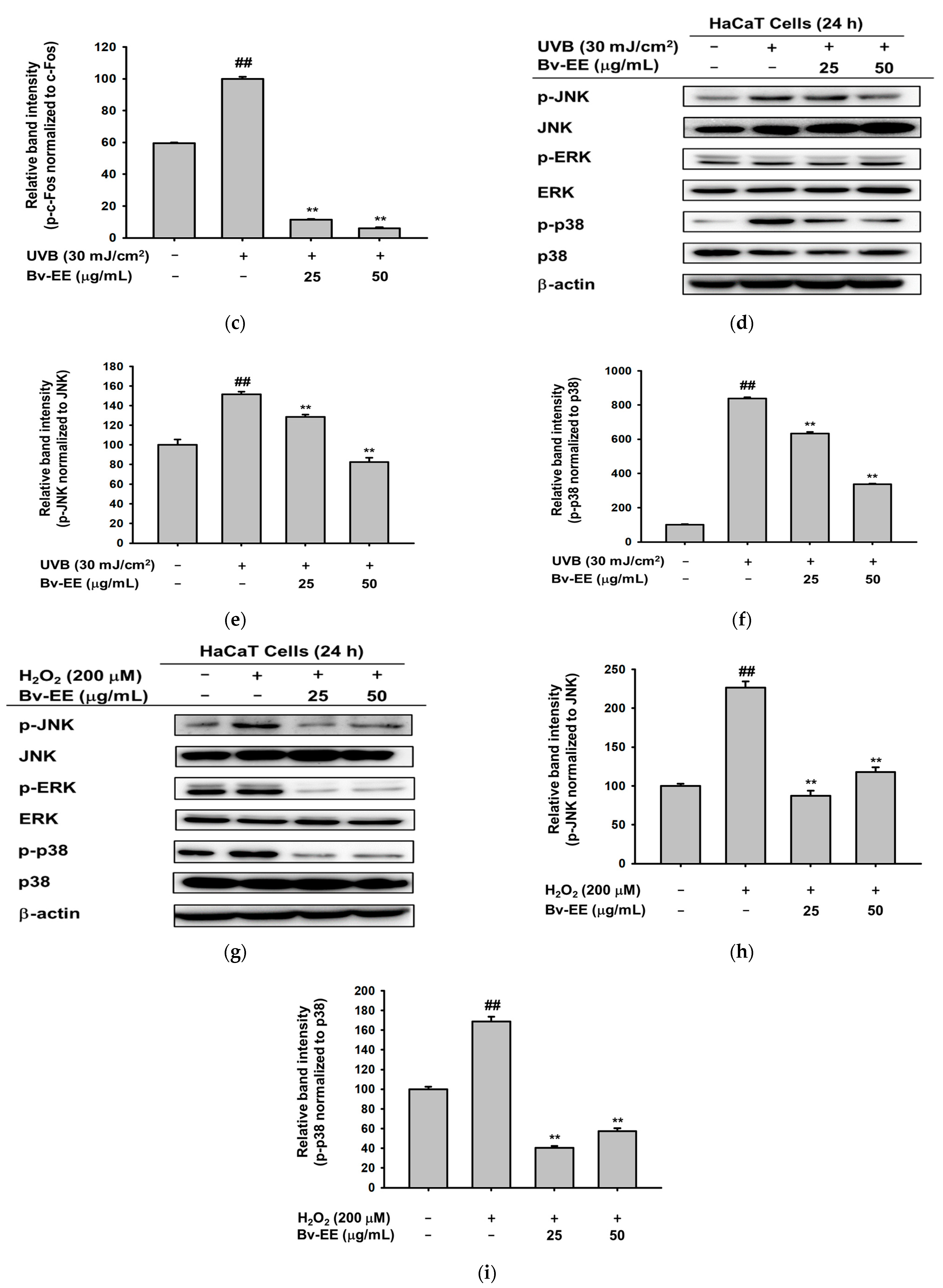
2.5. Bv-EE Inhibits the AP-1 Signaling Pathway
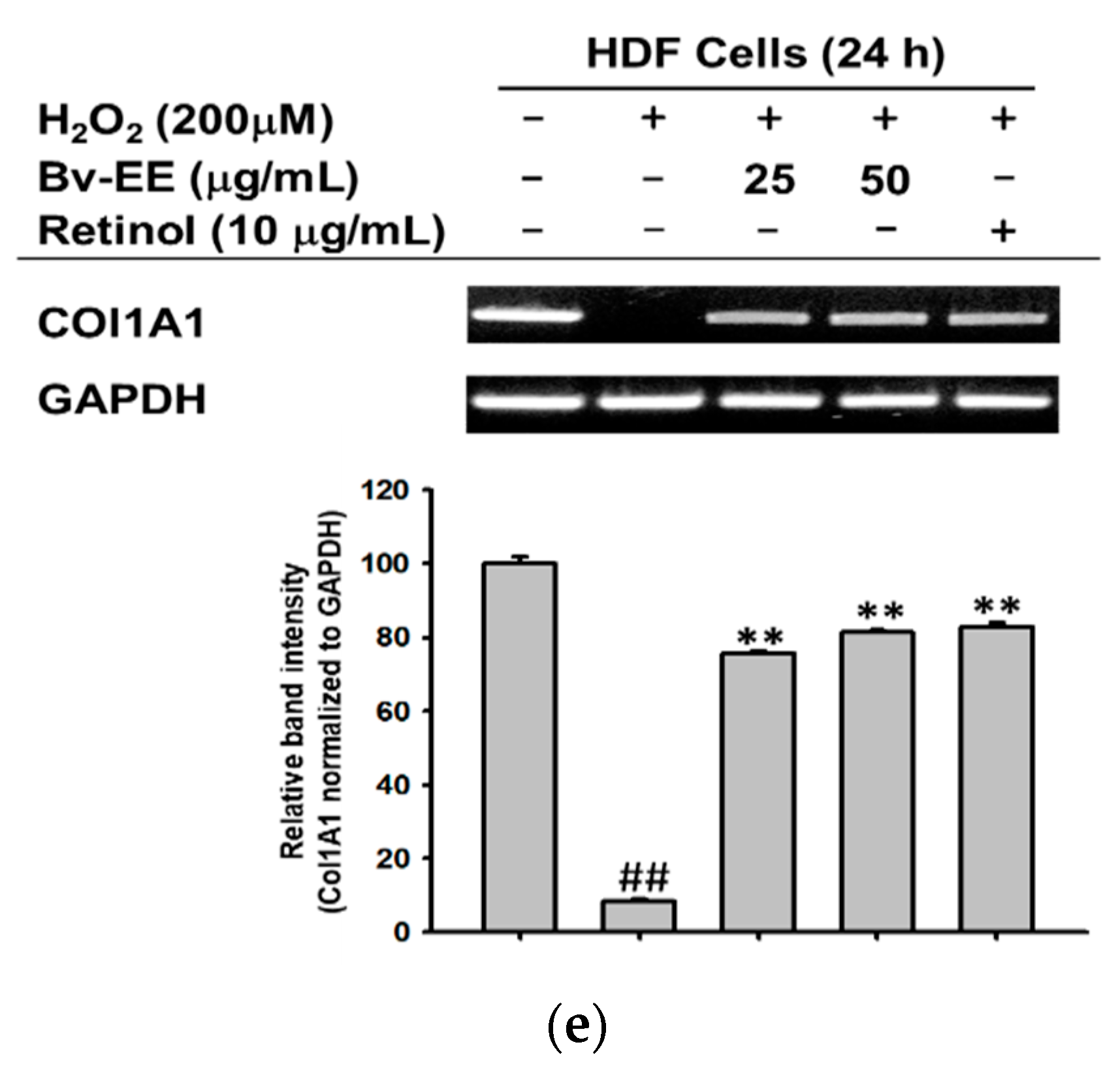
2.6. Bv-EE Promotes Collagen Generation in Human Dermal Fibroblast (HDF) Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Breynia vitis-idaea
4.3. GC–MS
4.4. Cell Culture
4.5. DPPH Radical Scavenging Activity Assay
4.6. ABTS Radical Scavenging Activity Assay
4.7. CUPRAC Assay
4.8. FRAP Assay
4.9. Determination of Total Phenolic Content
4.10. Determination of Total Flavonoid Content
4.11. ROS Generation Assay
4.12. Cell Viability Assay
4.13. H2O2 Treatment and UVB Irradiation
4.14. Semi-Quantitative Reverse Transcription-PCR (RT-PCR) and Quantitative Real-Time PCR (Real-Time PCR)
4.15. Luciferase Reporter Gene Assay
4.16. Preparation of Whole Cell Lysates and Western Blot Analysis
4.17. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kawakita, A.; Kato, M. Obligate pollination mutualism in Breynia (Phyllanthaceae): Further documentation of pollination mutualism involving Epicephala moths (Gracillariidae). Am. J. Bot. 2004, 91, 1319–1325. [Google Scholar] [CrossRef]
- Onyegbule, F.A.; Ilouno, I.O.; Eze, P.M.; Abba, C.C.; Chigozie, V.U. Evaluation of the analgesic, anti-inflammatory and antimicrobial activities of leaf extracts of Breynia nivosa. Chem. Sci. Rev. Lett. 2014, 3, 1126–1134. [Google Scholar]
- Murugan, R.; Prabu, J.; Chandran, R.; Sajeesh, T.; Iniyavan, M.; Parimelazhagan, T. Nutritional composition, in vitro antioxidant and anti-diabetic potentials of Breynia retusa (Dennst.) Alston. Food Sci. Hum. Wellness 2016, 5, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Saadullah, M.; Asif, M.; Uzair, M.; Afzal, S.; Rashid, S.A.; Rashad, M.; Bashir, R.; Mahmood, S. Pharmacological evaluation of the hypoglycemic and anti-Alzheimer’s activities of aerial parts of Breynia distachia (Phyllanthaceae). Trop. J. Pharm. Res. 2021, 21, 579–587. [Google Scholar] [CrossRef]
- Nguyen, T.-M.; Le, X.-T.; Nguyen, M.-T.K. Chemical constituents of Breynia vitis-idaea (Burm. f.) CEC Fischer. In AIP Conference Proceedings: 2017; AIP Publishing LLC: Melville, NY, USA, 2017; p. 020041. [Google Scholar]
- Jeyasankar, A.; Ramar, G. Larvicidal properties of Breyenia vitis-idaea (Burm. f.) Fischer (Euphorbiaceae) against important vector mosquitoes (Diptera: Culicidae). J. Vector Borne Dis. 2014, 51, 239. [Google Scholar]
- Meng, D.-H.; Wu, J.; Wang, L.-Y.; Zhao, W.-M. Two new glycosides from Breynia vitis-idaea. J. Asian Nat. Prod. Res. 2010, 12, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Nagar, J.C.; Chauhan, L.S. Evaluation of antihyperglycemic and antihyperlipidemic activity of leaf extracts of Breynia vitis-idaea in alloxan induced diabetic rats. Pharmacogn. J. 2016, 8, 259–263. [Google Scholar] [CrossRef] [Green Version]
- FISHER, C. Anti-cancer activity of aqueous and ethanol extracts of Breynia vistis-idaea (Burm.F) C. Fisher leaves by using HEPG2 cell line. World J. Pharm. Pharm Sci. 2014, 4, 830–840. [Google Scholar]
- Wang, Y.; Li, W.; Xu, S.; Hu, R.; Zeng, Q.; Liu, Q.; Li, S.; Lee, H.; Chang, M.; Guan, L. Protective skin aging effects of cherry blossom extract (Prunus yedoensis) on oxidative stress and apoptosis in UVB-irradiated HaCaT cells. Cytotechnology 2019, 71, 475–487. [Google Scholar] [CrossRef]
- You, L.; Cho, J.Y. The regulatory role of Korean ginseng in skin cells. J. Ginseng Res. 2021, 45, 363–370. [Google Scholar] [CrossRef]
- Maru, G.B.; Gandhi, K.; Ramchandani, A.; Kumar, G. The role of inflammation in skin cancer. Adv. Exp. Med. Biol. 2014, 816, 437–469. [Google Scholar]
- Lorz, L.R.; Kim, D.; Kim, M.Y.; Cho, J.Y. Panax ginseng-derived fraction BIOGF1K reduces atopic dermatitis responses via suppression of mitogen-activated protein kinase signaling pathway. J. Ginseng Res. 2020, 44, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Grillo, E.; Corsini, M.; Ravelli, C.; Zammataro, L.; Bacci, M.; Morandi, A.; Monti, E.; Presta, M.; Mitola, S. Expression of activated VEGFR2 by R1051Q mutation alters the energy metabolism of Sk-Mel-31 melanoma cells by increasing glutamine dependence. Cancer Lett. 2021, 507, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Kammeyer, A.; Luiten, R. Oxidation events and skin aging. Ageing Res. Rev. 2015, 21, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Joo, Y.H.; Karadeniz, F.; Ko, J.; Kong, C.-S. Syringaresinol inhibits UVA-induced MMP-1 expression by suppression of MAPK/AP-1 signaling in HaCaT keratinocytes and human dermal fibroblasts. Int. J. Mol. Sci. 2020, 21, 3981. [Google Scholar] [CrossRef]
- Chang, H.; Zou, Z.; Li, J.; Shen, Q.; Liu, L.; An, X.; Yang, S.; Xing, D. Photoactivation of mitochondrial reactive oxygen species-mediated Src and protein kinase C pathway enhances MHC class II-restricted T cell immunity to tumours. Cancer Lett. 2021, 523, 57–71. [Google Scholar] [CrossRef]
- Bode, A.M.; Dong, Z. Mitogen-activated protein kinase activation in UV-induced signal transduction. Sci. STKE 2003, 2003, re2. [Google Scholar] [CrossRef]
- Lee, M.E.; Kim, S.R.; Lee, S.; Jung, Y.-J.; Choi, S.S.; Kim, W.J.; Han, J.A. Cyclooxygenase-2 inhibitors modulate skin aging in a catalytic activity-independent manner. Exp. Mol. Med. 2012, 44, 536–544. [Google Scholar] [CrossRef]
- Buckman, S.; Gresham, A.; Hale, P.; Hruza, G.; Anast, J.; Masferrer, J.; Pentland, A.P. COX-2 expression is induced by UVB exposure in human skin: Implications for the development of skin cancer. Carcinogenesis 1998, 19, 723–729. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.H.; Cho, S.; Lee, S.; Kim, K.H.; Cho, K.H.; Eun, H.C.; Chung, J.H. Photoprotective and anti-skin-aging effects of eicosapentaenoic acid in human skin in vivo. J. Lipid Res. 2006, 47, 921–930. [Google Scholar] [CrossRef] [Green Version]
- Devary, Y.; Rosette, C.; DiDonato, J.A.; Karin, M. NF-κB activation by ultraviolet light not dependent on a nuclear signal. Science 1993, 261, 1442–1445. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Guo, J.-H.; Tu, X.-L.; Zhang, C.; Zhao, M.; Zhang, Q.-W.; Gao, F.-H. Tiron inhibits UVB-induced AP-1 binding sites transcriptional activation on MMP-1 and MMP-3 promoters by MAPK signaling pathway in human dermal fibroblasts. PLoS ONE 2016, 11, e0159998. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.H.; Shin, C.M.; Park, C.-H.; Kim, K.H.; Cho, K.H.; Eun, H.C.; Chung, J.H. Eicosapentaenoic acid inhibits UV-induced MMP-1 expression in human dermal fibroblasts. J. Lipid Res. 2005, 46, 1712–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.-O.; Baltes, W. On the role of 2, 3-dihydro-3, 5-dihydroxy-6-methyl-4 (H)-pyran-4-one in the Maillard reaction. J. Agric. Food Chem. 1996, 44, 282–289. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, Q.; Zhao, Z.; Bai, B.; Sun, Z.; Cai, L.; Fu, Y.; Ma, Y.; Wang, Q.; Xi, G. Effect of hydroxyl on antioxidant properties of 2, 3-dihydro-3, 5-dihydroxy-6-methyl-4 H-pyran-4-one to scavenge free radicals. RSC Adv. 2021, 11, 34456–34461. [Google Scholar] [CrossRef]
- Yu, X.; Zhao, M.; Liu, F.; Zeng, S.; Hu, J. Identification of 2, 3-dihydro-3, 5-dihydroxy-6-methyl-4H-pyran-4-one as a strong antioxidant in glucose–histidine Maillard reaction products. Food Res. Int. 2013, 51, 397–403. [Google Scholar] [CrossRef]
- Xie, Q.; Gu, X.; Chen, J.; Liu, M.; Xiong, F.; Wu, X.; Zhang, Y.; Chen, F.; Chen, H.; Li, M. Soyasaponins reduce inflammation and improve serum lipid profiles and glucose homeostasis in high fat diet-induced obese mice. Mol. Nutr. Food Res. 2018, 62, 1800205. [Google Scholar] [CrossRef]
- Kedare, S.B.; Singh, R. Genesis and development of DPPH method of antioxidant assay. J. Food Sci. Technol. 2011, 48, 412–422. [Google Scholar] [CrossRef] [Green Version]
- Roebuck, K.A. Oxidant stress regulation of IL-8 and ICAM-1 gene expression: Differential activation and binding of the transcription factors AP-1 and NF-kappaB. Int. J. Mol. Med. 1999, 4, 223–253. [Google Scholar] [CrossRef]
- Benbow, U.; Brinckerhoff, C.E. The AP-1 site and MMP gene regulation: What is all the fuss about? Matrix Biol. 1997, 15, 519–526. [Google Scholar] [CrossRef]
- Ding, M.; Zhao, J.; Bowman, L.; Lu, Y.; Shi, X. Inhibition of AP-1 and MAPK signaling and activation of Nrf2/ARE pathway by quercitrin. Int. J. Oncol. 2010, 36, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-T.; Lee, S.-Y.; Jeong, J.-H.; Jo, B.-K. New anti-wrinkle cosmetics. J. Soc. Cosmet. Sci. Korea 2002, 28, 71–79. [Google Scholar]
- Wenk, J.; Brenneisen, P.; Meewes, C.; Wlaschek, M.; Peters, T.; Blaudschun, R.; Ma, W.; Kuhr, L.; Schneider, L.; Scharffetter-Kochanek, K. UV-induced oxidative stress and photoaging. Curr. Probl. Dermatol. 2001, 29, 83–94. [Google Scholar] [PubMed]
- Harris, A.K.; Stopak, D.; Wild, P. Fibroblast traction as a mechanism for collagen morphogenesis. Nature 1981, 290, 249–251. [Google Scholar] [CrossRef]
- Redolfi Riva, E.; Micera, S. Progress and challenges of implantable neural interfaces based on nature-derived materials. Bioelectron. Med. 2021, 7, 1–10. [Google Scholar] [CrossRef]
- Birch-Machin, M.; Bowman, A. Oxidative stress and ageing. Br. J. Dermatol. 2016, 175, 26–29. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B. Antioxidant characterization: Methodology and mechanism. Biochem. Pharmacol. 1995, 49, 1341–1348. [Google Scholar] [CrossRef]
- Ou, S.; Kwok, K.C. Ferulic acid: Pharmaceutical functions, preparation and applications in foods. J. Sci. Food Agric. 2004, 84, 1261–1269. [Google Scholar] [CrossRef]
- Seibert, K.; Masferrer, J. Role of inducible cyclooxygenase (COX-2) in inflammation. Receptor 1994, 4, 17–23. [Google Scholar]
- Pillai, S.; Oresajo, C.; Hayward, J. Ultraviolet radiation and skin aging: Roles of reactive oxygen species, inflammation and protease activation, and strategies for prevention of inflammation-induced matrix degradation–A review. Int. J. Cosmet. Sci. 2005, 27, 17–34. [Google Scholar] [CrossRef]
- Borg, M.; Brincat, S.; Camilleri, G.; Schembri-Wismayer, P.; Brincat, M.; Calleja-Agius, J. The role of cytokines in skin aging. Climacteric 2013, 16, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Hwang, B.M.; Noh, E.M.; Kim, J.S.; Kim, J.M.; You, Y.O.; Hwang, J.K.; Kwon, K.B.; Lee, Y.R. Curcumin inhibits UVB-induced matrix metalloproteinase-1/3 expression by suppressing the MAPK-p38/JNK pathways in human dermal fibroblasts. Exp. Dermatol. 2013, 22, 371–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Schattenberg, J.r.M.; Rigoli, R.M.; Storz, P.; Czaja, M.J. Hepatocyte resistance to oxidative stress is dependent on protein kinase C-mediated down-regulation of c-Jun/AP-1. J. Biol. Chem. 2004, 279, 31089–31097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacKenna, D.; Summerour, S.R.; Villarreal, F.J. Role of mechanical factors in modulating cardiac fibroblast function and extracellular matrix synthesis. Cardiovasc. Res. 2000, 46, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Reilly, D.M.; Lozano, J. Skin collagen through the lifestages: Importance for skin health and beauty. Plast. Aesthet. Res. 2021, 8, 2. [Google Scholar] [CrossRef]
- Lee, H.P.; Kim, D.S.; Park, S.H.; Shin, C.Y.; Woo, J.J.; Kim, J.W.; An, R.-B.; Lee, C.; Cho, J.Y. Antioxidant capacity of Potentilla paradoxa nutt. and its beneficial effects related to anti-aging in hacat and B16F10 cells. Plants 2022, 11, 873. [Google Scholar] [CrossRef]
- Jang, W.Y.; Lee, H.P.; Kim, S.A.; Huang, L.; Yoon, J.H.; Shin, C.Y.; Mitra, A.; Kim, H.G.; Cho, J.Y. Angiopteris cochinchinensis de vriese ameliorates lps-induced acute lung injury via src inhibition. Plants 2022, 11, 1306. [Google Scholar] [CrossRef]
- Kim, J.W.; Kwon, K.W.; Kim, M.Y.; Cho, J.Y. Potentilla paradoxa Nutt. Ethanol Extract Exhibits Anti-Inflammatory Effects by Suppression of the Src/NF-kappaB Signaling Pathway. Plants 2022, 11, 1750. [Google Scholar] [CrossRef]
- Oh, J.; Kim, S.A.; Kwon, K.W.; Choi, S.R.; Lee, C.H.; Hossain, M.A.; Kim, E.S.; Kim, C.; Lee, B.H.; Lee, S.; et al. Sophora flavescens Aiton methanol extract exerts anti-inflammatory effects via reduction of Src kinase phosphorylation. J. Ethnopharmacol. 2023, 305, 116015. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.-E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Leb. Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Hwang, S.-H.; Kim, J.H.; Choi, E.; Park, S.H.; Cho, J.Y. Antioxidative and Skin Protective Effects of Canarium subulatum Methanol Extract on Keratinocytes. Evid. Based Complement. Alternat. Med. 2021, 2021, 6692838. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Apak, R.; Güçlü, K.; Özyürek, M.; Bener, M. Cupric ion reducing antioxidant capacity assay for food antioxidants: Vitamins, polyphenolics, and flavonoids in food extracts. In Advanced Protocols in Oxidative Stress I; Springer: Berlin/Heidelberg, Germany, 2008; pp. 163–193. [Google Scholar]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlesic, T.; Poljak, S.; Misetic Ostojic, D.; Lucin, I.; Reynolds, C.A.; Kalafatovic, D.; Saftic Martinovic, L. Mint (Mentha spp.) Honey: Analysis of the Phenolic Profile and Antioxidant Activity. Food Technol. Biotechnol. 2022, 60, 509–519. [Google Scholar] [CrossRef]
- Rahmawati, L.; Aziz, N.; Oh, J.; Hong, Y.H.; Woo, B.Y.; Hong, Y.D.; Manilack, P.; Souladeth, P.; Jung, J.H.; Lee, W.S.; et al. Cissus subtetragona Planch. Ameliorates Inflammatory Responses in LPS-induced Macrophages, HCl/EtOH-induced Gastritis, and LPS-induced Lung Injury via Attenuation of Src and TAK1. Molecules 2021, 26, 6073. [Google Scholar] [CrossRef] [PubMed]
- Ha, A.T.; Rahmawati, L.; You, L.; Hossain, M.A.; Kim, J.-H.; Cho, J.Y. Anti-inflammatory, antioxidant, moisturizing, and antimelanogenesis effects of quercetin 3-O-β-D-glucuronide in human keratinocytes and melanoma cells via activation of NF-κB and AP-1 pathways. Int. J. Mol. Sci. 2021, 23, 433. [Google Scholar] [CrossRef]
- Shin, K.K.; Park, S.H.; Lim, H.Y.; Lorza, L.R.; Qomaladewia, N.P.; You, L.; Aziz, N.; Kim, S.A.; Lee, J.S.; Choung, E.S. In Vitro Anti-Photoaging and Skin Protective Effects of Licania macrocarpa Cuatrec Methanol Extract. Plants 2022, 11, 1383. [Google Scholar] [CrossRef]
- Hong, Y.H.; Aziz, N.; Park, J.G.; Lee, D.; Kim, J.K.; Kim, S.A.; Choi, W.; Lee, C.Y.; Lee, H.P.; Huyen Trang, H.T.; et al. The EEF1AKMT3/MAP2K7/TP53 axis suppresses tumor invasiveness and metastasis in gastric cancer. Cancer Lett. 2022, 544, 215803. [Google Scholar] [CrossRef]
- Song, C.; Lorz, L.R.; Lee, J.; Cho, J.Y. In vitro photoprotective, anti-inflammatory, moisturizing, and antimelanogenic effects of a methanolic extract of Chrysophyllum lucentifolium Cronquist. Plants 2021, 11, 94. [Google Scholar] [CrossRef]
- Aziz, N.; Son, Y.-J.; Cho, J.Y. Thymoquinone suppresses IRF-3-mediated expression of type I interferons via suppression of TBK1. Int. J. Mol. Sci. 2018, 19, 1355. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-O.; Kim, E.; Kim, J.H.; Hong, Y.H.; Kim, H.G.; Jeong, D.; Kim, J.; Kim, S.H.; Park, C.; Seo, D.B. Antimelanogenesis and skin-protective activities of Panax ginseng calyx ethanol extract. J. Ginseng Res. 2018, 42, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.; Kang, Y.-G.; Hwang, S.-H.; Kim, J.K.; Hong, Y.D.; Park, W.-S.; Kim, D.; Kim, E.; Cho, J.Y. In vitro effects of dehydrotrametenolic acid on skin barrier function. Molecules 2019, 24, 4583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Jang, J.; Park, J.G.; Kim, K.-H.; Yoon, K.; Yoo, B.C.; Cho, J.Y. Protein arginine methyltransferase 1 (PRMT1) selective inhibitor, TC-E 5003, has anti-inflammatory properties in TLR4 signaling. Int. J. Mol. Sci. 2020, 21, 3058. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Kim, M.-Y.; Cho, J.Y. Protective effect of Potentilla glabra in UVB-induced photoaging process. Molecules 2021, 26, 5408. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak NO. | RT | Name of the Compound | Peak Height | Corr. Area | Corr. % Max | % of Total |
|---|---|---|---|---|---|---|
| 20 | 7.621 | 2,3-Dihydro-3,5-dihydroxy-6-methyl-4H-pyran-4-one | 876,767 | 17,904,906 | 100.00% | 13.46% |
| 28 | 9.349 | Lyxitol, 1-O-hexyl | 149,742 | 13,972,799 | 78.04% | 10.50% |
| 29 | 9.492 | 3-(2-Hydroxyethyl)imidazole-2-thione | 285,772 | 13,749,668 | 76.79% | 10.34% |
| 36 | 11.616 | Carbamic acid | 146,325 | 10,905,600 | 60.91% | 8.20% |
| 2 | 1.767 | Acetic acid | 251,106 | 8,966,954 | 50.08% | 6.74% |
| 26 | 8.985 | 1,2,3-Propanetriol 1-acetate | 241,168 | 7,172,538 | 40.06% | 5.39% |
| 4 | 2.185 | 2-Propenoic acid | 151,545 | 5,636,432 | 31.48% | 4.24% |
| 37 | 12.666 | Benzoic acid, 4-hydroxy- | 54,972 | 5,252,489 | 29.34% | 3.95% |
| 45 | 29.272 | Hexestrol | 111,097 | 4,721,710 | 26.37% | 3.55% |
| 40 | 13.965 | Ethyl .beta.-d-riboside | 99,327 | 3,979,699 | 22.23% | 2.99% |
| 30 | 9.606 | 1,8-Diamino-3,6-dioxaoctane | 70,016 | 3,407,960 | 19.03% | 2.56% |
| 41 | 14.127 | 4-O-beta-D-galactopyranosyl | 59,056 | 3,146,589 | 17.57% | 2.37% |
| 8 | 3.874 | Butanoic acid | 78,568 | 3,075,157 | 17.17% | 2.31% |
| 17 | 6.582 | 1,3,5-Triazine-2,4,6- | 95,026 | 2,726,316 | 15.23% | 2.05% |
| 19 | 7.497 | Ethanamine | 52,324 | 2,486,491 | 13.89% | 1.87% |
| 21 | 7.803 | Benzoic acid | 34,833 | 2,246,052 | 12.54% | 1.69% |
| 44 | 23.777 | 2-(Acetoxymethyl)-3-(methoxycarbonyl)biphenylene | 81,417 | 1,969,340 | 11.00% | 1.48% |
| 6 | 2.721 | 2-Propanone, 1-hydroxy- | 90,883 | 1,732,730 | 9.68% | 1.30% |
| 9 | 4.031 | Dihydroxyacetone | 39,401 | 1,686,793 | 9.42% | 1.27% |
| 24 | 8.821 | 5-Hydroxymethylfurfural | 37,226 | 1,395,927 | 7.80% | 1.05% |
| GC | Column | HP-5MS UI, 30 m × 250 μm × 0.25 μm |
| Injection volume | 1 μL of 100 mg/mL | |
| Inlet temperature | 250 °C | |
| Injection mode | Split, 50:1 | |
| Carrier gas | He, 1 mL/min | |
| Oven temperature | 50 °C → heating rate 10 °C/min → 300 °C (hold 15 min) | |
| Aux heater temperature | 300 °C | |
| MS | Ionization mode | Electron ionization |
| Detection temperature | 230 °C | |
| Quadrupole temperature | 150 °C | |
| Detection mode | Scan, m/z:33–150 |
| PCR Type | Gene Name | Sequence (5′-3′) | |
|---|---|---|---|
| RT-PCR (human) | Col1A1 | Forward | CAGGTACCATGACCGAGACG |
| Reverse | AGCACCATCATTTCCACGAG | ||
| GAPDH | Forward | GCACCGTCAAGGCTGAGAAC | |
| Reverse | ATGGTGGTGAAGACGCCAGT | ||
| Real-time PCR (human) | COX-2 | Forward | CAGCATTGTAAAGTTGGTGGACTGT |
| Reverse | GGGATTTTGGAACGTTGTGAA | ||
| MMP3 | Forward | TGTTAGGAGAAAGGACAGTGGTC | |
| Reverse | CGTCACCTCCAATCCAAGGA | ||
| MMP9 | Forward | GCCACTTGTCGGCGATAAGG | |
| Reverse | CACTGTCCACCCCTCAGAGC | ||
| GAPDH | Forward | GCGCCCAATACGACCAAATC | |
| Reverse | GACAGTCAGCCGCATCTTCT | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, C.Y.; Jang, J.; Lee, H.P.; Park, S.H.; Kry, M.; Keo, O.; Lee, B.-H.; Choi, W.; Lee, S.; Cho, J.Y. Anti-Oxidative and Anti-Aging Effects of Ethanol Extract of the Officinal Breynia (Breynia vitis-idaea) In Vitro. Plants 2023, 12, 1088. https://doi.org/10.3390/plants12051088
Shin CY, Jang J, Lee HP, Park SH, Kry M, Keo O, Lee B-H, Choi W, Lee S, Cho JY. Anti-Oxidative and Anti-Aging Effects of Ethanol Extract of the Officinal Breynia (Breynia vitis-idaea) In Vitro. Plants. 2023; 12(5):1088. https://doi.org/10.3390/plants12051088
Chicago/Turabian StyleShin, Chae Yun, Jiwon Jang, Hwa Pyoung Lee, Sang Hee Park, Masphal Kry, Omaliss Keo, Byoung-Hee Lee, Wooram Choi, Sarah Lee, and Jae Youl Cho. 2023. "Anti-Oxidative and Anti-Aging Effects of Ethanol Extract of the Officinal Breynia (Breynia vitis-idaea) In Vitro" Plants 12, no. 5: 1088. https://doi.org/10.3390/plants12051088
APA StyleShin, C. Y., Jang, J., Lee, H. P., Park, S. H., Kry, M., Keo, O., Lee, B. -H., Choi, W., Lee, S., & Cho, J. Y. (2023). Anti-Oxidative and Anti-Aging Effects of Ethanol Extract of the Officinal Breynia (Breynia vitis-idaea) In Vitro. Plants, 12(5), 1088. https://doi.org/10.3390/plants12051088