2. Results
2.1. Foliar Anatomical Description of the Laelia Group from Mexico
Dermal Tissue. Adaxial cuticle generally striated, except in L. rubescens and L. halbingeriana, which is smooth; adaxial cuticle thickness from 5 to 11 μm (<12 μm); except from L. rubescens, which is >12 μm, abaxial cuticle thickness 5–14 μm. Adaxial epidermal cells distributed in rows in L. gouldiana, L. rubescens and L. halbingeriana, the rest of the species with alternately distributed cells, the abaxial epidermal cells are in rows. Adaxial and abaxial epidermal cells are generally isodiametric polygonal in shape, except L. eyermaniana, L. rubescens and L. superbiens, where they are elongated polygonal. Area of adaxial epidermal cells <5000 μm2, cells abaxial from 801 to 1400 μm2, with the exception of L. aurea, where it is from 600 to 800 μm2. Stomata only in the abaxial epidermis, distributed in rows, except in L. albida and L. autumnalis, L. furfuracea, L. superbiens and L. halbingeriana, which is alternate, trichomes present at the margins of the adaxial surface, denoted by the permanence of basal cells, except in L. albida, L. crawshayana, L. eyermaniana and L. halbingeriana. Papillae present in the abaxial epidermis, except in L. albida, L. eyermaniana, L. furfuracea and L. halbingeriana, the papillae are epidermal, except in L. anceps and L. speciosa, where they are cuticular. The length of the guard cells is 30 to 40 μm, except in L. superbiens, which is larger than 45 μm. The development of the cuticular ridges is light in L. albida, L. anceps, L. eyermaniana and L. halbingeriana and deep in the rest of the species. The substomatal chamber is conspicuous, with the exception of L. aurea, L. rubescens and L. halbingeriana, which is inconspicuous. Stomata are generally semi-sunken, with the exception of that in L. autumnalis, L. eyermaniana, L. furfuracea and L. speciosa that are at the level of the epidermis and in L. aurea that are sunk. Stomatal index from 2 to 6. Height of adaxial epidermal cells 18–24 μm, except in L. albida, L. anceps, L. autumnalis, L. furfuracea and L. gouldiana ranging from 24 to 30 μm. Cell width adaxial epidermal 23–27 μm in L. anceps and L. crawshayana, 28–31 μm in L. aurea and in L. furfuracea, L. speciosa and L. superbiens, it Is 32–35 μm. Height of abaxial epidermal cells less than 30 μm as in Laelia furfuracea. Width of abaxial epidermal cells 25–37 μm as in L. aurea, except in L. speciosa which is 38 to 41 μm. Fundamental tissue. Hypodermis. Present in the leaf of all species, located both on the adaxial surface as abaxial, with the exception of L. speciosa which only occurs in the adaxial surface. Species with both hypodermises have a stratum in its abaxial hypodermis, the adaxial hypodermis generally presents one layer, with the exception of that in L. superbiens, which has two layers. Cell shape of the adaxial hypodermis is oblong except for L. rubescens, which presents cells with an elongated polygonal shape, and L. speciosa with polygonal cells isodiametric. Shape of cells of hypodermis abaxial, polygonal isodiametric except in the case of L. anceps, L. superbiens and L. halbingeriana, which are oblongs, and L. Rubescens that presents cells with a polygonal shape perpendicularly elongated. Thick cell walls in the adaxial hypodermis, except in L. crawshayana and L. rubescens, which are thin. In the hypodermis abaxial the walls are in some species thick and in others thin. The type of wall in both the adaxial and abaxial hypodermis is cellulosic, except in L. superbiens and L. halbingeriana, which is lignified. The distribution of the layers of the adaxial hypodermis is continuous for all species, while the distribution in the abaxial hypodermis is continuous only in some species, with the exception of that in L. crawshayana, L. aurea, L. anceps, L. eyermaniana and L. speciosa, which is discontinuous. On the other hand, in the basal region, the presence of hypodermis was found in all species. The location is on both surfaces (adaxial/abaxial), with the exception of L. eyermaniana, where it is abaxial only. There is one layer of the adaxial hypodermis, with the exception of L. superbiens and L. crawshayana, which have two or more layers. In the case of the abaxial hypodermis, there is only one layer for all species. Mesophyll with a thickness from 500 to 2500 μm, with L. superbiens being the species with the highest thickness (2500 μm). Heterogeneous mesophyll (palisade/spongy), except in L. autumnalis, L. gouldiana, L. rubescens and L. superbiens, which is homogeneous. The proportion of palisade parenchyma was generally 0 to 0.4, with the exception of that in L. albida, L. anceps and L. aurea, which is from 0.41 to 0.8, and in L. rubescens, L. superbiens and L. halbingeriana, which was 0.81 to 1. The presence of secondary thickenings was found only in the species L. aurea and L. superbiens. Water cells are absent in all species. Presence of calcium oxalate crystals with the exception of L. albida, L. anceps, L. aurea, L. autumnalis and L. eyermaniana. The shape of these crystals is mainly raphides. The ubication of the crystals is mainly in the center of the mesophyll. In the case of the basal region, the mesophyll is heterogeneous (spongy/palisade), with exception of the species that present a homogeneous mesophyll: L. anceps, L. autumnalis, L. eyermaniana, L. gouldiana and L. rubescens. The basal region also presents air spaces in the mesophyll, with the exception of L. crawshayana, L. furfuracea, L. speciosa, L. superbiens and L. halbingeriana. Extravascular fibers are found in the middle region in all species, the location is generally both abaxial and adaxial, with the exception of L. rubescens which has extravascular fibers present throughout the mesophyll. There are one to four layers of adaxial extravascular fiber bundles, with exception of L. rubescens and L. superbiens, which have four to five layers, make abaxial extravascular fibers of two to four layers, with the exception of L. aurea and L. furfuracea, which have four to six layers, and L. rubescens, which has six to nine layers. Extravascular fiber wall thickness of 2.5 μm, except for the species L. aurea, L. gouldiana, L. superbiens and L. halbingeriana with a wall thickness of 5 μm. Extravascular fibers with presence of stegmata. Central zone. In the central zone of the leaves, the presence of bulliform cells was found, with the exception of L. anceps, L. aurea, L. eyermaniana, L. gouldiana and L. halbingeriana. Species with bulliform cells have two to four strata. In a cross section, the leaves of the Mexican species of the genus Laelia present the projected central zone; this projection can be observed generally in convex shape, with the exception of that in L. anceps, L. aurea, L. rubescens and L. superbiens, which is keeled. Most species have a vascular bundle in the central zone. Regarding the presence of extravascular fiber bundles in the central zone, they were evident in all the species and generally had 2 to 4 strata. Vascular tissue. Four types of vascular bundles were found, except for L. crawshayana, L. superbiens and L. halbingeriana presenting three types and L. rubescens with five types of vascular bundles. In the blade, there are generally two layers of vascular bundles as in the case of L. albida, with the exception of L. eyermaniana and L. superbiens with one layer, and L. rubescens with three layers. Across the lamina there are 30 to 90 vascular bundles, with the exception of L. superbiens that presents more than 90 vascular bundles.
2.2. Evaluation of Quantitative Anatomical Characters of the Genus Laelia in Mexico
In particular, the characters evaluated were 11 quantitative anatomical characters with 30 repeated measurements per species. The summary of descriptive statistics for these characters, such as the mean (M) and the standard deviation (SD) are concentrated in
Table 1. Each letter represents a group derived from the post hoc analysis.
2.3. Canonical Discriminant Analysis (CDA)
The CDA result showed that the first three canonical functions contributed 90% of the variation (
Table 2). The eigenvalues and canonical correlation values were high (values greater than 1), which indicates that the evaluated parameters contributed greatly to the discrimination of groups and that the characters that displayed the greatest weight were the area of the abaxial epidermal cells, abaxial cuticle thickness and mesophyll thickness. On the other hand, some characters had values less than 1, in which we could observe that the height of the adaxial epidermal cells and the thickness of the adaxial cuticle could also be functional characters for the separation of the groups.
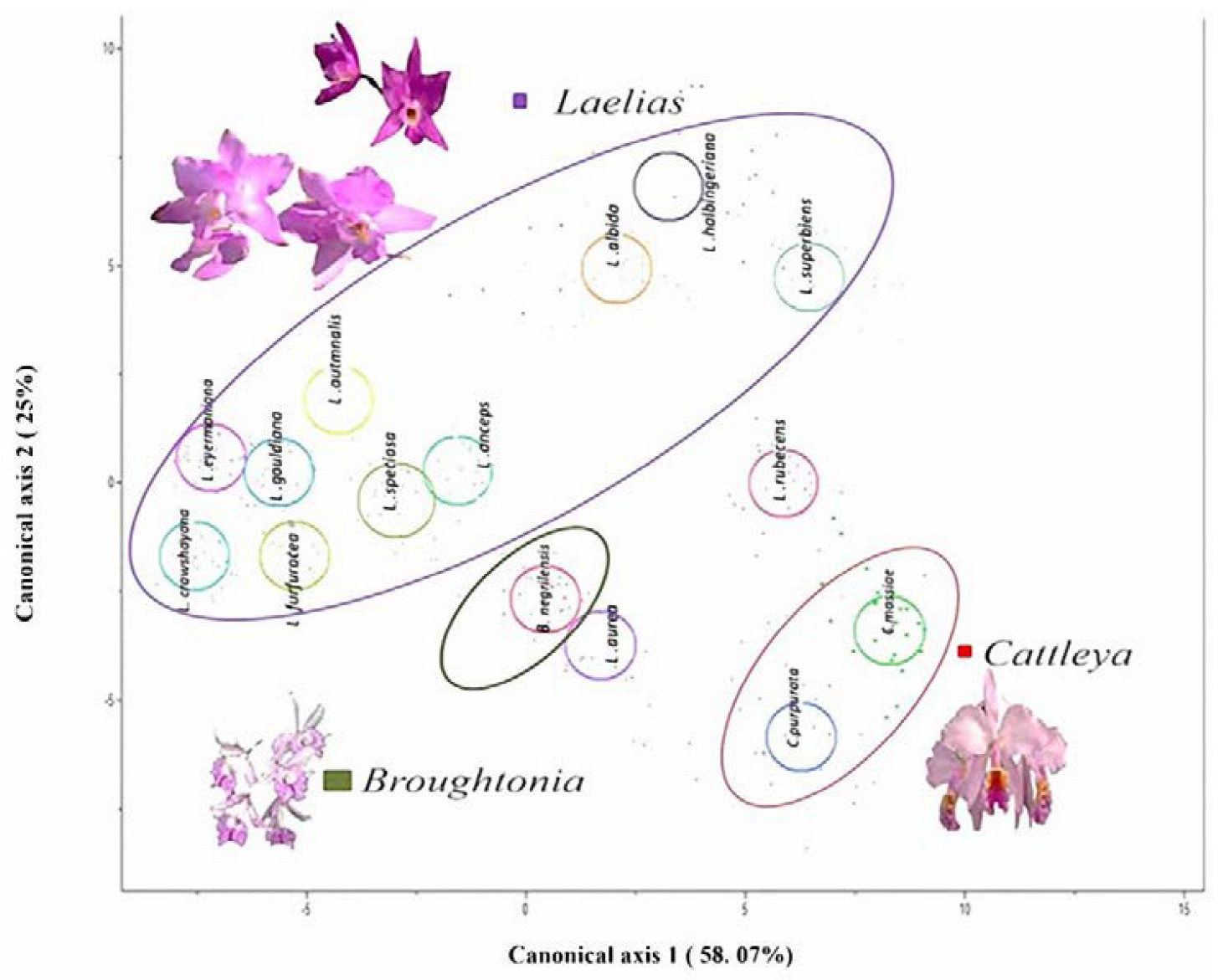
In the canonical discriminant analysis (CDA) (
Figure 1) it was found that in canonical axis 1 (58.07%), the species
Laelia superbiens and the genus
Cattleya displayed a very thick mesophyll (1501 to 2000 µm), while on the left side were the
Laelia species
L.
crawshayana,
L.
eyermaniana,
L.
gouldiana and
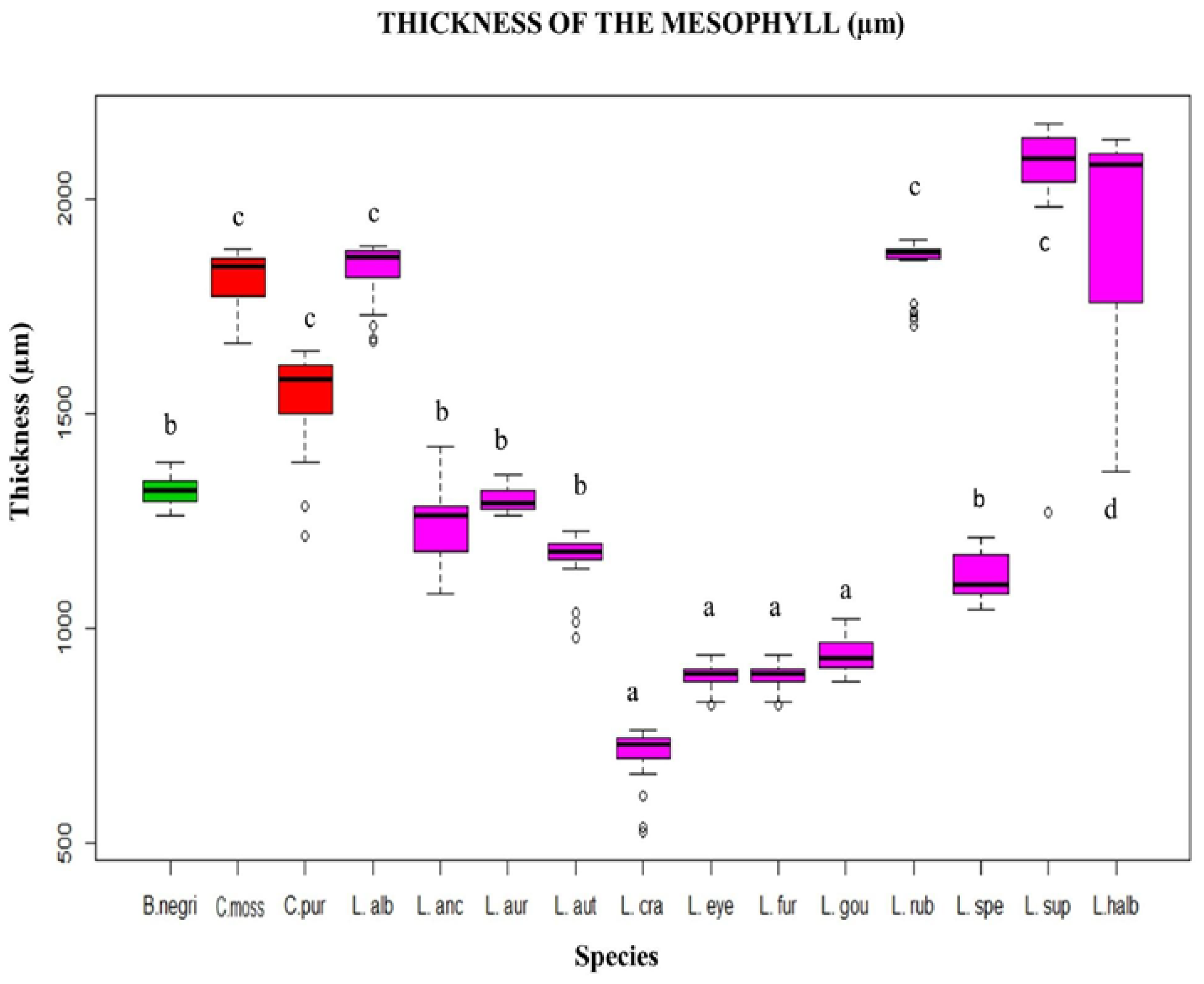
L. furfuracea, with lower thickness of the mesophyll (500 to 1000 µm). This quantitative anatomical character showed variations within the Mexican Laelias species (
Figure 2 and
Figure 3).
In canonical axis 2 (25%), the two species of
Cattleya had a smaller area of abaxial epidermal cells (600 to 800 µm
2) compared to those of
L.
crawshayana,
L.
eyermaniana,
L.
gouldiana,
L.
furfuracea,
L.
speciosa and
L.
anceps (
Figure 1) (800 to 1400 µm
2;
Figure 4 and
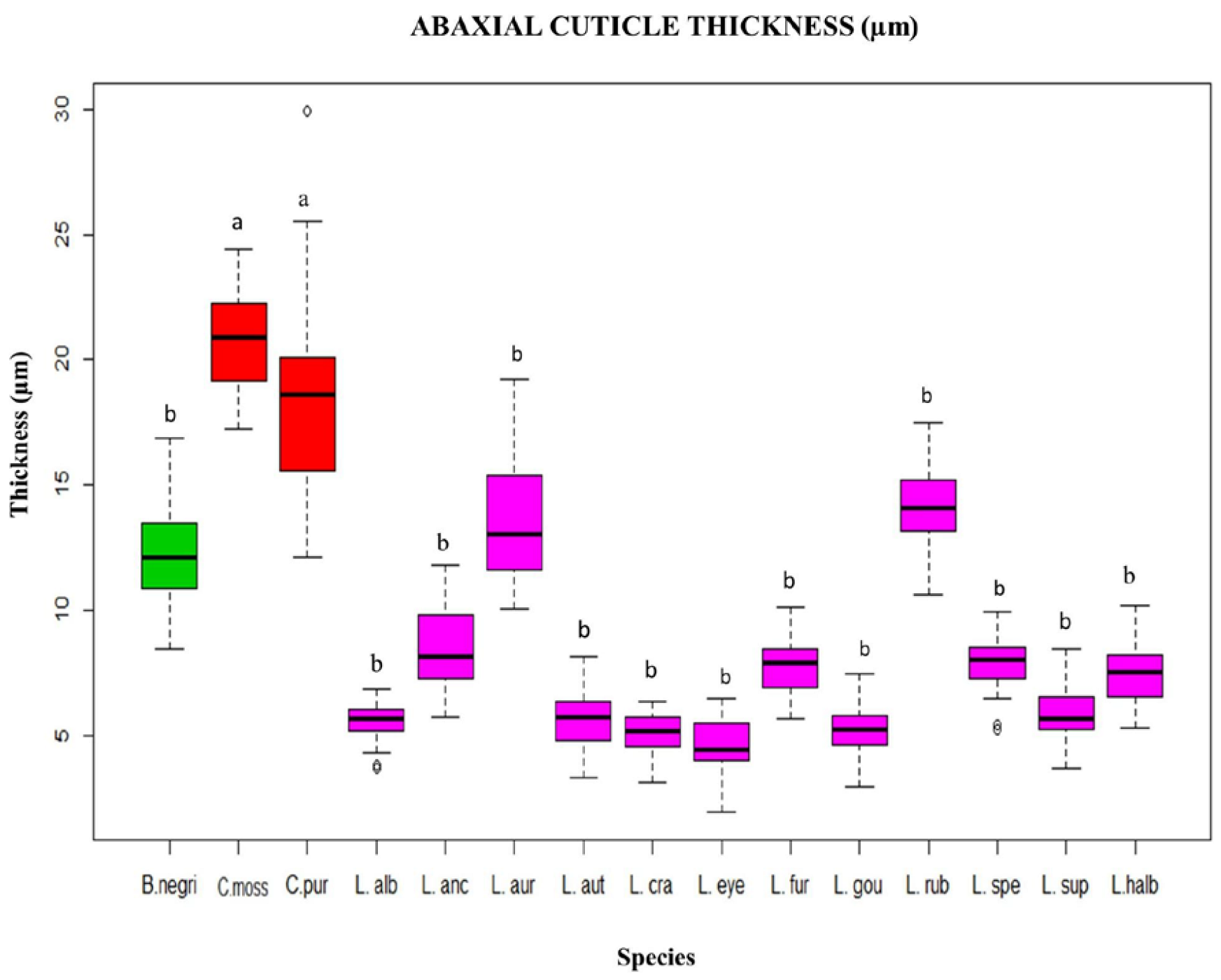
Figure 5). Regarding the thickness of the abaxial cuticle, we could see at the lower end of the canonical axis 2 the genus
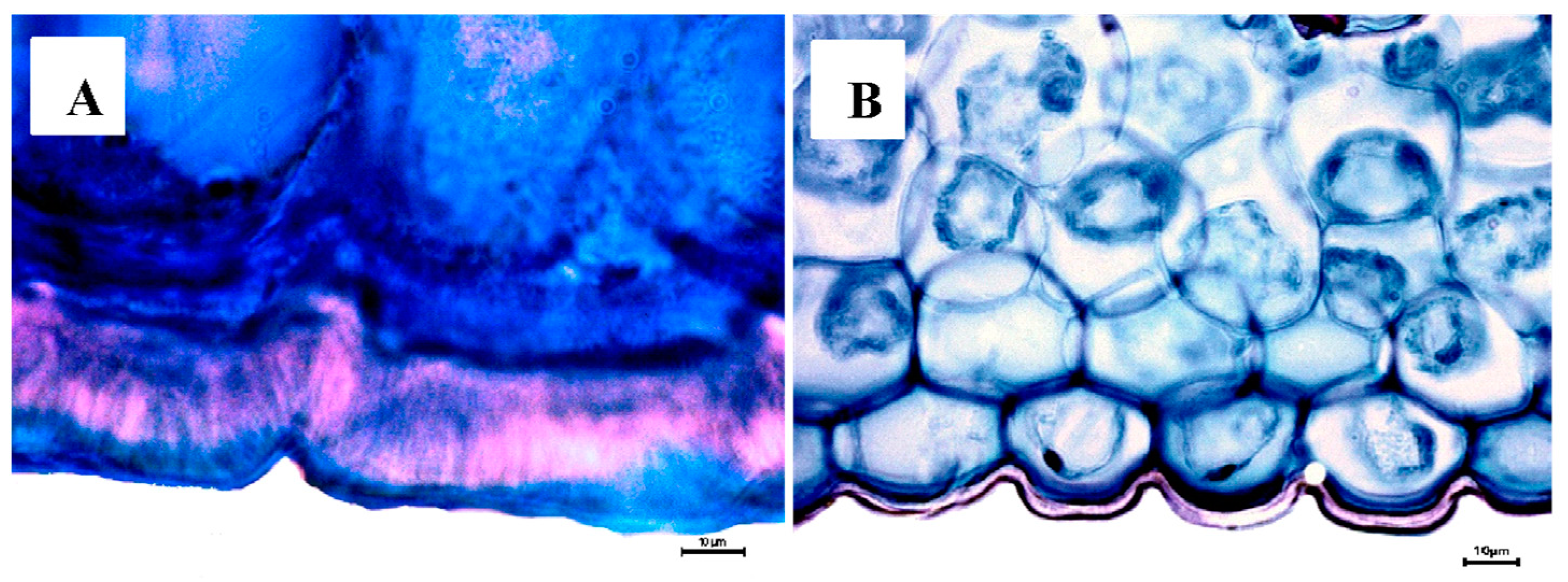
Cattleya with the greatest thickness of the abaxial cuticle (≥18 µm), and at the upper end of the axis, the Mexican species of the genus
Laelia with a smaller abaxial cuticle (≤18 µm;
Figure 6 and
Figure 7).
2.4. Cluster Analysis
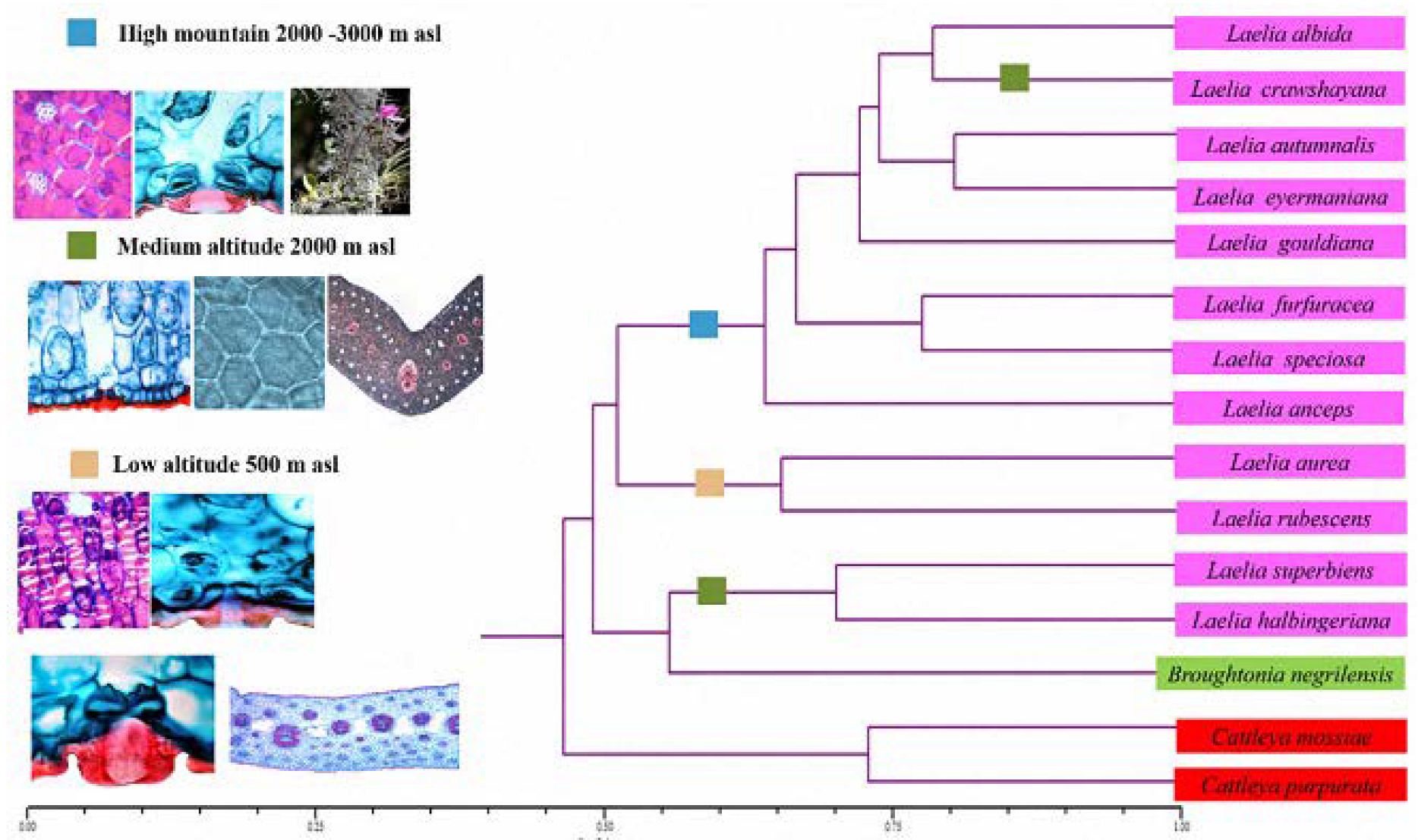
The cluster analysis was obtained using the taxonomic distance coefficient and the average bond (UPGMA) as a grouping algorithm, separating the group of Laelias (pink) from the
Broughtonia (green) and
Cattleya species (red) (
Figure 8). It was found that there was a similarity of structural characters in species that inhabited a certain altitude, particularly for the species of Mexican Laelias.
High-mountain Laelias (2000–3000 m asl), which include the species
Laelia anceps,
L.
albida,
L.
autumnalis,
L.
eyermaniana,
L.
furfuracea,
L.
gouldiana and
L.
speciosa, share the following anatomical characters: striated cuticular surface, conspicuous substomatal chamber, thick cells walls of adaxial hypodermis, distribution of continuous adaxial hypodermis strata, absence of cells with secondary thickenings in the mesophyll and four types of vascular bundles in the mesophyll (based on the number of vascular bundles along the lamina) (
Supplementary Materials, List S1). The morphological characters shared by this group of Laelias are fleshy leathery leaves and sulcate pseudobulbs, glabrous lip, peduncle bracts much shorter than the internodes, sub-distichal arrangement of the flowers, lip throat without a brown spot, stigmatic surface hidden by rostellum and flower arrangement in raceme (
Supplementary Materials, List S2).
At the other extreme are the orchids at low altitudinal distribution (500 m asl), which include Laelia aurea and L. rubescens species. They are similar in the following anatomical characters: epidermal papillae present in the abaxial epidermis of leaves, deep development of cuticular edges of the stoma, inconspicuous substomatal chamber, cellulosic cell wall in the abaxial hypodermis of leaves, presence of secondary thickenings in the mesophyll and hypodermis of the basal region of a stratum and air spaces present in the basal region of the leaf. Interesting morphological characters that can be highlighted for lowland orchids are: chartaceous leathery leaves and pseudobulbs with a discoid shape and more wrinkled, a leaf in each pseudobulb, an exclusively pubescent lip, a lip throat with a brown spot, scale-shaped floral bracts, and a stigmatic surface that is not hidden by the rostellum.
Regarding the species of the analyzed genus Cattleya, in the case of C. mossiae, which is found at altitudes of 800 to 2000 meters asl, and for C. purpurata 500 meters asl, anatomical characters were identified that can be functional for distinguishing them from the genus Laelia, such as: stomatal index (>7), rough cuticle, oblong adaxial and abaxial epidermal cell shape. The morphological characters in both Cattleya species are leaves with proportions greater than 7.41 cm.
3. Discussion
Here we present a study where anatomical and morphological characters were integrated to analyze the similarity relationships and the altitudinal distribution present in the species of the genus Laelia in Mexico. The importance of the results obtained lies in the fact that the present research work shows that the Laelias of México share several structural attributes by which its species can be recognized as a taxonomic group.
Previous studies mention the relevance of the taxonomic relationships that exist between some species of the Laeliinae Subtribe based on their vegetative and floral morphology [
11]. Stern and Carlsward [
12], pointed out that exhaustive anatomical studies in the Laeliinae subtribe are scarce and that plant anatomy is particularly important to understand the relationships of the species in conjunction with their morphology. Baker characterized in detail the morphology and foliar anatomy of 36 genera and 79 taxa of the Laeliinae [
13]. However, he did not include all species of Laelias from México.
This work is the first contribution on the relationship that exists between the thickness of the cuticle and the environment in the Mexican Laelias species. Particularly the species of the genus
Laelia in México, which inhabit more xeric environments, have thicker cuticles as a mechanism of tolerance to drought, as has been reported for other orchid species belonging to xeromorphic environments [
14].
In general, for the species of the Laeliinae subtribe, the shape of the cells in the epidermis is polygonal isodiametric, and their distribution is in rows [
13]. In the case of Mexican Laelias, it was found that they present adaxial and abaxial epidermal cells, generally of an isodiametric polygonal shape; however,
L.
eyermaniana,
L.
rubescens and
L.
superbiens have elongated polygonal cells, and adaxial and abaxial epidermal cells are distributed alternately. In species such as
L.
gouldiana,
L.
rubescens and
L.
halbingeriana, the distribution is in rows.
One of the characters that is not considered in previous studies but quantitatively analyzed in our study was the area of the abaxial and adaxial epidermal cells. In the Mexican Laelias, it was found that the area of the epidermal cells is a character that helps differentiate the Mexican species of the genus Laelia from other species. For example, in the case of Cattleya purpurata, the area of the adaxial epidermal cells is >5000 µm2. In Mexican Laelias, the adaxial epidermal cell area is <5000 µm2, while abaxial cell area is 801 to 1400 µm2, except for L. aurea, where it is 600 to 800 µm2. The area of the abaxial epidermal cells in both species of the genus Cattleya is smaller than in all the Mexican Laelias species with a value of 600 to 800 µm2.
For the Mexican Laelias species, the thickness of the mesophyll was quantitatively measured, and it was found to range from 500 to 2500 µm, with L. superbiens being the species in this study with the greatest thickness. Furthermore, the type of mesophyll is also heterogeneous, except for L. autumnalis, L. gouldiana, L. rubescens and L. superbiens, where it is homogeneous.
Laelia aurea and
L. rubescens are naturally exposed to a water deficit and are the only species with cells with secondary thickenings in the mesophyll. Baker reports the presence of secondary thickenings in the mesophyll as a common character in several species within the Laeliinae subtribe [
13]. Pridgeon mentions that the presence of these cells in the mesophyll can function as additional water reservoirs [
15,
16]. It has been reported that these cells are present in several species of epiphytic orchids of the Oncidiinae subtribe, particularly those that inhabit hot and dry environments [
17]. It is worth mentioning that cells with secondary thickenings in the mesophyll are not a particular character to all the Mexican species of
Laelia, since only two species display them; nevertheless, they were also found in the species of
Broughtonia and
Cattleya, confirming its presence in several species within the Laeliinae subtribe [
13].
Regarding vascular tissue, it has been reported that vascular bundles confer efficient water transport to the plant, necessary to respond to short periods of water availability [
18]. The Mexican Laelias generally present four types of vascular bundles, except for
L. crawshayana,
L. halbingeriana and
L. superbiens, which have three types, and species from more xeric environments such as
L. rubescens that have up to five types of vascular bundles. There are two vascular bundle strata in the lamina, except for
L. eyermaniana and
L. superbiens with one stratum and
L. rubescens with three strata. Throughout the lamina there are 30 to 90 vascular bundles, except in
L. superbiens, which has more than 90 vascular bundles.
It is noteworthy that in this work it was found that species from more xeric environments have a greater number of vascular bundles in the mesophyll, have shorter leaves and are wider. Such is the case of
L. rubescens, which has oblong-elliptical leaves up to 18 cm long and 4 cm wide [
7]. In addition, it is interesting that the species
L.
rubescens that inhabits lowlands and has a high tolerance to drought [
7], exhibits the greatest thickness of the mesophyll as well as three layers of vascular bundles in the mesophyll. On the other hand, the
Laelia species that live in the high mountains have fewer vascular bundles, more elongated and narrower leaves, as well as a less thick mesophyll in contrast to the low-altitude Mexican Laelias.
Plants have mineral inclusions in some of their cells, such as crystals of various kinds. In about 75% of angiosperms, calcium oxalate crystals are the most abundant and are significant in taxonomy [
19]. In Mexican Laelias, calcium oxalate crystals are present, except in
Laelia albida, L.
anceps, L.
aurea, L.
autumnalis and
L.
eyermaniana. The shape of these crystals is raphides and druse crystals; however, they are scarce, and their location is in the center of the mesophyll (
Figure 9). These crystals were reported by Baker for the Laeliinae [
13]. Calcium oxalate crystals are also present in other groups of Orchidaceae and raphidia, where they tend to be very abundant in both vegetative and reproductive structures [
20].
Laelia halbingeriana is a natural hybrid and differs from
L. superbiens by morphological peculiarities such as shorter and thicker pseudobulbs and a darkly bilobed anther [
21]. However, both species live at medium altitudes (600 to 2000 m asl) in semi-deciduous tropical forests [
7]. In this work, relevant characters were found in the foliar anatomy shared by these species, such as isodiametric polygonal shape of adaxial and abaxial epidermal cells, hypodermic cells with a lignified wall and heterogeneous mesophyll.
Finally,
Laelia aurea and
L. rubescens are species that stand out as phylogenetically related species that also share a morphological and anatomical similarity but are distinguished by the intense yellow color of the flowers and the shape of the more ovate pseudobulbs of
L.
aurea. Both species also inhabit xeric environments, mainly deciduous, dry tropical forests [
7]. In a more recent study, Peraza Flores and collaborators made a molecular phylogeny from seven regions of plastid DNA and the ITS region (5′ partial 18S, ITS1, 5.8S, ITS2, 3′ partial 26S) where they included the Mexican species of
Laelia [
22]. They propose that the
Laelia species need to be separated into two main clades, the
Laelia Lindl. clade and the clade
Schomburgkia Lindl.
L.
aurea and
L.
rubescens, the only Mexican species that grow in lowland environments, are immersed in the
Schomburgkia clade. It is convenient to expand the sampling for anatomical studies where species of
Schomburgkia are included, which will allow better understanding of the similarity between these two genera.
In the group of the Mexican Laelias species,
L.
aurea and
L.
rubescens were recently recognized as sufficiently different from the rest of
Laelia to deserve genus status, for which Archila and Szlachetko proposed the creation of the genus
Encabarcenia Archila & Szlach [
23]. Nevertheless, based on molecular phylogeny, Peraza Flores and collaborators placed them in
Schomburgkia for sharing low-elevation environments and other morphological characters [
22]. In the present work, we showed that these two species had several structural characters in common and were different from the rest of the Laelias; therefore, we support the proposal of the latter and emphasize the importance of extending anatomical studies to include species of
Schomburgkia.
From the combined analysis of morphological and anatomical data we were able to show that Laelia species from Mexico shared structural characters from which they can be distinguished from Broughtonia negrilensis, and to a greater extent from Cattleya mossiae and Cattleya purpurata, providing taxonomically useful information. It is important to emphasize that the present work provides information on the functional anatomical characters for the identification and distinction of Mexican Laelias such as area of abaxial epidermal cells, thickness of the abaxial cuticle and thickness of the mesophyll. This work also showed that within the Laelias of Mexico, there was a correlation between certain structural characters and the altitudinal distribution of their species. Taken together, these results contribute towards a better understanding of orchid evolution in Mexico.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}